Aqueous-Methanol Extracts of Orange-Fleshed Sweet Potato (Ipomoea batatas) Ameliorate Oxidative Stress and Modulate Type 2 Diabetes Associated Genes in Insulin Resistant C2C12 Cells


Abstract
1. Introduction
2. Results and Discussion
2.1. Quantification of Specific Polyphenols in the Extracts
2.2. Effect of Sweet Potato Extracts on Oxidative Stress in Insulin Resistant C2C12 Cells
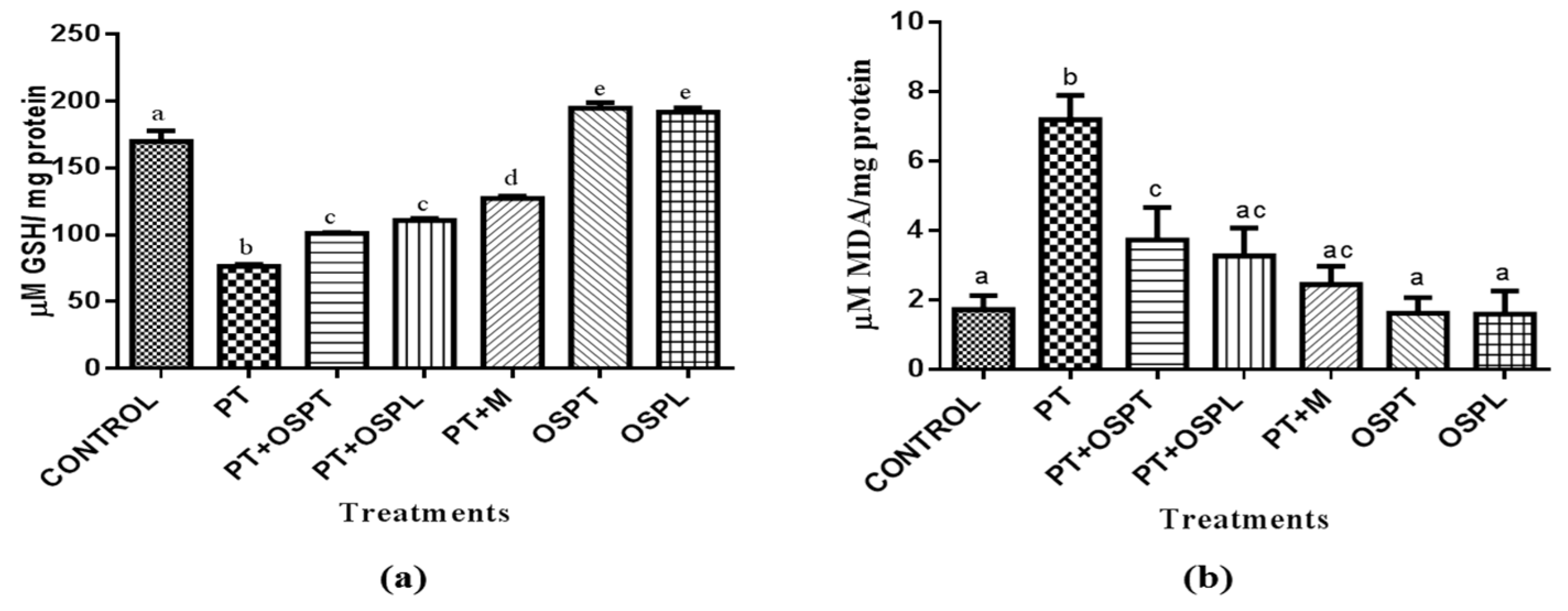
2.2.1. Effect of Sweet Potato Extracts on Total Glutathione Status and Lipid Peroxidation
2.2.2. Effect of Sweet Potato Extracts on Antioxidant Enzymes
2.2.3. Effect of Sweet Potato Extracts on Antioxidant Capacity
2.3. Effect of Sweet Potato Extracts on Expression Levels of Glut4, Nrf1, Mef2a, Cpt1 and Acc2 Genes
3. Materials and Methods
3.1. C2C12 Cells Subculture and Differentiation
3.2. Plant Samples and Preparation of Extracts
3.3. Quantification of Specific Polyphenols by HPLC Analysis
3.4. MTT Cytotoxicity Assay
3.5. Preparation of Palmitate and Induction of Insulin Resistance
3.6. Treatment of C2C12 Cells with Sweet Potato Extracts
3.7. Oxidative Stress and Antioxidant Activity Assays
3.7.1. Bradford Protein Assay
3.7.2. Glutathione Status and Glutathione Peroxidase Activity Assays
3.7.3. Catalase Activity Assay
3.7.4. Lipid Peroxidation Assay
3.7.5. Antioxidant Activity Assays
3.8. Total RNA Extraction and cDNA Synthesis
3.9. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- DeFronzo, R.A.; Tripathy, D. Skeletal muscle insulin resistance is the primary defect in type 2 diabetes. Diabetes Care 2009, 32, S157–S163. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R. Insulin resistance and type 2 diabetes. Diabetes 2012, 61, 778–779. [Google Scholar] [CrossRef] [PubMed]
- Morales-González, J.A. (Ed.) Oxidative Stress and Chronic Degenerative Diseases—A Role for Antioxidants; InTech: Rijeka, Croatia, 2013; 512p, ISBN 978-9-53-511123-8. [Google Scholar]
- Keane, K.N.; Cruzat, V.F.; Carlessi, R.; de Bittencourt, P.I.H.; Newsholme, P. Molecular events linking oxidative stress and inflammation to insulin resistance and β-cell dysfunction. Oxid. Med. Cell. Longev. 2015, 2015, 181643. [Google Scholar] [CrossRef] [PubMed]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J. Diabetes 2015, 6, 456–480. [Google Scholar] [CrossRef] [PubMed]
- Cahova, M.; Vavrinkova, H.; Kazdova, L. Glucose-fatty acid interaction in skeletal muscle and adipose tissue in insulin resistance. Physiol. Res. 2007, 56, 1–15. [Google Scholar] [PubMed]
- Pessin, J.E.; Saltiel, A.R. Signaling pathways in insulin action: Molecular targets of insulin resistance. J. Clin. Investig. 2000, 106, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Czech, M.P. The GLUT4 glucose transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed]
- McGee, S.L.; Hargreaves, M. Exercise and skeletal muscle glucose transporter 4 expression: Molecular mechanisms. Clin. Exp. Pharmacol. Physiol. 2006, 33, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, Z.; Ayeleso, A.; Mukwevho, E. Control of carbohydrate and lipid metabolism by NRF-1 and sirtuins: Implications on type 2 diabetes and obesity. Chem. Biol. Lett. 2014, 1, 66–76. [Google Scholar]
- Wu, H.; Deng, X.; Shi, Y.; Su, Y.; Wei, J.; Duan, H. PGC-1α, glucose metabolism and type 2 diabetes mellitus. J. Endocrinol. 2016, 229, R99–R115. [Google Scholar] [CrossRef] [PubMed]
- Wakil, S.J.; Abu-Elheiga, L.A. Fatty acid metabolism: Target for metabolic syndrome. J. Lipid Res. 2009, 50, S138–S143. [Google Scholar] [CrossRef] [PubMed]
- Fillmore, N.; Mori, J.; Lopaschuk, G.D. Mitochondrial fatty acid oxidation alterations in heart failure, ischaemic heart disease and diabetic cardiomyopathy. Br. J. Pharmacol. 2014, 171, 2080–2090. [Google Scholar] [CrossRef] [PubMed]
- Bruce, C.R.; Hoy, A.J.; Turner, N.; Watt, M.J.; Allen, T.L.; Carpenter, K.; Kraegen, E.W. Overexpression of carnitine palmitoyltransferase-1 in skeletal muscle is sufficient to enhance fatty acid oxidation and improve high-fat diet–induced insulin resistance. Diabetes 2009, 58, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Cui, F.; Meng, L.; Chen, G.; Li, Z. Carnitine Palmitoyltransferase Inhibitor in Diabetes. J. Mol. Genet. Med. 2016, 10, 238. [Google Scholar]
- Woo, Y.; Lee, H.; Jeong, Y.S.; Shin, G.Y.; Oh, J.G.; Kim, J.S.; Oh, J. Antioxidant Potential of Selected Korean Edible Plant Extracts. BioMed Res. Int. 2017, 2017, 7695605. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Pari, L. Ameliorative effect of diosmin, a citrus flavonoid against streptozotocin-nicotinamide generated oxidative stress induced diabetic rats. Chem. Biol. Interact. 2012, 195, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Nie, S.; Zhu, F. Chemical constituents and health effects of sweet potato. Food Res. Int. 2016, 89, 90–116. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Zeng, M.; Chen, J.; Jiao, Y.; Niu, F.; Tao, G.; Zhang, S.; Qin, F.; He, Z. Identification and quantitation of anthocyanins in purple-fleshed sweet potatoes cultivated in China by UPLC-PDA and UPLC-QTOF-MS/MS. J. Agric. Food Chem. 2015, 64, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.K.; Rana, Z.H.; Islam, S.N. Comparison of the proximate composition, total carotenoids and total polyphenol content of nine orange-fleshed sweet potato varieties grown in Bangladesh. Foods 2016, 5, 64. [Google Scholar] [CrossRef] [PubMed]
- El Gharras, H. Polyphenols: Food sources, properties and applications—A review. Int. J. Food Sci. Technol. 2009, 44, 2512–2518. [Google Scholar] [CrossRef]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.; Ferreira, I.C. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Ogunrinola, O.O.; Fajana, O.O.; Olaitan, S.N.; Adu, O.B.; Akinola, M.O. Anti-diabetic activity of Ipomoea batatas leaves extract: Effects on hepatic enzymes in alloxan-induced diabetic rats. Res. J. Med. Plants 2015, 9, 227–233. [Google Scholar] [CrossRef]
- Pal, S.; Gautam, S.; Mishra, A.; Maurya, R.; Srivastava, A.K. Antihyperglycemic and antidyslipidemic potential of Ipomoea batatas leaves in validated diabetic animal models. Int. J. Pharm. Pharm. Sci. 2015, 7, 176–186. [Google Scholar]
- Donado-Pestana, C.M.; Salgado, J.M.; de Oliveira Rios, A.; dos Santos, P.R.; Jablonski, A. Stability of carotenoids, total phenolics and in vitro antioxidant capacity in the thermal processing of orange-fleshed sweet potato (Ipomoea batatas Lam.) cultivars grown in Brazil. Plant Foods Hum. Nutr. 2012, 67, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Teow, C.C.; Truong, V.D.; McFeeters, R.F.; Thompson, R.L.; Pecota, K.V.; Yencho, G.C. Antioxidant activities, phenolic and β-carotene contents of sweet potato genotypes with varying flesh colours. Food Chem. 2007, 103, 829–838. [Google Scholar] [CrossRef]
- Koala, M.; Hema, A.; Somé, K.; Palé, E.; Sérémé, A.; Belem, J.; Nacro, M. Evaluation of Eight Orange Fleshed Sweetpotato (OFSP) Varieties for Their Total Antioxidant, Total Carotenoid and Polyphenolic. Evaluation 2013, 3, 67–72. [Google Scholar]
- Tang, Y.; Cai, W.; Xu, B. Profiles of phenolics, carotenoids and antioxidative capacities of thermal processed white, yellow, orange and purple sweet potatoes grown in Guilin, China. Food Sci. Hum. Wellness 2015, 4, 123–132. [Google Scholar] [CrossRef]
- Wang, L.; Yue, Z.; Guo, M.; Fang, L.; Bai, L.; Li, X.; Zhao, H. Dietary flavonoid hyperoside induces apoptosis of activated human LX-2 hepatic stellate cell by suppressing canonical NF-κB signaling. BioMed Res. Int. 2016, 2016, 1068528. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.F.; Tu, Z.C.; Zhang, L.; Wang, H.; Wen, Q.H.; Huang, T. Antioxidant activities and polyphenols of sweet potato (Ipomoea batatas L.) leaves extracted with solvents of various polarities. Food Biosci. 2016, 15, 11–18. [Google Scholar] [CrossRef]
- Harrison, H.F.; Peterson, J.K.; Snook, M.E.; Bohac, J.R.; Jackson, D.M. Quantity and potential biological activity of caffeic acid in sweet potato [Ipomoea batatas (L.) Lam.] storage root periderm. J. Agric. Food Chem. 2003, 51, 2943–2948. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, İ. Antioxidant activity of caffeic acid (3,4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Masek, A.; Chrzescijanska, E.; Latos, M. Determination of antioxidant activity of caffeic acid and p-coumaric acid by using electrochemical and spectrophotometric assays. Int. J. Electrochem. Sci. 2016, 11, 10644–10658. [Google Scholar] [CrossRef]
- Olivier, D.K.; van Wyk, B.E.; van Heerden, F.R. The chemotaxonomic and medicinal significance of phenolic acids in Arctopus and Alepidea (Apiaceae subfamily Saniculoideae). Biochem. Syst. Ecol. 2008, 36, 724–729. [Google Scholar] [CrossRef]
- Jo, S.H.; Ka, E.H.; Lee, H.S.; Apostolidis, E.; Jang, H.D.; Kwon, Y.I. Comparison of antioxidant potential and rat intestinal a-glucosidases inhibitory activities of quercetin, rutin, and isoquercetin. Int. J. Appl. Res. Nat. Prod. 2009, 2, 52–60. [Google Scholar]
- Dhanya, R.; Arya, A.D.; Nisha, P.; Jayamurthy, P. Quercetin, a lead compound against type 2 diabetes ameliorates glucose uptake via AMPK pathway in skeletal muscle cell line. Front. Pharmacol. 2017, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.R.; Nachbar, R.T.; Gorjao, R.; Vinolo, M.A.; Festuccia, W.T.; Lambertucci, R.H.; Hirabara, S.M. Mechanisms underlying skeletal muscle insulin resistance induced by fatty acids: Importance of the mitochondrial function. Lipids Health Dis. 2012, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Gross, M.; Lee, D.H.; Holvoet, P.; Himes, J.H.; Shikany, J.M.; Jacobs, D.R. Oxidative stress and insulin resistance. Diabetes Care 2009, 32, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Mazibuko, S.E.; Muller, C.J.F.; Joubert, E.; De Beer, D.; Johnson, R.; Opoku, A.R.; Louw, J. Amelioration of palmitate-induced insulin resistance in C2C12 muscle cells by rooibos (Aspalathus linearis). Phytomedicine 2013, 20, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Takamura, T.; Matsuzawa-Nagata, N.; Takayama, H.; Misu, H.; Noda, H.; Nabemoto, S.; Kurita, S.; Ota, T.; Ando, H.; et al. Palmitate induces insulin resistance in H4IIEC3 hepatocytes through reactive oxygen species produced by mitochondria. J. Biol. Chem. 2009, 284, 14809–14818. [Google Scholar] [CrossRef] [PubMed]
- Dai Ly, L.; Xu, S.; Choi, S.K.; Ha, C.M.; Thoudam, T.; Cha, S.K.; Wiederkehr, A.; Wollheim, C.B.; Lee, I.K.; Park, K.S. Oxidative stress and calcium dysregulation by palmitate in type 2 diabetes. Exp. Mol. Med. 2017, 49, e291. [Google Scholar]
- Diniz Vilela, D.; Gomes Peixoto, L.; Teixeira, R.R.; Belele Baptista, N.; Carvalho Caixeta, D.; Vieira de Souza, A.; Machado, H.L.; Pereira, M.N.; Sabino-Silva, R.; Espindola, F.S. The role of metformin in controlling oxidative stress in muscle of diabetic rats. Oxid. Med. Cell. Longev. 2016, 2016, 6978625. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Glutathione homeostasis and functions: potential targets for medical interventions. J. Amino. Acids 2012, 2012, 736837. [Google Scholar] [CrossRef] [PubMed]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Kalkan, I.H.; Suher, M. The relationship between the level of glutathione, impairment of glucose metabolism and complications of diabetes mellitus. Pak. J. Med. Sci. 2013, 29, 938–942. [Google Scholar] [CrossRef]
- Calabrese, V.; Cornelius, C.; Leso, V.; Trovato-Salinaro, A.; Ventimiglia, B.; Cavallaro, M.; Castellino, P. Oxidative stress, glutathione status, sirtuin and cellular stress response in type 2 diabetes. Biochim. Biophys. Acta 2012, 1822, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Goutzourelas, N.; Stagos, D.; Demertzis, N.; Mavridou, P.; Karterolioti, H.; Georgadakis, S.; Kerasioti, E.; Aligiannis, N.; Skaltsounis, L.; Statiri, A.; et al. Effects of polyphenolic grape extract on the oxidative status of muscle and endothelial cells. Hum. Exp. Toxicol. 2014, 33, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Goutzourelas, N.; Stagos, D.; Spanidis, Y.; Liosi, M.; Apostolou, A.; Priftis, A.; Haroutounian, S.; Spandidos, D.A.; Tsatsakis, A.M.; Kouretas, D. Polyphenolic composition of grape stem extracts affects antioxidant activity in endothelial and muscle cells. Mol. Med. Rep. 2015, 12, 5846–5856. [Google Scholar] [CrossRef] [PubMed]
- Niki, E.; Yoshida, Y.; Saito, Y.; Noguchi, N. Lipid peroxidation: Mechanisms, inhibition, and biological effects. Biochem. Biophys. Res. Commun. 2005, 338, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Umeno, A.; Shichiri, M. Lipid peroxidation biomarkers for evaluating oxidative stress and assessing antioxidant capacity in vivo. J. Clin. Biochem. Nutr. 2013, 52, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, V.K.; Agrawal, P.; Park, Y.H. Phytochemicals, Antioxidant and Anti-Lipid Peroxidation Activities of Ethanolic Extract of a Medicinal Plant, Andrographis paniculata. J. Food Biochem. 2014, 38, 584–591. [Google Scholar] [CrossRef]
- Kada, S.; Bouriche, H.; Senator, A.; Demirtaş, I.; Özen, T.; Çeken Toptanci, B.; Kızıl, G.; Kızıl, M. Protective activity of Hertia cheirifolia extracts against DNA damage, lipid peroxidation and protein oxidation. Pharm. Biol. 2017, 55, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Lee, T.H. Antioxidant enzymes as redox-based biomarkers: A brief review. BMB Rep. 2015, 48, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Tappel, A. Selenium-glutathione peroxidase: Properties and synthesis. Curr. Top. Cell. Regul. 2014, 24, 87–96. [Google Scholar]
- Torres, L.L.; Quaglio, N.B.; de Souza, G.T.; Garcia, R.T.; Dati, L.M.M.; Moreira, W.L.; Nitrini, R. Peripheral oxidative stress biomarkers in mild cognitive impairment and Alzheimer's disease. J. Alzheimer Dis. 2011, 26, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Kabel, A.M. Free radicals and antioxidants: Role of enzymes and nutrition. World J. Nutr. Health 2014, 2, 35–38. [Google Scholar]
- Doehner, W.; Gathercole, D.; Cicoira, M.; Krack, A.; Coats, A.J.; Camici, P.G.; Anker, S.D. Reduced glucose transporter GLUT4 in skeletal muscle predicts insulin resistance in non-diabetic chronic heart failure patients independently of body composition. Int. J. Cardiol. 2010, 138, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Kampmann, U.; Christensen, B.; Nielsen, T.S.; Pedersen, S.B.; Ørskov, L.; Lund, S.; Jessen, N. GLUT4 and UBC9 protein expression is reduced in muscle from type 2 diabetic patients with severe insulin resistance. PLoS ONE 2011, 6, e27854. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.M.; Hu, J.; Barnes, R.M.; Heidt, A.B.; Cornelissen, I.; Black, B.L. Myocyte enhancer factor 2C function in skeletal muscle is required for normal growth and glucose metabolism in mice. Skelet. Muscle 2015, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Schreurs, M.; Kuipers, F.; Van Der Leij, F.R. Regulatory enzymes of mitochondrial β-oxidation as targets for treatment of the metabolic syndrome. Obes. Rev. 2010, 11, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.S.; Savage, D.B.; Abu-Elheiga, L.; Liu, Z.X.; Kim, S.; Kulkarni, A.; Zhang, D. Continuous fat oxidation in acetyl–CoA carboxylase 2 knockout mice increases total energy expenditure, reduces fat mass, and improves insulin sensitivity. Proc. Natl. Acad. Sci. USA 2007, 104, 16480–16485. [Google Scholar] [CrossRef] [PubMed]
- Rosa, G.; Manco, M.; Vega, N. Insulin sensitivity inversely relates to skeletal muscle ACC2 mRNA in formerly obese subjects. Obes. Res. 2003, 11, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Yang, Y.R.Y.; Mo, Z.J.; Ding, Y.; Jiang, W.J. Silibinin improves palmitate-induced insulin resistance in C2C12 myotubes by attenuating IRS-1/PI3K/Akt pathway inhibition. Braz. J. Med. Biol. Res. 2015, 48, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Haghani, K.; Pashaei, S.; Vakili, S.; Taheripak, G.; Bakhtiyari, S. TNF-α knockdown alleviates palmitate-induced insulin resistance in C2C12 skeletal muscle cells. Biochem. Biophys. Res. Commun. 2015, 460, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W. Selenium: Biochemical role as a component of glutathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Hadwan, M.H.; Abed, H.N. Data supporting the spectrophotometric method for the estimation of catalase activity. Data Brief 2016, 6, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Method Enzymol. 1978, 52, 302–310. [Google Scholar]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
Sample Availability: Samples of the extracts are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Retention Time (min) | Parent Ion (m/z) | Product Ion (m/z) | Amount of Phenolic Acids and Flavonoids (mg/g of Extract) | |||
|---|---|---|---|---|---|---|---|
| LEAVES | TUBERS | ||||||
| AQ | AQ-ME | AQ | AQ-ME | ||||
| Caffeic acid | 14.59 | 181 | 163 | 0.98 ± 0.003 | 3.76 ± 0.003 | 1.7 ± 0.000 | 19.4 ± 0.042 |
| Catechin | 16.6 | 291.1 | 139 | ND | 0.01 ± 0.010 | ND | ND |
| Hyperoside | 15.69 | 465.1 | 303.1 | 1.39 ± 0.003 | 139.83 ± 0.006 | 0.22 ± 0.007 | 9.78 ± 0.003 |
| Iso-orientin | 14.8 | 449.1 | 299 | ND | ND | ND | ND |
| Isovanillic acid | 10.9 | 169 | 65.1 | 0.04 ± 0.005 | 0.13 ± 0.003 | 0.14 ± 0.007 | 0.04 ± 0.000 |
| Kaempferol | 15.8 | 287.1 | 167.2 | ND | 0.09 ± 0.007 | ND | ND |
| Orientin | 14.7 | 449.1 | 329 | ND | ND | ND | ND |
| Protocatechuic acid | 10.45 | 155 | 65.1 | 0.45 ± 0.004 | 1.22 ± 0.014 | 2.5 ± 0.008 | 0.08 ± 0.003 |
| Quercetin | 15.65 | 303.1 | 153 | 0.28 ± 0.003 | 23.36 ± 0.000 | 0.03 ± 0.001 | 1.76 ± 0.004 |
| Rutin | 15.34 | 611.2 | 303 | 0.73 ± 0.007 | 69.67 ± 0.007 | ND | 2.91 ± 0.007 |
| Vanyllic acid | 9.854 | 169 | 65.1 | 0.26 ± 0.0023 | 0.4 ± 0.014 | 0.45 ± 0.002 | 0.81 ± 0.127 |
| Vitexin | 15.4 | 433.1 | 313 | ND | ND | ND | ND |
| Concentration (µg/mL) | OSPT | OSPL |
|---|---|---|
| Cell Viability (%) | Cell Viability (%) | |
| 10 | 97.20 ± 1.56 a | 96.14 ± 0.08 a |
| 20 | 96.12 ± 1.04 a | 95.12 ± 1.72 a |
| 50 | 91.47 ± 2.16 a | 93.88 ± 0.09 a |
| 100 | 90.28 ± 2.12 a | 90.96 ± 2.34 a |
| 200 | 90.09 ± 1.87 a | 78.23 ± 1.11 b |
| 500 | 91.37 ± 1.68 a | 75.16 ± 1.46 b |
| 1000 | 76.31 ± 2.01 b | 77.91 ± 0.65 b |
| Treatment Groups | FRAP Values (µM AAE/mg Protein) | TEAC Values (µM TE/mg Protein) |
|---|---|---|
| CONTROL | 271.0 ± 4.17 a | 107.2 ± 1.68 a |
| PT | 102.2 ± 5.06 b | 39.8 ± 1.80 b |
| PT + OSPT | 167.7 ± 0.54 c | 71.94 ± 8.0 c |
| PT + OSPL | 172.1 ± 1.94 c | 76.19 ± 7.6 c |
| PT + M | 251.3 ± 2.50 a | 91.06 ± 1.40 a |
| OSPT | 299.8 ± 2.5 d | 127.9 ± 2.10 d |
| OSPL | 296.9 ± 7.4 d | 126.3 ± 2.51 d |
| Treatment Groups | Palmitate (PT) | Tuber Extracts (OSPT) | Leaf Extracts (OSPL) | Metformin (M) |
|---|---|---|---|---|
| CONTROL | − | − | − | − |
| PT | + | − | − | − |
| PT + OSPT | + | + | − | − |
| PT + OSPL | + | − | + | − |
| OSPT | − | + | − | − |
| OSPL | − | − | + | − |
| PT + M | + | − | − | + |
| Gene | Primer Sequence(5′-3′) |
|---|---|
| glut4 | Forward-AAGATGGCCACGGAGAGA Reverse-GTGGGTTGTGGCAGTGAGTC |
| nrf1 | Forward-AAACACAAACTCAGGCCACC Reverse-CCATCAGCCACAGCAGAGTA |
| mef2a | Forward-GTGTACTCAGCAATGCCGAC Reverse-AACCCTGAGATAACTGCCCTC |
| cpt1 | Forward-CCAGGCTACAGTGGGACATT Reverse-GAACTTGCCCATGTCCTTGT |
| acc2 | Forward-GTCCTCATCATGAACGGCTG Reverse-AGGACAGTGGGGTCGTTTTC |
| gapdh | Forward-GCACAGTCAAGGCCGAGAAT Reverse-GCCTTCTCCATGGTGGTGAA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayeleso, T.B.; Ramachela, K.; Mukwevho, E. Aqueous-Methanol Extracts of Orange-Fleshed Sweet Potato (Ipomoea batatas) Ameliorate Oxidative Stress and Modulate Type 2 Diabetes Associated Genes in Insulin Resistant C2C12 Cells. Molecules 2018, 23, 2058. https://doi.org/10.3390/molecules23082058
Ayeleso TB, Ramachela K, Mukwevho E. Aqueous-Methanol Extracts of Orange-Fleshed Sweet Potato (Ipomoea batatas) Ameliorate Oxidative Stress and Modulate Type 2 Diabetes Associated Genes in Insulin Resistant C2C12 Cells. Molecules. 2018; 23(8):2058. https://doi.org/10.3390/molecules23082058
Chicago/Turabian StyleAyeleso, Taiwo Betty, Khosi Ramachela, and Emmanuel Mukwevho. 2018. "Aqueous-Methanol Extracts of Orange-Fleshed Sweet Potato (Ipomoea batatas) Ameliorate Oxidative Stress and Modulate Type 2 Diabetes Associated Genes in Insulin Resistant C2C12 Cells" Molecules 23, no. 8: 2058. https://doi.org/10.3390/molecules23082058
APA StyleAyeleso, T. B., Ramachela, K., & Mukwevho, E. (2018). Aqueous-Methanol Extracts of Orange-Fleshed Sweet Potato (Ipomoea batatas) Ameliorate Oxidative Stress and Modulate Type 2 Diabetes Associated Genes in Insulin Resistant C2C12 Cells. Molecules, 23(8), 2058. https://doi.org/10.3390/molecules23082058