
The Effect of Chitosan Derivatives on the Compaction and Tension Generation of the Fibroblast-populated Collagen Matrix



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
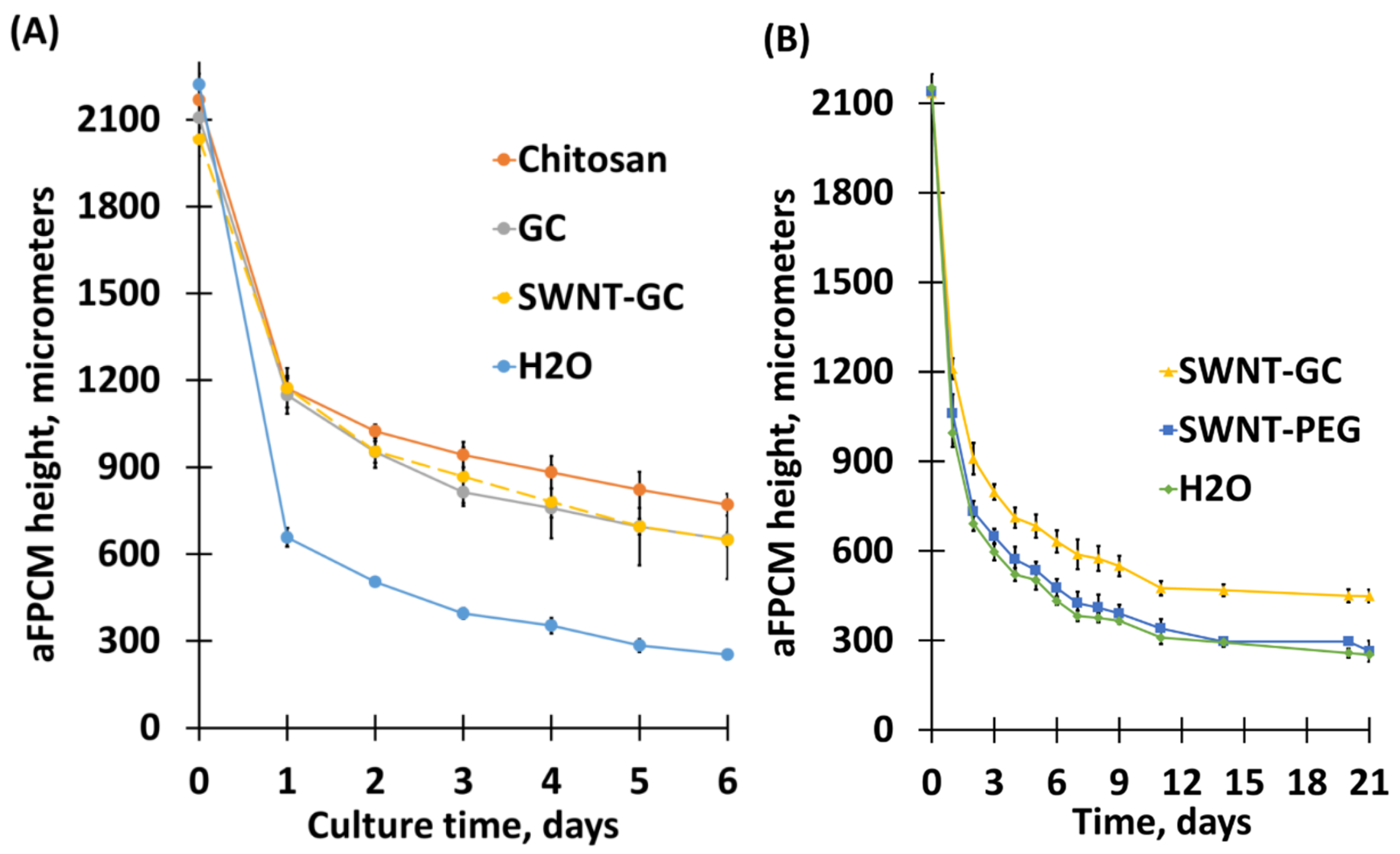
2.1. Compaction Analysis of aFPCM
2.1.1. Chitosan Derivatives, When Incorporated Into aFPCM, Reduce Their Compaction
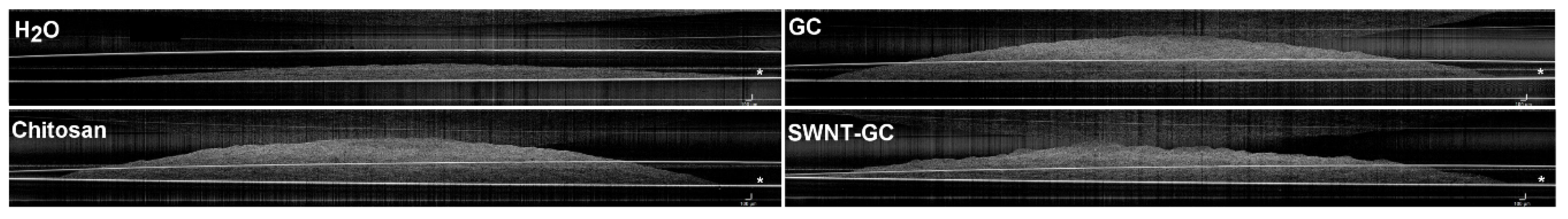
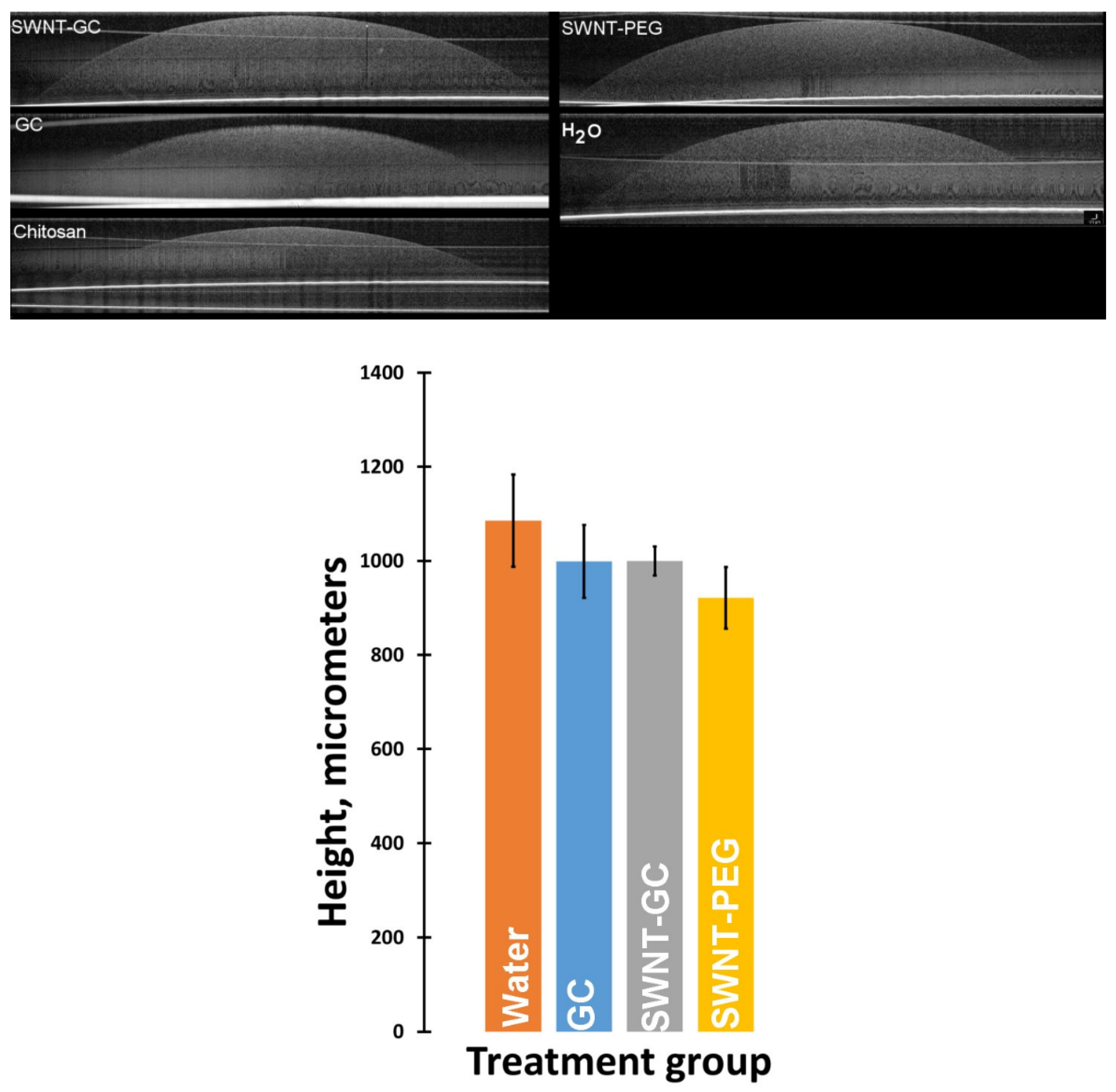
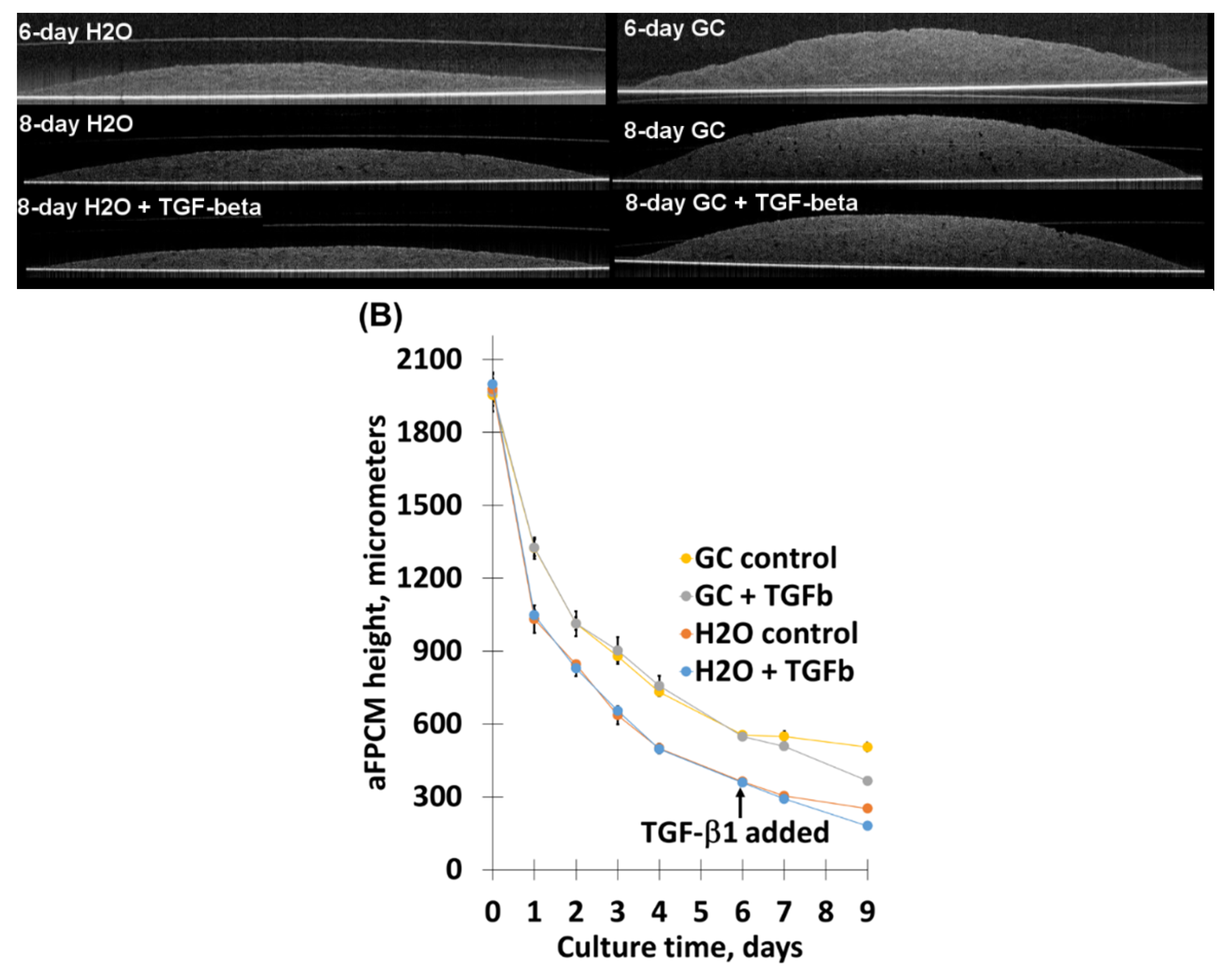
2.1.2. Optical Coherence Tomography (OCT) Imaging Shows Distinctive Morphology and Compaction of aFPCM
2.1.3. Tissue Swelling Was Not Observed When Chitosan Derivatives Were Present in The Matrix
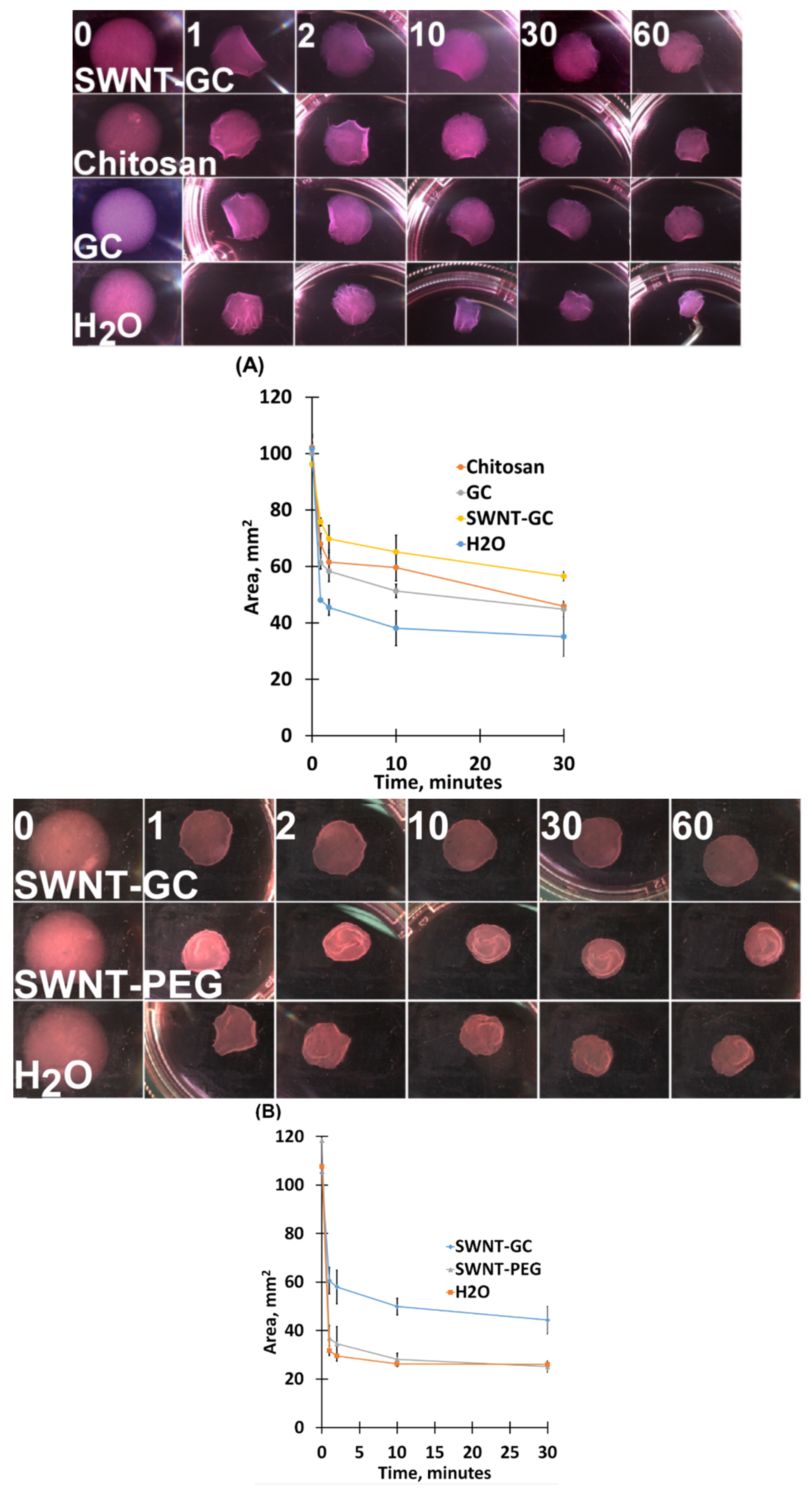
2.2. Chitosan Derivatives Reduce aFPCM Contraction (Diameter Reduction) When Released from Anchorage
2.3. Chitosan Derivative Presence Was Not Sufficient To Inhibit TGF-β1-Mediated Compaction Increase, Nor Its Correlated Contraction Increase
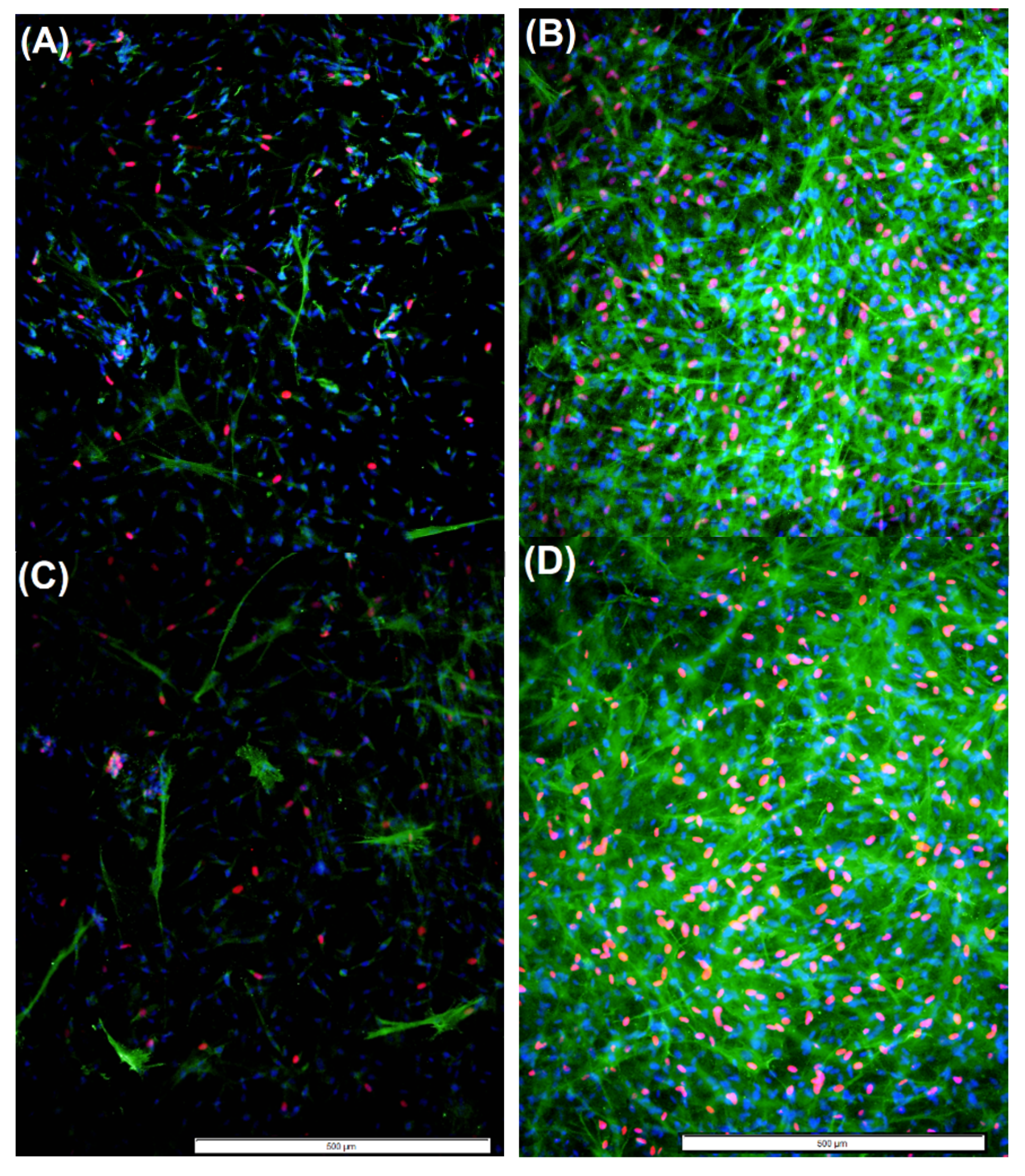
2.4. Cell Proliferation and Myofibroblast Presence Observed in TGF-β1-Treated Chitosan Derivative-incorporated aFPCM
3. Discussion
4. Materials and Methods
4.1. Human Cell Cultures
4.2. Preparation of aFPCM
Anchored Fibroblast-Populated Collagen Matrix (aFPCM) Assay
4.3. Compaction Measurements of aFPCM Assay
4.3.1. The aFPCM Height Measurement using Zeiss Primovert Microscope
4.3.2. Optical Coherence Tomography (OCT) Imaging on aFPCM
4.4. Released aFPCM Contraction Assay
4.5. Myofibroblast Proliferative Assay
4.6. Imaging and Analysis
4.7. Calculations/Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B.; McCulloch, C.A.; Coelho, N.M. Mechanical regulation of myofibroblast phenoconversion and collagen contraction. Exp. Cell Res. 2019, 379, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, M.B.; Odejimi, T.D.; Morris, T.L.; Sawalha, D.; Spencer, C.L. A new bioassay identifies proliferation ratios of fibroblasts and myofibroblasts. Cell Biol. Int. 2014, 38, 981–986. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, D.B.; Vi, L.; Gan, B.S. Molecular mechanisms and treatment strategies for Dupuytren’s disease. Ther. Clin. Risk Manag. 2010, 6, 383–390. [Google Scholar] [PubMed]
- Hindocha, S.; Stanley, J.K.; Watson, J.S.; Bayat, A. Revised Tubiana’s staging system for assessment of disease severity in Dupuytren’s disease-preliminary clinical findings. Hand (N Y) 2008, 3, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Warwick, D. Dupuytren’s disease: My personal view. J. Hand Surg. Eur. Vol. 2017, 42, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Kan, H.J.; Verrijp, F.W.; Hovius, S.E.R.; van Nieuwenhoven, C.A.; Dupuytren Delphi, G.; Selles, R.W. Recurrence of Dupuytren’s contracture: A consensus-based definition. PLoS ONE 2017, 12, e0164849. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Zhou, F.; Nordquist, R.E.; Carubelli, R.; Liu, H.; Chen, W.R. Glycated chitosan as a new non-toxic immunological stimulant. Immunopharmacol. Immunotoxicol. 2009, 31, 202–208. [Google Scholar] [CrossRef]
- Chen, W.R.; Carubelli, R.; Liu, H.; Nordquist, R.E. Detection of anti-tumor immunity induced by laser immunotherapy. Mol. Biotechnol. 2003, 25, 45–52. [Google Scholar] [CrossRef]
- Chen, W.R.; Carubelli, R.; Liu, H.; Nordquist, R.E. Laser immunotherapy: A novel treatment modality for metastatic tumors. Mol. Biotechnol. 2003, 25, 37–44. [Google Scholar] [CrossRef]
- Zhou, F.; Li, X.; Naylor, M.F.; Hode, T.; Nordquist, R.E.; Alleruzzo, L.; Raker, J.; Lam, S.S.; Du, N.; Shi, L.; et al. InCVAX—A novel strategy for treatment of late-stage, metastatic cancers through photoimmunotherapy induced tumor-specific immunity. Cancer Lett. 2015, 359, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Yang, J.; Zhang, Y.; Liu, M.; Lang, M.L.; Li, M.; Chen, W.R. Local Phototherapy Synergizes with Immunoadjuvant for Treatment of Pancreatic Cancer through Induced Immunogenic Tumor Vaccine. Clin. Cancer Res. 2018, 24, 5335–5346. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Doughty, A.; West, C.; Wang, L.; Zhou, F.; Nordquist, R.E.; Chen, W.R. Phototherapy using immunologically modified carbon nanotubes to potentiate checkpoint blockade for metastatic breast cancer. Nanomedicine 2019, 18, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Wang, C.Y.; Yang, F.Y.; Wang, B.S.; Chen, J.Y.; Lin, L.T.; Leu, J.D.; Chiu, S.J.; Chen, F.D.; Lee, Y.J.; et al. Synergistic effects of glycated chitosan with high-intensity focused ultrasound on suppression of metastases in a syngeneic breast tumor model. Cell Death Dis. 2014, 5, e1178. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.Y.; Leu, J.D.; Chang, C.Y.; Lee, Y.J.; Chen, W.R. Combination of Radiofrequency Ablation and Glycated Chitosan as Treatment on a Syngeneic Breast Tumor Model. Anticancer Res. 2017, 37, 2965–2974. [Google Scholar] [CrossRef]
- Kam, N.W.; Dai, H. Carbon nanotubes as intracellular protein transporters: Generality and biological functionality. J. Am. Chem. Soc. 2005, 127, 6021–6026. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Wu, S.; Song, S.; Chen, W.R.; Resasco, D.E.; Xing, D. Antitumor immunologically modified carbon nanotubes for photothermal therapy. Biomaterials 2012, 33, 3235–3242. [Google Scholar] [CrossRef] [PubMed]
- Layton, E.; McNamar, R.; Hasanjee, A.M.; McNair, C.; Stevens, B.; Vaughan, M.; Zhou, F.; Chen, W.R. Effects of Single-Walled Carbon Nanotubes on Cancer Cell Migration Using A Pancreatic Tumor Model. In Proceedings of the SPIE BiOS, San Francisco, CA, USA, 23 February 2017; Volume 10065. [Google Scholar]
- Okamoto, Y.; Watanabe, M.; Miyatake, K.; Morimoto, M.; Shigemasa, Y.; Minami, S. Effects of chitin/chitosan and their oligomers/monomers on migrations of fibroblasts and vascular endothelium. Biomaterials 2002, 23, 1975–1979. [Google Scholar] [CrossRef]
- Mariappan, M.R.; Alas, E.A.; Williams, J.G.; Prager, M.D. Chitosan and chitosan sulfate have opposing effects on collagen-fibroblast interactions. Wound Repair Regen. 1999, 7, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Grinnell, F.; Petroll, W.M. Cell motility and mechanics in three-dimensional collagen matrices. Annu Rev. Cell Dev. Biol. 2010, 26, 335–361. [Google Scholar] [CrossRef]
- Vaughan, M.B.; Xu, G.; Morris, T.L.; Kshetri, P.; Herwig, J.X. Predictable Fibroblast Tension Generation by Measuring Compaction of Anchored Collagen Matrices using Microscopy and Optical Coherence Tomography. Cell Adhes. Migr. 2019, 13, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Tan, F.J.; Marra, K.G.; Jan, S.S.; Liu, D.C. Synthesis and characterization of collagen/hyaluronan/chitosan composite sponges for potential biomedical applications. Acta Biomater. 2009, 5, 2591–2600. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Fu, Y.; Chung, W.; Zheng, X.; Cui, Y.; Hsu, I.C.; Kao, W.J. Thiol-ene-based biological/synthetic hybrid biomatrix for 3-D living cell culture. Acta Biomater. 2012, 8, 2504–2516. [Google Scholar] [CrossRef] [PubMed]
- Montesano, R.; Orci, L. Transforming growth factor beta stimulates collagen-matrix contraction by fibroblasts: Implications for wound healing. Proc. Natl. Acad. Sci. USA 1988, 85, 4894–4897. [Google Scholar] [CrossRef] [PubMed]
- Grinnell, F.; Ho, C.H. Transforming growth factor beta stimulates fibroblast-collagen matrix contraction by different mechanisms in mechanically loaded and unloaded matrices. Exp. Cell Res. 2002, 273, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Obioma, C.C.; Layton, E.; Doan, K.T.; Xu, G.; Zhou, F.; Chen, W.R.; Vaughan, M.B. Effect of near infrared lasers and glycated chitosan on myofibroblast differentiation and contraction. In Proceedings of the SPIE BiOS, San Francisco, CA, USA, 19 February 2018; p. 8. [Google Scholar]
- Desmouliere, A.; Geinoz, A.; Gabbiani, F.; Gabbiani, G. Transforming growth factor-beta 1 induces alpha-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. J. Cell Biol. 1993, 122, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, M.B.; Howard, E.W.; Tomasek, J.J. Transforming Growth Factor-β1 Promotes the Morphological and Functional Differentiation of the Myofibroblast. Exp. Cell Res. 2000, 257, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Grinnell, F. Fibroblasts, myofibroblasts, and wound contraction. J. Cell Biol. 1994, 124, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Dallon, J.C.; Ehrlich, H.P. A review of fibroblast-populated collagen lattices. Wound Repair Regen. 2008, 16, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Grinnell, F.; Zhu, M.; Carlson, M.A.; Abrams, J.M. Release of mechanical tension triggers apoptosis of human fibroblasts in a model of regressing granulation tissue. Exp. Cell Res. 1999, 248, 608–619. [Google Scholar] [CrossRef] [PubMed]
- Tomasek, J.J.; Haaksma, C.J.; Eddy, R.J.; Vaughan, M.B. Fibroblast contraction occurs on release of tension in attached collagen lattices: Dependency on an organized actin cytoskeleton and serum. Anat. Rec. 1992, 232, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Tomasek, J.; Rayan, G.M. Correlation of alpha-smooth muscle actin expression and contraction in Dupuytren’s disease fibroblasts. J. Hand Surg. Am. 1995, 20, 450–455. [Google Scholar] [CrossRef]
- Herwig, J.X.; Vaughan, M.B. Fibroblast contraction is dose-dependent on serum in an updated stress-relaxed collagen lattice model. UCO CMS Undergrad. J. 2013, 1, 25–31. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doan, K.T.; Kshetri, P.; Attamakulsri, N.; Newsome, D.R.; Zhou, F.; Murray, C.K.; Chen, W.R.; Xu, G.; Vaughan, M.B. The Effect of Chitosan Derivatives on the Compaction and Tension Generation of the Fibroblast-populated Collagen Matrix. Molecules 2019, 24, 2713. https://doi.org/10.3390/molecules24152713
Doan KT, Kshetri P, Attamakulsri N, Newsome DR, Zhou F, Murray CK, Chen WR, Xu G, Vaughan MB. The Effect of Chitosan Derivatives on the Compaction and Tension Generation of the Fibroblast-populated Collagen Matrix. Molecules. 2019; 24(15):2713. https://doi.org/10.3390/molecules24152713
Chicago/Turabian StyleDoan, K. Tu, Pratiksha Kshetri, Natthapume Attamakulsri, Derek R. Newsome, Feifan Zhou, Cynthia K. Murray, Wei R. Chen, Gang Xu, and Melville B. Vaughan. 2019. "The Effect of Chitosan Derivatives on the Compaction and Tension Generation of the Fibroblast-populated Collagen Matrix" Molecules 24, no. 15: 2713. https://doi.org/10.3390/molecules24152713
APA StyleDoan, K. T., Kshetri, P., Attamakulsri, N., Newsome, D. R., Zhou, F., Murray, C. K., Chen, W. R., Xu, G., & Vaughan, M. B. (2019). The Effect of Chitosan Derivatives on the Compaction and Tension Generation of the Fibroblast-populated Collagen Matrix. Molecules, 24(15), 2713. https://doi.org/10.3390/molecules24152713