
Anti-Inflammatory Compounds from Atractylodes macrocephala


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
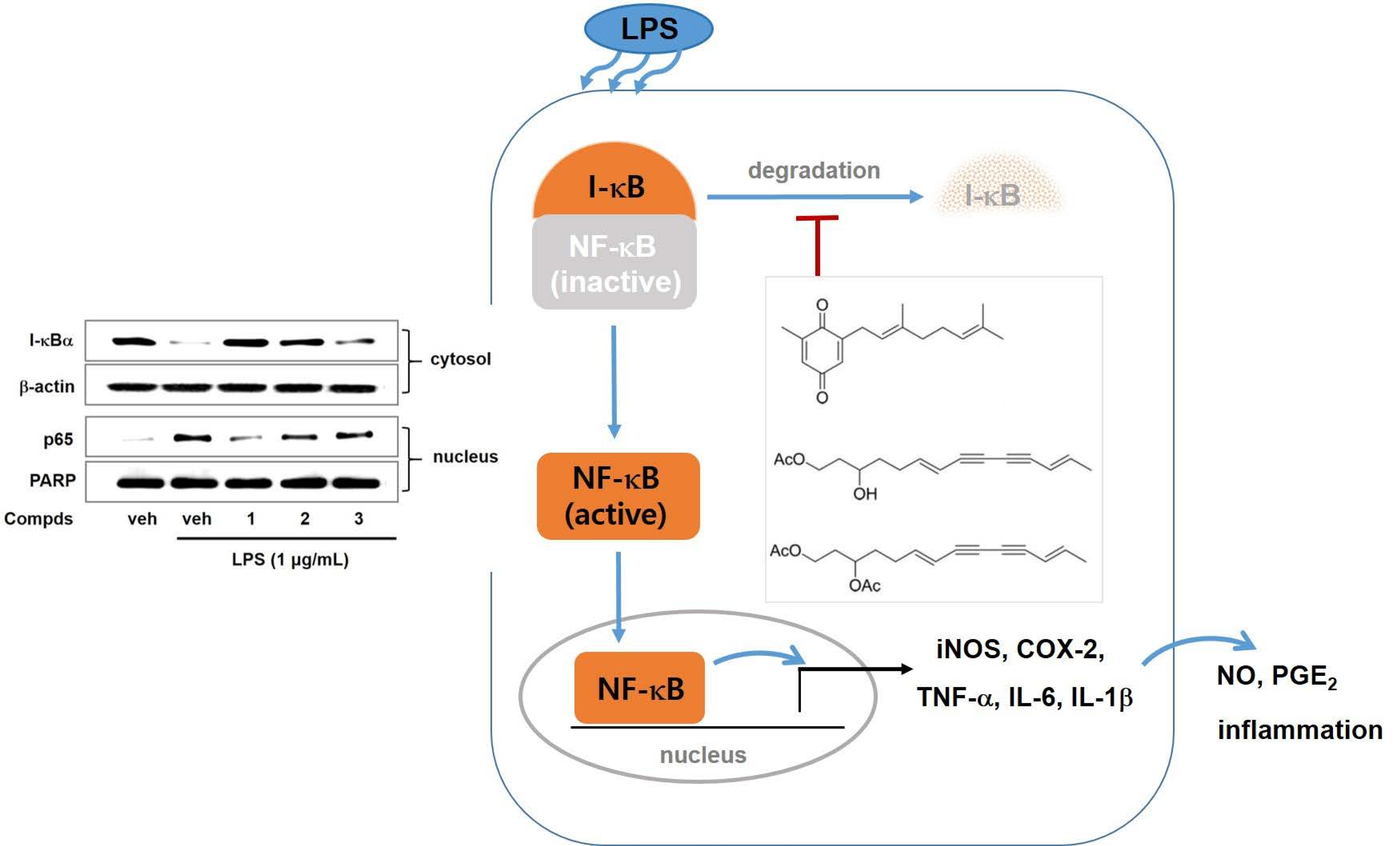
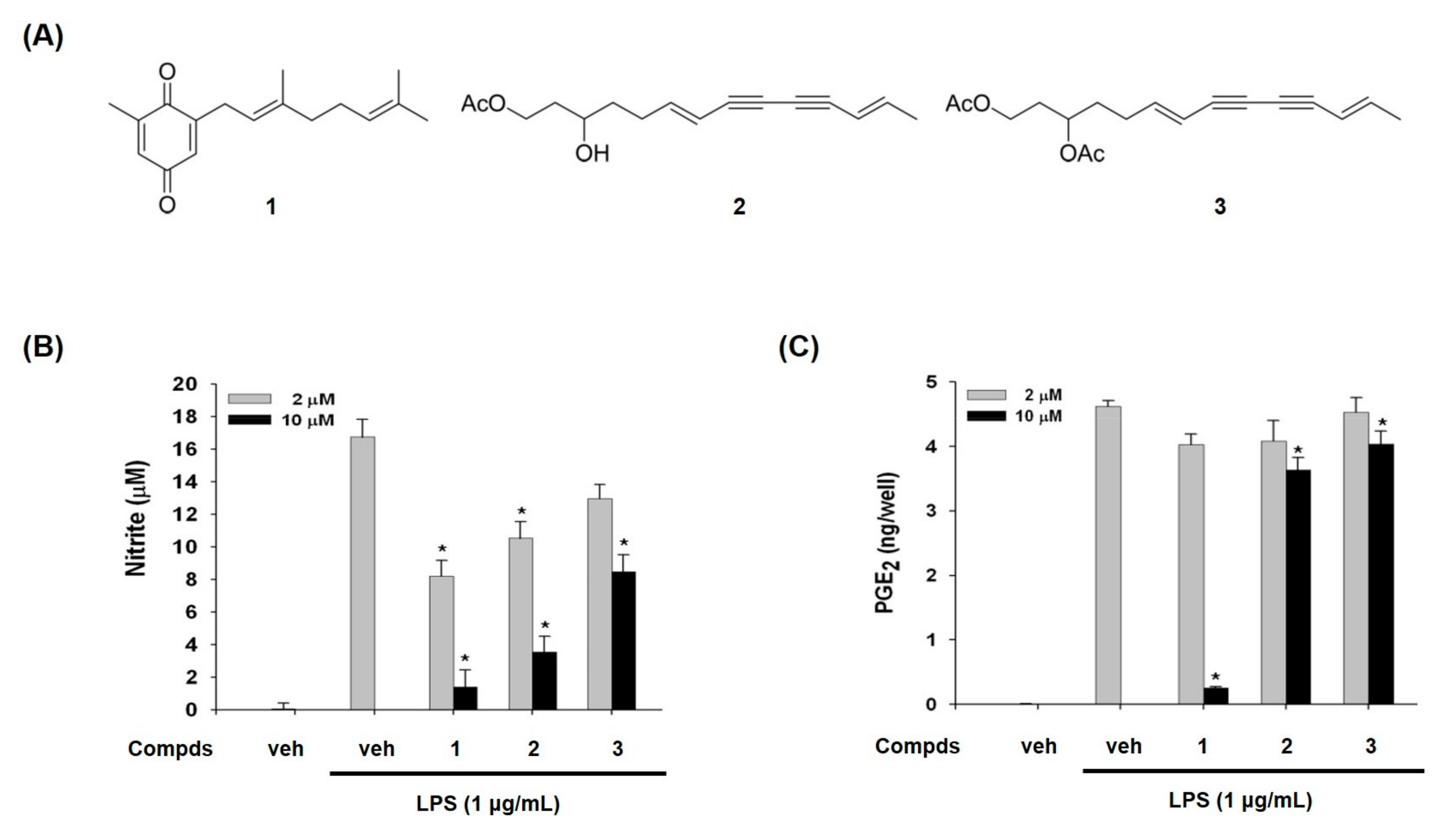
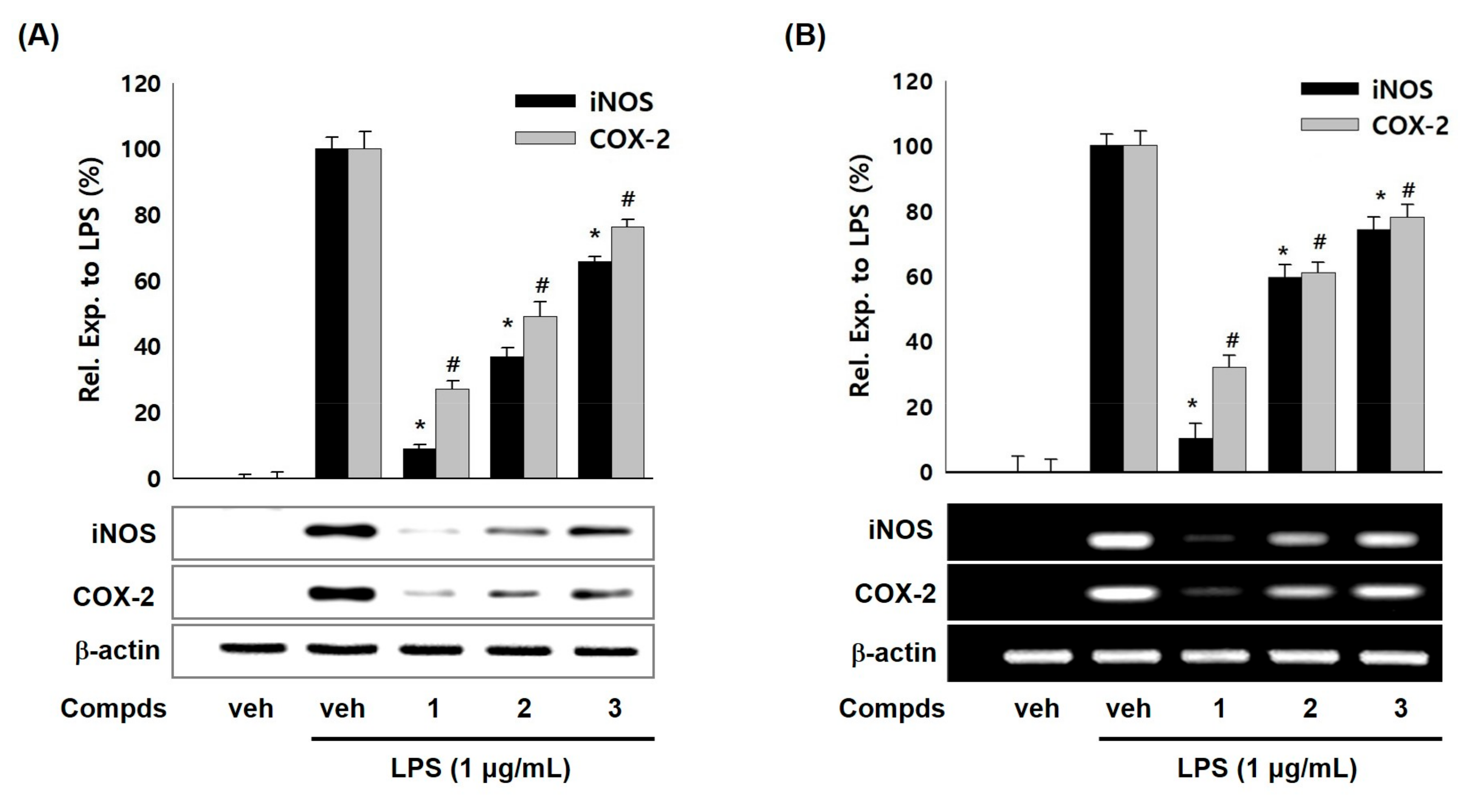
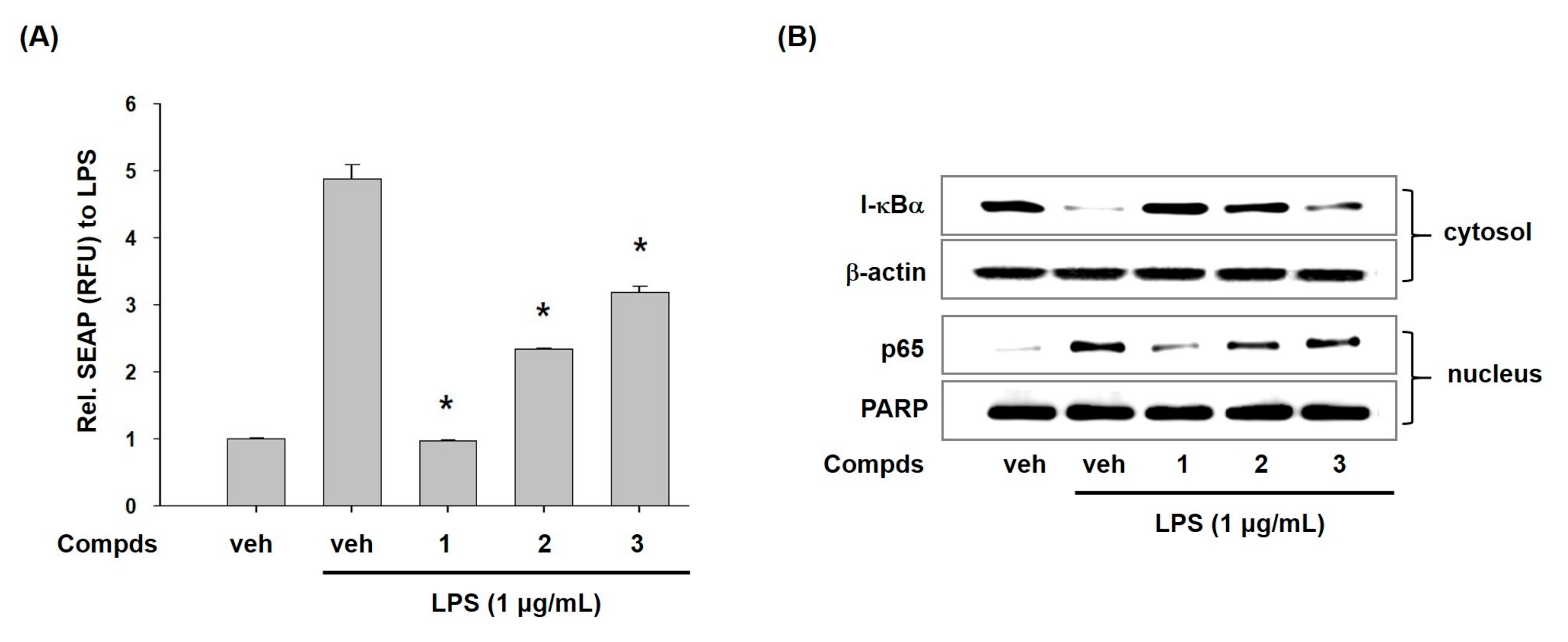
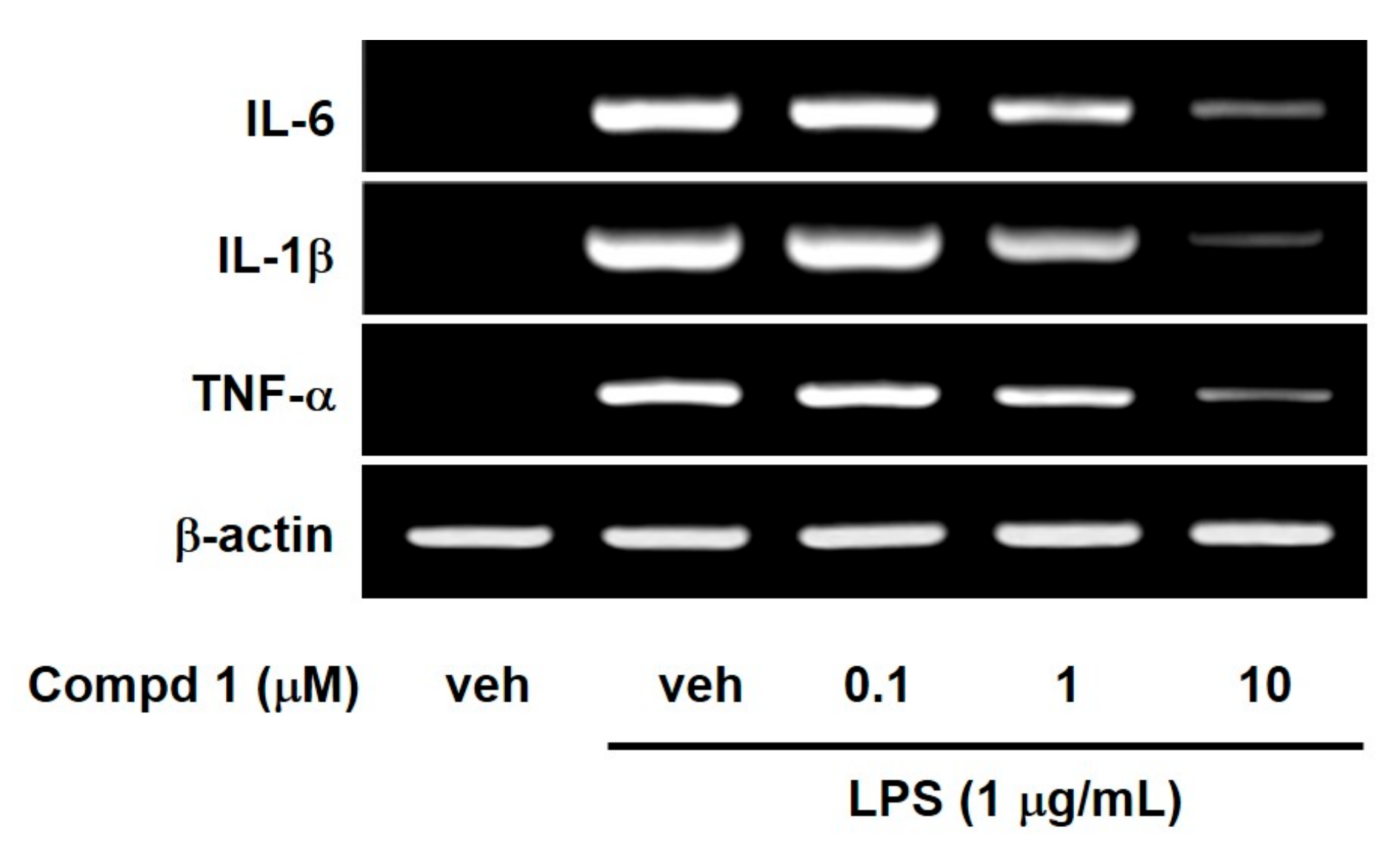
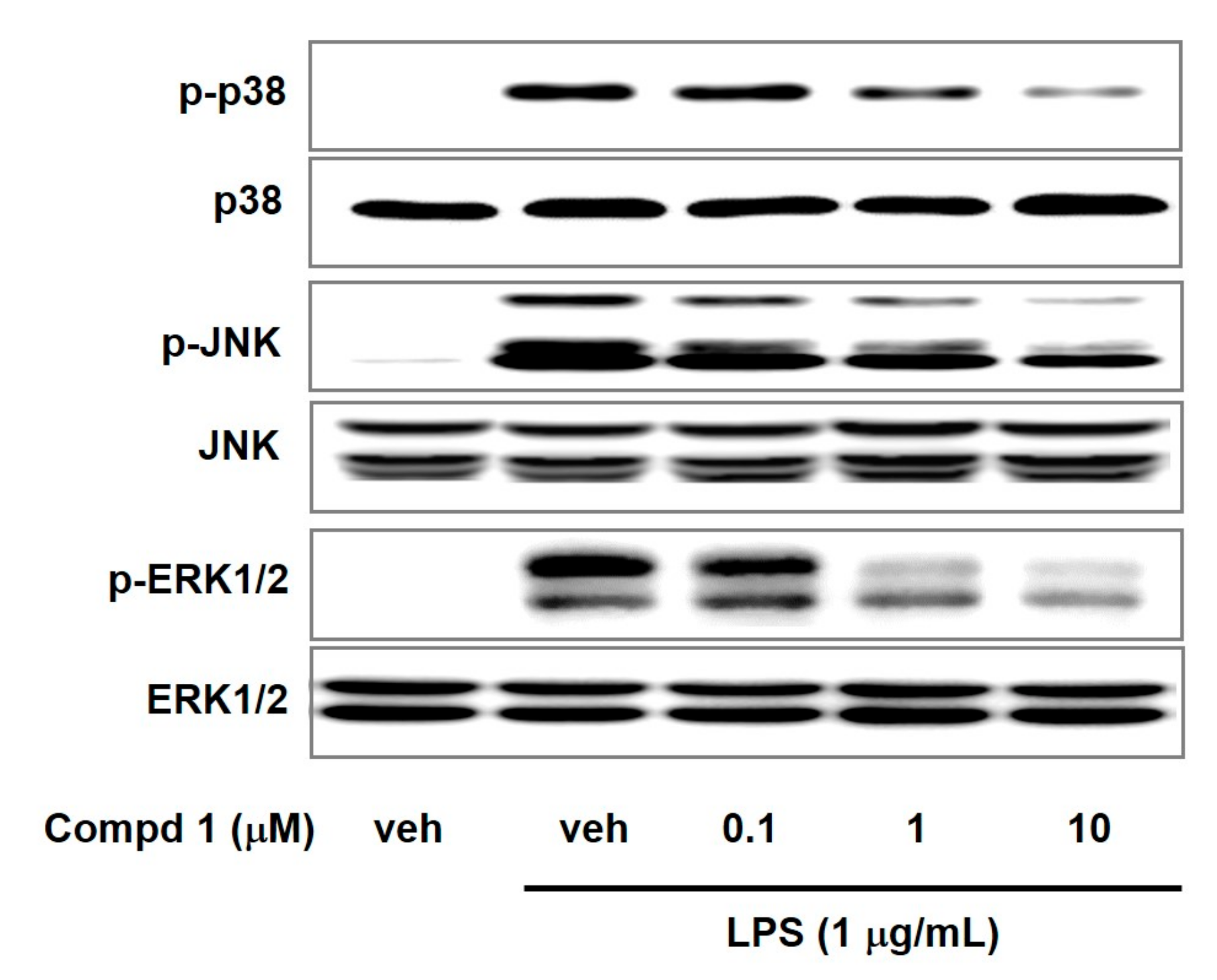
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Materials, Extraction and Isolation
3.3. Cell Culture
3.4. Nitrite Assay
3.5. PGE2 Assay
3.6. Western Blot Analysis
3.7. Reverse Transcription and Polymerase Chain Reaction (RT-PCR) Analysis
3.8. Measurement of NF-κB Transcriptional Activity
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kim, J.H.; Doh, E.J.; Lee, G. Evaluation of Medicinal Categorization of Atractylodes japonica Koidz. by Using Internal Transcribed Spacer Sequencing Analysis and HPLC Fingerprinting Combined with Statistical Tools. Evid. Based Complement. Alternat. Med. 2016, 2016, 2926819. [Google Scholar] [CrossRef] [PubMed]
- Li, L.S.; Qu, R.Y.; Wang, W.; Guo, H. Significance of changes of gastrointestinal peptides in blood and ileum of experimental spleen deficiency rats. World J. Gastroenterol. 2003, 9, 553–556. [Google Scholar] [CrossRef]
- Wang, R.; Zhou, G.; Wang, M.; Peng, Y.; Li, X. The Metabolism of Polysaccharide from Atractylodes macrocephala Koidz and Its Effect on Intestinal Microflora. Evid. Based Complement. Alternat. Med. 2014, 2014, 926381. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, F.; Ye, N.; Huang, C.; Yuan, X. Atractylodes macrocephala polysaccharides induces mitochondrial-mediated apoptosis in glioma C6 cells. Int. J. Biol. Macromol. 2014, 66, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Duan, H.; He, L. Inhibitory effect of atractylenolide I on angiogenesis in chronic inflammation in vivo and in vitro. Eur. J. Pharmacol. 2009, 612, 143–152. [Google Scholar] [CrossRef]
- Li, X.; Lin, J.; Han, W.; Mai, W.; Wang, L.; Li, Q.; Lin, M.; Bai, M.; Zhang, L.; Chen, D. Antioxidant ability and mechanism of rhizoma Atractylodes macrocephala. Molecules 2012, 17, 13457–13472. [Google Scholar] [CrossRef]
- Shu, Y.T.; Kao, K.T.; Weng, C.S. In vitro antibacterial and cytotoxic activities of plasma-modified polyethylene terephthalate nonwoven dressing with aqueous extract of Rhizome Atractylodes macrocephala. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 77, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Zhang, Q.L.; Hua, J.W.; Cheng, W.L.; Qin, L.P. The traditional uses, phytochemistry, and pharmacology of Atractylodes macrocephala Koidz.: A review. J. Ethnopharmacol. 2018, 226, 143–167. [Google Scholar] [CrossRef]
- Zhang, N.; Liu, C.; Sun, T.M.; Ran, X.K.; Kang, T.G.; Dou, D.Q. Two new compounds from Atractylodes macrocephala with neuroprotective activity. J. Asian Nat. Prod. Res. 2017, 19, 35–41. [Google Scholar] [CrossRef]
- Hoang, L.S.; Tran, M.H.; Lee, J.S.; Ngo, Q.M.; Woo, M.H.; Min, B.S. Inflammatory inhibitory activity of sesquiterpenoids from Atractylodes macrocephala rhizomes. Chem. Pharm. Bull. 2016, 64, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Q.; He, L.C.; Jin, J.Q. Atractylenolide I and atractylenolide III inhibit lipopolysaccharide-induced TNF-alpha and NO production in macrophages. Phytother. Res. 2007, 21, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.M.; Yang, X.W. Bioactivity-guided isolation of polyacetylenes with inhibitory activity against NO production in LPS-activated RAW264.7 macrophages from the rhizomes of Atractylodes macrocephala. J. Ethnopharmacol. 2014, 151, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Q.; He, L.C.; Dong, H.Y.; Jin, J.Q. Screening for the anti-inflammatory activity of fractions and compounds from Atractylodes macrocephala koidz. J. Ethnopharmacol. 2007, 114, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Murata, M. Inflammation and cancer. Environ. Health Prev. Med. 2018, 23, 50. [Google Scholar] [CrossRef] [PubMed]
- Bastard, J.P.; Jaachi, M.; Laqathu, C.; Kim, M.J.; Caron, M.; Vidal, H.; Capeau, J.; Feve, B. Recent advances in the relationship between obesity, inflammation, and insulin resistance. Eur. Cytokine Netw. 2006, 17, 4–12. [Google Scholar] [PubMed]
- De Virgilio, A.; Greco, A.; Fabbrini, G.; Inqhilleri, M.; Rizzo, M.I.; Gallo, A.; Conte, M.; Rosato, C.; Ciniqlio Appiani, M.; de Vincentiis, M. Parkinson’s disease: Autoimmunity and neuroinflammation. Autoimmun. Rev. 2016, 15, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Hamidzadeh, K.; Christensen, S.M.; Dalby, E.; Chandrasekaran, P.; Mosser, D.M. Macrophages and the Recovery from Acute and Chronic Inflammation. Annu. Rev. Physiol. 2017, 79, 567–592. [Google Scholar] [CrossRef]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar] [PubMed]
- Attiq, A.; Jalil, J.; Husain, K.; Ahmad, W. Raging the war against inflammation with natural products. Front. Pharmacol. 2018, 9, 976. [Google Scholar] [CrossRef]
- Lind, M.; Hayes, A.; Capmda, M.; Petrovic, D.; Rodrigo, L.; Kruzliak, P.; Zulli, A. Inducible nitric oxide synthase: Good or bad? Biomed. Pharmacother. 2017, 93, 370–375. [Google Scholar] [CrossRef]
- Kawahara, K.; Hohjoh, H.; Inazumi, T.; Tsuchiya, S.; Sugimoto, Y. Prostaglandin E2-induced inflammation: Relevance of prostaglandin E receptors. Biochim. Biophys. Acta 2015, 1851, 414–421. [Google Scholar] [CrossRef]
- Resch, M.; Steigel, A.; Chen, Z.L.; Bauer, R. 5-Lipoxygenase and cyclooxygenase-1 inhibitory active compounds from Atractylodes lancea. J. Nat. Prod. 1998, 61, 347–350. [Google Scholar] [CrossRef]
- Pachaly, P.; Lansing, A.; Neugebauer, M.; Sin, K.S. Acetylenes from Atractylis koreana. Planta Med. 1990, 56, 469–471. [Google Scholar] [CrossRef]
- Ahn, K.S.; Noh, E.J.; Zhao, H.S.; Jung, S.H.; Kang, S.S.; Kim, Y.S. Inhibition of inducible nitric oxide synthase and cyclooxygenase II by Platycodon grandiflorum saponins via suppression of nuclear factor-kappaB activation in RAW 264.7 cells. Life Sci. 2005, 76, 2315–2328. [Google Scholar] [CrossRef]
- Yamamoto, M.; Takeda, K. Role of nuclear IkappaB proteins in the regulation of host immune responses. J. Infect. Chemother. 2008, 14, 265–269. [Google Scholar] [CrossRef]
- Wan, F.; Lenardo, M.J. The nuclear signaling of NF-kappaB: Current knowledge, new insights, and future perspectives. Cell Res. 2010, 20, 24–33. [Google Scholar] [CrossRef]
- Liang, Y.; Zhou, Y.; Shen, P. NF-kappaB and its regulation on the immune system. Cell. Mol. Immunol. 2004, 1, 343–350. [Google Scholar]
- Chan, E.D.; Riches, D.W. IFN-gamma + LPS induction of iNOS is modulated by ERK, JNK/SAPK, and p38(mapk) in a mouse macrophage cell line. Am. J. Physiol. Cell Physiol. 2001, 280, C441–C450. [Google Scholar] [CrossRef]
- Chan, E.D.; Winston, B.W.; Uh, S.T.; Wynes, M.W.; Rose, D.M.; Riches, D.W. Evaluation of the role of mitogen-activated protein kinases in the expression of inducible nitric oxide synthase by IFN-gamma and TNF-alpha in mouse macrophages. J. Immunol. 1999, 162, 415–422. [Google Scholar]
- Kyriakis, J.M. Activation of the AP-1 transcription factor by inflammatory cytokines of the TNF family. Gene Expr. 1999, 7, 217–231. [Google Scholar]
- Mahtani, K.R.; Brook, M.; Dean, J.S.; Sully, G.; Saklatvala, J.; Clark, A.R. Mitogen-activated protein kinase p38 controls the expression and posttranslational modification of tristetraprolin, a regulator of tumor necrosis factor alpha mRNA stability. Mol. Cell. Biol. 2001, 21, 6461–6469. [Google Scholar] [CrossRef]
- Karunakaran, S.; Ravindranath, V. Activation of p38 MAPK in the substantia nigra leads to nuclear translocation of NF-kappaB in MPTP-treated mice: Implication in Parkinson’s disease. J. Neurochem. 2009, 109, 1791–1799. [Google Scholar] [CrossRef]
- Lee, S.M.; Kim, E.J.; Suk, K.; Lee, W.H. Stimulation of Fas (CD95) induces production of pro-inflammatory mediators through ERK/JNK-dependent activation of NF-kappaB in THP-1 cells. Cell. Immunol. 2011, 271, 157–162. [Google Scholar] [CrossRef]
- Li, L.; Zhao, R.; Li, Y.; Wang, W.H. Antitumor activity of 2-[(2E)-3,7-dimethyl-2,6-octadienyl]-6-methyl-2,5-cyclohexadiene-1,4-dione isolated from the aerial part of Atractylodes macrocephala in hepatocellular carcinoma. Mol. Med. Rep. 2017, 16, 6299–6305. [Google Scholar] [CrossRef][Green Version]
- Meng, H.; Li, G.; Dai, R.; Ma, Y.; Zhang, K.; Zhang, C.; Li, X.; Wang, J. Chemical constituents of Atractylodes chinensis(DC.) Koidz. Biochem. Syst. Ecol. 2010, 38, 1220–1223. [Google Scholar] [CrossRef]
- Moon, K.Y.; Hahn, B.S.; Lee, J.; Kim, Y.S. A cell-based assay system for monitoring NF-kappaB activity in human HaCat transfectant cells. Anal. Biochem. 2001, 292, 17–21. [Google Scholar] [CrossRef]
- Ahn, K.S.; Aggarwal, B.B. Transcription factor NF-kappaB: A sensor for smoke and stress signals. Ann. N. Y. Acad. Sci. 2005, 1056, 218–233. [Google Scholar] [CrossRef]
Sample Availability: Not available. |





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, D.; Dong, G.-z.; Lee, H.J.; Ryu, J.-H. Anti-Inflammatory Compounds from Atractylodes macrocephala. Molecules 2019, 24, 1859. https://doi.org/10.3390/molecules24101859
Jeong D, Dong G-z, Lee HJ, Ryu J-H. Anti-Inflammatory Compounds from Atractylodes macrocephala. Molecules. 2019; 24(10):1859. https://doi.org/10.3390/molecules24101859
Chicago/Turabian StyleJeong, Dawoon, Guang-zhi Dong, Hwa Jin Lee, and Jae-Ha Ryu. 2019. "Anti-Inflammatory Compounds from Atractylodes macrocephala" Molecules 24, no. 10: 1859. https://doi.org/10.3390/molecules24101859
APA StyleJeong, D., Dong, G.-z., Lee, H. J., & Ryu, J.-H. (2019). Anti-Inflammatory Compounds from Atractylodes macrocephala. Molecules, 24(10), 1859. https://doi.org/10.3390/molecules24101859