
Dendrobium officinale Polysaccharides Inhibit 1-Methyl-2-Nitro-1-Nitrosoguanidine Induced Precancerous Lesions of Gastric Cancer in Rats through Regulating Wnt/β-Catenin Pathway and Altering Serum Endogenous Metabolites

 ,
,
Abstract
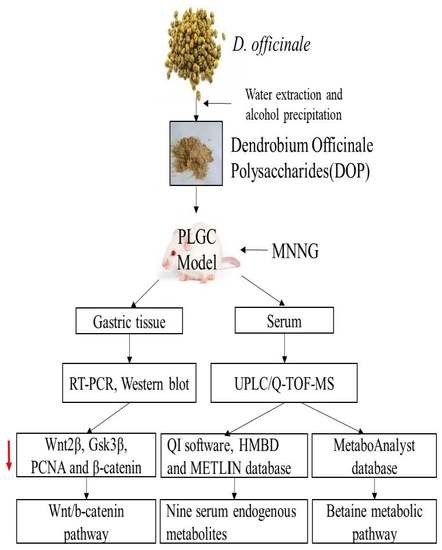
1. Introduction
2. Result
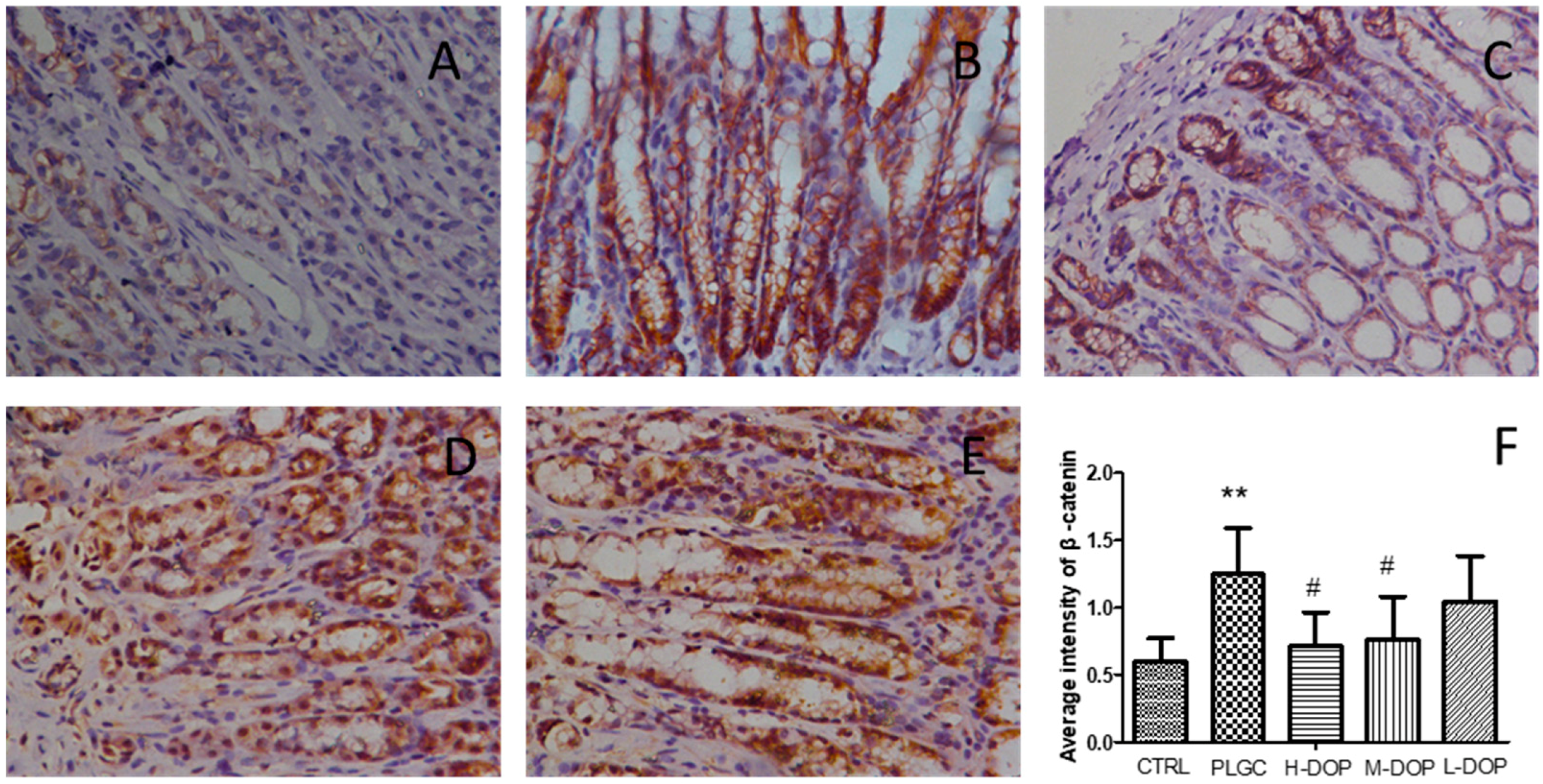
2.1. Immunohistochemical Analysis of β-Catenin
2.2. RT-PCR Analysis of Gene related to Wnt/β-Catenin Pathway
2.3. Western Blot Analysis of Wnt2β, PCNA, and β-Catenin Protein Expression
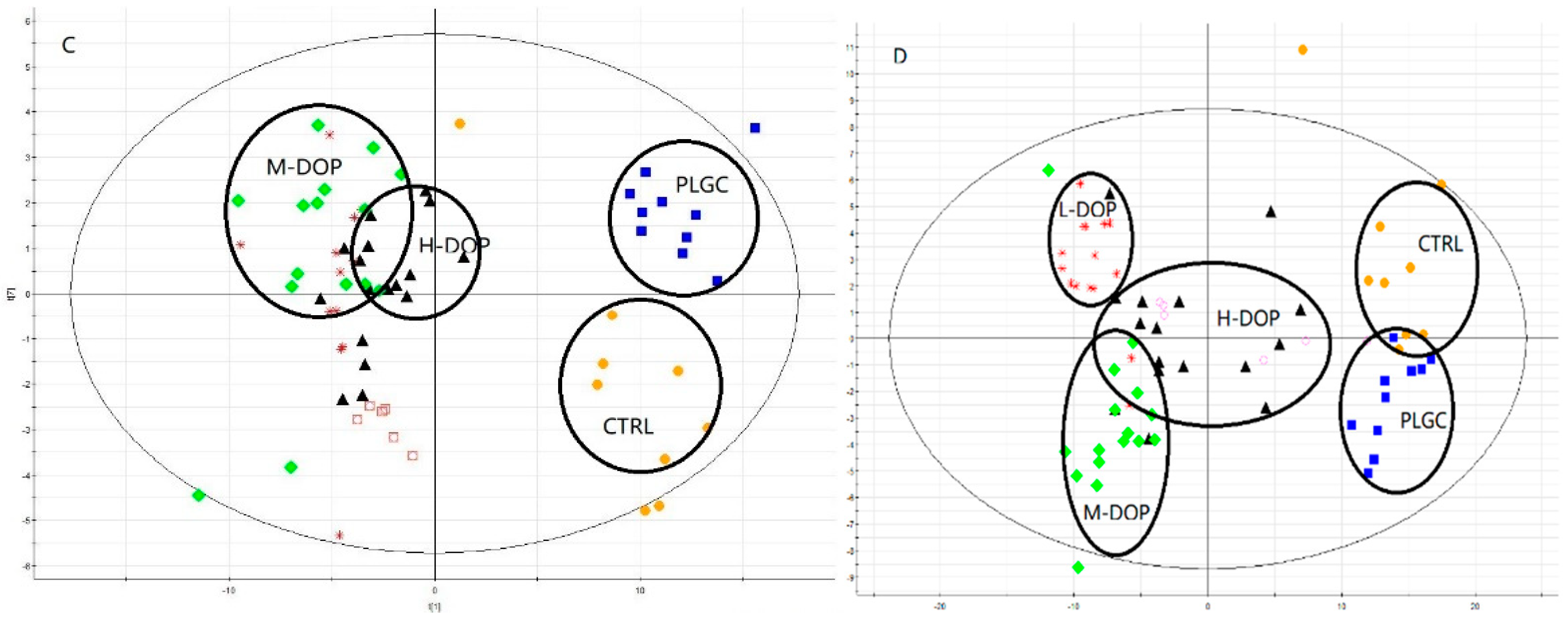
2.4. Metabolites Profiles in the Serum
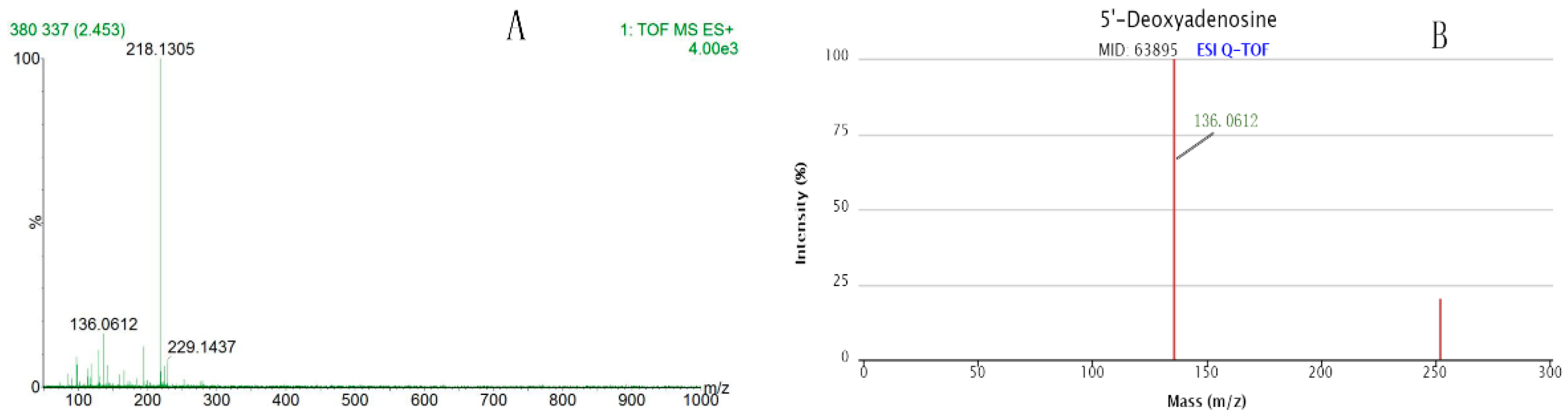
2.5. Potential Endogenous Metabolites Identification
2.6. Determination of Endogenous Metabolites
2.7. Enrichment Analysis of Differential Endogenous Metabolites
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Drugs and Reagents
4.3. Inhibition Effects of DOP on PLGC in Rats
4.4. Immunohistochemistry
4.5. Total RNA Extraction and Quantitative Real-Time RT-PCR
4.6. Western Blot Analysis
4.7. Preparation of Plasm and UPLC/Q-TOF-MS Analysis
4.8. Metabolomics Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PLGC | Precancerous lesions of gastric cancer |
| TCM | Traditional Chinese medicine |
| DOP | Dendrobium officinale polysaccharides |
| MNNG | 1-Methyl-2-nitro-1-nitrosoguanidine |
| HPGPC | High-Performance Gel-Permeation Chromatography |
| UPLC/Q-TOF-MS | Ultra-performance liquid chromatography-quadrupole-time-of-flight-mass spectrometry |
| PLS-DA | Partial least squares discriminant analysis |
| Wnt2β | Wnt Family Member 2B |
| Gsk3β | Glycogen Synthase Kinase 3 Beta |
| PCNA | Proliferating Cell Nuclear Antigen |
| β-catenin | Catenin beta 1 |
| CyclinD1 | G1/S-Specific Cyclin D1 |
| DAB | 3,3′-diaminobenzidine |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Leung, W.K.; Sung, J.J. Review article: Intestinal metaplasia and gastric carcinogenesis. Aliment Pharmacol Ther. 2002, 16, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Malik, T.H.; Sayahan, M.Y.; Al, A.H.; Hong, X. Gastric Intestinal Metaplasia: An Intermediate Precancerous Lesion in the Cascade of Gastric Carcinogenesis. J. Coll. Physicians Surg. Pak. 2017, 27, 166–172. [Google Scholar] [PubMed]
- Heo, S.H.; Jeong, E.S.; Lee, K.S.; Seo, J.H.; Jeong, D.G.; Won, Y.S.; Kwon, H.J.; Kim, H.C.; Kim, D.Y.; Choi, Y.K. Canonical Wnt signaling pathway plays an essential role in N-methyl-N-nitrosurea induced gastric tumorigenesis of mice. J. Vet. Med. Sci. 2013, 75, 299–307. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Oliveira, L.A.; Oshima, C.; Soffner, P.A.; Silva, M.S.; Lins, R.R.; Malinverni, A.; Waisberg, J. The canonal Wnt pathway in gastric carcinoma. Arq. Bras. Cir. Dig. 2019, 32, e1414. [Google Scholar] [CrossRef]
- Liesenfeld, D.B.; Habermann, N.; Owen, R.W.; Scalbert, A.; Ulrich, C.M. Review of mass spectrometry–based metabolomics in cancer research. Canc. Epidemiol. Biomarkers Prev. 2013, 22, 2182–2201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Sun, H.; Wang, X. Serum metabolomics as a novel diagnostic approach for disease: A systematic review. Anal. Bioanal. Chem. 2012, 404, 1239–1245. [Google Scholar] [CrossRef]
- Yan, M.; Xu, G. Current and future perspectives of functional metabolomics in disease studies—A review. Anal. Chim. Acta 2018, 1037, 41–54. [Google Scholar] [CrossRef]
- Slupsky, C.M.; Steed, H.; Wells, T.H.; Dabbs, K.; Schepansky, A.; Capstick, V.; Faught, W.; Sawyer, M.B. Urine metabolite analysis offers potential early diagnosis of ovarian and breast cancers. Clin. Cancer Res. 2010, 16, 5835–5841. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, Y.; Lan, X.M.; Xu, G.L.; Sun, Y.Z.; Li, F.; Liu, H.N. Effect of Dendrobium officinale Extraction on Gastric Carcinogenesis in Rats. Evid. Based Complement. Alternat. Med. 2016, 2016, e19647. [Google Scholar] [CrossRef]
- Wang, G.Y.; Liu, H.N.; Ge, S.C.; Jiang, Y.N.; Zhao, Y. Urine metabolomics analysis of Dendrobium Officinale water extraction on precancerous lesions of gastric cancer. Chin. J. Exp. Tradit. Med. Formul. 2018, 24, 77–85. [Google Scholar]
- Zhu, C.; Huang, Q.; Zhu, H. Melatonin Inhibits the Proliferation of Gastric Cancer Cells Through Regulating the miR-16-5p-Smad3 Pathway. DNA Cell Biol. 2018, 37, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Sun, Y.; Wang, G.; Ge, S.; Liu, H. Dendrobium Officinale Polysaccharides Protect against MNNG-Induced PLGC in Rats via Activating the NRF2 and Antioxidant Enzymes HO-1 and NQO-1. Oxid. Med. Cell. Longev. 2019. [Google Scholar] [CrossRef] [PubMed]
- Yoo, N.J.A.K. Mutational Analysis of WTX Gene in Wnt/beta-Catenin Pathway in Gastric, Colorectal, and Hepatocellular Carcinomas. Dig Dis Sci. 2009, 54, 1011–1014. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Xin, N.; Wang, W.; Zhao, C. Wnt/beta-catenin, an oncogenic pathway targeted by H. pylori in gastric carcinogenesis. Oncotarget 2015, 6, 35579–35588. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, J.; Dong, X. Wnt2 contributes to the progression of gastric cancer by promoting cell migration and invasion. Oncol. Lett. 2018, 16, 2857–2864. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, T.; Yamamoto, M.; Ogasawara, N.; Ushijima, T.; Nomoto, T.; Fujita, H.; Matsushima, T.; Nozaki, K.; Cao, X.; Tatematsu, M. beta-Catenin mutations and nuclear accumulation during progression of rat stomach adenocarcinomas. Cancer Sci. 2003, 94, 1046–1051. [Google Scholar] [CrossRef]
- Takasu, S.; Tsukamoto, T.; Ushijima, T.; Yamashita, S.; Ogasawara, N.; Ban, H.; Yanai, T.; Masegi, T.; Tatematsu, M. Cyclin D1 overexpression in N-methyl-N′-nitro-N-nitrosoguanidine-induced rat gastric adenocarcinomas. Exp. Toxicol. Pathol. 2007, 59, 171–175. [Google Scholar] [CrossRef]
- Bo Peng, H.L.X.P. Functional metabolomics: From biomarker discovery to metabolome reprogramming. Protein Cell. 2015, 9, 628–637. [Google Scholar] [CrossRef]
- Corona, G.; Cannizzaro, R.; Miolo, G.; Caggiari, L.; De Zorzi, M.; Repetto, O.; Steffan, A.; De Re, V. Use of Metabolomics as a Complementary Omic Approach to Implement Risk Criteria for First-Degree Relatives of Gastric Cancer Patients. Int. J. Mol. Sci. 2018, 19, 750. [Google Scholar] [CrossRef]
- Gu, J.; Hu, X.; Shao, W.; Ji, T.; Yang, W.; Zhuo, H.; Jin, Z.; Huang, H.; Chen, J.; Huang, C.; et al. Metabolomic analysis reveals altered metabolic pathways in a rat model of gastric carcinogenesis. Oncotarget 2016, 7, 60053–60073. [Google Scholar] [CrossRef] [PubMed]
- Craig, S.A. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Chen, S.; Zhu, G.; Huang, R.; Yin, Y.; Ren, W. Betaine Inhibits Interleukin-1beta Production and Release: Potential Mechanisms. Front. Immunol. 2018, 9, 2670. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, S.; Fang, S.; Yang, X.; Feng, J. Betaine Improves Intestinal Functions by Enhancing Digestive Enzymes, Ameliorating Intestinal Morphology, and Enriching Intestinal Microbiota in High-salt stressed Rats. Nutrients 2018, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.P.; Peng, J.S.; Sun, A.; Tang, Z.H.; Ling, W.H.; Zhu, H.L. Assessment of the effect of betaine on p16 and c-myc DNA methylation and mRNA expression in a chemical induced rat liver cancer model. BMC Cancer 2009, 9, 261. [Google Scholar] [CrossRef]
- Betancor-Fernandez, A.; Sies, H.; Stahl, W.; Polidori, M.C. In vitro antioxidant activity of 2,5,7,8-tetramethyl-2-(2′-carboxyethyl)-6-hydroxychroman (alpha-CEHC), a vitamin E metabolite. Free Radic. Res. 2002, 36, 915–921. [Google Scholar] [CrossRef]
- Galli, F.; Stabile, A.M.; Betti, M.; Conte, C.; Pistilli, A.; Rende, M.; Floridi, A.; Azzi, A. The effect of alpha- and gamma-tocopherol and their carboxyethyl hydroxychroman metabolites on prostate cancer cell proliferation. Arch. Biochem Biophys. 2004, 423, 97–102. [Google Scholar] [CrossRef]
- Luo, Q.L.; Tang, Z.H.; Zhang, X.F.; Zhong, Y.H.; Yao, S.Z.; Wang, L.S.; Lin, C.W.; Luo, X. Chemical properties and antioxidant activity of a water-soluble polysaccharide from Dendrobium officinale. Int. J. Biol. Macromol. 2016, 89, 219–227. [Google Scholar] [CrossRef]
- Mali, A.V.; Joshi, A.A.; Hegde, M.V.; Kadam, S.S. Enterolactone modulates the ERK/NF-kappaB/Snail signaling pathway in triple-negative breast cancer cell line MDA-MB-231 to revert the TGF-beta-induced epithelial-mesenchymal transition. Cancer Biol. Med. 2018, 15, 137–156. [Google Scholar]
- Liu, H.; Liu, J.; Wang, S.; Zeng, Z.; Li, T.; Liu, Y.; Mastriani, E.; Li, Q.H.; Bao, H.X.; Zhou, Y.J.; et al. Enterolactone has stronger effects than enterodiol on ovarian cancer. J. Ovarian Res. 2017, 10, 49. [Google Scholar] [CrossRef]
- Eriksen, A.K.; Kyro, C.; Norskov, N.; Bolvig, A.K.; Christensen, J.; Tjonneland, A.; Overvad, K.; Landberg, R.; Olsen, A. Prediagnostic enterolactone concentrations and mortality among Danish men diagnosed with prostate cancer. Eur. J. Clin. Nutr. 2017, 71, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Chikara, S.; Lindsey, K.; Borowicz, P.; Christofidou-Solomidou, M.; Reindl, K.M. Enterolactone alters FAK-Src signaling and suppresses migration and invasion of lung cancer cell lines. BMC Complement. Altern. Med. 2017, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Mali, A.V.; Padhye, S.B.; Anant, S.; Hegde, M.V.; Kadam, S.S. Anticancer and antimetastatic potential of enterolactone: Clinical, preclinical and mechanistic perspectives. Eur. J. Pharmacol. 2019, 852, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Ichikawa, H.; Fujita, T.; Takagi, T.; Osada-Oka, M.; Minamiyama, Y. Histidine deficiency attenuates cell viability in rat intestinal epithelial cells by apoptosis via mitochondrial dysfunction. J. Nutr. 2017, 8, 21–28. [Google Scholar] [CrossRef]
- Hunting, D.; Henderson, J.F. Effects of 5′-deoxyadenosine on phosphoribosyl pyrophosphate and purine nucleotide synthesis in Ehrlich ascites tumor cells in vitro. Biochem. Pharmacol. 1978, 27, 2163–2169. [Google Scholar] [CrossRef]
- Rao, D.; Peitzsch, M.; Prejbisz, A.; Hanus, K.; Fassnacht, M.; Beuschlein, F.; Brugger, C.; Fliedner, S.; Langton, K.; Pamporaki, C. Plasma Methoxytyramine: Clinical Utility with Metanephrines for Diagnosis of Pheochromocytoma and Paraganglioma. Eur. J. Endocrinol. 2017, 177, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Eisenhofer, G.; Lenders, J.W.; Timmers, H.; Mannelli, M.; Grebe, S.K.; Hofbauer, L.C.; Bornstein, S.R.; Tiebel, O.; Adams, K.; Bratslavsky, G.; et al. Measurements of plasma methoxytyramine, normetanephrine, and metanephrine as discriminators of different hereditary forms of pheochromocytoma. Clin. Chem. 2011, 57, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Heinzmann, S.S.; Brown, I.J.; Chan, Q.; Bictash, M.; Dumas, M.; Kochhar, S.; Stamler, J.; Holmes, E.; Elliott, P.; Nicholson, J.K. Metabolic profiling strategy for discovery of nutritional biomarkers: Proline betaine as a marker of citrus consumption. Am. J. Clin. Nutr. 2010, 92, 436–443. [Google Scholar] [CrossRef]
- Manjunatha, S.G.; Rajappa, S. From N-nitroacetylproline to leucylproline. J. Chem. Soc. Chem. Commun. 1991, 372–373. [Google Scholar] [CrossRef]
- Shearer, G.C.; Harris, W.S.; Pedersen, T.L.; Newman, J.W. Detection of omega-3 oxylipins in human plasma and response to treatment with omega-3 acid ethyl esters. J. Lipid Res. 2010, 51, 2074–2081. [Google Scholar] [CrossRef]
- Yi, Z.; Yan, L.; Lan, X.M.; Guo-Liang, X.U.; Liu, H.N. Study on inhibition of Dendrobium officinale extract on gastric carcinogenesis. Chin. Tradit. Herbal Drugs 2015, 46, 3704–3709. [Google Scholar]
- Yamachika, T.; Nakanishi, H.; Inada, K.; Tsukamoto, T.; Shimizu, N.; Kobayashi, K.; Fukushima, S.; Tatematsu, M. N-methyl-N-nitrosourea concentration-dependent, rather than total intake-dependent, induction of adenocarcinomas in the glandular stomach of BALB/c mice. Jpn. J. Cancer Res. 1998, 89, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Kokubo, T.; Furukawa, F.; Kurokawa, Y.; Tatematsu, M.; Hayashi, Y. Effect of high salt diet on rat gastric carcinogenesis induced by N-methyl-N′-nitro-N-nitrosoguanidine. Jpn. J. Cancer Res. 1983, 74, 28–34. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ion Model | Retention Time (min) | M/Z | Component ID | Molecular Formula | Component Name | P Value |
|---|---|---|---|---|---|---|
| Positive | 9.521 | 235.1691 | HMDB00043 | C5H11NO2 | Betaine | 2.48 × 10−10 |
| 10.305 | 395.2206 | HMDB04063 | C10H15NO3 | Metanephrine | 1.21 × 10−11 | |
| 6.889 | 299.1313 | HMDB06101 | C18H18O4 | Enterolactone | 0.000245555 | |
| 2.431 | 252.1095 | HMDB01983 | C10H13N5O3 | 5′-Deoxyadenosine | 0.0000987 | |
| 1.168 | 144.1020 | HMDB04827 | C7H13NO2 | Proline betaine | 0.011885662 | |
| 4.556 | 157.0608 | HMDB02320 | C6H8N2O3 | Imidazolelactic acid | 0.000000336 | |
| Negative | 3.679 | 227.1416 | HMDB11175 | C11H20N2O3 | L-leucyl-L-proline | 1.32 × 10−9 |
| 6.989 | 311.2237 | HMDB10208 | C18H32O4 | 15,16-DiHODE | 0.000509626 | |
| 4.920 | 277.1455 | HMDB01518 | C16H22O4 | Alpha-CEHC | 0.00000215 |
| Pathway Name | Total | Expected | Hits | Raw p | Holm p | FDR |
|---|---|---|---|---|---|---|
| Betaine Metabolism | 10 | 0.109 | 1 | 0.105 | 1 | 1 |
| Methionine Metabolism | 24 | 0.262 | 1 | 0.235 | 1 | 1 |
| Glycine, Serine and Threonine Metabolism | 26 | 0.284 | 1 | 0.252 | 1 | 1 |
| Tyrosine Metabolism | 38 | 0.415 | 1 | 0.348 | 1 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Li, B.; Wang, G.; Ge, S.; Lan, X.; Xu, G.; Liu, H. Dendrobium officinale Polysaccharides Inhibit 1-Methyl-2-Nitro-1-Nitrosoguanidine Induced Precancerous Lesions of Gastric Cancer in Rats through Regulating Wnt/β-Catenin Pathway and Altering Serum Endogenous Metabolites. Molecules 2019, 24, 2660. https://doi.org/10.3390/molecules24142660
Zhao Y, Li B, Wang G, Ge S, Lan X, Xu G, Liu H. Dendrobium officinale Polysaccharides Inhibit 1-Methyl-2-Nitro-1-Nitrosoguanidine Induced Precancerous Lesions of Gastric Cancer in Rats through Regulating Wnt/β-Catenin Pathway and Altering Serum Endogenous Metabolites. Molecules. 2019; 24(14):2660. https://doi.org/10.3390/molecules24142660
Chicago/Turabian StyleZhao, Yi, Bingtao Li, Gaoyu Wang, Shuchao Ge, Ximing Lan, Guoliang Xu, and Hongning Liu. 2019. "Dendrobium officinale Polysaccharides Inhibit 1-Methyl-2-Nitro-1-Nitrosoguanidine Induced Precancerous Lesions of Gastric Cancer in Rats through Regulating Wnt/β-Catenin Pathway and Altering Serum Endogenous Metabolites" Molecules 24, no. 14: 2660. https://doi.org/10.3390/molecules24142660
APA StyleZhao, Y., Li, B., Wang, G., Ge, S., Lan, X., Xu, G., & Liu, H. (2019). Dendrobium officinale Polysaccharides Inhibit 1-Methyl-2-Nitro-1-Nitrosoguanidine Induced Precancerous Lesions of Gastric Cancer in Rats through Regulating Wnt/β-Catenin Pathway and Altering Serum Endogenous Metabolites. Molecules, 24(14), 2660. https://doi.org/10.3390/molecules24142660