Structure-Immunogenicity Relationship of α- and β-Tetrasaccharide Glycoforms from Bacillus anthracis Exosporium and Fragments Thereof


Abstract
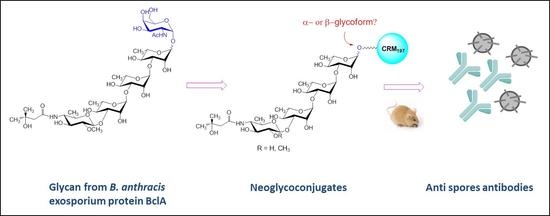
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Synthesis of the Glycoconjugates
2.2. Immunological Evalution
3. Materials and Methods
3.1. Glycoconjugation
3.2. MALDI-TOF MS Analysis
3.3. Immunogenicity of Conjugates in Mice
3.4. ELISA Analysis Using Bovine Serum Albumin (BSA) Conjugates as Coating Reagent
3.5. ELISA Analysis to Evaluate Anti-Glycan Binding to B. anthracis Spores
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanna, P. Anthrax pathogenesis and host response. Curr. Top. Microbiol. Immunol. 1998, 225, 13–35. [Google Scholar] [PubMed]
- Wang, J.Y.; Roehrl, M.H. Anthrax vaccine design: Strategies to achieve comprehensive protection against spore, bacillus, and toxin. Med. Immunol. 2005, 4. [Google Scholar] [CrossRef] [PubMed]
- Webb, G. Being prepared: Modeling the response to an anthrax attack. Ann. Intern. Med. 2005, 142, 667–668. [Google Scholar] [CrossRef] [PubMed]
- Young, J.A.; Collier, R.J. Anthrax toxin: Receptor binding, internalization, pore formation, and translocation. Annu. Rev. Biochem. 2007, 76, 243–265. [Google Scholar] [CrossRef] [PubMed]
- Bouzanias, D.G. Medical countermeasures to protect humans from anthrax bioterrorism. Trends Microbiol. 2009, 17, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Daubenspeck, J.M.; Zeng, H.; Chen, P.; Dong, S.; Steichen, C.T.; Krishna, N.R.; Pritchard, D.G.; Turnbough, C.L., Jr. Novel oligosaccharide side chains of the collagen-like region of bcla, the major glycoprotein of the bacillus anthracis exosporium. J. Biol. Chem. 2004, 279, 30945–30953. [Google Scholar] [CrossRef] [PubMed]
- Berti, F.; Adamo, R. Recent mechanistic insights on glycoconjugate vaccines and future perspectives. ACS Chem. Biol. 2013, 8, 1653–1663. [Google Scholar] [CrossRef] [PubMed]
- Astronomo, R.D.; Burton, D.R. Carbohydrate vaccines: Developing sweet solutions to sticky situations? Nat. Rev. Drug Discov. 2010, 9, 308–324. [Google Scholar] [CrossRef] [PubMed]
- Adamo, R. Glycan surface antigens from Bacillus anthracis as vaccine targets: Current status and future perspectives. Exp. Rev. Vaccines 2014, 13, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Saksena, R.; Adamo, R.; Kováč, P. Studies toward a conjugate vaccine for anthrax. Synthesis and characterization of anthrose [4,6-dideoxy-4-(3-hydroxy-3-methylbutanamido)-2-o-methyl-d-glucopyranose] and its methyl glycosides. Carbohydr. Res. 2005, 340, 1591–1600. [Google Scholar] [CrossRef] [PubMed]
- Milhomme, O.; Grandjean, C. Synthetic efforts towards glycoconjugate-based vaccines active against anthrax. Curr. Org. Synth. 2014, 18, 291–311. [Google Scholar] [CrossRef]
- Oberli, M.A.; Horlacher, T.; Werz, D.B.; Seeberger, P.H. Synthetic oligosaccharide bacterial antigens to produce monoclonal antibodies for diagnosis and treatment of disease using bacillus anthracis as a case study. In Anticarbohydrate Antibodies; Kosma, P., Müller-Loennies, S., Eds.; Springer: Vienna, Austria, 2012; pp. 37–54. [Google Scholar]
- Werz, D.B.; Seeberger, P.H. Total synthesis of antigen bacillus anthracis tetrasaccharide—Creation of an anthrax vaccine candidate. Angew. Chem. Int. Ed. Engl. 2005, 44, 6315–6318. [Google Scholar] [CrossRef] [PubMed]
- Adamo, R.; Saksena, R.; Kováč, P. Synthesis of the beta anomer of the spacer-equipped tetrasaccharide side chain of the major glycoprotein of the bacillus anthracis exosporium. Carbohydr. Res. 2005, 340, 2579–2582. [Google Scholar] [CrossRef] [PubMed]
- Saksena, R.; Adamo, R.; Kováč, P. Synthesis of the tetrasaccharide side chain of the major glycoprotein of the bacillus anthracis exosporium. Bioorg. Med. Chem. Lett. 2006, 16, 615–617. [Google Scholar] [CrossRef] [PubMed]
- Adamo, R.; Saksena, R.; Kováč, P. Studies towards a conjugate vaccine for anthrax: Synthesis of the tetrasaccharide side chain of the bacillus anthracis exosporium. Helv. Chim. Acta 2006, 89, 1075–1088. [Google Scholar] [CrossRef]
- Saksena, R.; Adamo, R.; Kováč, P. Immunogens related to the synthetic tetrasaccharide side chain of the bacillus anthracis exosporium. Bioorg. Med. Chem. 2007, 15, 4283–4310. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.S.; Saile, E.; Zhong, W.; Buskas, T.; Carlson, R.; Kannenberg, E.; Reed, Y.; Quinn, C.P.; Boons, G.J. Synthesis and antigenic analysis of the Bcla glycoprotein oligosaccharide from the Bacillus anthracis exosporium. Chemistry 2006, 12, 9136–9149. [Google Scholar] [CrossRef] [PubMed]
- Crich, D.; Vinogradova, O. Synthesis of the antigenic tetrasaccharide side chain from the major glycoprotein of Bacillus anthracis exosporium. J. Org. Chem. 2007, 72, 6513–6520. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; O’Doherty, G.A. De novo asymmetric synthesis of the anthrax tetrasaccharide by a palladium-catalyzed glycosylation reaction. Angew. Chem. Int. Ed. Engl. 2007, 46, 5206–5208. [Google Scholar] [CrossRef] [PubMed]
- Dhenin, S.G.; Moreau, V.; Morel, N.; Nevers, M.C.; Volland, H.; Creminon, C.; Djedaini-Pilard, F. Synthesis of an anthrose derivative and production of polyclonal antibodies for the detection of anthrax spores. Carbohydr. Res. 2008, 343, 2101–2110. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Kováč, P. A convenient synthesis of furanose-free d-fucose per-o-acetates and a precursor for anthrose. Eur. J. Org. Chem. 2008, 2008, 1947–1952. [Google Scholar] [CrossRef]
- Milhomme, O.; Dhenin, S.G.; Djedaini-Pilard, F.; Moreau, V.; Grandjean, C. Synthetic studies toward the anthrax tetrasaccharide: Alternative synthesis of this antigen. Carbohydr. Res. 2012, 356, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Kováč, P. Conjugation-amenable tetrasaccharide of the side chain of the major glycoprotein of the bacillus anthracis exosporium: A large-scale preparation. Synthesis 2009, 545–550. [Google Scholar]
- Werz, D.B.; Adibekian, A.; Seeberger, P.H. Synthesis of a spore surface pentasaccharide of bacillus anthracis. Eur. J. Org. Chem. 2007, 2007, 1976–1982. [Google Scholar] [CrossRef]
- Tamborrini, M.; Werz, D.B.; Frey, J.; Pluschke, G.; Seeberger, P.H. Anticarbohydrate antibodies for the detection of anthrax spores. Angew. Chem. Int. Ed. Engl. 2006, 45, 6581–6582. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Carroll, G.T.; Turro, N.J.; Koberstein, J.T.; Kovac, P.; Saksena, R.; Adamo, R.; Herzenberg, L.A.; Steinman, L. Photogenerated glycan arrays identify immunogenic sugar moieties of bacillus anthracis exosporium. Proteomics 2007, 7, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Oberli, M.A.; Tamborrini, M.; Tsai, Y.-H.; Werz, D.B.; Horlacher, T.; Adibekian, A.; Gauss, D.; Moller, H.M.; Pluschke, G.; Seeberger, P.H. Molecular analysis of carbohydrate-antibody interactions: Case study using a bacillus anthracis tetrasaccharide. J. Am. Chem. Soc. 2010, 132, 10239–10241. [Google Scholar] [CrossRef] [PubMed]
- Milhomme, O.; Köhler, S.M.; Ropartz, D.; Lesur, D.; Pilard, S.; Djedaïni-Pilard, F.; Beyerb, W.; Grandjean, C. Synthesis and immunochemical evaluation of a non-methylated disaccharide analogue of the anthrax tetrasaccharide. Org. Biomol. Chem. 2012, 10, 8524–8532. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Adamo, R.; Costantino, P. Protein carriers for glycoconjugate vaccines: History, selection criteria, characterization and new trends. Molecules 2018, 23, 1451. [Google Scholar] [CrossRef] [PubMed]
- Adamo, R.; Romano, M.R.; Berti, F.; Leuzzi, R.; Tontini, M.; Danieli, E.; Cappelletti, E.; Cakici, O.S.; Swennen, E.; Pinto, V.; et al. Phosphorylation of the synthetic hexasaccharide repeating unit is essential for the induction of antibodies to Clostridium difficile PSII cell wall polysaccharide. ACS Chem. Biol. 2012, 7, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Trinh, M.N.; Kováč, P. Conjugation of carbohydrates to proteins using di (triethylene glycol monomethyl ether) squaric acid ester revisited. Carbohydr. Res. 2018, 456, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Swiecki, M.K.; Lisanby, M.W.; Shu, F.; Turnbough, C.L.; Kearney, J.F. Monoclonal antibodies for bacillus anthracis spore detection and functional analyses of spore germination and outgrowth. J. Immunol. 2006, 176, 6076–6084. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds 1–5 are available from the authors. |






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Ricco, R.; Ventura, C.L.; Carboni, F.; Saksena, R.; Kováč, P.; Adamo, R. Structure-Immunogenicity Relationship of α- and β-Tetrasaccharide Glycoforms from Bacillus anthracis Exosporium and Fragments Thereof. Molecules 2018, 23, 2079. https://doi.org/10.3390/molecules23082079
De Ricco R, Ventura CL, Carboni F, Saksena R, Kováč P, Adamo R. Structure-Immunogenicity Relationship of α- and β-Tetrasaccharide Glycoforms from Bacillus anthracis Exosporium and Fragments Thereof. Molecules. 2018; 23(8):2079. https://doi.org/10.3390/molecules23082079
Chicago/Turabian StyleDe Ricco, Riccardo, Christy L. Ventura, Filippo Carboni, Rina Saksena, Pavol Kováč, and Roberto Adamo. 2018. "Structure-Immunogenicity Relationship of α- and β-Tetrasaccharide Glycoforms from Bacillus anthracis Exosporium and Fragments Thereof" Molecules 23, no. 8: 2079. https://doi.org/10.3390/molecules23082079
APA StyleDe Ricco, R., Ventura, C. L., Carboni, F., Saksena, R., Kováč, P., & Adamo, R. (2018). Structure-Immunogenicity Relationship of α- and β-Tetrasaccharide Glycoforms from Bacillus anthracis Exosporium and Fragments Thereof. Molecules, 23(8), 2079. https://doi.org/10.3390/molecules23082079