The Liver Protection Effects of Maltol, a Flavoring Agent, on Carbon Tetrachloride-Induced Acute Liver Injury in Mice via Inhibiting Apoptosis and Inflammatory Response

 ,
, 
Abstract

1. Introduction
2. Results
2.1. Effect of Maltol on Body Weight and Organ Index in Mice
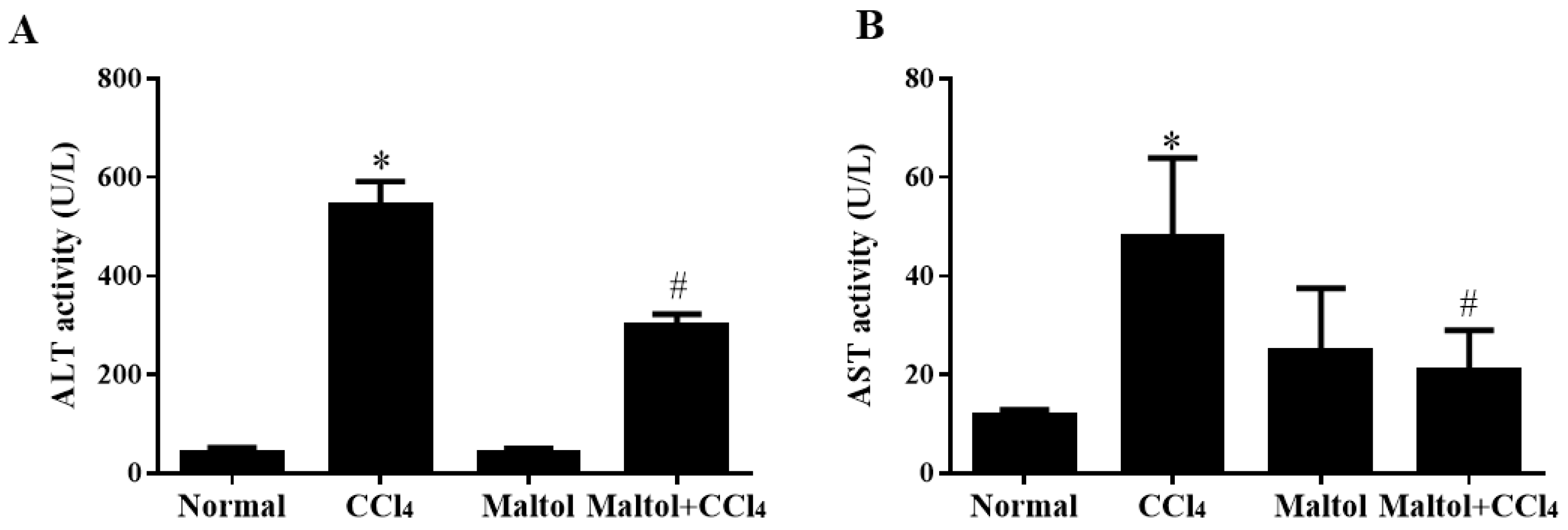
2.2. Effect of Maltol on Serum Aspartate Transaminase (Alt) and Alanine Transaminase (Ast) Activities
2.3. Maltol Ameliorated CCl4-Induced Hepatic Histopathological Changes in Mice
2.4. Maltol Inhibited Cell Apoptosis in CCl4-Induced Acute Liver Injury in Mice
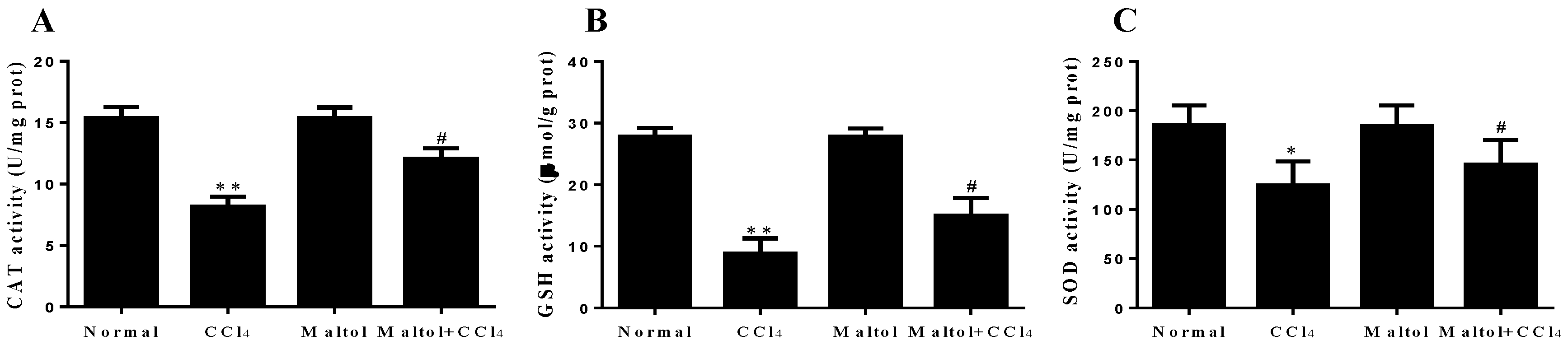
2.5. Effects of Maltol on Oxidative Stress Markers
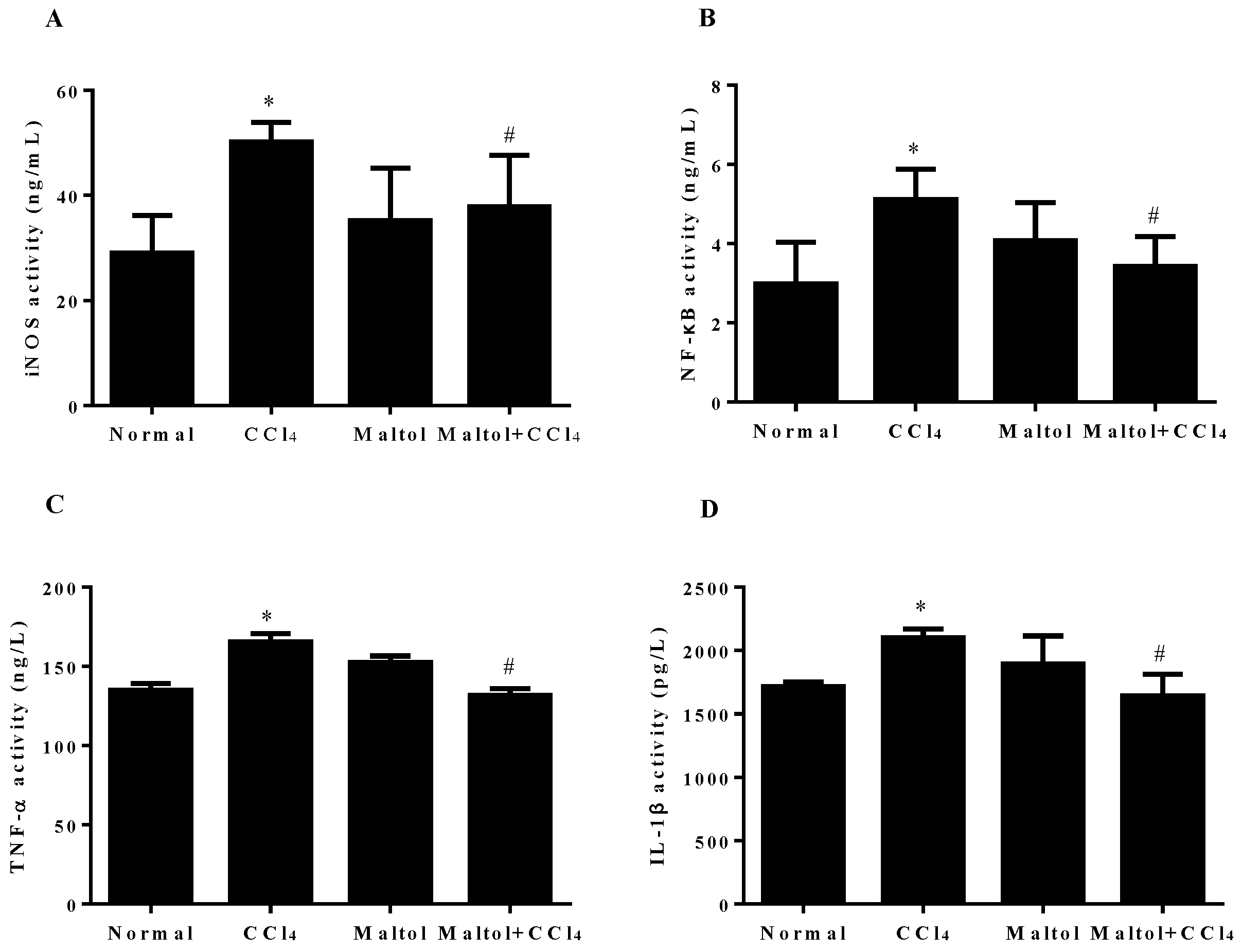
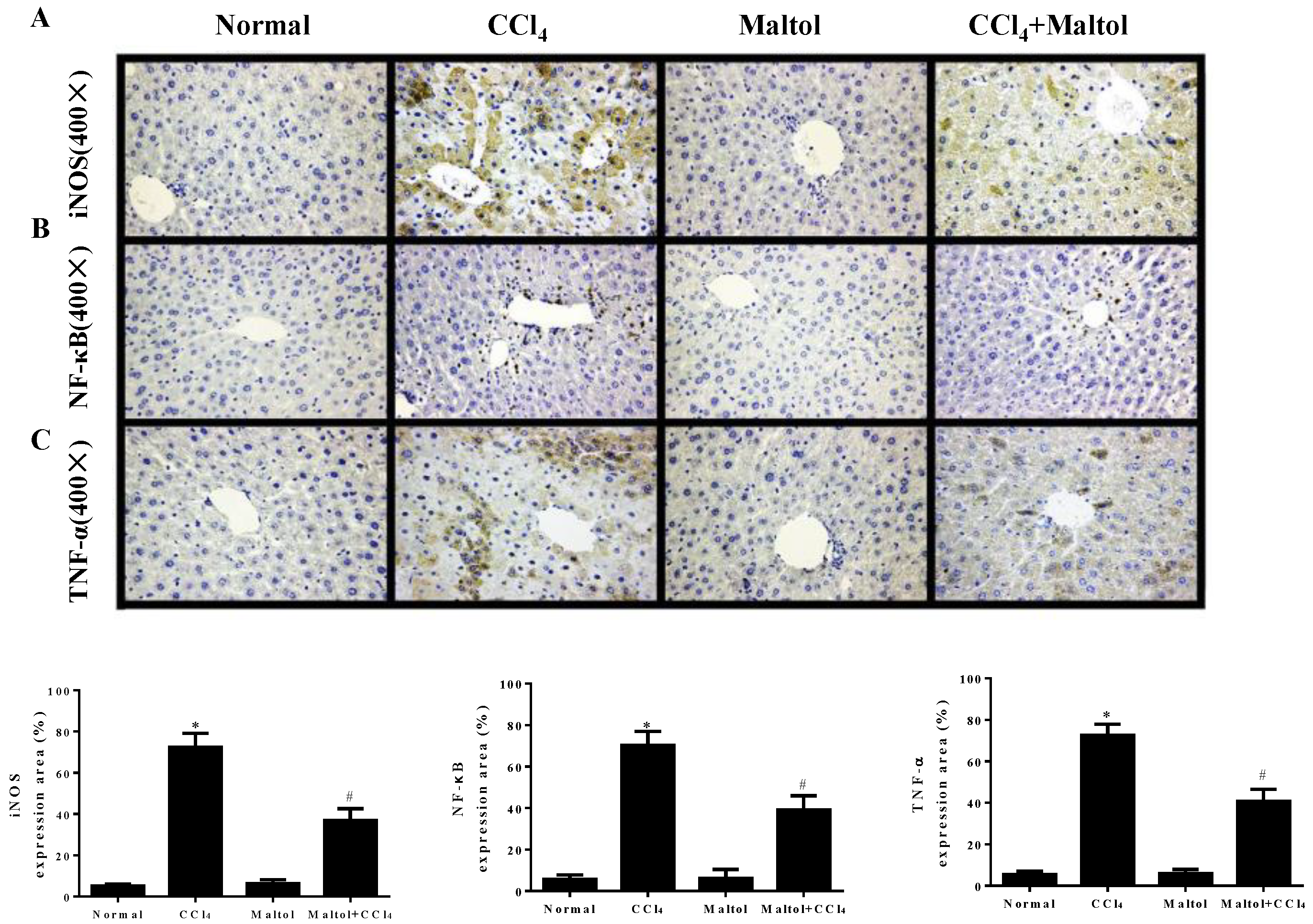
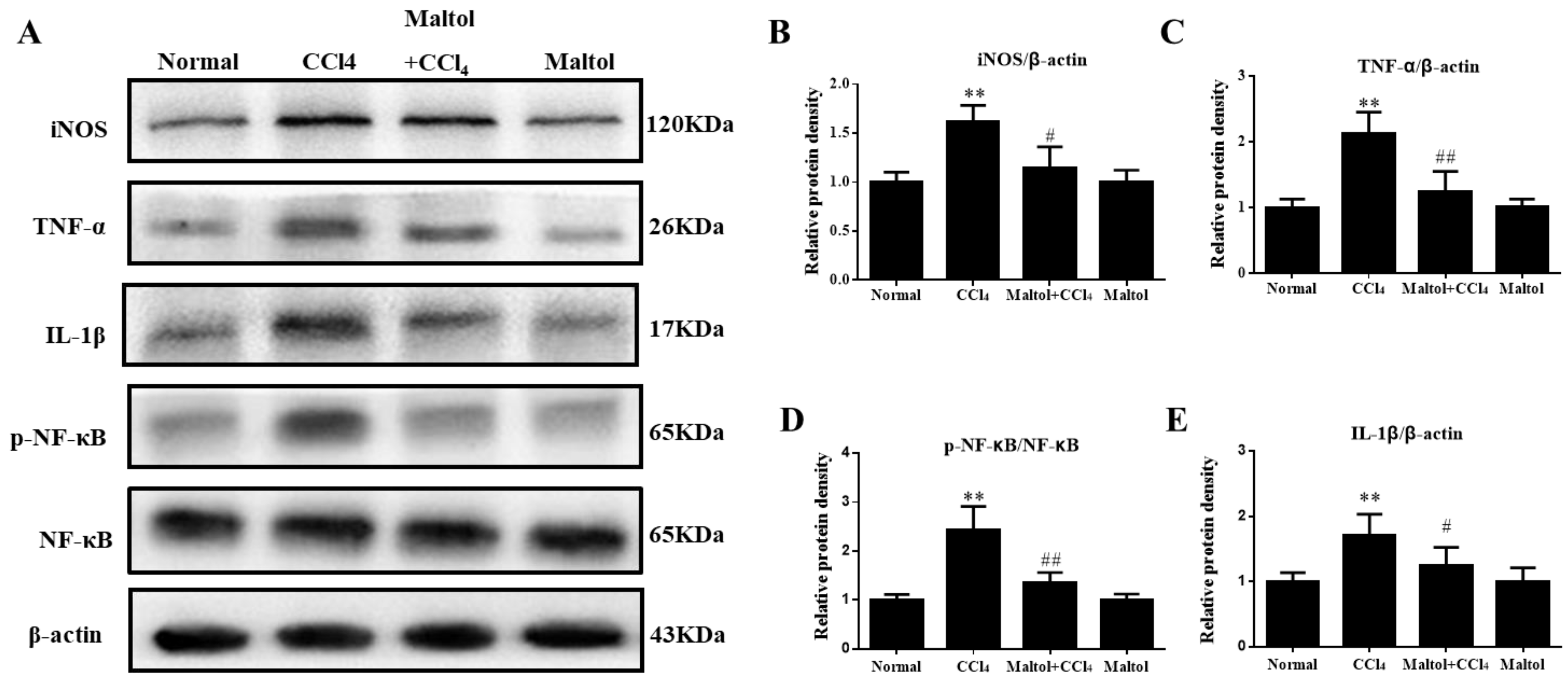
2.6. Effect of Maltol on CCl4-Induced Inflammatory Response
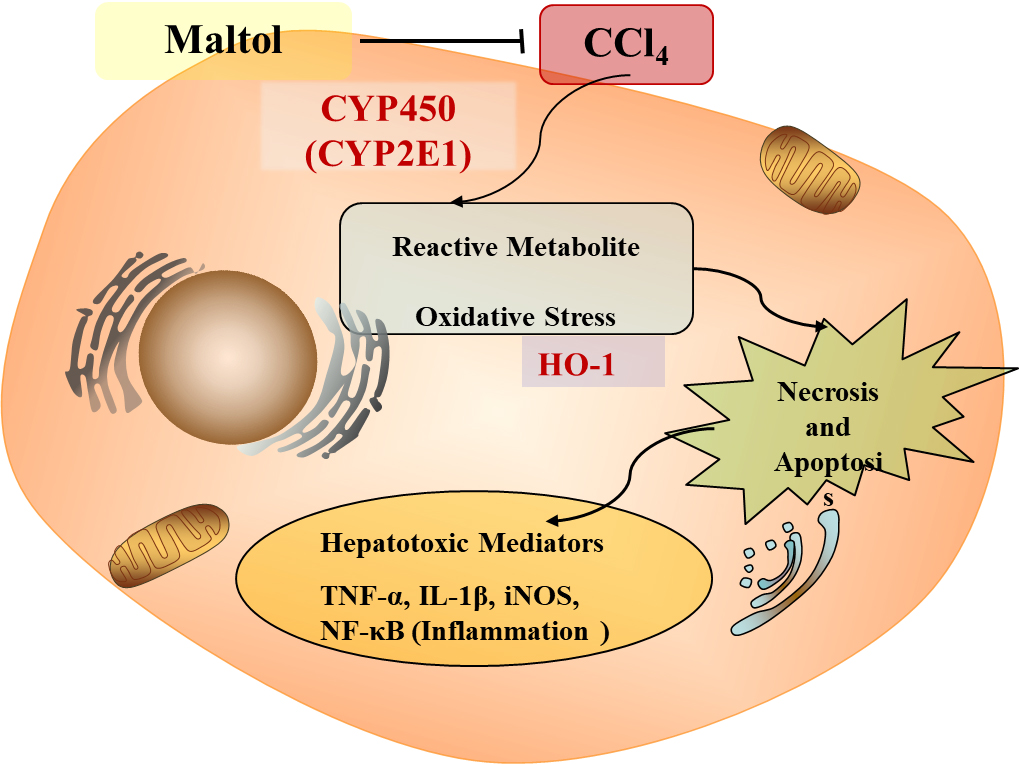
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. Animal Treatment and Experimental Protocol
4.4. Determination of Liver Enzymes
4.5. Assay for Inflammatory Markers
4.6. Histopathological Analysis
4.7. Hoechst 33258 Staining
4.8. TUNEL Assay
4.9. Immunohistochemistry Analysis
4.10. Immunofluorescence Analysis
4.11. Western Blot Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ALT | Alanine aminotransferase |
| iNOS | Inducible nitric oxide synthase |
| TNF-α | Tumor necrosis factor-α |
| NO | Nitric oxide |
| HO-1 | Heme oxygenase-1 |
| CAT | Catalase |
| SOD | Superoxide dismutase |
| AST | Aspartate aminotransferase |
| NF-κB | Nuclear factor-kappa B |
| IL-1β | Interleukin-1β |
| CYP2E1 | Cytochrome P450 E1 |
| CCl4 | Carbon tetrachloride |
| GSH | Glutathione |
References
- Zamora Nava, L.E.; Aguirre Valadez, J.; Chavez-Tapia, N.C.; Torre, A. Acute-on-chronic liver failure: A Review. Ther. Clin. Risk Manag. 2014, 10, 295–303. [Google Scholar] [PubMed]
- Chiu, Y.J.; Chou, S.C.; Chiu, C.S.; Kao, C.P.; Wu, K.C.; Chen, C.J.; Tsai, J.C.; Peng, W.H. Hepatoprotective effect of the ethanol extract of polygonum orientale on carbon tetrachloride-induced acute liver injury on mice. J. Food Drug Anal. 2018, 26, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Muriel, P.; Rivera-Espinoza, Y. Beneficial drugs for liver diseases. J. Appl. Toxicol. 2008, 28, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Amat, N.; Upur, H.; Blazekovic, B. In vivo hepatoprotective activity of the aqueous extract of Artemisia absinthium L. Against chemically and immunologically induced liver injuries in mice. J. Ethnopharmacol. 2010, 131, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Wang, F.; Tang, J.; Mao, A.; Liao, S.; Wang, Q. Dicranostiga leptopodu (maxim.) fedde extracts attenuated CCl4-induced acute liver damage in mice through increasing anti-oxidative enzyme activity to improve mitochondrial function. Biomed. Pharmacother. 2017, 85, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Recknagel, R.O.; Glende, E.A., Jr.; Dolak, J.A.; Waller, R.L. Mechanisms of carbon tetrachloride toxicity. Pharmacol. Ther. 1989, 43, 139–154. [Google Scholar] [CrossRef]
- Li, S.; Hong, M.; Tan, H.Y.; Wang, N.; Feng, Y. Insights into the role and interdependence of oxidative stress and inflammation in liver diseases. Oxid. Med. Cell. Longev. 2016, 2016, 4234061. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.; Kodani, S.; Rand, A.; Yang, J.; Imai, D.; Hwang, S.; Hammock, B. Celecoxib does not protect against fibrosis and inflammation in a carbon tetrachloride-induced model of liver injury. Mol. Pharmacol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Hou, J.; Liu, W.; Ren, S.; Wang, Y.; Zhang, R.; Chen, C.; Wang, Z.; Li, W. 20(r)-ginsenoside rg3, a rare saponin from red ginseng, ameliorates acetaminophen-induced hepatotoxicity by suppressing pi3k/akt pathway-mediated inflammation and apoptosis. Int. Immunopharmacol. 2018, 59, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Acaroz, U.; Ince, S.; Arslan-Acaroz, D.; Gurler, Z.; Kucukkurt, I.; Demirel, H.; Arslan, H.; Varol, N.; Zhu, K. The ameliorative effects of boron against acrylamide-induced oxidative stress, inflammatory response, and metabolic changes in rats. Food Chem. Toxicol. 2018, 118, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Altunay, N.; Gurkan, R.; Orhan, U. Indirect determination of the flavor enhancer maltol in foods and beverages through flame atomic absorption spectrometry after ultrasound assisted-cloud point extraction. Food Chem. 2017, 235, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Hong, S.; Iizuka, Y.; Kim, C.Y.; Seong, G.J. The neuroprotective effect of maltol against oxidative stress on rat retinal neuronal cells. Korean J. Ophthalmol. 2015, 29, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Corrochano, A.R.; Buckin, V.; Kelly, P.M.; Giblin, L. Invited review: Whey proteins as antioxidants and promoters of cellular antioxidant pathways. J. Dairy Sci. 2018, 101, 4747–4761. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, J.; Xu, C.; Pan, H.; Zhang, Z. Maltol inhibits apoptosis of human neuroblastoma cells induced by hydrogen peroxide. J. Biochem. Mol. Biol. 2006, 39, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Iizuka, Y.; Lee, T.; Kim, C.; Seong, G. Neuroprotective and neurite outgrowth effects of maltol on retinal ganglion cells under oxidative stress. Mol. Vis. 2014, 20, 1456–1462. [Google Scholar] [PubMed]
- Liu, Y.; Wen, P.; Zhang, X.; Dai, Y.; He, Q. Breviscapine ameliorates ccl4-induced liver injury in mice through inhibiting inflammatory apoptotic response and ros generation. Int. J. Mol. Med. 2018, 42, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Yamabe, N.; Kim, H.Y.; Yokozawa, T. Role of maltol in advanced glycation end products and free radicals: In-vitro and in-vivo studies. J. Pharm. Pharmacol. 2008, 60, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Stallmach, A.; Buning, C. Ferric maltol (st10): A novel oral iron supplement for the treatment of iron deficiency anemia in inflammatory bowel disease. Expert Opin. Pharmacother. 2015, 16, 2859–2867. [Google Scholar] [CrossRef] [PubMed]
- Enyedy, E.A.; Domotor, O.; Bali, K.; Hetenyi, A.; Tuccinardi, T.; Keppler, B.K. Interaction of the anticancer gallium(iii) complexes of 8-hydroxyquinoline and maltol with human serum proteins. JBJC J. Biol. Inorg. Chem. 2015, 20, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Domotor, O.; Aicher, S.; Schmidlehner, M.; Novak, M.S.; Roller, A.; Jakupec, M.A.; Kandioller, W.; Hartinger, C.G.; Keppler, B.K.; Enyedy, E.A. Antitumor pentamethylcyclopentadienyl rhodium complexes of maltol and allomaltol: Synthesis, solution speciation and bioactivity. J. Inorg. Biochem. 2014, 134, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Kandioller, W.; Hartinger, C.G.; Nazarov, A.A.; Bartel, C.; Skocic, M.; Jakupec, M.A.; Arion, V.B.; Keppler, B.K. Maltol-derived ruthenium-cymene complexes with tumor inhibiting properties: The impact of ligand-metal bond stability on anticancer activity in vitro. Chemistry 2009, 15, 12283–12291. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Lu, Y.; Hu, Y.; Zhai, X.; Xu, W.; Jing, H.; Tian, X.; Lin, Y.; Gao, D.; Yao, J. Salvianolic acid b protects against acute ethanol-induced liver injury through sirt1-mediated deacetylation of p53 in rats. Toxicol. Lett. 2014, 228, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Qi, X.; Cao, S.; Li, P. Protective effect of flavonoid extract from chinese bayberry (myrica rubra sieb. Et zucc.) fruit on alcoholic liver oxidative injury in mice. J. Nat. Med. 2014, 68, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Abdelmegeed, M.A.; Song, B.J. Preventive effects of indole-3-carbinol against alcohol-induced liver injury in mice via antioxidant, anti-inflammatory, and anti-apoptotic mechanisms: Role of gut-liver-adipose tissue axis. J. Nutr. Biochem. 2018, 55, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.C.; Chiu, C.S.; Chen, Y.C.; Lee, M.S.; Hao, X.Y.; Hsieh, M.T.; Kao, C.P.; Peng, W.H. Hepatoprotective effect of coreopsis tinctoria flowers against carbon tetrachloride-induced liver damage in mice. BMC Complement. Altern. Med. 2017, 17, 139. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.Y.; Li, G.Y.; Lou, M.M.; Li, X.Y.; Wei, X.Y.; Wang, J.H. Hepatoprotective effect of matrine salvianolic acid b salt on carbon tetrachloride-induced hepatic fibrosis. J. Inflamm. 2012. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.S.; Wang, Y.H.; Lai, Y.Y.; Tsou, H.K.; Liou, G.G.; Ko, J.L.; Wang, S.H. Kaempferol protects against propacetamol-induced acute liver injury through cyp2e1 inactivation, ugt1a1 activation, and attenuation of oxidative stress, inflammation and apoptosis in mice. Toxicol. Lett. 2018, 290, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Radulovic, N.S.; Randjelovic, P.J.; Stojanovic, N.M.; Ilic, I.R.; Miltojevic, A.B.; Stojkovic, M.B.; Ilic, M. Effect of two esters of n-methylanthranilic acid from rutaceae species on impaired kidney morphology and function in rats caused by CCl4. Life Sci. 2015, 135, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Chakraborty, A.; Dey, P.; Kar, P.; Guha, P.; Sen, S.; Kumar, A.; Sen, A.; Chaudhuri, T. Amelioration of ccl4 induced liver injury in swiss albino mice by antioxidant rich leaf extract of croton bonplandianus baill. PLoS ONE 2018. [Google Scholar] [CrossRef] [PubMed]
- Basu, S. Carbon tetrachloride-induced lipid peroxidation: Eicosanoid formation and their regulation by antioxidant nutrients. Toxicology 2003, 189, 113–127. [Google Scholar] [CrossRef]
- Ai, G.; Liu, Q.; Hua, W.; Huang, Z.; Wang, D. Hepatoprotective evaluation of the total flavonoids extracted from flowers of Abelmoschus manihot (L.) medic: In vitro and in vivo studies. J. Ethnopharmacol. 2013, 146, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Dong, Z.; Chang, X.; Zhang, C.; Rong, G.; Gao, X.; Zeng, Z.; Wang, C.; Chen, Y.; Rong, Y.; et al. Protective effect of the total flavonoids from apocynum venetum l. On carbon tetrachloride-induced hepatotoxicity in vitro and in vivo. J. Physiol. Biochem. 2018, 74, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Xu, Q.; Hu, J.N.; Han, X.Y.; Li, W.; Zhao, L.C. Maltol, a food flavoring agent, attenuates acute alcohol-induced oxidative damage in mice. Nutrients 2015, 7, 682–696. [Google Scholar] [CrossRef] [PubMed]
- Mendes-Braz, M.; Martins, J.O. Diabetes mellitus and liver surgery: The effect of diabetes on oxidative stress and inflammation. Mediat. Inflamm. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Tao, X.; Liang, S.; Pan, Y.; He, L.; Sun, J.; Wenbo, J.; Li, X.; Chen, J.; Wang, C. Protective effect of acidic polysaccharide from schisandra chinensis on acute ethanol-induced liver injury through reducing cyp2e1-dependent oxidative stress. Biomed. Pharmacother. 2018, 99, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Utaipan, T.; Suksamrarn, A.; Kaemchantuek, P.; Chokchaisiri, R.; Stremmel, W.; Chamulitrat, W.; Chunglok, W. Diterpenoid trigonoreidon b isolated from trigonostemon reidioides alleviates inflammation in models of lps-stimulated murine macrophages and inflammatory liver injury in mice. Biomed. Pharmacother. 2018, 101, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Germoush, M.O.; Othman, S.I.; Al-Qaraawi, M.A.; Al-Harbi, H.M.; Hussein, O.E.; Al-Basher, G.; Alotaibi, M.F.; Elgebaly, H.A.; Sandhu, M.A.; Allam, A.A.; et al. Umbelliferone prevents oxidative stress, inflammation and hematological alterations, and modulates glutamate-nitric oxide-cgmp signaling in hyperammonemic rats. Biomed. Pharmacother. 2018, 102, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, L.; Li, Q.; Zhang, Z.; Xu, L.; Lin, C.; Gao, L.; Zhao, K.; Liang, F.; Zhang, Q.; et al. Madecassoside prevents acute liver failure in lps/d-galn-induced mice by inhibiting p38/nf-kappab and activating nrf2/ho-1 signaling. Biomed. Pharmacother. 2018, 103, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Sun, X.; Xia, T.; Mao, Q.; Zhong, L. Pretreatment with dihydroquercetin, a dietary flavonoid, protected against concanavalin a-induced immunological hepatic injury in mice and tnf-alpha/actd-induced apoptosis in hepg2 cells. Food Funct. 2018, 9, 2341–2352. [Google Scholar] [CrossRef] [PubMed]
- Simeonova, P.P.; Gallucci, R.M.; Hulderman, T.; Wilson, R.; Kommineni, C.; Rao, M.; Luster, M.I. The role of tumor necrosis factor-alpha in liver toxicity, inflammation, and fibrosis induced by carbon tetrachloride. Toxicol. Appl. Pharmacol. 2001, 177, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Al-Tamimi, J.; Alhazza, I.M.; Al-Khalifa, M.; Metwalli, A.; Rady, A.; Ebaid, H. Potential effects of samsum ant, brachyponera sennaarensis, venom on tnf-alpha/nf-kappab mediated inflammation in ccl4-toxicity in vivo. Lipids Health Dis. 2016, 15, 198. [Google Scholar] [CrossRef] [PubMed]
- Hatano, E. Tumor necrosis factor signaling in hepatocyte apoptosis. J. Gastroenterol. Hepatol. 2007, 23, 501–502. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, W.; Jantan, I.; Kumolosasi, E.; Haque, M.A.; Bukhari, S.N.A. Immunomodulatory effects of tinospora crispa extract and its major compounds on the immune functions of raw 264.7 macrophages. Int. Immunopharmacol. 2018, 60, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.A.; Han, A.R.; Choi, J.; Ahn, J.Y.; Choi, S.Y.; Cho, S.W. Anti-inflammatory mechanisms of n-adamantyl-4-methylthiazol-2-amine in lipopolysaccharide-stimulated bv-2 microglial cells. Int. Immunopharmacol. 2014, 22, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Moita, E.; Gil-Izquierdo, A.; Sousa, C.; Ferreres, F.; Silva, L.R.; Valentao, P.; Dominguez-Perles, R.; Baenas, N.; Andrade, P.B. Integrated analysis of cox-2 and inos derived inflammatory mediators in lps-stimulated raw macrophages pre-exposed to echium plantagineum l. Bee pollen extract. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.T.; Tseng, Y.T.; Hsu, Y.Y.; Cheng, K.I.; Chou, S.H.; Lo, Y.C. Propofol attenuates lipopolysaccharide-induced reactive oxygen species production through activation of nrf2/gsh and suppression of nadph oxidase in human alveolar epithelial cells. Inflammation 2015, 38, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, B.; Li, J.; Feng, M.; Zhang, Y.; Yao, W.; Zhang, C.; Wan, L. Celastrol attenuates incision-induced inflammation and pain associated with inhibition of the nf-kappab signalling pathway via sarm. Life Sci. 2018, 15, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zeng, Y.; Wu, A.; Yu, C.; Tang, Y.; Wang, X.; Xiong, R.; Chen, H.; Wu, J.; Qin, D. Lychee seed fraction inhibits aβ(1-42)-induced neuroinflammation in bv-2 cells via nf-κb signaling pathway. Front. Pharmacol. 2018, 9, 380. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Q.; Liu, C.M.; Qin, Z.H.; Jiang, J.H.; Sun, Y.Z. Ganoderma applanatum terpenes protect mouse liver against benzo(alpha)pyren-induced oxidative stress and inflammation. Environ. Toxicol. Pharmacol. 2011, 31, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Pervez, M.A.; Khan, D.A.; Ijaz, A.; Khan, S. Effects of delta-tocotrienol supplementation on liver enzymes, inflammation, oxidative stress and hepatic steatosis in patients with nonalcoholic fatty liver disease. Turk. J. Gastroenterol. 2018, 29, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Tornatore, L.; Thotakura, A.K.; Bennett, J.; Moretti, M.; Franzoso, G. The nuclear factor kappa b signaling pathway: Integrating metabolism with inflammation. Trends Cell Biol. 2012, 22, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Hua, F.; Ma, J.; Ha, T.; Xia, Y.; Kelley, J.; Williams, D.; Kao, R.; Browder, I.; Schweitzer, J.; Kalbfleisch, J.; et al. Activation of toll-like receptor 4 signaling contributes to hippocampal neuronal death following global cerebral ischemia/reperfusion. J. Neuroimmunol. 2007, 190, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Leng, J.; Wang, Z.; Fu, C.; Zhang, J.; Ren, S.; Hu, J.; Jiang, S.; Wang, Y.; Chen, C.; Li, W. NF-κB and AMPK/PI3K/Akt signaling pathways are involved in the protective effects of platycodon grandiflorum saponins against acetaminophen-induced acute hepatotoxicity in mice. Phytother. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liu, Z.; Wang, Z.; Ren, S.; Tang, S.; Wang, Y.; Xiao, S.; Chen, C.; Li, W. Supplementation of American ginseng berry extract mitigated cisplatin-evoked nephrotoxicity by suppressing ros-mediated activation of MAPK and NF-κB signaling pathways. Food Chem. Toxicol. 2017, 110, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, L.; Han, Y.; Wu, Z.; Chen, P.; Zhang, H.; Zhou, J. Protective effects of hepatocyte-specific glycyrrhetic derivatives against carbon tetrachloride-induced liver damage in mice. Bioorg. Chem. 2017, 72, 42–50. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Dosage (mg/kg) | Weight (g) | Organ Indices (mg/g × 100) | ||
|---|---|---|---|---|---|
| Initial | Final | Liver | Kidney | ||
| Normal | — | 30.7 ± 1.06 | 30.34 ± 1.02 | 1.41 ± 0.06 | 0.45 ± 0.02 |
| CCl4 | — | 30.4 ± 1.14 | 28.78 ± 1.06 | 1.55 ± 0.20 * | 0.50 ± 0.04 * |
| Maltol | 100 | 30.3 ± 1.20 | 29.96 ± 0.89 | 1.45 ± 0.05 | 0.43 ± 0.06 # |
| CCl4 + Maltol | 100 | 30.1 ± 1.13 | 30.25 ± 1.36 | 1.43 ± 0.44 # | 0.45 ± 0.03 # |
| Groups | Dosage (mg/kg) | n | Necrocytosis Grade | Score | Ridit Analysis | ||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |||||
| Normal | — | 8 | 8 | 0 | 0 | 0 | 0 | 0 | 0.28 |
| CCl4 | — | 8 | 0 | 2 | 3 | 3 | 0 | 17 | 0.84 * |
| Maltol | 100 | 8 | 7 | 1 | 0 | 0 | 0 | 1 | 0.33 |
| CCl4 + Maltol | 100 | 8 | 3 | 4 | 1 | 0 | 0 | 6 | 0.55 # |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Wang, Z.; Hou, J.-g.; Zhou, Y.-d.; He, Y.-f.; Jiang, S.; Wang, Y.-p.; Ren, S.; Li, W. The Liver Protection Effects of Maltol, a Flavoring Agent, on Carbon Tetrachloride-Induced Acute Liver Injury in Mice via Inhibiting Apoptosis and Inflammatory Response. Molecules 2018, 23, 2120. https://doi.org/10.3390/molecules23092120
Liu W, Wang Z, Hou J-g, Zhou Y-d, He Y-f, Jiang S, Wang Y-p, Ren S, Li W. The Liver Protection Effects of Maltol, a Flavoring Agent, on Carbon Tetrachloride-Induced Acute Liver Injury in Mice via Inhibiting Apoptosis and Inflammatory Response. Molecules. 2018; 23(9):2120. https://doi.org/10.3390/molecules23092120
Chicago/Turabian StyleLiu, Wei, Zi Wang, Jin-gang Hou, Yan-dan Zhou, Yu-fang He, Shuang Jiang, Ying-ping Wang, Shen Ren, and Wei Li. 2018. "The Liver Protection Effects of Maltol, a Flavoring Agent, on Carbon Tetrachloride-Induced Acute Liver Injury in Mice via Inhibiting Apoptosis and Inflammatory Response" Molecules 23, no. 9: 2120. https://doi.org/10.3390/molecules23092120
APA StyleLiu, W., Wang, Z., Hou, J.-g., Zhou, Y.-d., He, Y.-f., Jiang, S., Wang, Y.-p., Ren, S., & Li, W. (2018). The Liver Protection Effects of Maltol, a Flavoring Agent, on Carbon Tetrachloride-Induced Acute Liver Injury in Mice via Inhibiting Apoptosis and Inflammatory Response. Molecules, 23(9), 2120. https://doi.org/10.3390/molecules23092120