Neuroprotective and Anti-Amyloid β Effect and Main Chemical Profiles of White Tea: Comparison Against Green, Oolong and Black Tea

 , , and
, , and
Abstract
1. Introduction
2. Results
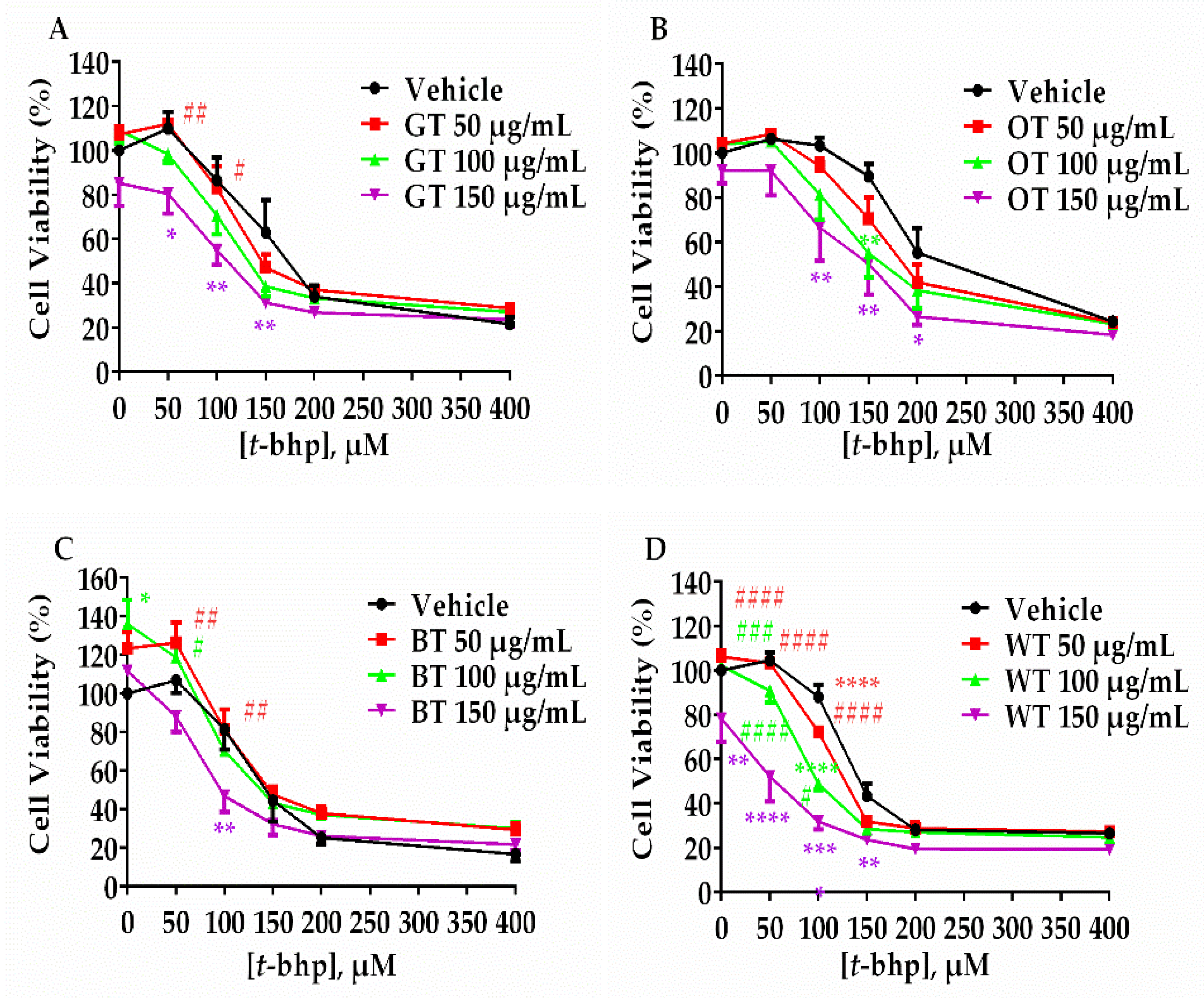
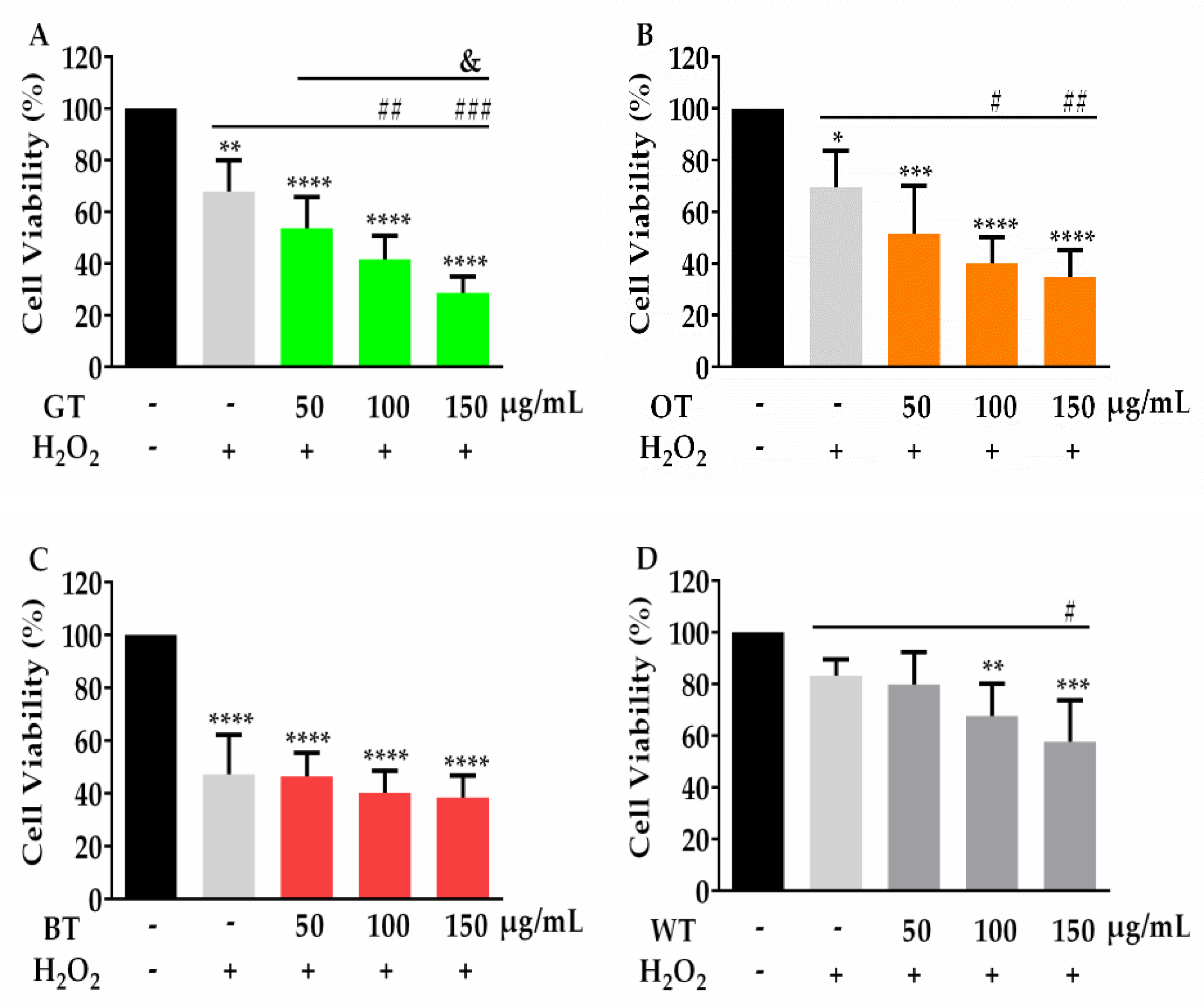
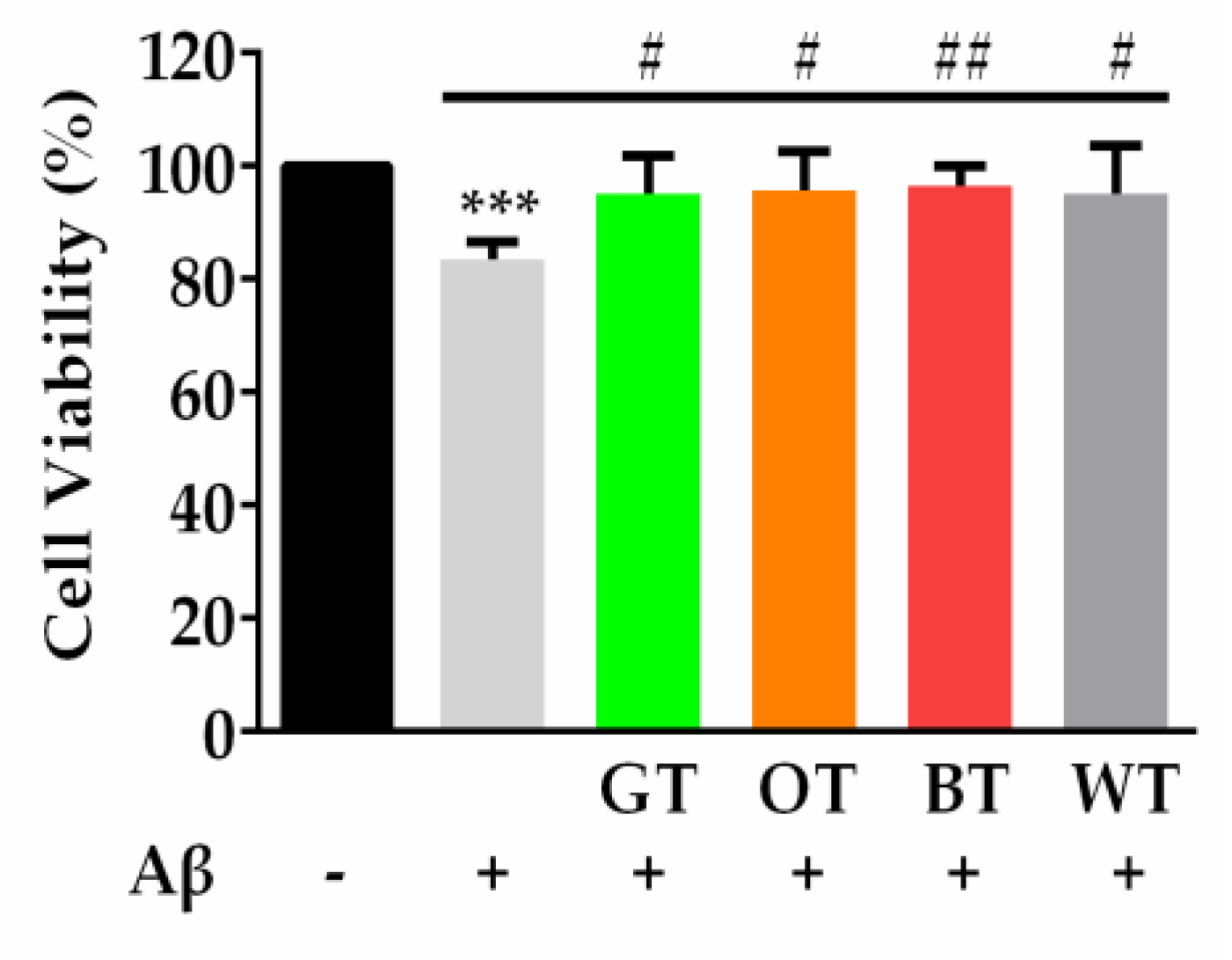
2.1. WT Significantly Inhibited Aβ-Evoked Neuronal Cell Death but not Oxidative Stress
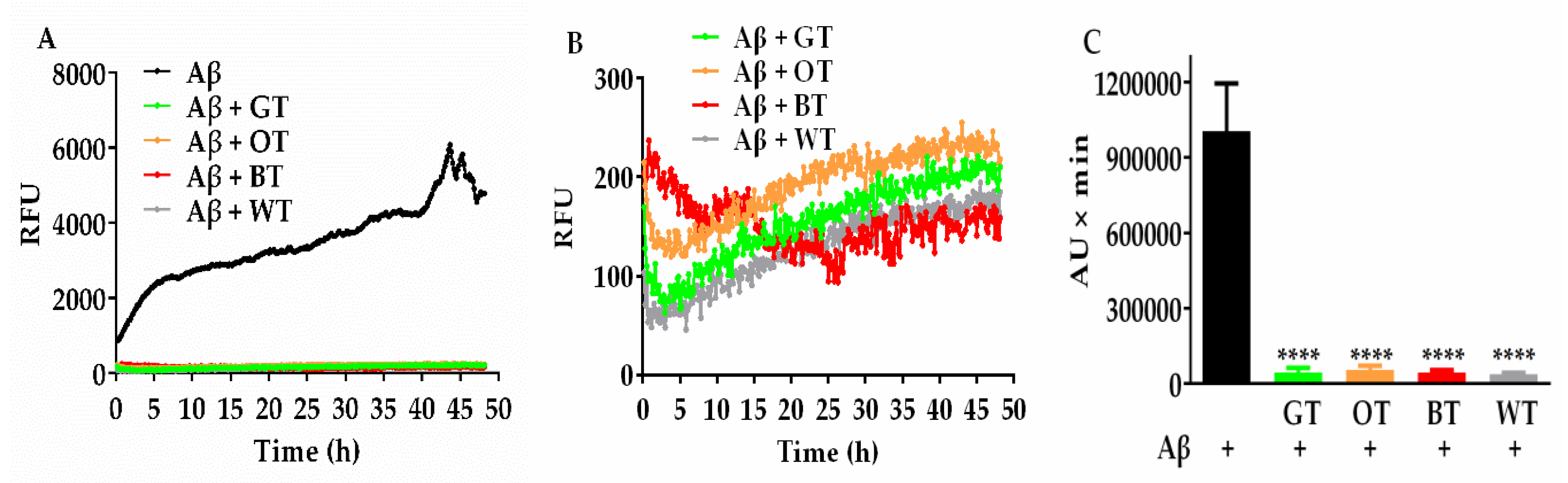
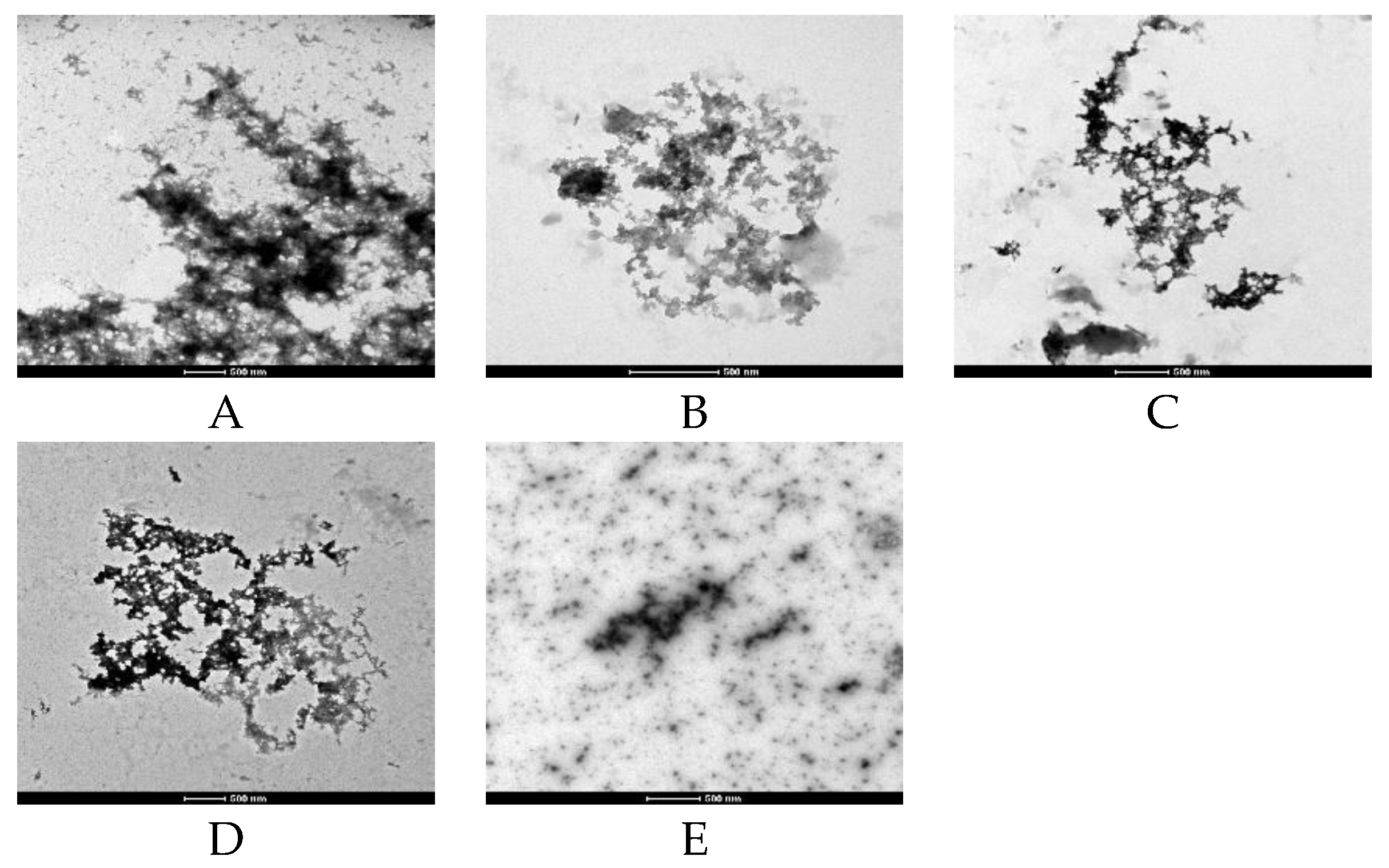
2.2. Effects of WT on Aβ Fibril Kinetics and Aggregate Morphology (ThT and TEM Assays)
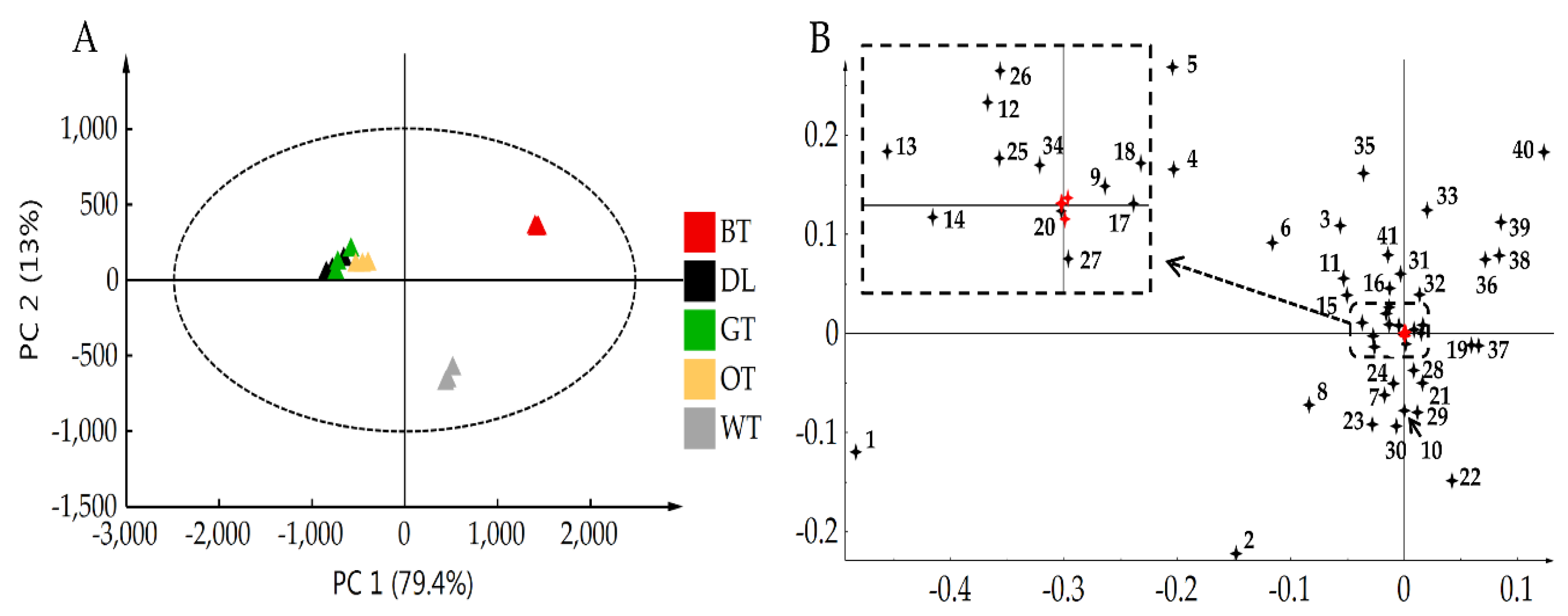
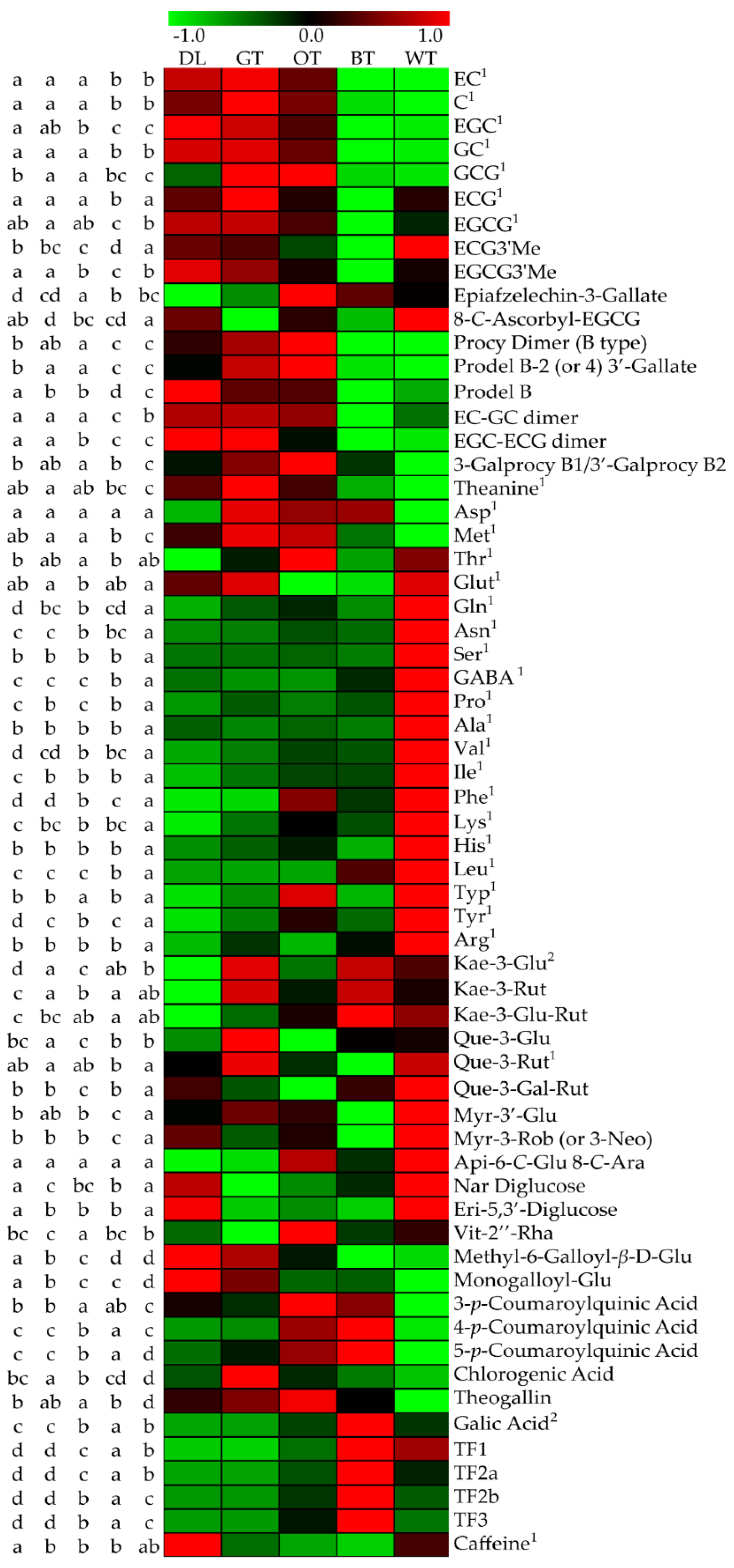
2.3. UPLC-QTOF-MS and UPLC-QqQ-MS Results Showed That Catechin Derivatives, Flavonol or Flavone Glycosides and Amino Acids were Characteristic Compounds of WT
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Tea Samples Preparation for Cell Culture and Chemical Analysis
4.3. Cell Culture
4.4. Cell Treatment and Cell Viability Measurements
4.5. Aβ Preparation
4.6. ThT Assay and TEM of Aβ Fibril and Aggregate Formation
4.7. UPLC-QqQ-MS Based Targeted Quantification of Catechins, Que-3-Rut, Caffeine and Amino Acids in Different Tea Types
4.8. UPLC-QTOF MS-Based Non-Targeted Metabolite Analysis of Different Tea Types
4.9. Data Processing, Metabolite Identification, and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- John, H.; Dennis J, S. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef]
- Yankner, B.A.; Lu, T. Amyloid β-protein toxicity and the pathogenesis of Alzheimer Disease. J. Biol. Chem. 2009, 284, 4755–4759. [Google Scholar] [CrossRef] [PubMed]
- Naghma, K.; Hasan, M. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef]
- Harvey, B.S.; Musgrave, I.F.; Ohlsson, K.S.; Fransson, Å.; Smid, S.D. The green tea polyphenol (−)-epigallocatechin-3-gallate inhibits amyloid-β evoked fibril formation and neuronal cell death in vitro. Food Chem. 2011, 129, 1729–1736. [Google Scholar] [CrossRef]
- Stéphane, B.; Zhi-Xing, Y.; Vassilios, P.; Rémi, Q. Neuroprotective effects of green and black teas and their catechin gallate esters against β-amyloid-induced toxicity. Eur. J. Neurosci. 2006, 23, 55–64. [Google Scholar] [CrossRef]
- Grelle, G.; Otto, A.; Lorenz, M.; Frank, R.; Wanker, E.E.; Bieschke, J. Black tea theaflavins inhibit formation of toxic amyloid-β and α-synuclein fibrils. Biochemistry 2011, 50, 10624–10636. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Stark, L.; Musgrave, I.F.; Pukala, T.L.; Smid, S.D. Bioactive polyphenol interactions with β amyloid: A comparison of binding modelling, effects on fibril and aggregate formation and neuroprotective capacity. Food Func. 2016, 7, 1138–1146. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Y.; Su, Y.; Zhou, W.; Yang, S.; Zhang, R.; Zhao, M.; Li, Y.; Zhang, Z.; Zhan, D. Rutin inhibits β-amyloid aggregation and cytotoxicity, attenuates oxidative stress, and decreases the production of nitric oxide and proinflammatory cytokines. Neurotoxicology 2012, 33, 482–490. [Google Scholar] [CrossRef]
- Marsh, D.T.; Das, S.; Ridell, J.; Smid, S.D. Structure-activity relationships for flavone interactions with amyloid β reveal a novel anti-aggregatory and neuroprotective effect of 2’,3’,4’-trihydroxyflavone (2-D08). Bioorg. Med. Chem. 2017, 25, 3827–3834. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.; Feng, L.; Niti, M.; Kua, E.H.; Yap, K.B. Tea consumption and cognitive impairment and decline in older Chinese adults. Am. J. Clin. Nutr. 2008, 88, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Noguchishinohara, M.; Yuki, S.; Dohmoto, C.; Ikeda, Y.; Samuraki, M.; Iwasa, K.; Yokogawa, M.; Asai, K.; Komai, K.; Nakamura, H. Consumption of green tea, but not black tea or coffee, is associated with reduced risk of cognitive decline. PLoS ONE 2014, 9, 1–8. [Google Scholar] [CrossRef]
- Huang, C.; Dong, B.; Zhang, Y.; Wu, H.; Liu, Q. Association of cognitive impairment with smoking, alcohol consumption, tea consumption, and exercise among Chinese nonagenarians/centenarians. Cogn. Behav. Neurol. 2009, 22, 190–196. [Google Scholar] [CrossRef]
- Yue, W.; Sun, W.; Rao, R.S.P.; Ye, N.; Yang, Z.; Chen, M. Non-targeted metabolomics reveals distinct chemical compositions among different grades of Bai Mudan white tea. Food Chem. 2019, 277, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, P.; Liu, P.; Song, X.; Guo, F.; Li, Y.; Ni, D.; Jiang, C. Novel insight into the role of withering process in characteristic flavor formation of teas using transcriptome analysis and metabolite profiling. Food Chem. 2019, 272, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.R.; Alves, M.G.; Tomas, G.D.; Socorro, S.; Silva, B.M.; Oliveira, P. White tea as a promising antioxidant medium additive for sperm storage at room temperature: A comparative study with green tea. J. Agric Food Chem. 2014, 62, 608–617. [Google Scholar] [CrossRef]
- Zhao, L.; La, V.D.; Grenier, D. Antibacterial, antiadherence, antiprotease, and anti-inflammatory activities of various tea extracts: Potential benefits for periodontal diseases. J. Med. Food 2013, 16, 428–436. [Google Scholar] [CrossRef]
- Crispo, J.A.; Ansell, D.R.; Piche, M.; Eibl, J.K.; Khaper, N.; Ross, G.M.; Tai, T.C. Protective effects of polyphenolic compounds on oxidative stress-induced cytotoxicity in PC12 cells. Can. J. Physiol. Pharmacol. 2010, 88, 429–438. [Google Scholar] [CrossRef]
- Shi, X.; Ye, J.; Leonard, S.S.; Ding, M.; Vallyathan, V.; Castranova, V.; Rojanasakul, Y.; Dong, Z. Antioxidant properties of (-)-epicatechin-3-gallate and its inhibition of Cr(VI)-induced DNA damage and Cr(IV)- or TPA-stimulated NF-kappaB activation. Mol. Cell Biochem. 2000, 206, 125–132. [Google Scholar] [CrossRef]
- Carloni, P.; Tiano, L.; Padella, L.; Bacchetti, T.; Customu, C.; Kay, A.; Damiani, E. Antioxidant activity of white, green and black tea obtained from the same tea cultivar. Food Res. Int. 2013, 53, 900–908. [Google Scholar] [CrossRef]
- Ranzato, E.; Martinotti, S.; Magnelli, V.; Murer, B.; Biffo, S.; Mutti, L.; Burlando, B. Epigallocatechin-3-gallate induces mesothelioma cell death via H2O2−dependent T-type Ca2+ channel opening. J. Cell Mol. Med. 2012, 16, 2667–2678. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Y.; Sun, W.; Zhang, M.; Zheng, Y.; Shen, H.; Yang, J.; Zhu, X. FBS or BSA inhibits EGCG induced cell death through covalent binding and the reduction of intracellular ROS production. Biomed Res. Int. 2016, 2016, 5013409. [Google Scholar] [CrossRef]
- Jimenezaliaga, K.L.; Bermejobescos, P.; Benedi, J.; Martinaragon, S. Quercetin and rutin exhibit antiamyloidogenic and fibril-disaggregating effects in vitro and potent antioxidant activity in APPswe cells. Life Sci. 2011, 89, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.R.; Chang, J.Y.; Chang, H.C. Production of gamma-aminobutyric acid (GABA) by Lactobacillus buchneri isolated from kimchi and its neuroprotective effect on neuronal cells. J. Microbiol. Biotechnol. 2007, 17, 104–109. [Google Scholar] [CrossRef]
- Ehrnhoefer, D.E.; Bieschke, J.; Boeddrich, A.; Herbst, M.; Masino, L.; Lurz, R.; Engemann, S.; Pastore, A.; Wanker, E.E. EGCG redirects amyloidogenic polypeptides into unstructured, off-pathway oligomers. Nat. Struct. Mol. Biol. 2008, 15, 558–566. [Google Scholar] [CrossRef]
- Bieschke, J.; Russ, J.; Friedrich, R.P.; Ehrnhoefer, D.E.; Wobst, H.; Neugebauer, K.; Wanker, E.E. EGCG remodels mature alpha-synuclein and amyloid-beta fibrils and reduces cellular toxicity. Proc. Natl. Acad. Sci. USA 2010, 107, 7710–7715. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.; VanSchouwen, B.; Jafari, N.; Ni, X.; Ortega, J.; Melacini, G. Molecular mechanism for the (−)-epigallocatechin gallate-induced toxic to nontoxic remodeling of Aβ oligomers. J. Am. Chem. Soc. 2017, 139, 13720–13734. [Google Scholar] [CrossRef]
- Dai, W.; Xie, D.; Lu, M.; Li, P.; Lv, H.; Yang, C.; Peng, Q.; Zhu, Y.; Guo, L.; Zhang, Y. Characterization of white tea metabolome: Comparison against green and black tea by a nontargeted metabolomics approach. Food Res. Int. 2017, 96, 40–45. [Google Scholar] [CrossRef]
- Liu, Y.; Pukala, T.L.; Musgrave, I.F.; Williams, D.M.; Dehle, F.C.; Carver, J.A. Gallic acid is the major component of grape seed extract that inhibits amyloid fibril formation. Bioorg. Med. Chem. Lett. 2013, 23, 6336–6340. [Google Scholar] [CrossRef]
- He, H. Research progress on theaflavins: Efficacy, formation, and preparation. Food Nutr. Res. 2017, 61, 1344521. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, Y.; Wang, P.; Ahmedna, M.; Ho, C.T.; Sang, S. Tea flavanols block advanced glycation of lens crystallins induced by dehydroascorbic acid. Chem. Res. Toxicol. 2015, 28, 135–143. [Google Scholar] [CrossRef]
- Wang, W.; Fu, X.; Dai, X.; Hua, F.; Chu, G.; Chu, M.; Hu, F.; Ling, T.; Gao, L.; Xie, Z. Novel acetylcholinesterase inhibitors from Zijuan tea and biosynthetic pathway of caffeoylated catechin in tea plant. Food Chem. 2017, 237, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, C.; Idehen, E.; Shi, L.; Lv, L.; Sang, S. Novel theaflavin-type chlorogenic acid derivatives identified in black tea. J. Agric. Food Chem. 2018, 66, 3402–3407. [Google Scholar] [CrossRef]
- Dai, W.; Tan, J.; Lu, M.; Zhu, Y.; Li, P.; Peng, Q.; Guo, L.; Zhang, Y.; Xie, D.; Hu, Z.; Lin, Z. Metabolomics Investigation reveals That 8-C N-ethyl-2-pyrrolidinone-substituted flavan-3-ols are potential marker compounds of stored white teas. J. Agric. Food Chem. 2018, 66, 7209–7218. [Google Scholar] [CrossRef] [PubMed]
- Guzzi, C.; Colombo, L.; De Luigi, A.; Salmona, M.; Nicotra, F.; Airoldi, C. Flavonoids and their glycosides as anti-amyloidogenic compounds: Aβ1–42 interaction studies to gain new insights into their potential for Alzheimer’s Disease prevention and therapy. Chem. Asian J. 2017, 12, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Choi, R.C.Y.; Zhu, J.T.T.; Leung, K.W.; Chu, G.K.Y.; Xie, H.Q.; Chen, V.P.; Zheng, K.Y.Z.; Lau, D.T.W.; Dong, T.T.X.; Chow, P.C.Y. A flavonol glycoside, isolated from roots of Panax notoginseng, reduces amyloid-β-induced neurotoxicity in cultured neurons: Signaling transduction and drug development for Alzheimer’s disease. J. Alzheimers Dis. 2010, 19, 795–811. [Google Scholar] [CrossRef]
- Li, P.; Zhu, Y.; Lu, M.; Yang, C.; Xie, D.; Tan, J.; Peng, Q.; Zhang, Y.; Ni, D.; Dai, W.; Lin, Z. Variation patterns in the content of glycosides during green tea manufacturing by a modification-specific metabolomics approach: Enzymatic reaction promoting an increase in the glycosidically bound volatiles at the pan firing stage. Food Chem. 2019, 279, 80–87. [Google Scholar] [CrossRef]
- Tan, J.; Dai, W.; Lu, M.; Lv, H.; Guo, L.; Zhang, Y.; Zhu, Y.; Peng, Q.; Lin, Z. Study of the dynamic changes in the non-volatile chemical constituents of black tea during fermentation processing by a non-targeted metabolomics approach. Food Res. Int. 2016, 79, 106–113. [Google Scholar] [CrossRef]
- Fraser, K.; Lane, G.A.; Otter, D.E.; Harrison, S.J.; Quek, S.Y.; Hemar, Y.; Rasmussen, S. Non-targeted analysis by LC-MS of major metabolite changes during the oolong tea manufacturing in New Zealand. Food Chem. 2014, 151, 394–403. [Google Scholar] [CrossRef]
- Xing, S.G.; Jun, Y.B.; Hau, Z.W.; Liang, L.Y. Higher accumulation of γ-aminobutyric acid induced by salt stress through stimulating the activity of diamine oxidases in Glycine max (L.) Merr. roots. Plant Physiol. Biochem. 2007, 45, 560–566. [Google Scholar] [CrossRef]
- Deewatthanawong, R.; Rowell, P.; Watkins, C.B. γ-Aminobutyric acid (GABA) metabolism in CO2 treated tomatoes. Postharvest Biol. Technol. 2010, 57, 97–105. [Google Scholar] [CrossRef]
- Hou, C. Pu-Erh tea and GABA attenuates oxidative stress in kainic acid-induced status epilepticus. J. Biomed. Sci. 2011, 18, 75. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, D. Amyloid-β impairs synaptic inhibition via GABAA receptor endocytosis. J. Neurosci. 2015, 35, 9205–9210. [Google Scholar] [CrossRef]
- Limon, A.; Reyesruiz, J.M.; Miledi, R. Loss of functional GABA(A) receptors in the Alzheimer diseased brain. Proc. Natl. Acad. Sci. USA 2012, 109, 10071–10076. [Google Scholar] [CrossRef]
- Sun, X.; Meng, X.; Zhang, J.; Li, Y.; Wang, L.; Qin, X.; Sui, N.; Zhang, Y. GABA attenuates amyloid toxicity by downregulating its endocytosis and improves cognitive impairment. J. Alzheimers Dis. 2012, 31, 635–649. [Google Scholar] [CrossRef]
- Wang, H.; Tang, C.; Jiang, Z.; Zhou, X.; Chen, J.; Na, M.; Shen, H.; Lin, Z. Glutamine promotes Hsp70 and inhibits α-Synuclein accumulation in pheochromocytoma PC12 cells. Exp. Ther. Med. 2017, 14, 1253–1259. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tsai, Y.; Lee, Y.; Lam, K.; Lin, J.; Wang, J.; Yen, M.H.; Cheng, P. The role of heat shock protein 70 in the protective effect of YC-1 on β-amyloid-induced toxicity in differentiated PC12 cells. PLoS ONE 2013, 8, e69320. [Google Scholar] [CrossRef]
- Chen, J.; Herrup, K. Glutamine acts as a neuroprotectant against DNA damage, beta-amyloid and H2O2-induced stress. PLoS ONE 2012, 7, e33177. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, N.; Jana, N.R.; Jana, N.R. Inhibition of protein aggregation by iron oxide nanoparticles conjugated with glutamine- and proline-based osmolytes. ACS Appl. Nano Mater. 2018, 1, 1094–1103. [Google Scholar] [CrossRef]
- Li, C.; Yao, M.; Ma, C.; Ma, J.; Jin, J.; Chen, L. Differential metabolic profiles during the Albescent stages of ‘Anji Baicha’ (Camellia sinensis). PLoS ONE 2015, 10, e69320. [Google Scholar] [CrossRef]
- Chen, S.; Lin, J.; Liu, H.; Gong, Z.; Wang, X.; Li, M.; Aharoni, A.; Yang, Z.; Yu, X. Insights into tissue-specific specialized metabolism in Tieguanyin Tea cultivar by untargeted metabolomics. Molecules 2018, 23, 1817. [Google Scholar] [CrossRef]
- Dixon, D.N.; Loxley, R.A.; Barron, A.M.; Cleary, S.; Phillips, J.K. Comparative studies of PC12 and mouse pheochromocytoma-derived rodent cell lines as models for the study of neuroendocrine systems. In Vitro Cell. Dev. Biol. Anim. 2005, 41, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, Q.; Wang, W.; Li, X.; Zhang, J. A polysaccharide isolated from Cynomorium songaricum Rupr. protects PC12 cells against H2O2-induced injury. Int. J. Biol. Macromol. 2016, 87, 222–228. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, L.; Guo, X.; Li, C.; Li, H.; Lou, H.; Ren, D. Chemical constituents from Phyllanthus emblica and the cytoprotective effects on H2O2-induced PC12 cell injuries. Arch. Pharm. Res. 2016, 39, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, M.; Zheng, G.; Wang, T.; Lin, J.; Wang, S.; Wang, X.; Chao, Q.; Cao, S.; Yang, Z. Metabolite profiling of 14 Wuyi rock tea cultivars using UPLC-QTOF MS and UPLC-QqQ MS combined with chemometrics. Molecules 2018, 23, 104. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Tentative Assignment | RT (min) | Detected [M − H]− (m/z) | Theoretical [M − H]− (m/z) | Mass Error (ppm) | Formula | MS/MS Fragments |
|---|---|---|---|---|---|---|---|
| 1 | EGCG a | 6.34 | 457.0816 | 457.0771 | −3.28 | C22H18O11 | 457.0781, 305.0669, 169.0147 |
| 2 | ECG a | 7.83 | 441.0832 | 441.0822 | −2.27 | C22H18O10 | 289.0720, 169.0144 |
| 3 | C a | 5.36 | 289.072 | 289.0712 | −2.77 | C15H14O6 | 245.0816, 203.0709 |
| 4 | EGC a | 4.92 | 305.0704 | 305.0661 | −3.93 | C15H14O7 | 305.0672, 219.0661 |
| 5 | EC a | 6.25 | 289.0723 | 289.0712 | −3.81 | C15H14O6 | 245.082, 203.0711 |
| 6 | GC a | 3.85 | 305.0667 | 305.0661 | −1.97 | C15H14O7 | 219.0659, 125.0242 |
| 7 | ECG3’’Me | 8.88 | 455.0983 | 455.0978 | −1.10 | C23H20O10 | 290.0750, 183.0298 |
| 8 | EGCG3’’Me a | 7.41 | 471.0933 | 471.0927 | −1.27 | C23H20O11 | 287.0534, 183.0298 |
| 9 | Epiafzelechin-3-Gallate | 8.93 | 425.0878 | 425.0873 | −1.25 | C22H18O9 | 125.0243, 137.0245, 273.0761, 169.0140, 151.0029, |
| 10 | 8-C-Ascorbyl-EGCG | 6.66 | 631.0945 | 631.0935 | −1.58 | C28H24O17 | 479.0816, 423.0706, 316.0225 |
| 11 | Procy Dimer (B type) | 5.68 | 577.1353 | 577.1346 | −1.21 | C30H26O12 | 425.0872, 407.0768, 289.0716 |
| 12 | Prodel B-2 (or 4) 3’-O-Gallate | 5.13 | 761.1356 | 761.1354 | −0.26 | C37H30O18 | 423.0718, 609.1236, 591.1135, 577.1348, |
| 13 | Prodel B Isomer 2 | 4.13 | 609.1249 | 609.1244 | −0.82 | C30H26O14 | 441.0827, 423.0717, 305.0665 |
| 14 | EC-GC Dimer | 4.81 | 593.1300 | 593.1295 | −0.84 | C30H26O13 | 423.0712 |
| 15 | EGC-ECG Dimer | 6.03 | 745.1407 | 745.1405 | −0.27 | C37H30O17 | 593.1300, 407.0767 |
| 16 | 3-Galloyl-Procy B1/3’-Galloyl-Procy B2 | 6.78 | 729.1458 | 729.1456 | −0.27 | C37H30O16 | 577.1228, 407.0768, 289.0715 |
| 17 | Kae-3-Glu a | 8.76 | 447.0925 | 447.0927 | 0.45 | C21H20O11 | 255.0298, 227.0348 |
| 18 | Kae-3-Rut | 8.42 | 593.1509 | 593.1506 | −0.51 | C27H30O15 | 285.0399 |
| 19 | Kae-3-Glu-Rut | 8.00 | 755.2038 | 755.2035 | −0.40 | C33H40O20 | 285.0404 |
| 20 | Que-3-Glu | 8.00 | 463.0879 | 463.0877 | −0.43 | C21H20O12 | 300.0275, 271.0252 |
| 21 | Que-3-Rut a | 7.69 | 609.1445 | 609.1456 | 1.81 | C27H30O16 | 271.0248 |
| 22 | Que-3-Glu-Rut | 7.35 | 771.1988 | 771.1984 | −0.52 | C33H40O21 | 609.1461, 301.0348 |
| 23 | Myr-3’-Glu | 7.11 | 479.0819 | 479.0826 | 1.46 | C21H20O13 | 287.0201, 271.0247 |
| 24 | Myr-3-Rob (or 3-Neo) | 6.93 | 625.1400 | 625.1405 | 0.80 | C27H30O17 | 527.1541, 307.1396 |
| 25 | Methyl-6-Galloyl-β-d-Glu | 3.68 | 345.0818 | 345.0822 | 1.16 | C14H18O10 | 285.0611, 225.0401, 183,0296 |
| 26 | Monogalloyl Glu | 2.49 | 331.0669 | 331.0665 | −1.21 | C13H16O10 | 169.0142 |
| 27 | Vitexin-2’’-Rha | 7.67 | 577.1551 | 577.1557 | 1.04 | C27H30O14 | 323.0557, 282.0533 |
| 28 | Api-6-C-Glu 8-C-Ara | 6.90 | 563.1405 | 563.1401 | −0.71 | C26H28O14 | 473.1089, 383.0770, 353.0665 |
| 29 | Naringenin Diglucose | 6.13 | 595.1660 | 595.1663 | 0.50 | C27H32O15 | 475.1245, 433.1347, 313.0925 |
| 30 | Eri-5,3’-Diglucose | 6.05 | 611.1617 | 611.1612 | −0.82 | C27H32O16 | 491.1194, 449.1288, 329.0868 |
| 31 | 3-p-Coumaroylquinic Acid | 5.16 | 337.0930 | 337.0923 | −2.08 | C16H18O8 | 191.0559, 163.0399, 119.0500 |
| 32 | 4-p-Coumaroylquinic Acid | 6.12 | 337.0930 | 337.0923 | −2.08 | C16H18O8 | 173.0455, 163.0397, 1193.0502 |
| 33 | 5-p-Coumaroylquinic Acid | 6.40 | 337.0931 | 337.0923 | −2.37 | C16H18O8 | 173.0460, 163.0400, 119.0502 |
| 34 | Chlorogenic Acid | 5.51 | 353.0877 | 353.0873 | −1.13 | C16H18O9 | 191.0558, 179.0349, 173.0455, 135.0449 |
| 35 | Theogallin | 2.94 | 343.0666 | 343.0665 | −0.29 | C14H16O10 | 343.0670, 191.0562 |
| 36 | Galic Acid a | 2.78 | 169.0142 | 169.0137 | −2.96 | C7H6O5 | 125.0242, 95.0138 |
| 37 | TF1 | 10.57 | 563.1196 | 563.1190 | −1.07 | C29H24O12 | 425.092 |
| 38 | TF2a | 10.94 | 715.1303 | 715.1299 | −0.56 | C36H28O16 | 563.119 |
| 39 | TF2b | 11.18 | 715.1301 | 715.1299 | −0.28 | C36H28O16 | 289.0721, 245.0820 |
| 40 | TF3 | 11.12 | 867.1416 | 867.1409 | −0.81 | C43H32O20 | 867.1414 |
| 41 | l-Theanine | 1.36 | 173.0932 | 173.0926 | −3.58 | C7H14N2O3 | 155.0830, 128.0354 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Smid, S.D.; Lin, J.; Gong, Z.; Chen, S.; You, F.; Zhang, Y.; Hao, Z.; Lin, H.; Yu, X.; et al. Neuroprotective and Anti-Amyloid β Effect and Main Chemical Profiles of White Tea: Comparison Against Green, Oolong and Black Tea. Molecules 2019, 24, 1926. https://doi.org/10.3390/molecules24101926
Li X, Smid SD, Lin J, Gong Z, Chen S, You F, Zhang Y, Hao Z, Lin H, Yu X, et al. Neuroprotective and Anti-Amyloid β Effect and Main Chemical Profiles of White Tea: Comparison Against Green, Oolong and Black Tea. Molecules. 2019; 24(10):1926. https://doi.org/10.3390/molecules24101926
Chicago/Turabian StyleLi, Xinlei, Scott D. Smid, Jun Lin, Zhihong Gong, Si Chen, Fangning You, Yan Zhang, Zhilong Hao, Hongzheng Lin, Xiaomin Yu, and et al. 2019. "Neuroprotective and Anti-Amyloid β Effect and Main Chemical Profiles of White Tea: Comparison Against Green, Oolong and Black Tea" Molecules 24, no. 10: 1926. https://doi.org/10.3390/molecules24101926
APA StyleLi, X., Smid, S. D., Lin, J., Gong, Z., Chen, S., You, F., Zhang, Y., Hao, Z., Lin, H., Yu, X., & Jin, X. (2019). Neuroprotective and Anti-Amyloid β Effect and Main Chemical Profiles of White Tea: Comparison Against Green, Oolong and Black Tea. Molecules, 24(10), 1926. https://doi.org/10.3390/molecules24101926