Identification of Nutritional Components in Black Sesame Determined by Widely Targeted Metabolomics and Traditional Chinese Medicines


 ,
, 

Abstract

1. Introduction
2. Results and Discussion
2.1. Widely Targeted Metabolic Profiling of Sesame Seeds Based on LC-MS/MS
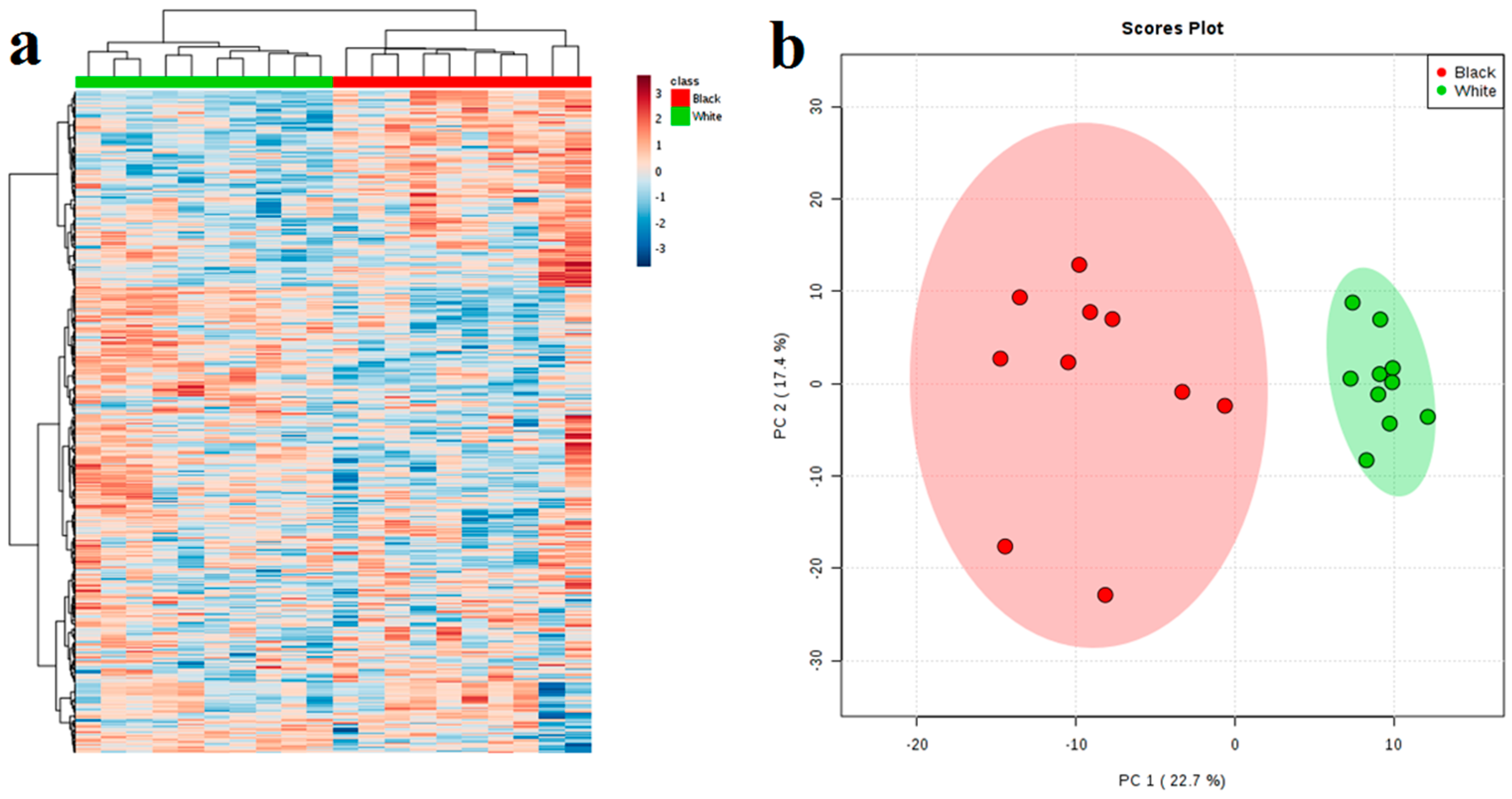
2.2. Multivariate and Cluster Analysis of Black and White Sesame Seeds
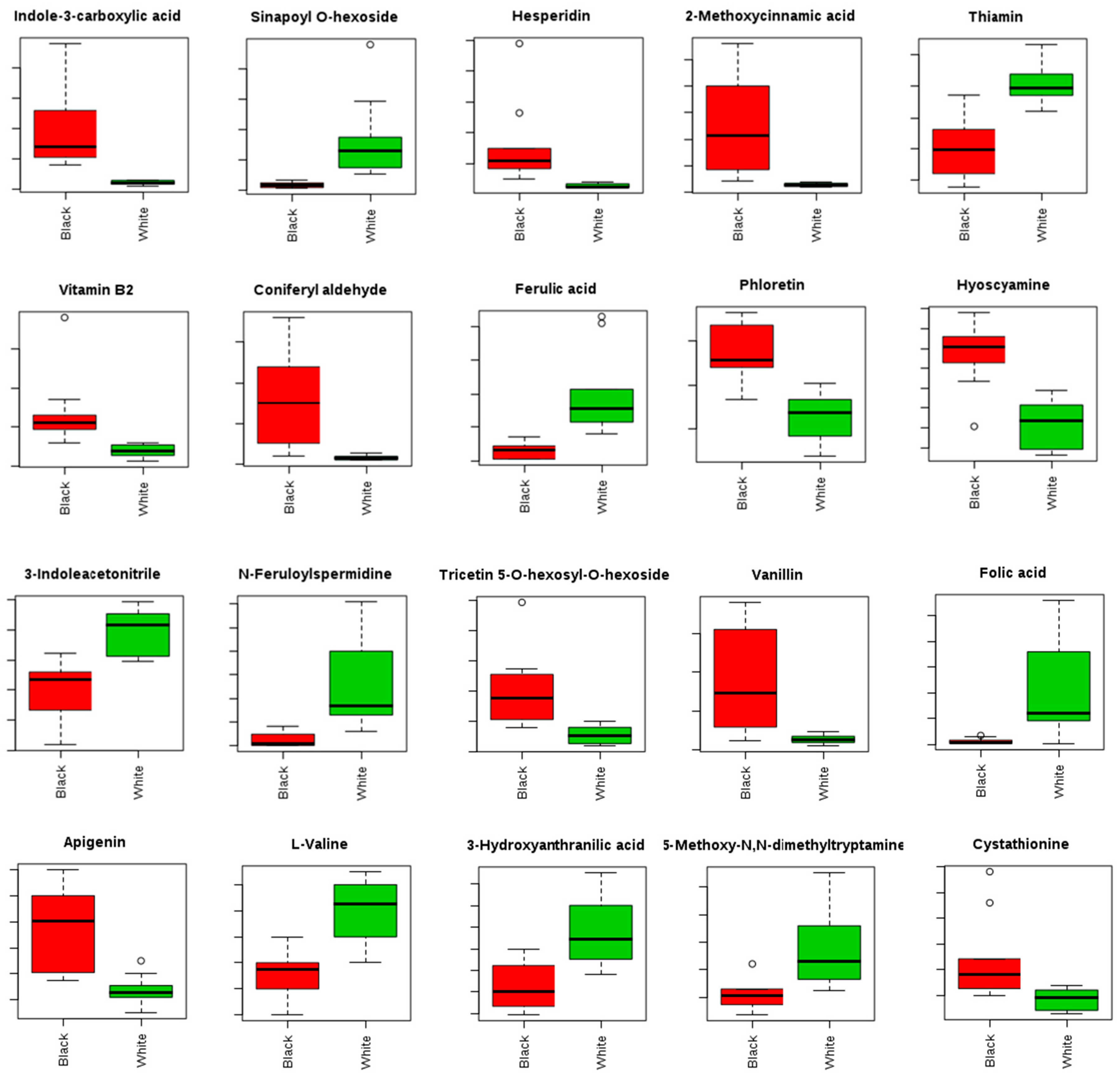
2.3. Identification of Biomarkers of Black Sesame Seeds
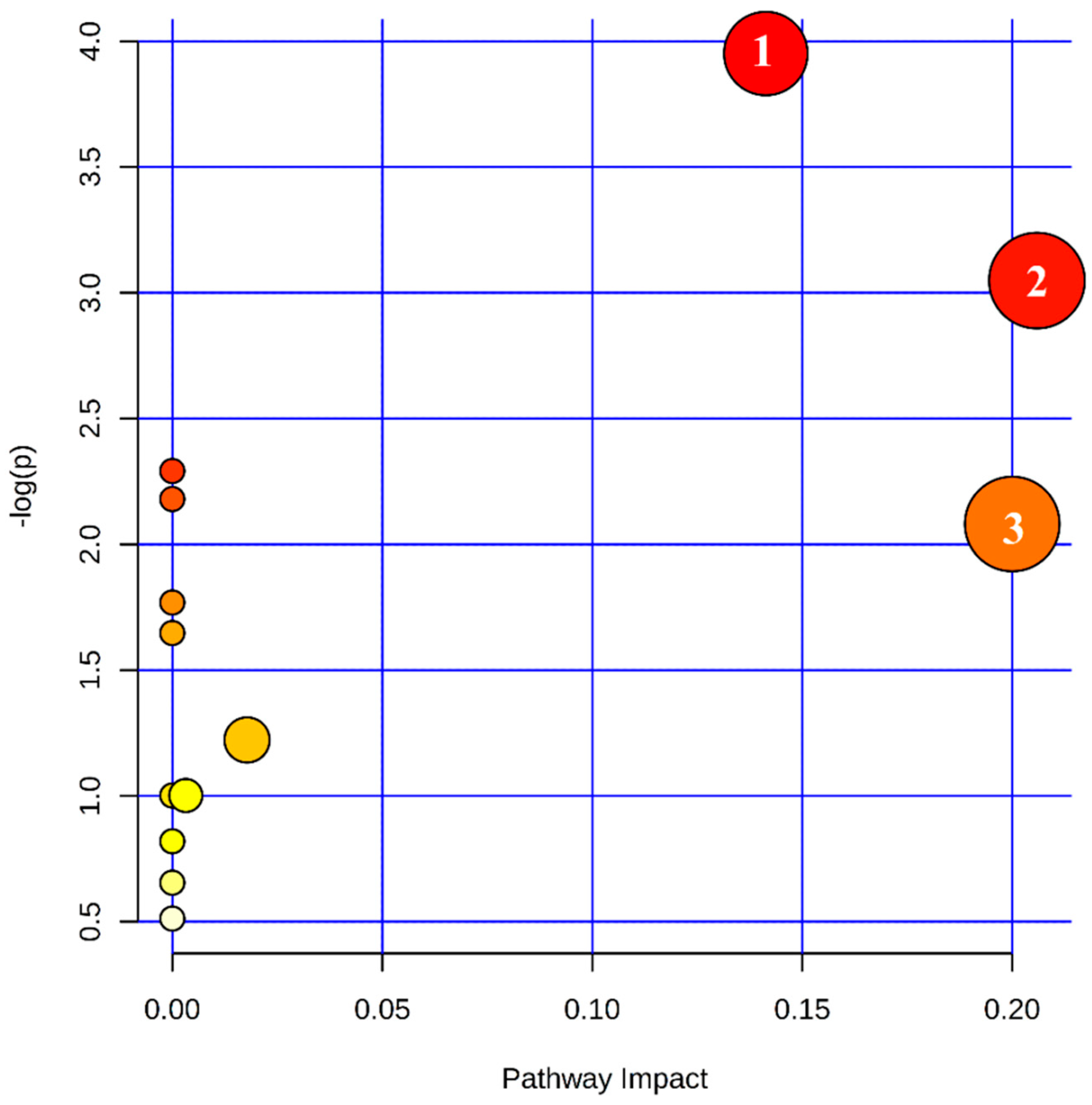
2.4. Metabolic Pathway Analysis of Differential Metabolites
2.5. Nutritional Components Identified in Black Sesame Seeds
3. Conclusion
4. Methods
4.1. Analysis of Sesame Metabolomics Based on LC-MS Data
4.1.1. Sample Preparation
4.1.2. Liquid Chromatographic Mass Spectrometry
4.2. Qualitative and Quantitative Analysis of Metabolites
4.3. Analysis of Metabolite Differences and Metabolic Pathways
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bedigian, D.; Harlan, J.R. Evidence for cultivation of sesame in the ancient world. Econ. Bot. 1986, 40, 137–154. [Google Scholar] [CrossRef]
- Pham, T.D.; Bui, T.M.; Werlemark, G.; Bui, T.C.; Merker, A.; Carlsson, A.S. A study of genetic diversity of sesame (Sesamum indicum L.) in Vietnam and Cambodia estimated by RAPD markers. Genet. Resour. Crop Evol. 2009, 56, 679–690. [Google Scholar] [CrossRef]
- Fan, F.F.; Li, N.W.; Wang, J.; Liu, X.D.; Liu, J.F.; Zhu, Y.G.; Li, S.Q. Molecular marker-directed development of a novel cytoplasmic male sterile line in rice. Mol. Breeding 2015, 35, 212. [Google Scholar] [CrossRef]
- Wei, X.; Zhu, X.D.; Yu, J.Y.; Wang, L.H.; Zhang, Y.X.; Li, D.H.; Zhou, R.; Zhang, X.R. Identification of sesame genomic variations from genome comparison of landrace and variety. Front. Plant Sci. 2016, 7, 1169. [Google Scholar] [CrossRef] [PubMed]
- Laurentin, H.E.; Karlovsky, P. AFLP fingerprinting of sesame (Sesamum indicum L.) cultivars: Identification, genetic relationship and comparison of AFLP informativeness parameters. Genet. Resour. Crop Evol. 2007, 54, 1437–1446. [Google Scholar] [CrossRef]
- Yan, R.; Liang, C.Z.; Meng, Z.G.; Malik, W.; Zhu, T.; Zong, X.F.; Guo, S.D.; Zhang, R. Progress in genome sequencing will accelerate molecular breeding in cotton (Gossypium spp.). 3 Biotech 2016, 6, 217. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.P.K.; McKeown, P.C.; Boualem, A.; Ryder, P.; Brychkova, G.; Bendahmane, A.; Sarkar, A.; Chatterjee, M.; Spillane, C. Tilling by Sequencing (TbyS) for targeted genome mutagenesis in crops. Mol. Breeding 2017, 37, 14. [Google Scholar] [CrossRef]
- Suwarno, W.B.; Pixley, K.V.; Palacios-Rojas, N.; Kaeppler, S.M.; Babu, R. Formation of heterotic groups and understanding genetic effects in a provitamin a biofortified maize breeding program. Crop Sci. 2014, 54, 14–24. [Google Scholar] [CrossRef]
- Qu, J.; Liu, S.Y.; Wang, P.W.; Guan, S.Y.; Fan, Y.G.; Yao, D.; Zhang, L.; Dai, J.L. Agrobacterium-mediated transformation of the beta-subunit gene in 7S globulin protein in soybean using RNAi technology. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.C.; Li, S.G.; Wang, Z.L.; Chang, F.G.; Kong, J.J.; Cai, J.Y.; Zhao, T.J. Identification of major quantitative trait loci for seed oil content in soybeans by combining linkage and genome-wide association mapping. Front. Plant Sci. 2017, 8, 1222. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, E.; Matsunaga, H.; Onogi, A.; Kajiya-Kanegae, H.; Minamikawa, M.; Suzuki, A.; Shirasawa, K.; Hirakawa, H.; Nunome, T.; Yamaguchi, H.; et al. A simulation-based breeding design that uses whole-genome prediction in tomato. Sci. Rep. 2016, 6, 19454. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, H.C.; Kashyap, S.P.; Krishna, R.; Sinha, D.P.; Reddy, S.; Malathi, V.G. Marker assisted selection of Ty-2 and Ty-3 carrying tomato lines and their implications in breeding tomato leaf curl disease resistant hybrids. Euphytica 2015, 204, 407–418. [Google Scholar] [CrossRef]
- Lee, K.R.; Kim, E.H.; Roh, K.H.; Kim, J.B.; Kang, H.C.; Go, Y.S.; Suh, M.C.; Kim, H.U. High-oleic oilseed rapes developed with seed-specific suppression of FAD2 gene expression. Appl. Biol. Chem. 2016, 59, 669–676. [Google Scholar] [CrossRef]
- Mondal, N.; Bhat, K.V.; Srivastava, P.S. Variation in fatty acid composition in Indian germplasm of sesame. J. Am. Oil Chem. Soc. 2010, 87, 1263–1269. [Google Scholar] [CrossRef]
- Weckwerth, W. Metabolomics in systems biology. Ann. Rev. Plant. Biol. 2003, 54, 669–689. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L. The nutritional metabolomics crossroads: How to ensure success for dietary biomarkers. Am. J. Clin. Nutr. 2015, 105, 293–294. [Google Scholar] [CrossRef] [PubMed]
- Mallick, S.; Singh, S.K.; Sarkar, C.; Saha, B.; Bhadra, R. Human placental lipid induces melanogenesis by increasing the expression of tyrosinase and its related proteins in vitro. Pigm. Cell Res. 2005, 1, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O.; Kopka, J.; Dörmann, P.; Altmann, T.; Trethewey, R.N.; Willmitzer, L. Metabolite profiling for plant functional genomics. Nat. Biotechnol. 2000, 18, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- German, J.B.; Roberts, M.A.; Watkins, S.M. Personal metabolomics as a next generation nutritional assessment. J. Nutr. 2003, 133, 4260–4266. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y. Application of the concepts of risk assessment to the study of amino acid supplements. J. Nutr. 2003, 133, 2021S–2024S. [Google Scholar] [CrossRef] [PubMed]
- Kanu, P.J.; Zhu, K.R.; Kanu, J.B.; Zhou, H.M.; Qian, H.F.; Zhu, K.X. Biologically active components and nutraceuticals in sesame and related products: A review and prospect. Trends Food Sci. Technol. 2007, 18, 599–608. [Google Scholar] [CrossRef]
- Bose, U.; Hewavitharana, A.K.; Ng, Y.K.; Shaw, P.N.; Fuerst, J.A.; Hodson, M.P. LC-MS-Based metabolomics study of marine bacterial secondary metabolite and antibiotic production in Salinispora arenicola. Mar. Drugs 2015, 3, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.S.; Zhang, H.Y.; Liu, X.Q.; Yu, S.B.; Xiong, L.Z.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the Study of Rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Akiyama, K.; Sakata, A.; Kuwahara, A.; Otsuki, H.; Sakurai, T.; Saito, K.; Hirai, M.Y. Widely targeted metabolomics based on large-scale MS/MS data for elucidating metabolite accumulation patterns in plants. Plant Cell Physiol. 2009, 50, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Liu, K.Y.; Zhang, Y.X.; Feng, Q.; Wang, L.H.; Zhao, Y.; Li, D.H.; Zhao, Q.; Zhu, X.D.; Zhu, X.F.; et al. Genetic discovery for oil production and quality in sesame. Nat. Commun. 2015, 6, 8609. [Google Scholar] [CrossRef] [PubMed]
- Takayama, S.; Iwasaki, K. Systematic review of traditional Chinese medicine for geriatrics. Geriatr. Gerontol. Int. 2017, 17, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Normile, D. Asian medicine: The new face of traditional Chinese medicine. Science 2003, 299, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.J.; Lee, M.H.; Seo, W.D.; Baek, I.Y.; Kang, J.E.; Lee, J.H. Changes occurring in nutritional components (phytochemicals and free amino acid) of raw and sprouted seeds of white and black sesame (Sesamum indicum L.) and screening of their antioxidant activities. Food Sci. Biotechnol. 2017, 26, 71–78. [Google Scholar] [CrossRef]
- Narasimhulu, C.A.; Selvarajan, K.; Litvinov, D.; Parthasarathy, S. Anti-atherosclerotic and Anti-inflammatory actions of sesame oil. J. Med. Food 2015, 18, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Namiki, M. Nutraceutical Functions of sesame: A review. Crit. Rev. Food Sci. 2007, 47, 651–673. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Mudgal, J.; Parihar, V.K.; Nayak, P.G.; Kutty, N.G.; Rao, C.M. Sesamol treatment reduces plasma cholesterol and triacylglycerol levels in mouse models of acute and chronic hyperlipidemia. Lipids 2013, 48, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, L.D.V.; Cardoso, C.A.; de Oliveira, G.M.M.; Rosa, G.; Moreira, A.S.B. Effects of the intake of sesame seeds (Sesamum indicum L.) and derivatives on oxidative stress: A systematic review. J. Med. Food 2016, 19, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Majdalawieh, A.F.; Massri, M.; Nasrallah, G.K. A comprehensive review on the anti-cancer properties and mechanisms of action of sesamin, a lignan in sesame seeds (Sesamum indicum). Eur. J. Pharmacol. 2017, 815, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.Y.; Cai, F.; Liu, X.F.; Guo, L.L. Sesamin inhibits lipopolysaccharide-induced proliferation and invasion through the p38-MAPK and NF-kappa B signaling pathways in prostate cancer cells. Oncol. Rep. 2015, 33, 3117–3123. [Google Scholar] [CrossRef] [PubMed]
- Kocher, A.; Schiborr, C.; Behnam, D.; Frank, J. The oral bioavailability of curcuminoids in healthy humans is markedly enhanced by micellar solubilisation but not further improved by simultaneous ingestion of sesamin, ferulic acid, naringenin and xanthohumol. J. Funct. Foods 2015, 14, 183–191. [Google Scholar] [CrossRef]
- Park, S.H.; Ryu, S.N.; Bu, Y.; Kim, H.; Simon, J.E.; Kim, K.S. Antioxidant components as potential neuroprotective agents in sesame (Sesamum indicum L.). Food Rev. Int. 2010, 26, 103–121. [Google Scholar] [CrossRef]
- Liu, C.T.; Chien, S.P.; Hsu, D.Z.; Periasamy, S.; Liu, M.Y. Curtive effect of sesame oil in a rat model of chronic kidney disease. Nephrology 2015, 20, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Wanachewin, O.; Boonmaleerat, K.; Pothacharoen, P.; Reutrakul, V.; Kongtawelert, P. Sesamin stimulates osteoblast differentiation through p38 and ERK1/2 MAPK signaling pathways. BMC Complem. Altern. Med. 2012, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Srisuthtayanont, W.; Pruksakorn, D.; Kongtawelert, P.; Pothacharoen, P. Effects of sesamin on chondroitin sulfate proteoglycan synthesis induced by interleukin-1beta in human chondrocytes. BMC Complem. Altern. Med. 2017, 17, 286. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gao, Y.Q.; Xie, W.B.; Gong, L.; Lu, K.; Wang, W.S.; Li, Y.; Liu, X.Q.; Zhang, H.Y.; Dong, H.X.; et al. Genome-wide association analyses provide genetic and biochemical insights into natural variation in rice metabolism. Nat. Genet. 2014, 46, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0-making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed]
- Slominski, R.M.; Zmijewski, M.A.; Slominski, A.T. The role of melanin pigment in melanoma. Exp. Dermatol. 2015, 24, 258–259. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Kita, S.; Tanida, Y.; Taguchi, Y.; Morimoto, S.; Akimoto, K.; Tanaka, T. Antihypertensive effect of sesamin. III. Protection against development and maintenance of hypertension in stroke-prone spontaneously hypertensive rats. Biol. Pharm. Bull. 1998, 21, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Camachohübner, A.; Beermann, F. Cellular and molecular features of mammalian pigmentation-tyrosinase and TRPs. Pathol. Biol. 2000, 6, 577–583. [Google Scholar]
- Wang, L.H.; Xia, Q.J.; Zhang, Y.X.; Zhu, X.D.; Zhu, X.F.; Li, D.H.; Ni, X.M.; Gao, Y.; Xiang, H.T.; Wei, X.; et al. Updated sesame genome assembly and fine mapping of plant height and seed coat color QTLs using a new high-density genetic map. BMC Genomics. 2016, 17, 31. [Google Scholar] [CrossRef] [PubMed]
- Haidari, F.; Mohammad-Shahi, M.; Keshavarz, S.A.; Rashidi, M.R. Effect of orange juice and hesperidin on biomarkers of oxidative stress and liver xanthine oxidoreductase activity in hyperuricemic rats. J. Gastroen. Hepatol. 2012, 27, 91. [Google Scholar]
- Roohbakhsh, A.; Parhiz, H.; Soltani, F.; Rezaee, R.; Iranshahi, M. Molecular mechanisms behind the biological effects of hesperidin and hesperetin for the prevention of cancer and cardiovascular diseases. Life Sci. 2015, 124, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Chou, S.T.; Chang, W.L.; Chang, C.T.; Hsu, S.L.; Lin, Y.C.; Shih, Y. Cinnamomum cassia essential oil inhibits alpha-MSH-induced melanin production and oxidative stress in murine B16 melanoma cells. Int. J. Mol. Sci. 2013, 14, 19186–19201. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.L.; Gooden, D.; Liu, L.H.; Zhao, S.M.; Soderblom, E.J.; Toone, E.J.; Beyer, W.F., Jr.; Walder, H.; Spector, N.L. Photo-activated psoralen binds the ErbB2 catalytic kinase domain, blocking ErbB2 signaling and triggering tumor cell apoptosis. PLoS ONE 2014, 9, e88983. [Google Scholar] [CrossRef]
- Yu, L.T.; Tan, Y.Y.; Zhu, L. Dieta ry vitamin B2 intake and breast cancer risk: A systematic review and meta-analysis. Arch. Gynecol. Obstet. 2017, 295, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.; Kniazeva, M.; Han, M. A vitamin-B2-sensing mechanism that regulates gut protease activity to impact animal’s food behavior and growth. eLife 2017, 6, e26243. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: not available. |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolic Pathway | Total Number of Metabolites | Detected Metabolites |
|---|---|---|
| taurine and hypotaurine metabolism | 5 | 1 |
| alanine, aspartate and glutamate metabolism | 22 | 5 |
| arginine and proline metabolism | 38 | 9 |
| isoquinoline alkaloid biosynthesis | 6 | 2 |
| phenylalanine metabolism | 8 | 2 |
| tyrosine metabolism | 18 | 2 |
| tryptophan metabolism | 27 | 7 |
| sulfur metabolism | 12 | 3 |
| indole alkaloid biosynthesis | 7 | 2 |
| cysteine and methionine metabolism | 34 | 8 |
| glycine, serine and threonine metabolism | 30 | 6 |
| purine metabolism | 61 | 11 |
| riboflavin metabolism | 10 | 1 |
| phenylpropanoid biosynthesis | 45 | 7 |
| methane metabolism | 11 | 1 |
| pantothenate and CoA biosynthesis | 14 | 2 |
| flavonoid biosynthesis | 43 | 2 |
| aminoacyl-tRNA biosynthesis | 67 | 19 |
| pyrimidine metabolism | 38 | 3 |
| lysine degradation | 17 | 2 |
| glutathione metabolism | 26 | 3 |
| lysine biosynthesis | 10 | 3 |
| histidine metabolism | 16 | 2 |
| starch and sucrose metabolism | 30 | 2 |
| glycerophospholipid metabolism | 25 | 3 |
| galactose metabolism | 26 | 1 |
| valine, leucine and isoleucine biosynthesis | 26 | 4 |
| steroid biosynthesis | 36 | 1 |
| porphyrin and chlorophyll metabolism | 29 | 2 |
| selenoamino acid metabolism | 19 | 2 |
| Number | Compound | p | Number | Compound | p |
|---|---|---|---|---|---|
| 1 | indole-3-carboxylic acid | 1.37 × 10−8 | 11 | 3-indoleacetonitrile | 4.66 × 10−5 |
| 2 | 1-O-Sinapoyl-β-d-glucose | 2.73 × 10−7 | 12 | N-feruloylspermidine | 5.28 × 10−5 |
| 3 | hesperidin | 3.50 × 10−6 | 13 | tricetin 5-O-hexosyl-O-hexoside | 8.43 × 10−5 |
| 4 | 2-methoxycinnamic acid | 4.73 × 10−6 | 14 | vanillin | 1.8 × 10−3 |
| 5 | thiamine | 1.35 × 10−5 | 15 | folic acid | 1.9 × 10−3 |
| 6 | vitamin B2 | 1.44 × 10−5 | 16 | apigenin | 2.4 × 10−3 |
| 7 | coniferyl aldehyde | 1.65 × 10−5 | 17 | L-valine | 3.2 × 10−3 |
| 8 | ferulic acid | 2.18 × 10−5 | 18 | 3-hydroxyanthranilic acid | 4.2 × 10−3 |
| 9 | phloretin | 2.54 × 10−5 | 19 | 5-methoxy-N,N-dimethyltryptamine | 7.0 × 10−3 |
| 10 | hyoscyamine | 2.59 × 10−5 | 20 | l-cystathionine | 9.6 × 10−3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Zhang, L.; Huang, X.; Wang, X.; Yang, R.; Mao, J.; Wang, X.; Wang, X.; Zhang, Q.; Li, P. Identification of Nutritional Components in Black Sesame Determined by Widely Targeted Metabolomics and Traditional Chinese Medicines. Molecules 2018, 23, 1180. https://doi.org/10.3390/molecules23051180
Wang D, Zhang L, Huang X, Wang X, Yang R, Mao J, Wang X, Wang X, Zhang Q, Li P. Identification of Nutritional Components in Black Sesame Determined by Widely Targeted Metabolomics and Traditional Chinese Medicines. Molecules. 2018; 23(5):1180. https://doi.org/10.3390/molecules23051180
Chicago/Turabian StyleWang, Dandan, Liangxiao Zhang, Xiaorong Huang, Xiao Wang, Ruinan Yang, Jin Mao, Xuefang Wang, Xiupin Wang, Qi Zhang, and Peiwu Li. 2018. "Identification of Nutritional Components in Black Sesame Determined by Widely Targeted Metabolomics and Traditional Chinese Medicines" Molecules 23, no. 5: 1180. https://doi.org/10.3390/molecules23051180
APA StyleWang, D., Zhang, L., Huang, X., Wang, X., Yang, R., Mao, J., Wang, X., Wang, X., Zhang, Q., & Li, P. (2018). Identification of Nutritional Components in Black Sesame Determined by Widely Targeted Metabolomics and Traditional Chinese Medicines. Molecules, 23(5), 1180. https://doi.org/10.3390/molecules23051180