
PPARα Agonist Suppresses Inflammation after Corneal Alkali Burn by Suppressing Proinflammatory Cytokines, MCP-1, and Nuclear Translocation of NF-κB

 , and
, and
Abstract
1. Introduction
2. Results
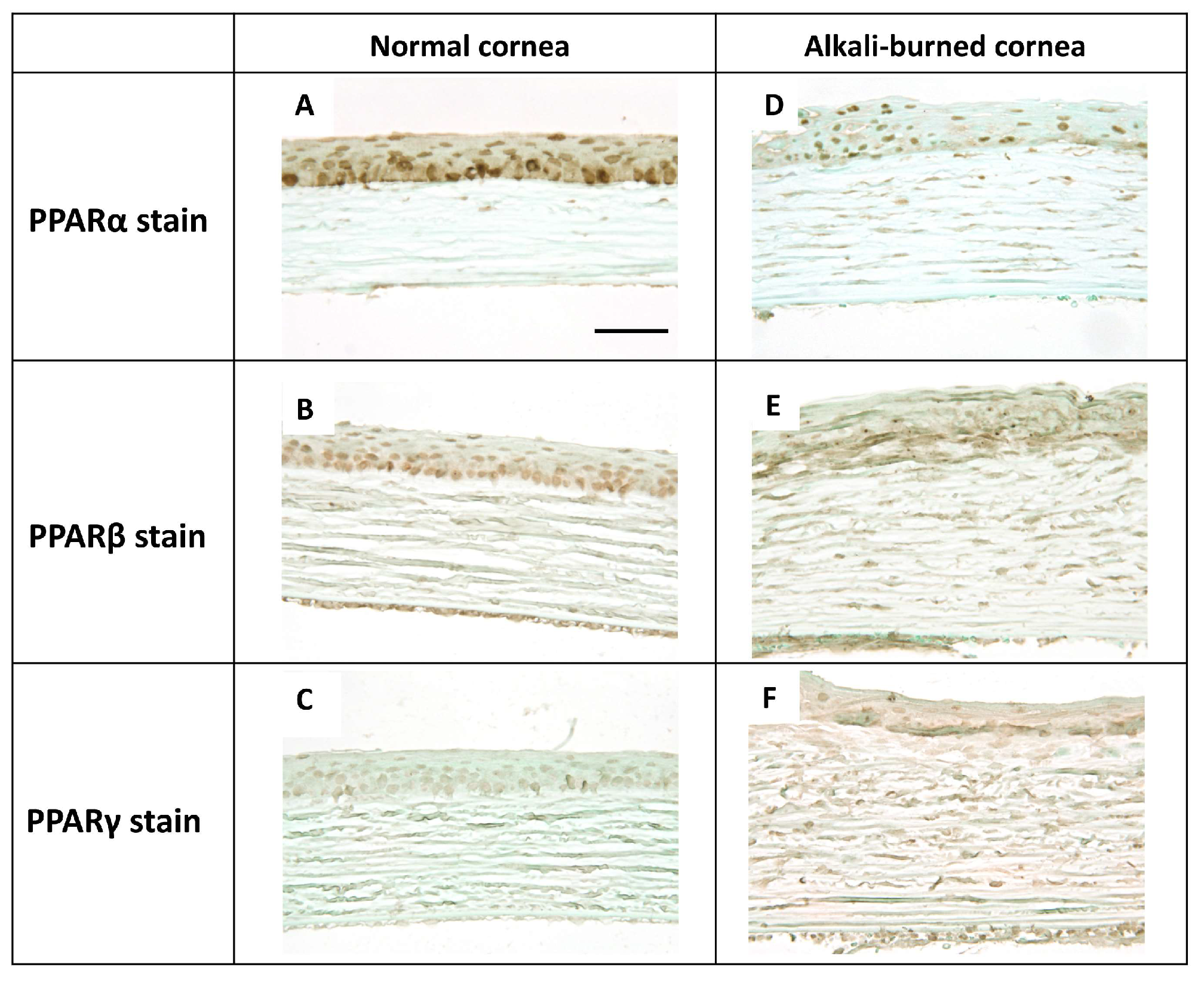
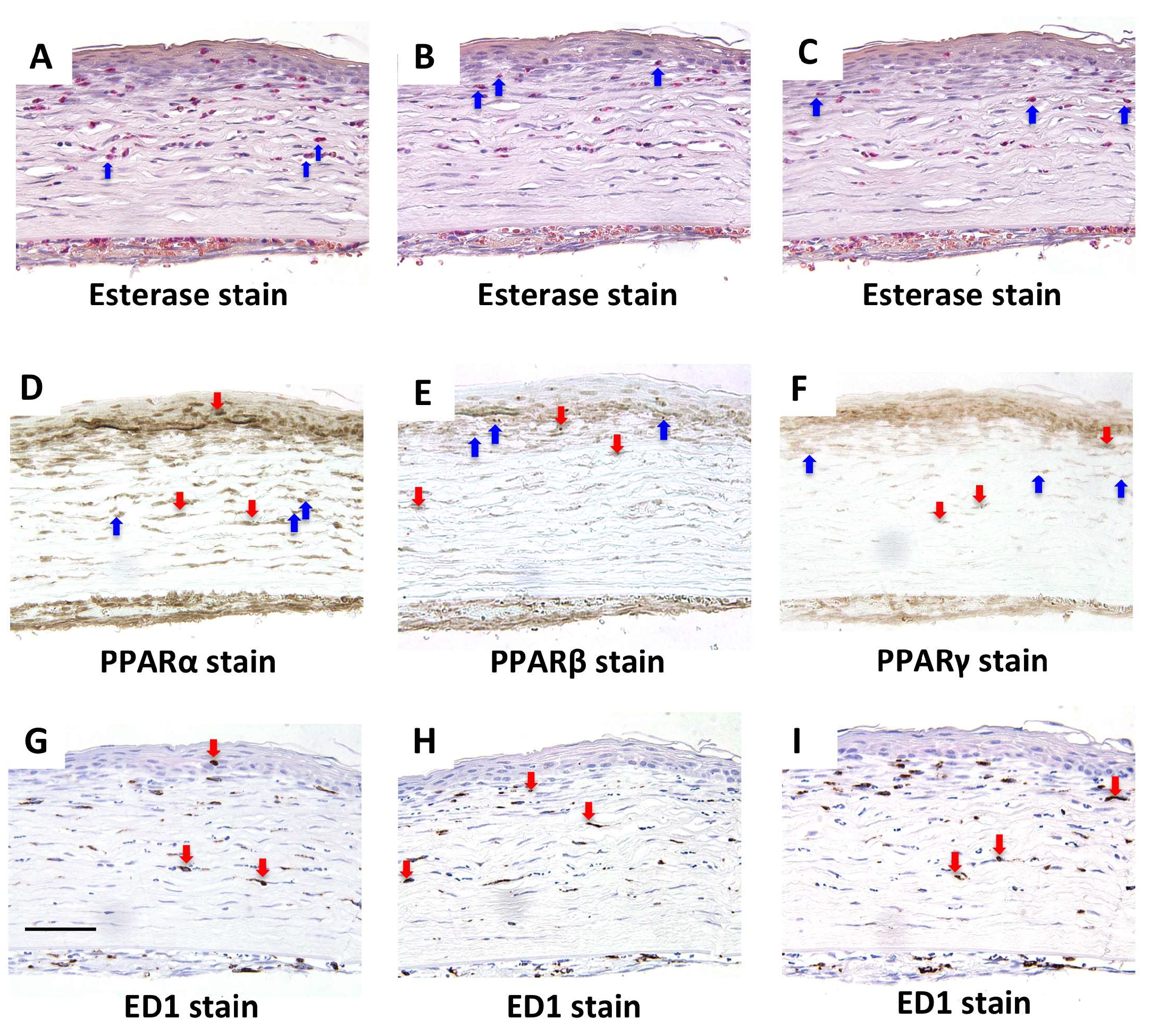
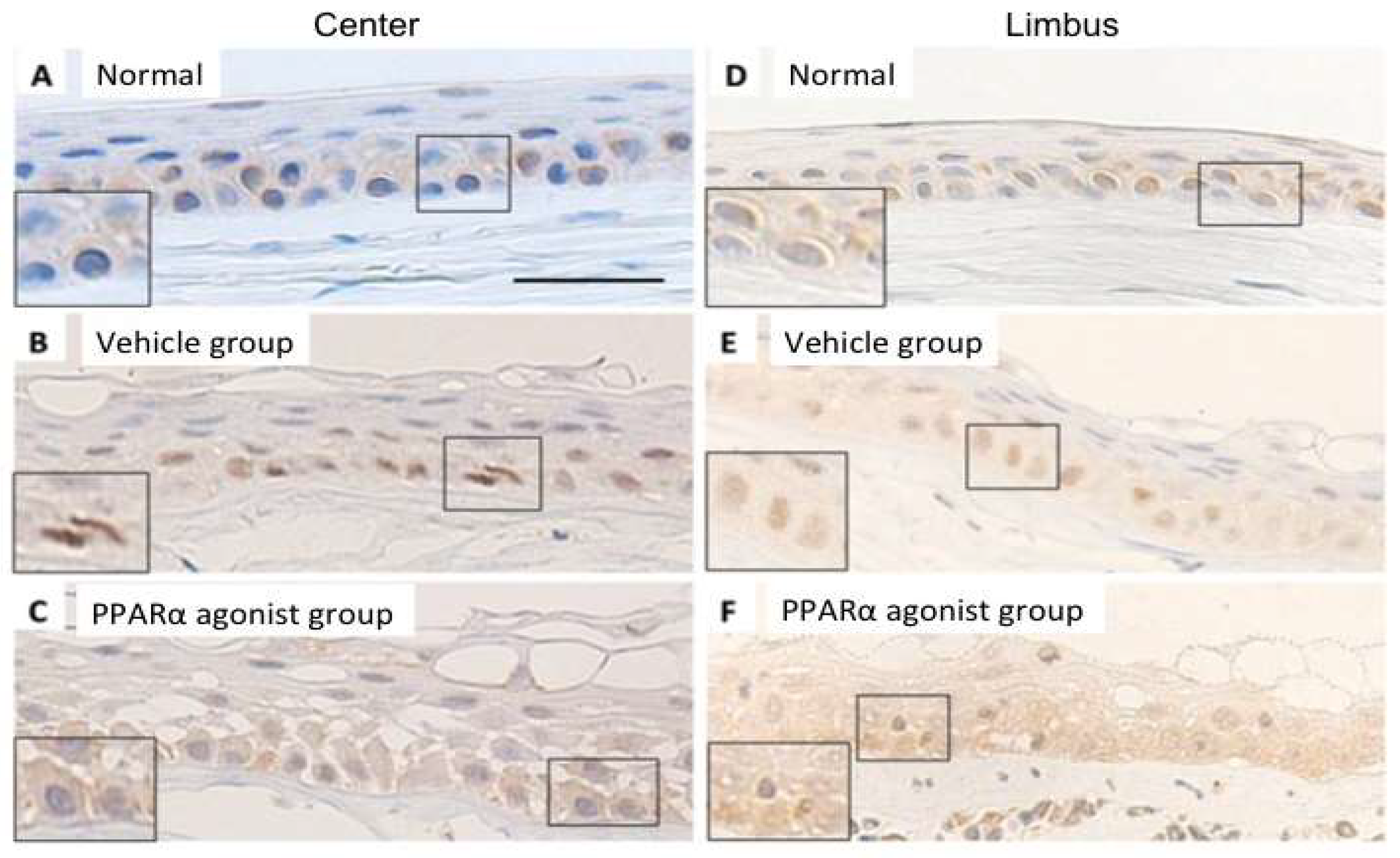
2.1. Localization of PPARs in the Rat Cornea
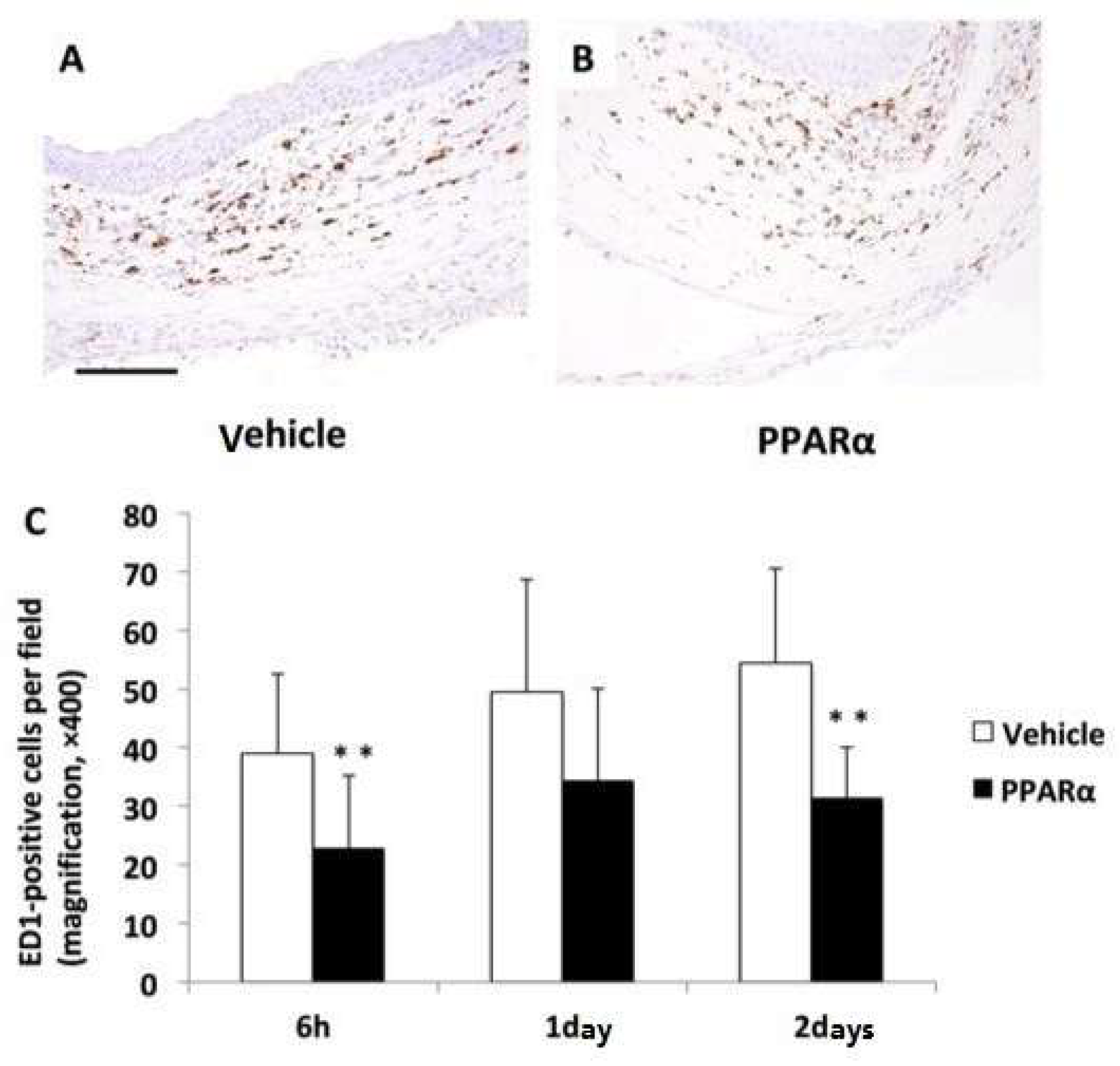
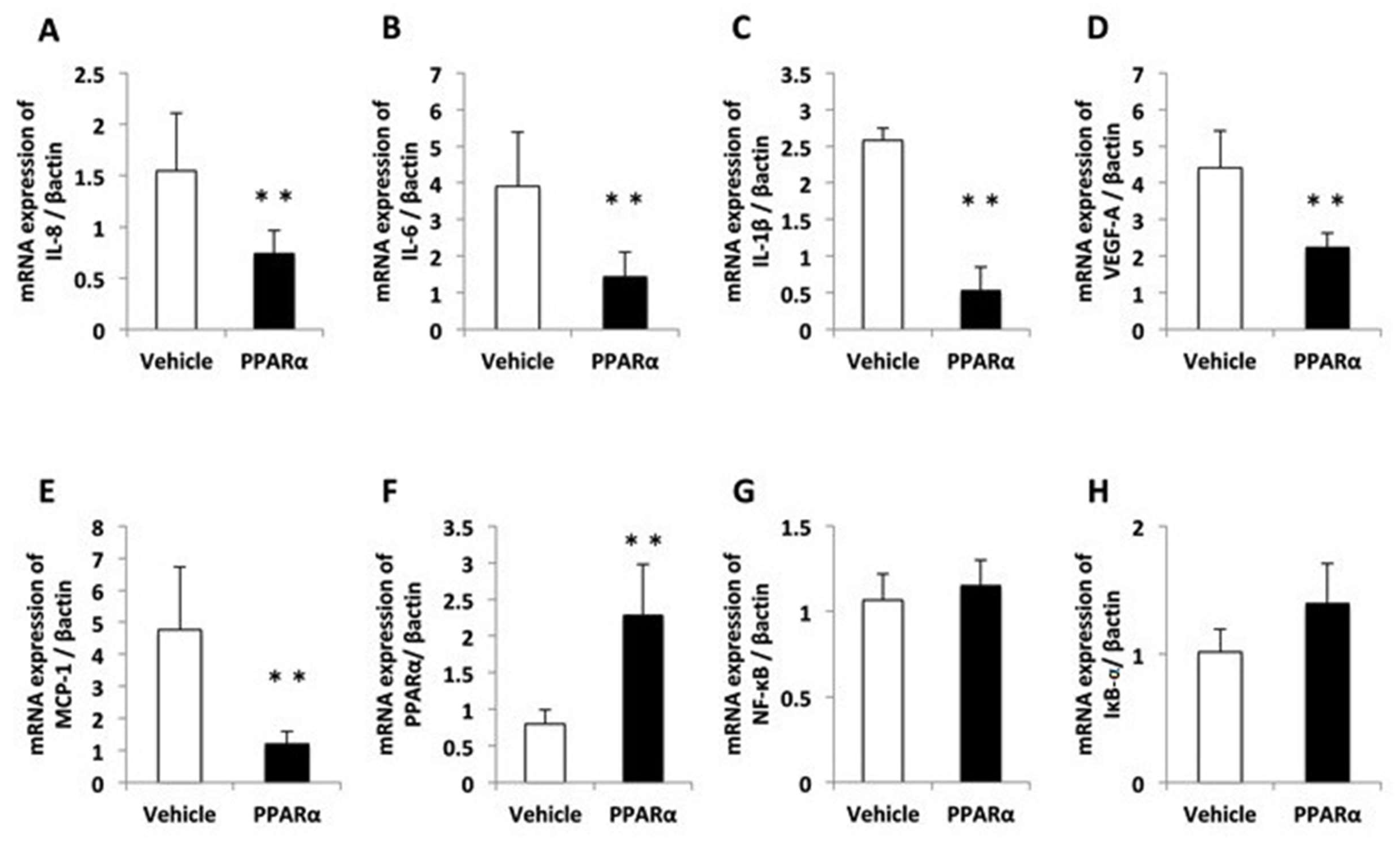
2.2. Effects of an Ophthalmic Solution of a PPARα Agonist after Alkali Burn Injury
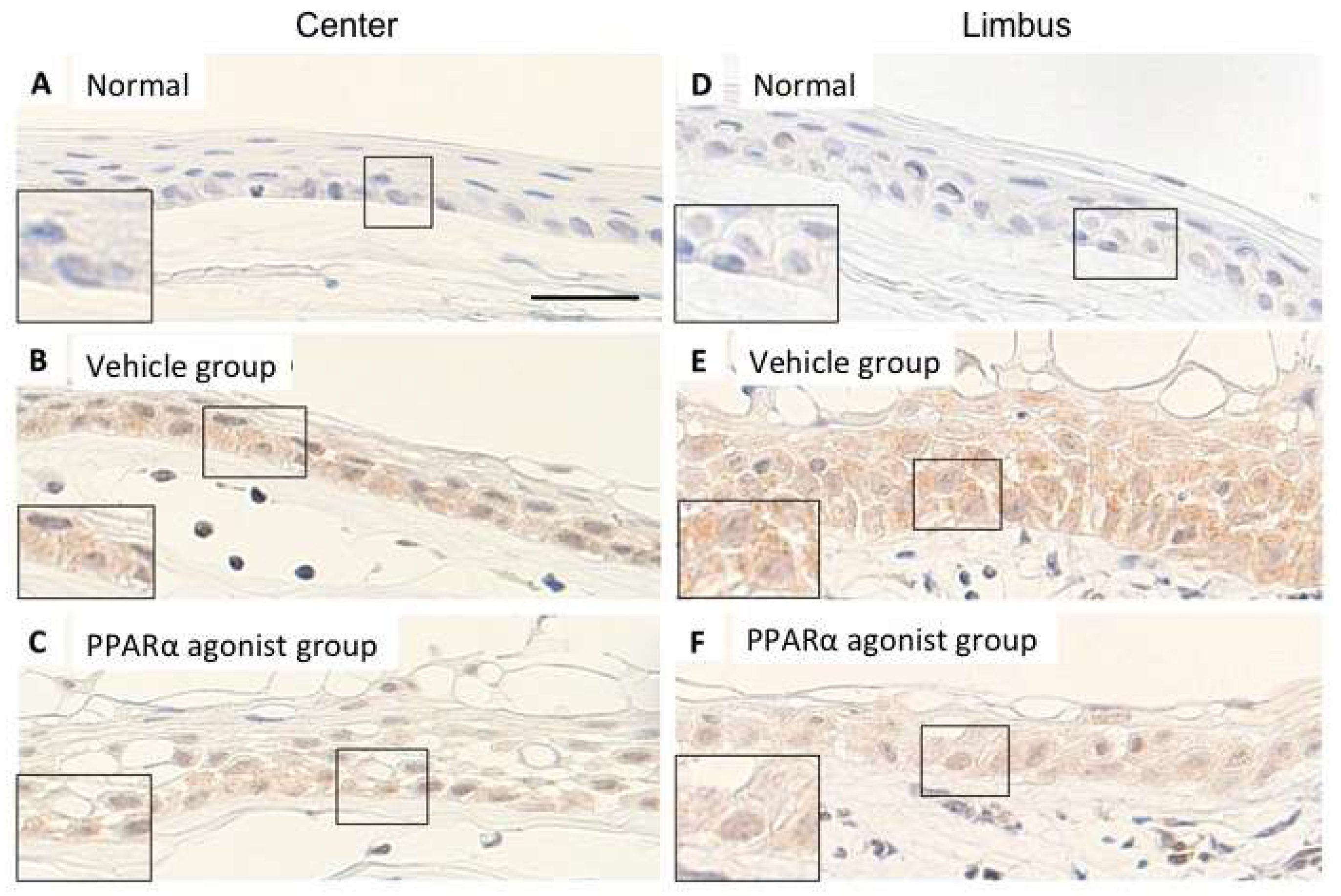
2.3. NF-κB, IκB-α, and MCP-1 Staining on Day 1
3. Discussion
4. Materials and Methods
4.1. Animal Model of Corneal Alkali Burn
4.2. Ophthalmic Solution of PPARα Agonist
4.3. Treatment with the Ophthalmic Solutions
4.4. Histological and Immunohistochemical Analyses
4.5. Real-Time RT–PCR
4.6. Statistical Analyses
5. Conclusions
Reference
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gottlicher, M.; Widmark, E.; Li, Q.; Gustafsson, J.A. Fatty acids activate a chimera of the clofibric acid-activated receptor and the glucocorticoid receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 4653–4657. [Google Scholar] [CrossRef] [PubMed]
- Issemann, I.; Green, S. Activation of a member of the steroid hormone receptor superfamily by peroxisome proliferators. Nature 1990, 347, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Hu, E.; Graves, R.A.; Budavari, A.I.; Spiegelman, B.M. mPPAR gamma 2: Tissue-specific regulator of an adipocyte enhancer. Genes Dev. 1994, 8, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Olson, P.; Evans, R.M. Minireview: Lipid metabolism, metabolic diseases, and peroxisome proliferator-activated receptors. Endocrinology 2003, 144, 2201–2207. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Nagothu, K.K.; Desai, V.; Lee, T.; Branham, W.; Moland, C.; Megyesi, J.K.; Crew, M.D.; Portilla, D. Transgenic expression of proximal tubule peroxisome proliferator-activated receptor-alpha in mice confers protection during acute kidney injury. Kidney Int. 2009, 76, 1049–1062. [Google Scholar] [CrossRef] [PubMed]
- Desvergne, B.; Wahli, W. Peroxisome proliferator-activated receptors: Nuclear control of metabolism. Endocr. Rev. 1999, 20, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, L.S.; Lione, L. Harnessing the benefits of PPARβ/δ agonists. Life Sci. 2013, 93, 963–967. [Google Scholar] [CrossRef]
- Zhang, S.; Gu, H.; Hu, N. Role of Peroxisome Proliferator-Activated Receptor γ in Ocular Diseases. J. Ophthalmol. 2015, 2015, 275435. [Google Scholar] [CrossRef]
- Balachandar, S.; Katyal, A. Peroxisome proliferator activating receptor (PPAR) in cerebral malaria (CM): A novel target for an additional therapy. Eur. J. Clin. Microbiol. 2011, 30, 483–498. [Google Scholar] [CrossRef]
- Nakamura, Y.; Nakamura, T.; Tarui, T.; Inoue, J.; Kinoshita, S. Functional role of PPARdelta in corneal epithelial wound healing. Am. J. Pathol. 2012, 180, 583–598. [Google Scholar] [CrossRef]
- Uchiyama, M.; Shimizu, A.; Masuda, Y.; Nagasaka, S.; Fukuda, Y.; Takahashi, H. An ophthalmic solution of a peroxisome proliferator-activated receptor gamma agonist prevents corneal inflammation in a rat alkali burn model. Mol. Vis. 2013, 19, 2135–2150. [Google Scholar] [PubMed]
- Arima, T.; Uchiyama, M.; Nakano, Y.; Nagasaka, S.; Kang, D.; Shimizu, A.; Takahashi, H. Peroxisome proliferator-activated receptor alpha agonist suppresses neovascularization by reducing both vascular endothelial growth factor and angiopoietin-2 in corneal alkali burn. Sci. Rep. 2017, 7, 17763. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Gao, Y.; Lu, B.; Zhang, Q.; Hu, Y.; Chen, Y. Negatively regulating TLR4/NF-κB signaling via PPARα in endotoxin-induced uveitis. BBA-Mol. Basis Dis. 2014, 1842, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Delerive, P.; Gervois, P.; Fruchart, J.C.; Staels, B. Induction of IκBα expression as a mechanism contributing to the anti-inflammatory activities of PPARα activators. J. Biol. Chem. 2000, 275, 36703–36707. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.C.; Ding, X.; Daynes, R.A. Nuclear receptor peroxisome proliferator-activated receptor α (PPARα) is expressed in resting murine lymphocytes. The PPARα in T and B lymphocytes is both transactivation and transrepression competent. J. Biol. Chem. 2002, 277, 6838–6845. [Google Scholar] [CrossRef] [PubMed]
- Cunard, R.; DiCampli, D.; Archer, D.C.; Stevenson, J.L.; Ricote, M.; Glass, C.K.; Kelly, C.J. WY14,643, a PPARα ligand, has profound effects on immune responses in vivo. J. Immunol. 2002, 169, 6806–6812. [Google Scholar] [CrossRef] [PubMed]
- Delerive, P.; De Bosscher, K.; Besnard, S.; Vanden Berghe, W.; Peters, J.M.; Gonzalez, F.J.; Fruchart, J.C.; Tedgui, A.; Haegeman, G.; Staels, B. Peroxisome proliferator-activated receptor α negatively regulates the vascular inflammatory gene response by negative cross-talk with transcription factors NF-κB and AP-1. J. Biol. Chem. 1999, 274, 32048–32054. [Google Scholar] [CrossRef] [PubMed]
- Chinetti, G.; Lestavel, S.; Bocher, V.; Remaley, A.T.; Neve, B.; Torra, I.P.; Teissier, E.; Minnich, A.; Jaye, M.; Duverger, N.; et al. PPAR-α and PPAR-γ activators induce cholesterol removal from human macrophage foam cells through stimulation of the ABCA1 pathway. Nat. Med. 2001, 7, 53–58. [Google Scholar] [CrossRef]
- Panigrahy, D.; Kaipainen, A.; Huang, S.; Butterfield, C.E.; Barnés, C.M.; Fannon, M.; Laforme, A.M.; Chaponis, D.M.; Folkman, J.; Kieran, M.W. PPARα agonist fenofibrate suppresses tumor growth through direct and indirect angiogenesis inhibition. Proc. Natl. Acad. Sci. USA 2008, 105, 985–990. [Google Scholar] [CrossRef]
- Srivastava, S.K.; Ramana, K.V. Focus on molecules: Nuclear factor-κB. Exp. Eye Res. 2009, 88, 2–3. [Google Scholar] [CrossRef]
- Lan, W.; Petznick, A.; Heryati, S.; Rifada, M.; Tong, L. Nuclear Factor-kappaB: Central regulator in ocular surface inflammation and diseases. Ocul. Surf. 2012, 10, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zeng, L.; Zhang, T.; Liu, J.; Wang, W. Tenuigenin Prevents IL-1β-induced Inflammation in Human Osteoarthritis Chondrocytes by Suppressing PI3K/AKT/NF-κB Signaling Pathway. Inflammation 2016, 39, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Verma, I.M. NF-κB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Silverman, N.; Maniatis, T. NF-κB signaling pathways in mammalian and insect innate immunity. Gene. Dev. 2001, 15, 2321–2342. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Mihara, Y.; Kanai, K.; Yamashita, Y.; Kimura, Y.; Itoh, N. Tyrosol ameliorates lipopolysaccharide-induced ocular inflammation in rats via inhibition of nuclear factor (NF)-κB activation. J. Vet. Med. Sci. 2016, 78, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, N.; Yamagami, S.; Yokoo, S.; Amano, S.; Murakami, A. Involvement of C-C chemokine ligand 2-CCR2 interaction in monocyte-lineage cell recruitment of normal human corneal stroma. J. Immunol. 2007, 178, 3288–3292. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.W.; Liu, J.J.; Lee, J.S.; Mohan, R.R.; Mohan, R.R.; Woods, D.J.; He, Y.G.; Wilson, S.E. Proinflammatory chemokine induction in keratocytes and inflammatory cell infiltration into the cornea. Investig. Ophth. Vis. Sci. 2001, 42, 2795–2803. [Google Scholar]
- Goede, V.; Brogelli, L.; Ziche, M.; Augustin, H.G. Induction of inflammatory angiogenesis by monocyte chemoattractant protein-1. Int. J. Cancer 1999, 82, 765–770. [Google Scholar] [CrossRef]
- O’Brien, T.P.; Li, Q.; Ashraf, M.F.; Matteson, D.M.; Stark, W.J.; Chan, C.C. Inflammatory response in the early stages of wound healing after excimer laser keratectomy. Arch. Ophthalmol.-Chic. 1998, 116, 1470–1474. [Google Scholar] [CrossRef]
- Liu, Q.; Smith, C.W.; Zhang, W.; Burns, A.R.; Li, Z. NK cells modulate the inflammatory response to corneal epithelial abrasion and thereby support wound healing. Am. J. Pathol. 2012, 181, 452–462. [Google Scholar] [CrossRef]
Sample Availability: There was no sample of the compounds by the authors in this experiment. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| IL-8 (CXCL8) | CCCCCATGGTTCAGAAGATTG | TTGTCAGAAGCCAGCGTTCAC |
| IL-6 | GTCAACTCCATCTGCCCTTCAG | GGCAGTGGCTGTCAACAACAT |
| IL-1β | TACCTATGTCTTGCCCGTGGAG | ATCATCCCACGAGTCACAGAGG |
| VEGF-A | TGTGCGGGCTGCTGCAATGAT | TGTGCTGGCTTTGGTGAGGTTTGA |
| MCP-1 (CCL2) | AGCCAGATGCAGTTAATGCCC | ACACCTGCTGCTGGTGATTCTC |
| PPARα | TGAACAAAGACGGGATG | TCAAACTTGGGTTCCATGAT |
| NF-κB | GGCAGCACTCCTTATCAA | GGTGTCGTCCCATCGTAG |
| IκB-α | TGACCATGGAAGTGATTGGTCAG | GATCACAGCCAAGTGGAGTGGA |
| β-actin | ACCACCATGTACCCAGGCATT | CCACACAGAGTACTTGCGCTCA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakano, Y.; Uchiyama, M.; Arima, T.; Nagasaka, S.; Igarashi, T.; Shimizu, A.; Takahashi, H. PPARα Agonist Suppresses Inflammation after Corneal Alkali Burn by Suppressing Proinflammatory Cytokines, MCP-1, and Nuclear Translocation of NF-κB. Molecules 2019, 24, 114. https://doi.org/10.3390/molecules24010114
Nakano Y, Uchiyama M, Arima T, Nagasaka S, Igarashi T, Shimizu A, Takahashi H. PPARα Agonist Suppresses Inflammation after Corneal Alkali Burn by Suppressing Proinflammatory Cytokines, MCP-1, and Nuclear Translocation of NF-κB. Molecules. 2019; 24(1):114. https://doi.org/10.3390/molecules24010114
Chicago/Turabian StyleNakano, Yuichiro, Masaaki Uchiyama, Takeshi Arima, Shinya Nagasaka, Tsutomu Igarashi, Akira Shimizu, and Hiroshi Takahashi. 2019. "PPARα Agonist Suppresses Inflammation after Corneal Alkali Burn by Suppressing Proinflammatory Cytokines, MCP-1, and Nuclear Translocation of NF-κB" Molecules 24, no. 1: 114. https://doi.org/10.3390/molecules24010114
APA StyleNakano, Y., Uchiyama, M., Arima, T., Nagasaka, S., Igarashi, T., Shimizu, A., & Takahashi, H. (2019). PPARα Agonist Suppresses Inflammation after Corneal Alkali Burn by Suppressing Proinflammatory Cytokines, MCP-1, and Nuclear Translocation of NF-κB. Molecules, 24(1), 114. https://doi.org/10.3390/molecules24010114