
Essential Oils as Feed Additives—Future Perspectives


Abstract
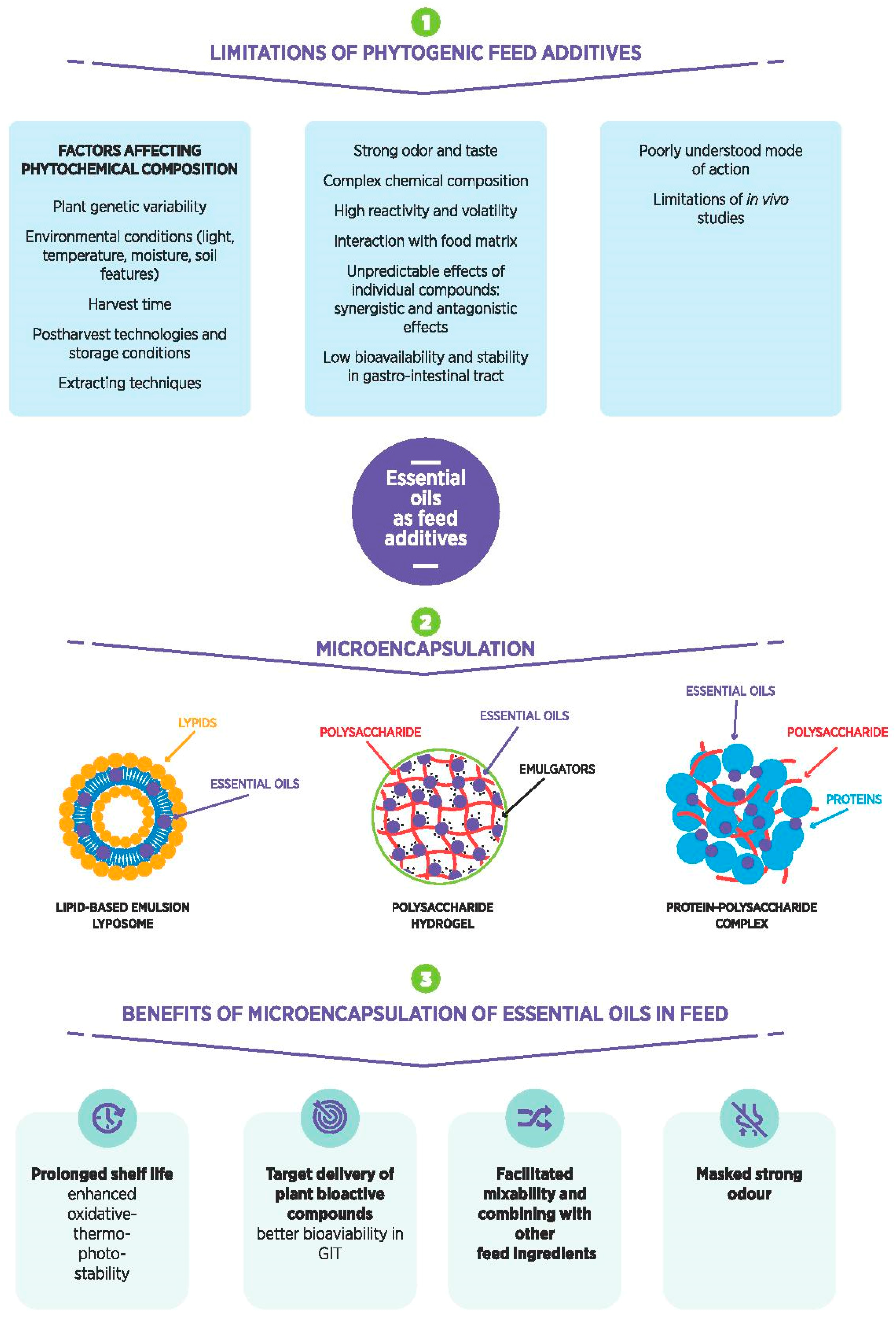
1. Introduction: Phytogenic Feed Additives
2. Essential Oils as Bioactive Plant Metabolites and Phytogenic Additives
2.1. Essential Oil Composition and Biosynthesis
2.2. Isolation and Identification of EO Components
2.3. Biological Effects of Essential Oils and Mechanisms of Activity
3. Effects of PFA and EOs on Poultry and Pigs’ Production
4. Limitations of PFA Application
5. Microencapsulation of EOs and Its Importance in Feed Additives
5.1. Polymer-Based Particles
5.2. Lipid-Based Particles
5.3. OilCarriers in Pig and Poultry Production
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Durmic, Z.; Blache, D. Bioactive plants and plant products: Effects on animal function, health and welfare. Anim. Feed Sci. Technol. 2012, 176, 150–162. [Google Scholar] [CrossRef]
- Puvača, N.; Stanaćev, V.; Glamočić, D.; Lević, J.; Perić, L.; Milić, D. Beneficial effects of phytoadditives in broiler nutrition. World Poult. Sci. J. 2013, 69, 27–34. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O.; et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef] [PubMed]
- Karásková, K.; Suchý, P.; Straková, E. Current use of phytogenic feed additives in animal nutrition: A review. Czech J. Anim. Sci. 2015, 60, 521–530. [Google Scholar] [CrossRef]
- World Health Organization. Legal Status of Traditional Medicine and Complementary/Alternative Medicine: A Worldwide Review; World Health Organization: Geneva, Switzerland, 2001. [Google Scholar]
- Windisch, W.; Schedle, K.; Plitzner, C.; Kroismayr, A. Use of phytogenic products as feed additives for swine and poultry. J. Anim. Sci. 2008, 86 (Suppl. 14), E140–E148. [Google Scholar] [CrossRef] [PubMed]
- Franz, C.; Baser, K.H.C.; Windisch, W. Essential oils and aromatic plants in animal feeding—An European perspective: A review. Flavour Fragr. J. 2010, 25, 327–340. [Google Scholar] [CrossRef]
- Casewell, M.; Friis, C.; Marco, E.; McMullin, P.; Phillips, I. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.B.; Ferasyi, T.R. Clean, Green, Ethical (CGE) Management: What Research Do We Really Need? Int. J. Trop. Vet. Biomed. Res. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Burdock, G.A.; Carabin, I.G. Generally recognized as safe (GRAS): History and description. Toxicol. Lett. 2004, 150, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Déciga-Campos, M.; Rivero-Cruz, I.; Arriaga-Alba, M.; Castañeda-Corral, G.; Angeles-López, G.E.; Navarrete, A.; Mata, R. Acute toxicity and mutagenic activity of Mexican plants used in traditional medicine. J. Ethnopharmacol. 2007, 110, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.A.; Carson, C.F. Antibacterial and Antifungal Activities of Essential Oils. In Lipids and Essential Oils as Antimicrobial Agents; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011; pp. 255–306. ISBN 9780470976616. [Google Scholar]
- Miguel, M.G. Antioxidant and anti-inflammatory activities of essential oils: A short review. Molecules 2010, 15, 9252–9287. [Google Scholar] [CrossRef] [PubMed]
- Brenes, A.; Roura, E. Essential oils in poultry nutrition: Main effects and modes of action. Anim. Feed Sci. Technol. 2010, 158, 1–14. [Google Scholar] [CrossRef]
- Zeng, Z.; Zhang, S.; Wang, H.; Piao, X. Essential oil and aromatic plants as feed additives in non-ruminant nutrition: A review. J. Anim. Sci. Biotechnol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Chowdhury, M.A.K.; Hou, Y.; Gong, J. Phytogenic Compounds as Alternatives to In-Feed Antibiotics: Potentials and Challenges in Application. Pathogens 2015, 4, 137–156. [Google Scholar] [CrossRef] [PubMed]
- Kis, R.K.; Bilkei, G. Effect of a phytogenic feed additive on weaning-to-estrus interval and farrowing rate in sows. J. Swine Health Prod. 2003, 11, 296–299. [Google Scholar]
- Hristov, A.N.; Lee, C.; Cassidy, T.; Heyler, K.; Tekippe, J.A.; Varga, G.A.; Corl, B.; Brandt, R.C. Effect of Origanum vulgare L. leaves on rumen fermentation, production, and milk fatty acid composition in lactating dairy cows. J. Dairy Sci. 2013, 96, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.H.; Anjum, M.A.; Parveen, A.; Khawaja, T.; Ashraf, N.M. Effects of black cumin seed (Nigella sativa L.) on performance and immune system in newly evolved crossbred laying hens. Vet. Quart. 2013, 33, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Nasir, Z.; Grashorn, M.A. Effects of Echinacea purpurea and Nigella sativa supplementation on broiler performance, carcass and meat quality. J. Anim. Feed Sci. 2010, 19, 94–104. [Google Scholar] [CrossRef]
- Bartoš, P.; Dolan, A.; Smutný, L.; Šístková, M.; Celjak, I.; Šoch, M.; Havelka, Z. Effects of phytogenic feed additives on growth performance and on ammonia and greenhouse gases emissions in growing-finishing pigs. Anim. Feed Sci. Technol. 2016, 212, 143–148. [Google Scholar] [CrossRef]
- Banik, B.K.; Durmic, Z.; Erskine, W.; Revell, C.K.; Vadhanabhuti, J.; McSweeney, C.S.; Padmanabha, J.; Flematti, G.R.; Algreiby, A.A.; Vercoe, P.E. Bioactive fractions from the pasture legume Biserrulapelecinus L. have an anti-methanogenic effect against key rumen methanogens. Anaerobe 2016, 39, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Broom, D.M.; Reefmann, N. Chicken welfare as indicated by lesions on carcases in supermarkets. Br. Poult. Sci. 2005, 46, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, M.; Che, T.M.; Bravo, D.; Maddox, C.W.; Pettigrew, J.E. Effects of capsicum oleoresin, garlic botanical, and turmeric oleoresin on gene expression profile of ileal mucosa in weaned pigs. J. Anim. Sci. 2014, 92, 3426–3440. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Matsuda, F. Metabolomics for functional genomics, systems biology, and biotechnology. Annu. Rev. Plant Biol. 2010, 61, 463–489. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, J.; Erehman, B.O.; Gohlke, T.; Wilhelm, R.; Preissner, M.; Dunkel, D. Super natural II-a database of natural products. Nucleic Acids Res. 2015, 43, 935–939. [Google Scholar] [CrossRef] [PubMed]
- Jamwa, K.; Bhattacharya, S.; Puri, S. Plant growth regulator mediated consequences of secondary metabolites in medicinal plants. J. Appl. Res. Med. Aromat. Plants 2018, 9, 26–38. [Google Scholar]
- Verpoorte, R. Exploration of nature’s chemodiversity: The role of secondary metabolites as leads in drug development. Drug Discov. Today 1998, 3, 232–238. [Google Scholar] [CrossRef]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Champagne, A.; Boutry, M. Proteomics of terpenoid biosynthesis and secretion in trichomes of higher plant species. Biochim. Biophys. Acta 2016, 1864, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Jacela, J.Y.; DeRouchey, J.M.; Tokach, M.D.; Goodband, R.D.; Nelssen, J.L.; Renter, D.G.; Dritz, S.S. Feed additives for swine: Fact sheets-prebiotics and probiotics, and phytogenics. J. Swine Health Prod. 2010, 18, 132–136. [Google Scholar] [CrossRef]
- Prakash, B.; Singh, P.; Kedia, A.; Dubey, N.K. Assessment of some essential oils as food preservatives based on antifungal, antiaflatoxin, antioxidant activities and in vivo efficacy in food system. Food Res. Int. 2012, 49, 201–208. [Google Scholar] [CrossRef]
- Prakash, B.; Kedia, A.; Mishra, P.K.; Dubey, N.K. Plant essential oils as food preservatives to control moulds, mycotoxin contamination and oxidative deterioration of agri-food commodities—Potentials and challenges. Food Control 2015, 47, 381–391. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential oils as ecofriendly biopesticides? Challenges and constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Ambrosio, C.M.S.; de Alencar, M.; de Sousa, R.L.M.; Moreno, A.M.; Da Gloria, E.M. Antimicrobial activity of several essential oils on pathogenic and beneficial bacteria. Ind. Crops Prod. 2017, 97, 128–136. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J.C. Factors affecting secondary metabolite production in plants: Volatile components and essential oils. Flavour Fragr. J. 2008, 23, 213–226. [Google Scholar] [CrossRef]
- Maenner, K.; Vahjen, W.; Simon, O. Studies on the effects of essential-oil-based feed additives on performance, ileal nutrient digestibility, and selected bacterial groups in the gastrointestinal tract of piglets. J. Anim. Sci. 2011, 89, 2106–2112. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.; Campi, E.M.; Jackson, W.R.; Patti, A.F. Effect of oxidative deterioration on flavour and aroma components of lemon oil. Food Chem. 2009, 112, 388–393. [Google Scholar] [CrossRef]
- Piva, A.; Pizzamiglio, V.; Morlacchini, M.; Tedeschi, M.; Piva, G. Lipid microencapsulation allows slow release of organic acids and natural identical flavors along the swine intestine. J. Anim. Sci. 2007, 85, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Bini, M.L.; Giuliani, C. The glandular trichomes of the labiatae. A review. Acta Hort. 2006, 723, 85–90. [Google Scholar] [CrossRef]
- Ciccarelli, D.; Andreuci, C.A.; Pagni, A.M. Translucent glands and secretory canals in Hypericum perforatum L. (Hypericaceae): Morphological, anatomical and histochemical studies during the course of ontogenesis. Ann. Bot. 2001, 88, 637–644. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Dajic Stevanovic, Z.; Nastovski, T.; Ristic, M.; Radanovic, D. Variability of essential oil composition of cultivated Feverfew (Tanacetum parthenium (L). Schultz Bip.) populations. J. Essent. Oil Res. 2009, 21, 292–294. [Google Scholar] [CrossRef]
- Stesevic, D.; Bozovic, M.; Tadic, V.; Rancic, D.; Dajic-Stevanovic, Z. Plant-part anatomy related composition of essential oils and phenolic compounds in Chaerophyllumcoloratum, a Balkan endemic species. Flora 2016, 220, 37–51. [Google Scholar] [CrossRef]
- Seow, Y.X.; Yeo, C.R.; Chung, H.L.; Yuk, H.-G. Plant essential oils as active antimicrobial agents. Crit. Rev. Food Sci. Nutr. 2014, 54, 625–644. [Google Scholar] [CrossRef] [PubMed]
- El Asbahani, A.; Miladi, K.; Badri, W.; Sala, M.; Addi, E.H.; Casabianca, H.; El Mousadik, A.; Hartmann, D.; Jilale, A.; Renaud, F.N.R.; et al. Essential oils: From extraction to encapsulation. Int. J. Pharm. 2015, 483, 220–243. [Google Scholar] [CrossRef] [PubMed]
- McCaskill, D.; Croteau, R. Some caveats for bioengineering terpenoid metabolism in plants. Trends Biotechnol. 1998, 16, 349–355. [Google Scholar] [CrossRef]
- Voon, C.H.; Bhat, R.; Rusul, G. Flower extracts and their essential oils as potential antimicrobial agents for fooduses and pharmaceutical applications. Compr. Rev. Food Sci. Food Saf. 2012, 11, 34–55. [Google Scholar] [CrossRef]
- Gang, D.R.; Wang, J.; Dudareva, N.; Nam, K.H.; Simon, J.E.; Lewinsohn, E.; Pichersky, E. An investigation of the storage and biosynthesis of phenylpropenes in sweet basil. Plant Physiol. 2001, 125, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Julsing, M.K.; Koulman, A.; Woerdenbag, H.J.; Quax, W.J.; Kayser, O. Combinatorial biosynthesis of medicinal plant secondary metabolites. Biomol. Eng. 2006, 23, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yang, C.; Li, C.; Zhao, Q.; Liu, L.; Fang, X.; Chen, X.Y. Recent advances in biosynthesis of bioactive compounds in traditional Chinese medicinal plants. Sci. Bull. 2016, 61, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Schuurink, R.C.; Caissard, J.C.; Hugueney, P.; Baudino, S. My Way: Noncanonical Biosynthesis Pathways for Plant Volatiles. Trends Plant Sci. 2016, 21, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Ludwiczuk, A.; Skalicka-Woźniak, K.; Georgiev, M.I. Terpenoids. In Pharmacognosy: Fundamentals, Applications and Strategies; Badal, S., Delgoda, R., Eds.; Elsevier/AP: Boston, MA, USA, 2017; pp. 233–266. [Google Scholar]
- Degenhardt, J.; Köllner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef] [PubMed]
- Ferhat, M.A.; Meklati, B.Y.; Chemat, F. Comparison of different isolation methods of essential oil from Citrus fruits: Cold pressing, hydrodistillation and microwave ‘dry’ distillation. Flavour Fragr. J. 2007, 22, 494–504. [Google Scholar] [CrossRef]
- Smelcerovic, A.; Djordjevic, A.; Lazarevic, J.; Stojanovic, G. Recent advances in analysis of essential oils. Curr. Anal. Chem. 2013, 9, 61–70. [Google Scholar] [CrossRef]
- Do, T.K.T.; Hadji-Minaglou, F.; Antoniotti, S.; Fernandez, X. Authenticity of essential oils. Trends Anal. Chem. 2015, 66, 146–157. [Google Scholar] [CrossRef]
- Edris, A.E. Pharmaceutical and therapeutic potentials of essential oils and their individual volatile constituents: A review. Phytother. Res. 2007, 21, 308–323. [Google Scholar] [CrossRef] [PubMed]
- George, D.R.; Smith, T.J.; Shiel, R.S.; Sparagano, O.A.E.; Guy, J.H. Mode of action and variability in efficacy of plant essential oils showing toxicity against the poultry red mite, Dermanyssusgallinae. Vet. Parasitol. 2009, 161, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Calo, J.R.; Crandall, F.G.; O’Bryan, C.A.; Ricke, S.C. Essential oils as antimicrobials in food systems—A review. Food Control 2015, 54, 111–119. [Google Scholar] [CrossRef]
- Dwivedy, A.K.; Kumar, M.; Upadhyay, N.; Prakash, B.; Dubey, N.K. Plant essential oils against food borne fungi and mycotoxins. Curr. Opin. Food Sci. 2016, 11, 16–21. [Google Scholar] [CrossRef]
- Lambert, R.J.W.; Skandamis, P.N.; Coote, P.; Nychas, G.J.E. A study of the minimum inhibitory concentration and mode of action of oregano essential oil, thymol and carvacrol. J. Appl. Microbiol. 2001, 91, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Kwon, Y.I.; Labbe, R.G.; Shetty, K. Inhibition of Helicobacter pylori and associated urease by Oregano and Cranberry phytochemical synergies. Appl. Environ. Microbiol. 2005, 71, 8558–8564. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zheng, H.; Tang, Y.; Yu, W.; Gong, Q. Eugenol inhibits quorum sensing at sub-inhibitory concentrations. Biotechnol. Lett. 2013, 35, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Defoirdt, T.; Brackman, G.; Coenye, T. Quorum sensing inhibitors: How strong is the evidence? Trends Microbiol. 2013, 21, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Bassolé, I.H.N.; Juliani, H.R. Essential oils in combination and their antimicrobial properties. Molecules 2012, 17, 3989–4006. [Google Scholar] [CrossRef] [PubMed]
- Becerril, R.; Nerin, C.; Gomez-Lus, R. Evaluation of bacterial resistance to essential oils and antibiotics after exposure to oregano and cinnamon essential oils. Foodborne Pathog. Dis. 2012, 9, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Langeveld, W.T.; Veldhuizen, E.J.; Burt, S.A. Synergy between essential oil components and antibiotics: A review. Crit. Rev. Microbiol. 2014, 40, 76–94. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Shen, S.C.; Chow, J.M.; Ko, C.H.; Tseng, S.W. Flavone inhibition of tumor growth via apoptosis in vitro and in vivo. Int. J. Oncol. 2004, 25, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, B.C.; Chang, C.J. Chemistry and biology of reactive oxygen species in signaling or stress responses. Nat. Chem. Biol. 2011, 7, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Fernandes, E.; Lima, J.L.F.C.; Mira, L.; Corvo, M.L. Molecular mechanisms of anti-inflammatory activity mediated by flavonoids. Curr. Med. Chem. 2008, 15, 1586–1605. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Liu, H.; Wang, S.; Wu, J.; Kluenter, A.-M. Potential of essential oils for poultry and pigs. Anim. Nutr. J. 2018. [Google Scholar] [CrossRef]
- Steiner, T.; Syed, B. Phytogenic Feed Additives in Animal Nutrition. In Medicinal and Aromatic Plants of the World. Medicinal and Aromatic Plants of the World; Máthé, Á., Ed.; Springer: Dordrecht, The Netherlands, 2015; Volume 1, pp. 403–423. ISBN 978-94-017-9810-5. [Google Scholar]
- Frankič, T.; Voljč, M.; Salobir, J.; Rezar, V. Use of herbs and spices and their extracts in animal nutrition. Acta Agric. Slov. 2009, 94, 95–102. [Google Scholar]
- Roper, T.J. Olfaction in birds. In Advances in the Study of Behavior; Slater, P.J.B., Rosenblat, J.S., Snowden, C.T., Roper, T.J., Eds.; Elsevier: San Diego, CA, USA; London, UK, 1999; Volume 28, pp. 247–332. ISBN 9780120045280. [Google Scholar]
- Janz, J.A.M.; Morel, P.C.H.; Wilkinson, B.H.P.; Purchas, R.W. Preliminary investigation of the effects of low-level dietary inclusion of fragrant essential oils and oleoresins on pig performance and pork quality. Meat Sci. 2007, 75, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Jugl-Chizzola, M.; Ungerhofer, E.; Gabler, C.; Hagmüller, W.; Chizzola, R.; Zitterl-Eglseer, K.; Franz, C. Testing of the palatability of Thymus vulgaris L. and Origanum vulgare L. as flavouring feed addititve for weaner pigs on the basis of a choice experiment. Berl. Munch. Tierarztl. Wochenschr. 2006, 119, 238–243. [Google Scholar] [PubMed]
- Oostindjer, M.; Bolhuis, J.E.; Simon, K.; van den Brand, H.; Kemp, B. Perinatal flavour learning and adaptation to being weaned: All the pig needs is smell. PLoS ONE 2011, 6, e25318. [Google Scholar] [CrossRef] [PubMed]
- Solà-Oriol, D.; Roura, E.; Torrallardona, D. Feed preference in pigs: Effect of selected protein, fat, and fiber sources at different inclusion rates. J. Anim. Sci. 2011, 89, 3219–3227. [Google Scholar] [CrossRef] [PubMed]
- Emami, N.K.; Samie, A.; Rahmani, H.R.; Ruiz-Feria, C.A. The effect of peppermint essential oil and fructooligosaccharides, as alternatives to virginiamycin, on growth performance, digestibility, gut morphology and immune response of male broilers. Anim. Feed Sci. Technol. 2012, 175, 57–64. [Google Scholar] [CrossRef]
- Ahmed, S.T.; Hossain, M.E.; Kim, G.M.; Hwang, J.A.; Ji, H.; Yang, C.J. Effects of resveratrol and essential oils on growth performance, immunity, digestibility and fecal microbial shedding in challenged piglets. Asian-Aust. J. Anim. Sci. 2013, 26, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Platel, K.; Srinivasan, K. Stimulatory influence of select spices on bile secretion in rats. Nutr. Res. 2000, 20, 1493–1503. [Google Scholar] [CrossRef]
- Platel, K.; Srinivasan, K. Studies on the influence of dietary spices on food transit time in experimental rats. Nutr. Res. 2001, 21, 1309–1314. [Google Scholar] [CrossRef]
- Muhl, A.; Liebert, F. No impact of a phytogenic feed additive on digestion and unspecific immune reaction in piglets. J. Anim. Physiol. Anim. Nutr. 2007, 91, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.C.; Steiner, T.; Aufy, A.; Lien, T.F. Effects of supplemental essential oil on growth performance, lipid metabolites and immunity, intestinal characteristics, microbiota and carcass traits in broilers. Livest. Sci. 2012, 144, 253–262. [Google Scholar] [CrossRef]
- Namkung, H.; Li, M.; Gong, J.; Yu, H.; Cottrill, M.; Lange, C.F.M. Impact of feeding blends of organic acids and herbal extracts on growth performance, gut microbiota and digestive function in newly weaned pigs. Can. J. Anim. Sci. 2004, 84, 697–704. [Google Scholar] [CrossRef]
- Jamroz, D.; Wertelecki, T.; Houszka, M.; Kamel, C. Influence of diet type on the inclusion of plant origin active substances on morphological and histochemical characteristics of the stomach and jejunum walls in chicken. J. Anim. Physiol. Anim. Nutr. 2006, 90, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Mitsch, P.; Zitterl-Eglseer, K.; Kohler, B.; Gabler, C.; Losa, R.; Zimpernik, I. The effect of two different blends of essential oil components on the proliferation of Clostridium perfringens in the intestines of broiler chickens. Poult. Sci. 2004, 83, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.S.; Ko, Y.H.; Kang, S.Y.; Lee, C.Y. Effect of a commercial essential oil on growth performance, digestive enzyme activity and intestinal microflora population in broiler chickens. Anim. Feed Sci. Technol. 2007, 134, 304–315. [Google Scholar] [CrossRef]
- Reisinger, N.; Steiner, T.; Nitsch, S.; Schatzmayr, G.; Applegate, T.J. Effects of a blend of essential oils on broiler performance and intestinal morphology during coccidial vaccine exposure. J. Appl. Poult. Res. 2011, 20, 272–283. [Google Scholar] [CrossRef]
- Kroismayr, A.; Schedle, K.; Sehm, J.; Pfaffl, M.W.; Plitzner, C.; Foissy, H.; Ettle, T.; Mayer, H.; Schreiner, M.; Windisch, W. Effects of antimicrobial feed additives on gut microbiology and blood parameters of weaned piglets. Bodenkultur 2008, 59, 111–120. [Google Scholar]
- Draskovic, V.; Bosnjak-Neumuller, J.; Vasiljevic, M.; Petrujkic, B.; Aleksic, N.; Kukolj, V.; Stanimirovic, Z. Influence of phytogenic feed additive on Lawsoniaintracellularis infection in pigs. Prev. Vet. Med. 2018. [Google Scholar] [CrossRef]
- Li, S.Y.; Ru, Y.J.; Liu, M.; Xu, B.; Péron, A.; Shi, X.G. The effect of essential oils on performance, immunity and gut microbial population in weaner pigs. Livest. Sci. 2012, 145, 119–123. [Google Scholar] [CrossRef]
- Horošová, K.; Bujňáková, D.; Kmeť, V. Effect of oregano essential oil on chicken lactobacilli and E. coli. Folia Microbiol. 2006, 51, 278–280. [Google Scholar] [CrossRef]
- Gopi, M.; Karthik, K.; Manjunathachar, H.V.; Tamilmahan, P.; Kesavan, M.; Dashprakash, M.; Balaraju, B.L.; Purushothaman, M.R. Essential Oils as a Feed Additive in Poultry Nutrition. Adv. Anim. Vet. Sci. 2013, 2, 1–7. [Google Scholar] [CrossRef]
- Çabuk, M.; Bozkurt, M.; Alcicek, A.; Çatli, A.U.; Baser, K.H.C. Effect of a dietary essential oil mixture on performance of laying hens in the summer season. S. Afr. J. Anim. Sci. 2006, 36, 215–221. [Google Scholar]
- Özek, K.; Wellmann, K.T.; Ertekin, B.; Tarım, B. Effects of dietary herbal essential oil mixture and organic acid preparation on laying traits, gastrointestinal tract characteristics, blood parameters and immune response of laying hens in a hot summer season. J. Anim. Feed Sci. 2011, 20, 575–586. [Google Scholar] [CrossRef]
- Bozkurt, M.; Küçükyilmaz, K.; Catli, A.U.; Çınar, M.; Bintaş, E.; Çöven, F. Performance, egg quality, and immune response of laying hens fed diets supplemented with mannan-oligosaccharide or an essential oil mixture under moderate and hot environmental conditions. Poult. Sci. 2012, 91, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, R.; Zarei, M.; Torki, M. Adding medicinal herbs including garlic (Allium sativum) and thyme (Thymus vulgaris) to diet of laying hens and evaluating productive performance and egg quality characteristics. Am. J. Anim. Vet. Sci. 2010, 5, 151–154. [Google Scholar] [CrossRef]
- Abdel-Wareth, A.A.A.; Lohakare, J.D. Effect of dietary supplementation of peppermint on performance, egg quality, and serum metabolic profile of Hy-Line Brown hens during the late laying period. Anim. Feed Sci. Technol. 2014, 197, 114–120. [Google Scholar] [CrossRef]
- Aydin, R.; Karaman, M.; Cicek, T.; Yardibi, H. Black cumin (Nigella sativa L.) supplementation into the diet of the laying hen positively influences egg yield parameters, shell quality, and decreases egg cholesterol. Poult. Sci. 2008, 87, 2590–2595. [Google Scholar] [CrossRef] [PubMed]
- Florou-Paneri, P.; Nikolakakis, I.; Giannenas, I.; Koidis, A.; Botsoglou, E.; Dotas, V.; Mitsopoulos, I. Hen performance and egg quality as affected by dietary oregano essential oil and tocopheryl acetate supplementation. Int. J. Poult. Sci. 2005, 4, 449–454. [Google Scholar]
- Cullen, S.P.; Monahan, F.J.; Callan, J.J.; O’doherty, J.V. The effect of dietary garlic and rosemary on grower-finisher pig performance and sensory characteristics of pork. Ir. J. Agric. Food Res. 2005, 44, 57–67. [Google Scholar]
- Stanaćev, V.; Glamočić, D.; Miloscaron, N.; Puvača, N.; Stanaćev, V.; Plavscaron, N. Effect of garlic (Allium sativum L.) in fattening chicks nutrition. Afr. J. Agric. Res. 2011, 6, 943–948. [Google Scholar] [CrossRef]
- Rossi, R.; Pastorelli, G.; Cannata, S.; Tavaniello, S.; Maiorano, G.; Corino, C. Effect of long term dietary supplementation with plant extract on carcass characteristics meat quality and oxidative stability in pork. Meat Sci. 2013, 95, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Fascina, V.B.; Sartori, J.R.; Gonzales, E.; Carvalho, F.B.; Souza, I.M.G.P.; Polycarpo, G.V.; Stradiotti, A.C.; Pelícia, V.C. Phytogenic additives and organic acids in broiler chicken diets. Rev. Bras. Zootec. 2012, 41, 2189–2197. [Google Scholar] [CrossRef]
- Soltan, M.A.; Shewita, R.S.; El-Katcha, M.I. Effect of dietary anise seeds supplementation on growth performance, immune response, carcass traits and some blood parameters of broiler chickens. Int. J. Poult. Sci. 2008, 7, 1078–1088. [Google Scholar] [CrossRef]
- Lange, B.M.; Croteau, R. Genetic engineering of essential oil production in mint. Curr. Opin. Plant Biol. 1999, 2, 139–144. [Google Scholar] [CrossRef]
- Turek, C.; Stintzing, F.C. Stability of essential oils: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 40–53. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; McGinn, S.M. Effects of various feed additives on the methane emissions from beef cattle. Int. Congr. Ser. 2006, 1293, 152–155. [Google Scholar] [CrossRef]
- Śliwiński, B.J.; Soliva, C.R.; Machmüller, A.; Kreuzer, M. Efficacy of plant extracts rich in secondary constituents to modify rumen fermentation. Anim. Feed Sci. Technol. 2002, 101, 101–114. [Google Scholar] [CrossRef]
- Gallardo, G.; Guida, L.; Martinez, V.; López, C.M.; Bernhardt, D.; Blasco, R.; Pedroza-Islas, R.; Hermid, L.G. Microencapsulation of linseed oil by spray drying for functional food application. Food Res. Int. 2013, 52, 473–482. [Google Scholar] [CrossRef]
- Xiao, Z.; Liu, W.; Zhu, G.; Zhou, R.; Niu, Y. A review of the preparation and application of flavour and essential oils microcapsules based on complex coacervation technology. J. Sci. Food Agric. 2014, 94, 1482–1494. [Google Scholar] [CrossRef] [PubMed]
- Martins, I.M.; Barreiro, M.F.; Coelho, M.; Rodrigues, A.E. Microencapsulation of essential oils with biodegradable polymeric carriers for cosmetic applications. Chem. Eng. J. 2014, 245, 191–200. [Google Scholar] [CrossRef]
- Walia, K.; Arguello, H.; Lynch, H.; Leonard, F.C.; Grant, J.; Yearsley, D.; Kelly, S.; Duffy, G.; Gardiner, G.E.; Lawlor, P.G. Effect of strategic administration of an encapsulated blend of formic acid, citric acid, and essential oils on Salmonella carriage, seroprevalence, and growth of finishing pigs. Prev. Vet. Med. 2017, 137, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Zhao, M.; McClements, D.J. Improving the stability of wheat protein-stabilized emulsions: Effect of pectin and xanthan gum addition. Food Hydrocoll. 2015, 43, 377–387. [Google Scholar] [CrossRef]
- Hebishy, E.; Buffa, M.; Juan, B.; Blasco-Moreno, A.; Trujillo, A.J. Ultra high-pressure homogenized emulsions stabilized by sodium caseinate: Effects of protein concentration and pressure on emulsions structure and stability. LWTFood Sci. Technol. 2017, 76, 57–66. [Google Scholar] [CrossRef]
- Zhang, Y.; Gong, J.; Yu, H.; Guo, Q.; Defelice, C.; Hernandez, M.; Yin, Y.; Wang, Q. Alginate-whey protein dry powder optimized for target delivery of essential oils to the intestine of chickens. Poult. Sci. 2014, 93, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, R.; Zou, L.; McClements, D.J. Protein encapsulation in alginate hydrogel beads: Effect of pH on microgel stability, protein retention and protein release. Food Hydrocoll. 2016, 58, 308–315. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Q.C.; Yu, H.; Zhu, J.; de Lange, K.; Yin, Y.; Wang, Q.; Gong, J. Evaluation of alginate–whey protein microcapsules for intestinal delivery of lipophilic compounds in pigs. J. Sci. Food Agric. 2016, 96, 2674–2681. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.S. Preparation of Ca-alginate beads containing high oil content: Influence of process variables on encapsulation efficiency and bead properties. Carbohydr. Polym. 2011, 84, 1267–1275. [Google Scholar] [CrossRef]
- George, M.; Abraham, T.E. Polyionic hydrocolloids for the intestinal delivery of protein drugs: Alginate and chitosan—A review. J. Control. Release 2006, 114, 1–14. [Google Scholar] [CrossRef] [PubMed]
- De Oliveria, E.F.; Paula, H.C.B.; de Paula, R.C.M. Alginate cashew gum nanoparticles for essential oil encapsulation. Colloids Surf. B Biointerfaces 2014, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, F.; Wang, Y.; Zhang, W.; Peng, X.; Pepe, F. Silica modified calcium alginate–xanthan gum hybrid bead composites for the removal and recovery of Pb(II) from aqueous solution. Chem. Eng. J. 2013, 234, 33–42. [Google Scholar] [CrossRef]
- Wang, W.; Waterhouse, G.I.N.; Sun-Waterhouse, D. Co-extrusion encapsulation of canola oil with alginate: Effect of quercetin addition to oil core and pectin addition to alginate shell on oil stability. Food Res. Int. 2013, 54, 837–851. [Google Scholar] [CrossRef]
- Lam, R.S.H.; Nickerson, M.T. Food proteins: A review on their emulsifying properties using a structure–function approach. Food Chem. 2013, 141, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Torcello-Gomez, A.; Maldonado-Valderrama, J.; Martın-Rodrıguez, A.; McClements, D.J. Physicochemical properties and digestibility of emulsified lipids in simulated intestinal fluids: Influence of interfacial characteristics. Soft Matter 2011, 7, 6167–6177. [Google Scholar] [CrossRef]
- Xu, X.; Luo, L.; Liu, C.; McClements, D.J. Utilization of anionic polysaccharides to improve the stability of rice glutelin emulsions: Impact of polysaccharide type, pH, salt, and temperature. Food Hydrocoll. 2017, 64, 112–122. [Google Scholar] [CrossRef]
- Bugarski, B.; Obradovic, B.; Nedovic, V.; Goosen, M.F.A. Electrostatic droplet generation technique for cell immobilization. In Finely Dispersed Systems; Shu, J.P., Spasic, A., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 869–886. [Google Scholar]
- Manojlovic, V.; Rajic, N.; Djonlagic, J.; Obradovic, B.; Nedovic, V.; Bugarski, B. Application of Electrostatic Extrusion—Flavour Encapsulation and Controlled Release. Sensors 2008, 8, 1488–1496. [Google Scholar] [CrossRef] [PubMed]
- Yara-Varon, E.; Li, Y.; Balcells, M.; Canela-Garayoa, R.; Fabiano-Tixier, A.S.; Chemat, F. Vegetable Oils as Alternative Solvents for Green Oleo-Extraction, Purification and Formulation of Food and Natural Products. Molecules 2017, 22, 1474. [Google Scholar] [CrossRef] [PubMed]
- Majeed, H.; Antoniou, J.; Hategekimana, J.; Sharif, H.R.; Haider, J.; Liu, F.; Ali, B.; Rong, L.; Ma, J.; Zhong, F. Influence of carrier oil type, particle size on in vitro lipid digestion and eugenol release in emulsion and nanoemulsions. Food Hydrocol. 2015, 52, 415–422. [Google Scholar] [CrossRef]
- Nazzaro, F.; Orlando, P.; Fratianni, F.; Coppola, R. Microencapsulation in food science and biotechnology. Curr. Opin. Biotechnol. 2012, 23, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, P.T.; Fries, L.L.M.; de Menezes, C.R.; Holkem, A.T.; Schwan, C.L.; Wigmann, E.F.; de Oliveria Bastos, J.; da Silva, C.D.B. Microencapsulation: Concepts, mechanisms, methods and some applications in food technology. Ciênc. Rural 2014, 44, 1304–1311. [Google Scholar] [CrossRef]
- Bilia, A.R.; Guccion, C.; Isacchi, B.; Righasch, C.; Firenzuoli, F.; Berqonizi, M.C. Essential Oils Loaded in Nanosystems: A Developing Strategy for a Successful Therapeutic Approach. Evid. Based Complement. Altern. Med. 2014, 2014, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Akbarzadeh, A.; Rezaei-Sadabady, R.; Davaran, S.; Joo, S.W.; Zarghami, N.; Hanifehpour, Y.; Samiei, M.; Kouhi, M.; Nejati-Koshki, K. Liposome: Classification, preparation, and applications. Nanoscale Res. Lett. 2013, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Bento, M.H.L.; Ouwehand, A.C.; Tiihonen, K.; Lahtinen, S.; Nurminen, P.; Saarinen, M.T.; Schulze, H.; Mygind, T.; Fischer, J. Essential oils and their use in animal feeds for monogastric animals—Effects on feed quality, gut microbiota, growth performance and food safety: A review. Vet. Med. 2013, 58, 449–458. [Google Scholar] [CrossRef]
- MohammadiGheisar, M.; Hosseindoust, A.; Kim, I.H. Evaluating the effect of microencapsulated blends of organic acids and essential oils in broiler chickens diet. J. Appl. Poult. Res. 2015, 24, 511–519. [Google Scholar]
- Palou, R.; Bhandari, B.; Castro, C.; Roura, E. In vivo digestion of encapsulated essential oils in weaned pigs. Anim. Prod. Sci. 2017, 57, 2434. [Google Scholar] [CrossRef]
- Azevedo, I.L.; Martins, E.R.; Almeida, A.C.D.; Nogueira, W.C.L.; FariaFilho, D.E.D.; Santos, V.K.F.D.R.; Lara, L.J.C. Use of Lippiarotundifolia and Cymbopogon flexuosus essential oils, individually or in combination, in broiler diets. RevistaBrasileira de Zootecnia 2017, 46, 13–19. [Google Scholar] [CrossRef]
- Cairo, P.L.G.; Gois, F.D.; Sbardella, M.; Silveira, H.; de Oliveira, R.M.; Allaman, I.B.; Cantarelli, V.S.; Costa, L.B. Effects of dietary supplementation of red pepper (Schinus terebinthifolius Raddi) essential oil on performance, small intestinal morphology and microbial counts of weanling pigs. J. Sci. Food Agric. 2018, 98, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Gois, F.D.; Cairo, P.L.G.; de Souza Cantarelli, V.; do BomfimCosta, L.C.; Fontana, R.; Allaman, I.B.; Sbardellac, M.; de Carvalho Júnior, F.M.; Costa, L.B. Effect of Brazilian red pepper (Schinus terebinthifolius Raddi) essential oil on performance, diarrhea and gut health of weanling pigs. Livest. Sci. 2016, 183, 24–27. [Google Scholar] [CrossRef]
- Méndez Zamora, G.; Durán Meléndez, L.A.; Hume, M.E.; Silva Vázquez, R. Performance, blood parameters, and carcass yield of broiler chickens supplemented with Mexican oregano oil. Rev. Bras. Zootecn. 2017, 46, 515–520. [Google Scholar] [CrossRef][Green Version]


{kind=link}
| Feed Additive | Dose | Major Components | Animal Product | Treatment Effects (Difference with Untreated Group) | Reference |
|---|---|---|---|---|---|
| Egg Production | |||||
| EO blend: oregano, laurel leaf oil, sage leaf oil, myrtle leaf oil, fennel seed oil, and citrus peel oil | 24 mg essential oils/kg complete feed | Not specified | Consumable eggs (Nick-Brown hens) | Increased egg production and decrease incidence of broken-cracked eggs | [100] |
| EO blend: oregano, laurel leaf oil, sage leaf oil, myrtle leaf oil, fennel seed oil, and citrus peel oil | 36 mg essential oils/kg complete feed | carvacrol, thymol, 1:8-cineole, p-cymene and limonene | Consumable eggs | No effects in hen-day egg production, egg weight, egg mass, feed intake, feed conversion ratio, livability, liveweight gain (LWG) and cracked-broken egg ratio and tendency to increase egg weight Increased albumen height and Haugh unit | [101] |
| EO blend (24 mg/kg): oregano, laurel leaf oil, sage leaf oil, myrtle leaf oil, fennel seed oil, and citrus peel oil | 24 mg essential oils/kg complete feed | carvacrol, thymol, 1:8-cineole, p-cymene, and limonene | Consumable eggs (Lohmann LSL-classic) | No effects on egg production, egg weight, egg mass, feed consumption, feed conversion ratio (g feed/g egg), and shell-less egg Increased eggshell weight | [102] |
| Powder mixture of garlic and thyme | 0.1% | Not specified | Consumable eggs | improved means of egg weight | [103] |
| 0.2% | increased egg yolk color as well as blood lymphocyte counts and decreased egg shell weigh | ||||
| Dried peppermint leaves | 0.5, 10, 15, 20 g/kg | Not specified | Consumable eggs (Hy-Line Brown) | Improved egg shell percentage, eggshell thickness and Haugh units. No effects on albumen and yolk percentages and albumen height. Decreased serum cholesterol significantly decreased | [104] |
| Black cumin seeds | 1, 2, 3% | Not specified | Consumable eggs (Hyline-5 White) | Yolk weights of the eggs from hens fed diets containing 1, 2, and 3% black cumin were higher, 2 and 3% increased egg weight and shell thickness and 3% increased egg production and shell strength. | [105] |
| EO Origanum vulgare subsp. hirtum (5%) | 100 mg/kg and 200 mg/kg | Not specified | Consumable eggs (Lohmann laying hens) | No effects on egg production, feed consumption, feed conversion ratio, egg weight and shape, yolk diameter, height and color, Haugh units, and shell thickness | [99] |
| Meat Quality and Production | |||||
| Freeze-dried rosemary at 1 g/kg | 1 g/kg | Not specified | Pork meat (Large White × Landrace) | No effects on sensory properties or carcass characteristics | [106] |
| 10 g/kg | |||||
| Dried garlic | 1 g/kg | Slightly better carcass feed conversion ratio (kg/kg) | |||
| 10 g/kg | Better sensory properties of cooked pork | ||||
| Garlic | 2% | Not specified | Poultry meat (Hubbard hybrid broilers) | No effect on carcass quality parameters | [107] |
| water-soluble extract of Verbenaceae (Lippia spp.) leaves | 5 mg verbascoside/kg feed | Verbascoside | Pork meat Dalland pigs | No effects on carcass characteristics, Longissimus dorsi (LD) meat quality parameters and collagen characteristics Reduced fat odor and rancid flavor intensity in cooked LD muscle stored | [108] |
| Turmeric extracts, citrus extract, grape seed extract Chinese cinnamon essential oil, Chile Boldo leaves and fenugreek seeds | As substitution for inert material | Not specified | Poultry meat | Higher carcass yield, no effects on abdominal fat | [109] |
| Anise seed supplementation | 0.25, 0.5, 0.75, 1.0, 1.25, 1.5 g/kg | Not specified | Poultry meat (Hubbard broilers) | No effects on carcass dressing | [110] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stevanović, Z.D.; Bošnjak-Neumüller, J.; Pajić-Lijaković, I.; Raj, J.; Vasiljević, M. Essential Oils as Feed Additives—Future Perspectives. Molecules 2018, 23, 1717. https://doi.org/10.3390/molecules23071717
Stevanović ZD, Bošnjak-Neumüller J, Pajić-Lijaković I, Raj J, Vasiljević M. Essential Oils as Feed Additives—Future Perspectives. Molecules. 2018; 23(7):1717. https://doi.org/10.3390/molecules23071717
Chicago/Turabian StyleStevanović, Zora Dajić, Jasna Bošnjak-Neumüller, Ivana Pajić-Lijaković, Jog Raj, and Marko Vasiljević. 2018. "Essential Oils as Feed Additives—Future Perspectives" Molecules 23, no. 7: 1717. https://doi.org/10.3390/molecules23071717
APA StyleStevanović, Z. D., Bošnjak-Neumüller, J., Pajić-Lijaković, I., Raj, J., & Vasiljević, M. (2018). Essential Oils as Feed Additives—Future Perspectives. Molecules, 23(7), 1717. https://doi.org/10.3390/molecules23071717