
Identification by Molecular Docking of Homoisoflavones from Leopoldia comosa as Ligands of Estrogen Receptors

 ,
,  ,
, 


Abstract

1. Introduction
2. Results and Discussion
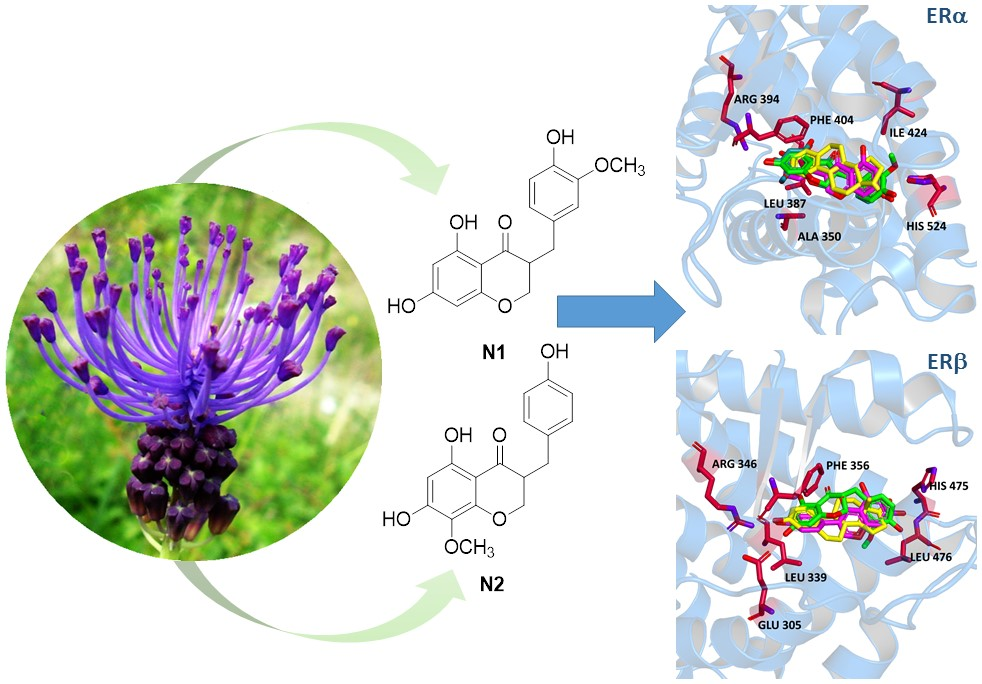
2.1. Homoisoflavones from Leopoldia comosa
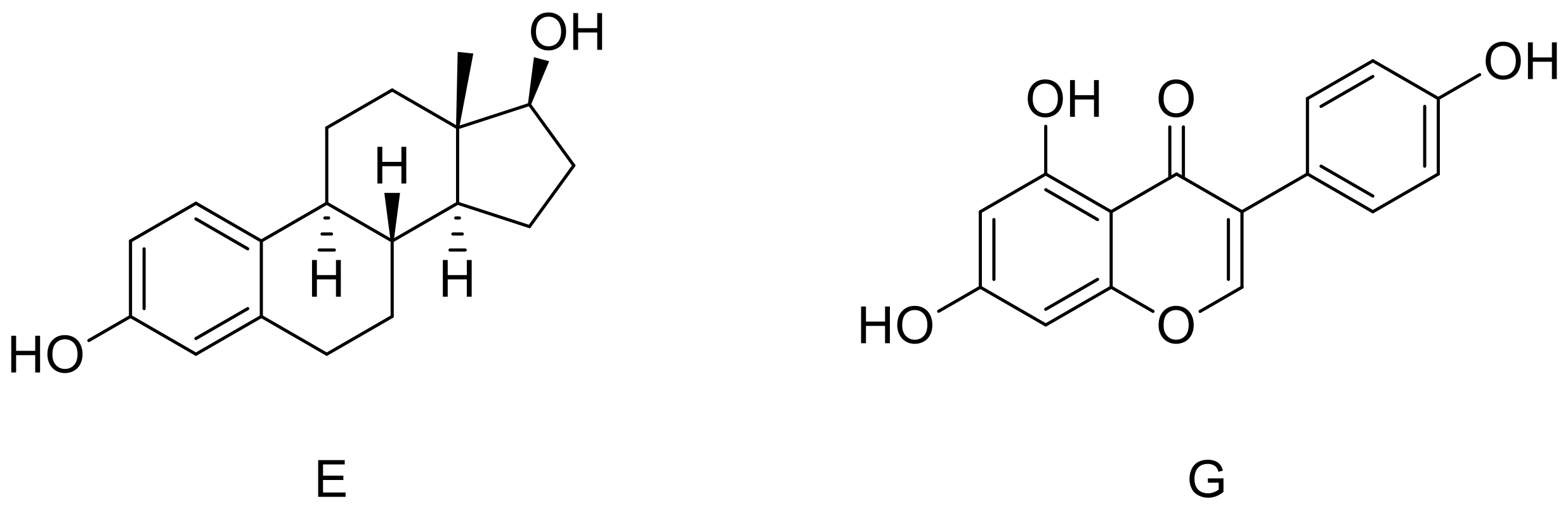
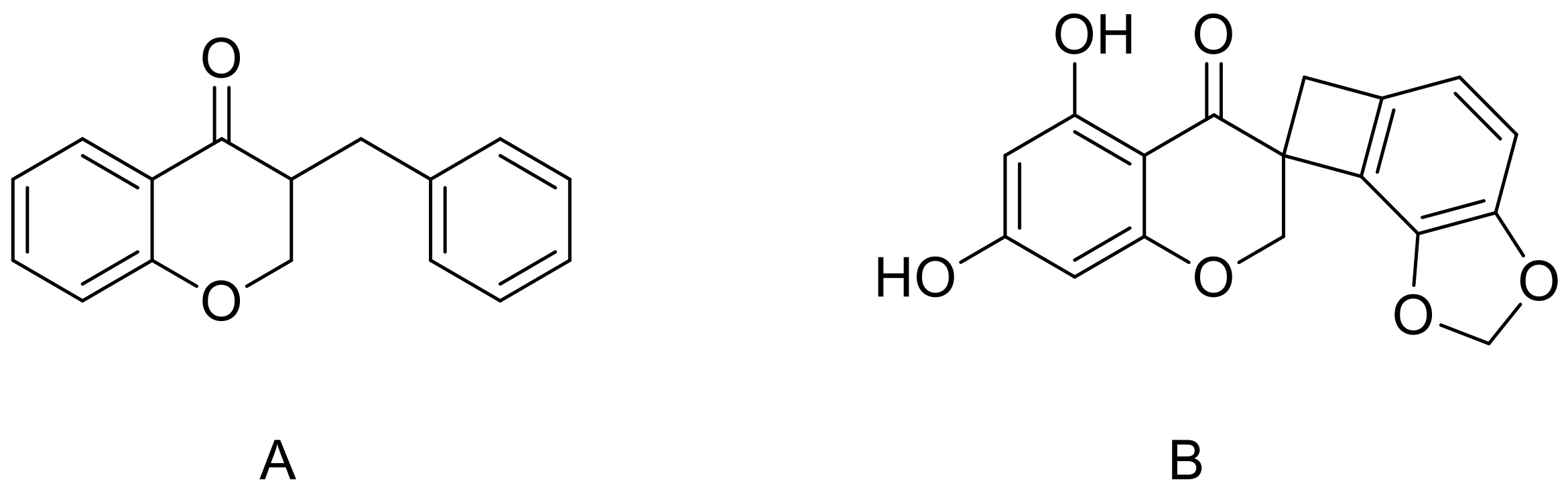
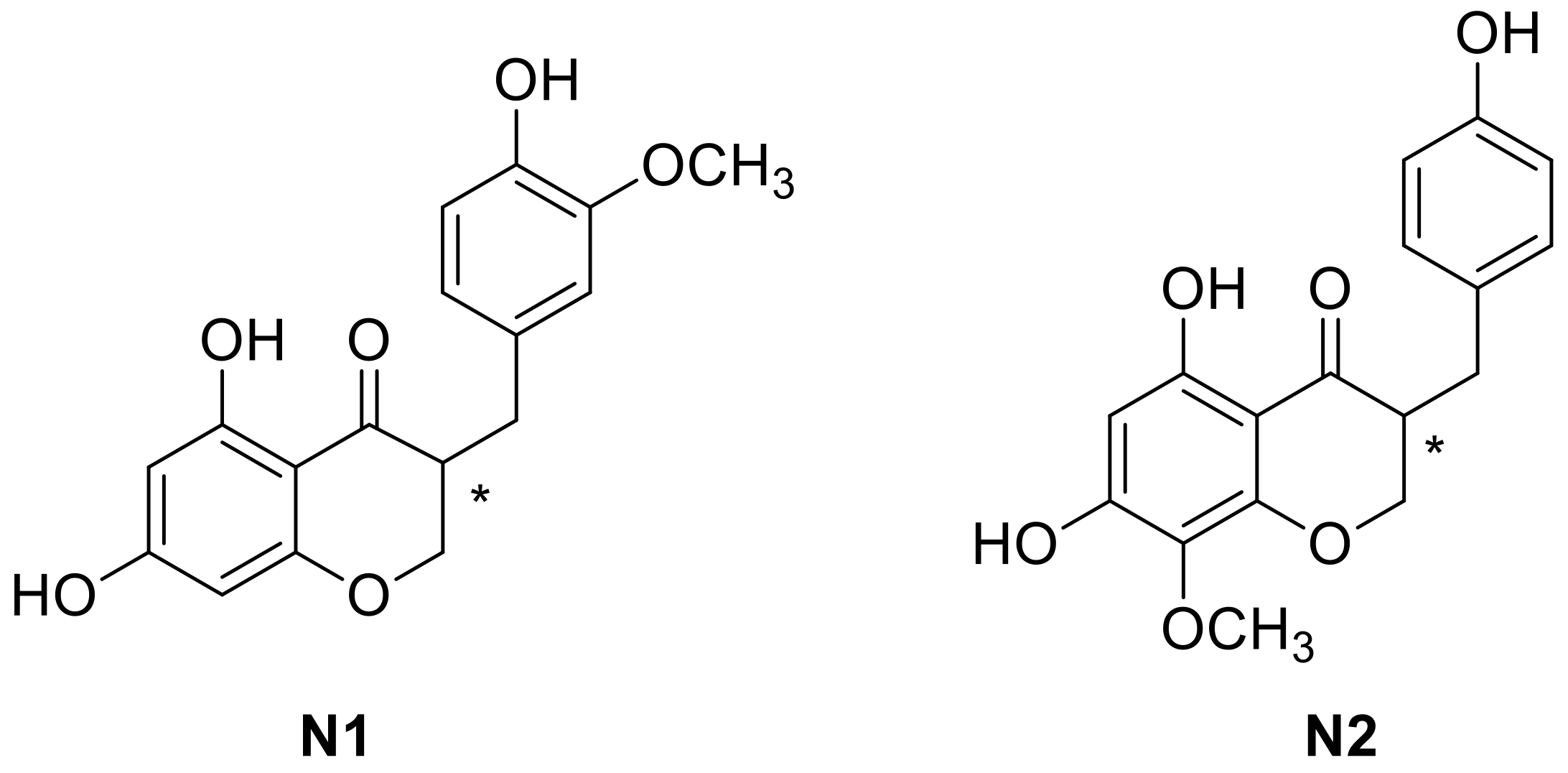
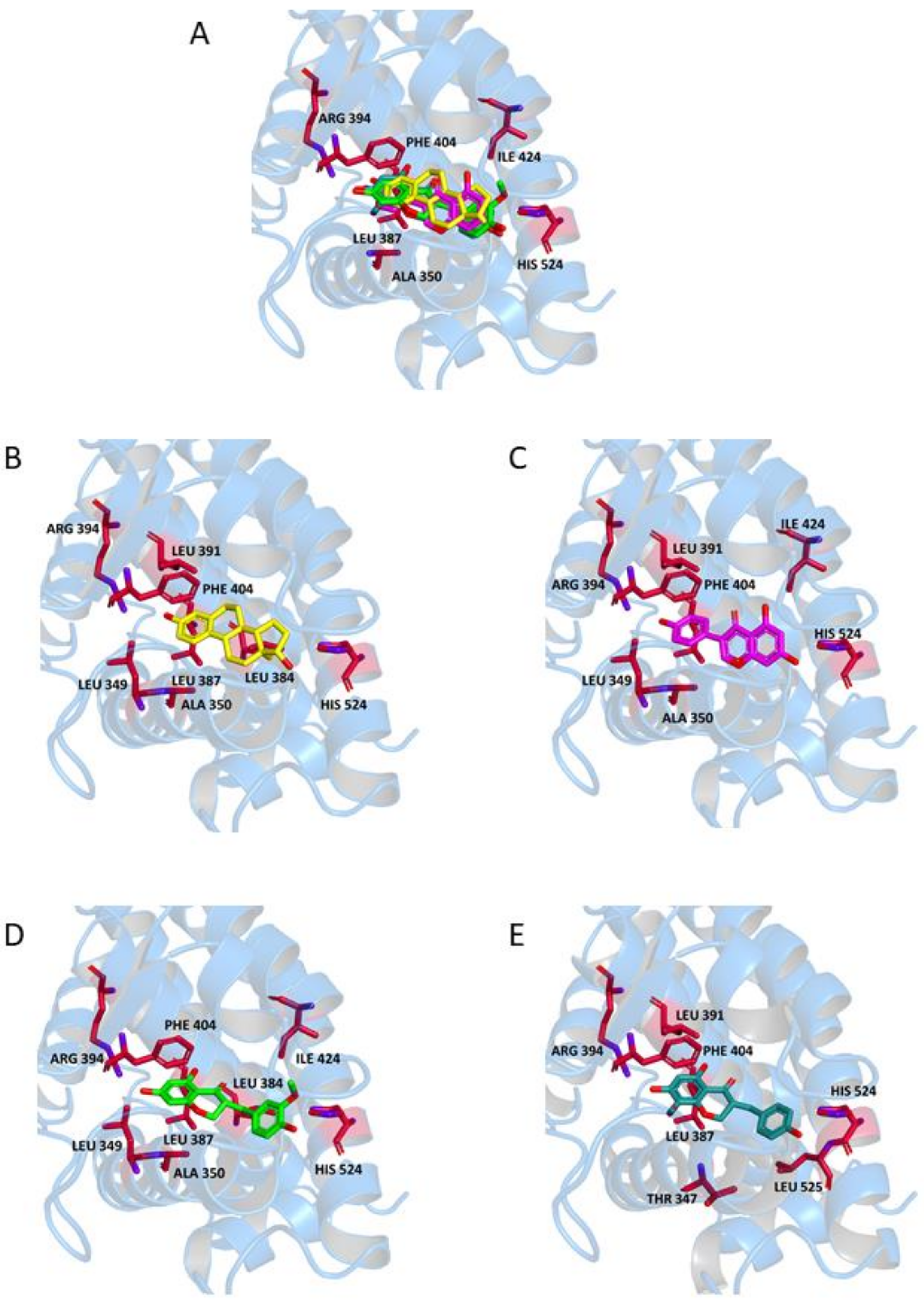
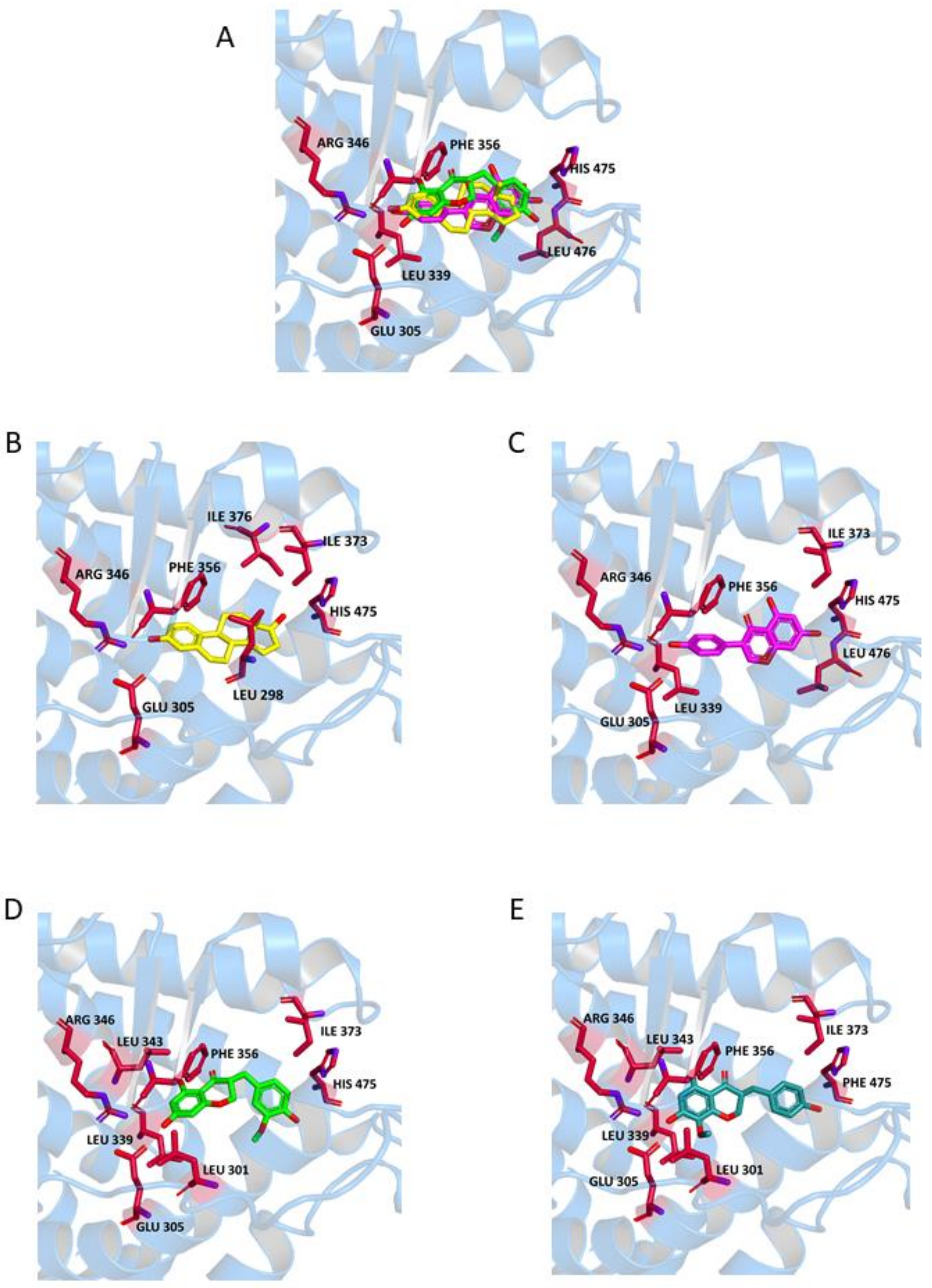
2.2. Binding to Estrogen Receptor (ER) Proteins
3. Materials and Methods
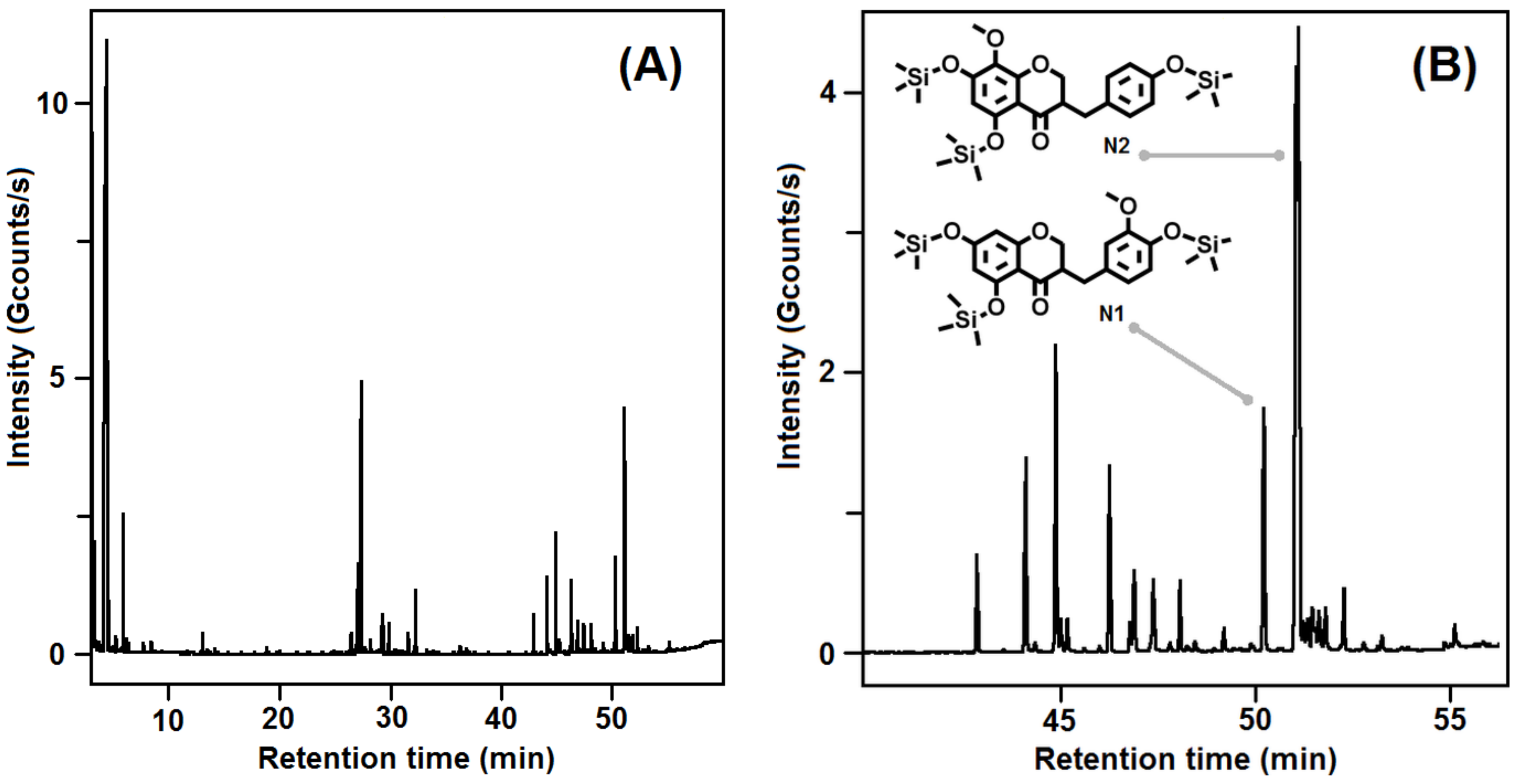
3.1. Plant Material and Phytochemical Profile
3.2. Molecular Docking
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, P.; McInnes, C.; Zhu, B.T. Structural characterization of the binding interactions of various endogenous estrogen metabolites with human estrogen receptor α and β subtypes: A molecular modeling study. PLoS ONE 2013, 8, e74615. [Google Scholar] [CrossRef] [PubMed]
- Sukocheva, O.A. Estrogen, estrogen receptors, and hepatocellular carcinoma: Are we there yet? World J. Gastroenterol. 2018, 24, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Guillaume, M.; Montagner, A.; Fontaine, C.; Lenfant, F.; Arnal, J.F.; Gourdy, P. Nuclear and membrane actions of estrogen receptor alpha: contribution to the regulation of energy and glucose homeostasis. Adv. Exp. Med. Biol. 2017, 1043, 401–426. [Google Scholar] [PubMed]
- Jia, M.; Dahlman-Wright, K.; Gustafsson, J.A. Estrogen receptor alpha and beta in health and disease. Best Pract. Res. Clin. Endoc. Metab. 2015, 29, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, K.; Delaunay, F.; Gustafsson, J.A. Estrogen receptor beta acts as a dominant regulator of estrogen signaling. Oncogene 2000, 19, 4970–4978. [Google Scholar] [CrossRef] [PubMed]
- Paech, K.; Webb, P.; Kuiper, G.G.; Nilsson, S.; Gustafsson, J.; Kushner, P.J.; Scanlan, T.S. Differential ligand activation of estrogen receptors ERalpha and ERbeta at AP1 sites. Science 1997, 277, 1508–1510. [Google Scholar] [CrossRef] [PubMed]
- Cowley, S.M.; Parker, M.G. A comparison of transcriptional activation by ER alpha and ER beta. J. Steroid Biochem. Mol. Biol. 1999, 69, 165–175. [Google Scholar] [CrossRef]
- Chakraborty, S.; Willett, H.; Biswas, P.K. Insight into estrogen receptor beta-beta and alpha-beta homo- and heterodimerization: A combined molecular dynamics and sequence analysis study. Biophys. Chem. 2012, 170, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh, S.; Zarghi, A. Estrogen Receptor Ligands: A Review (2013–2015). Sci. Pharm. 2016, 84, 409–427. [Google Scholar] [CrossRef] [PubMed]
- Paterni, I.; Granchi, C.; Katzenellenbogen, J.A.; Minutolo, F. Estrogen receptors alpha (ERalpha) and beta (ERbeta): Subtype-selective ligands and clinical potential. Steroids 2014, 90, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Zakharov, M.N.; Khan, S.H.; Miki, R.; Jang, H.; Toraldo, G.; Singh, R.; Bhasin, S.; Jasuja, R. The dynamic structure of the estrogen receptor. J. Amino Acids 2011, 2011, 812540. [Google Scholar] [CrossRef] [PubMed]
- Traboulsi, T.; El Ezzy, M.; Gleason, J.L.; Mader, S. Antiestrogens: Structure-activity relationships and use in breast cancer treatment. J. Mol. Endocrinol. 2017, 58, R15–R31. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; Bocedi, A.; Marino, M. Structure-function relationship of estrogen receptor alpha and beta: Impact on human health. Mol. Asp. Med. 2006, 27, 299–402. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Omoto, Y.; Iwase, H.; Yamashita, H.; Toyama, T.; Coombes, R.C.; Filipovic, A.; Warner, M.; Gustafsson, J.A. Differential expression of estrogen receptor α, β1, and β2 in lobular and ductal breast cancer. Proc. Natl. Acad. Sci. USA 2014, 111, 1933–1938. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Lam, E.W.; Sunters, A.; Enmark, E.; de Bella, M.T.; Coombes, R.C.; Gustafsson, J.A.; Dahlman-Wright, K. Expression of estrogen receptor beta isoforms in normal breast epithelial cells and breast cancer: Regulation by methylation. Oncogene 2003, 22, 7600–7606. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kim, H.S.; Song, Y.S. Genistein as a potential anticancer agent against ovarian cancer. J. Tradit. Complement. Med. 2012, 2, 96–104. [Google Scholar] [CrossRef]
- Zhang, L.L.; Li, L.; Wu, D.P.; Fan, J.H.; Li, X.; Wu, K.J.; Wang, X.Y.; He, D.L. A novel anti-cancer effect of genistein: Reversal of epithelial mesenchymal transition in prostate cancer cells. Acta Pharmacol. Sin. 2008, 29, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Peterson, G.; Barnes, S. Genistein inhibition of the growth of human breast cancer cells: independence from estrogen receptors and the multi-drug resistance gene. Biochem. Biophys. Res. Commun. 1991, 179, 661–667. [Google Scholar] [CrossRef]
- Lentini, F.; Venza, F. Wild food plants of popular use in Sicily. J. Ethnobiol. Ethnomed. 2007, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Pieroni, A.; Nebel, S.; Santoro, R.F.; Heinrich, M. Food for two seasons: culinary uses of non-cultivated local vegetables and mushrooms in a south Italian village. Int. J. Food Sci. Nutr. 2005, 56, 245–272. [Google Scholar] [CrossRef] [PubMed]
- Pieroni, A.; Nebel, S.; Quave, C.; Munz, H.; Heinrich, M. Ethnopharmacology of liakra: Traditional weedy vegetables of the Arbereshe of the Vulture area in southern Italy. J. Ethnopharmacol. 2002, 81, 165–185. [Google Scholar] [CrossRef]
- Lin, L.G.; Liu, Q.Y.; Ye, Y. Naturally occurring homoisoflavonoids and their pharmacological activities. Planta Med. 2014, 80, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Casacchia, T.; Scavello, F.; Rocca, C.; Granieri, M.C.; Beretta, G.; Amelio, D.; Gelmini, F.; Spena, A.; Mazza, R.; Toma, C.C.; et al. Leopoldia comosa prevents metabolic disorders in rats with high-fat diet-induced obesity. Eur. J. Nutr. 2018, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nirmal, N.P.; Rajput, M.S.; Prasad, R.G.; Ahmad, M. Brazilin from Caesalpinia sappan heartwood and its pharmacological activities: A review. Asian Pac. J. Trop. Med. 2015, 8, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Tundis, R.; Menichini, F.; Pugliese, A.; Bonesi, M.; Solimene, U.; Menichini, F. Chelating, antioxidant and hypoglycaemic potential of Muscari comosum (L.) Mill. bulb extracts. Int. J. Food Sci. Nutr. 2010, 61, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Harinantenaina, L.; Brodie, P.J.; Goetz, M.; Shen, Y.; TenDyke, K.; Kingston, D.G. Antiproliferative homoisoflavonoids and bufatrienolides from Urginea depressa. J. Nat. Prod. 2013, 76, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, D.A.; Schwikkard, S.L.; Crouch, N.R. The chemistry and biological activity of the Hyacinthaceae. Nat. Prod. Rep. 2013, 30, 1165–1210. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, K.; Elgorashi, E.E.; Malan, S.F.; Drewes, S.E.; van Staden, J.; Crouch, N.R.; Mulholland, D.A. Anti-inflammatory activity and QSAR studies of compounds isolated from Hyacinthaceae species and Tachiadenus longiflorus Griseb. (Gentianaceae). Bioorg. Med. Chem. 2005, 13, 2561–2568. [Google Scholar] [CrossRef] [PubMed]
- Abegaz, B.M.; Mutanyatta-Comar, J.N. Naturally occurring homoisoflavonoids: Phytochemistry, biological activities and synthesis. Nat. Prod. Commun. 2007, 2, 475–498. [Google Scholar]
- Castelli, M.V.; López, S.N. Homoisoflavonoids: Occurrence, biosynthesis, and biological activity. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, Netherlands, 2017; Volume 54, pp. 315–354. [Google Scholar]
- Mann, J.; Davidson, R.S.; Hobbs, J.B.; Banthorpe, D.V.; Harbome, J.B. Natural products: Their chemistry and biological significance. In Longman Scientific and Technical; Harlow, E., Ed.; Longman Group UK Ltd.: Harlow, UK, 1994; p. 372. [Google Scholar]
- De Vincenzi, S.; Lupattelli, M.; Cestola, E.; Liponi, G.B. Effect of variety and agronomical conditions on the level of polyphenols and antinutritional factors of Vicia Faba Minor. Vet. Res. Commun. 2006, 30, 371–374. [Google Scholar] [CrossRef]
- Casacchia, T.; Sofo, A.; Casaburi, I.; Marrelli, M.; Conforti, F.; Statti, G.A. Antioxidant, enzyme-inhibitory and antitumor activity of the wild dietary plant Muscari comosum (L.). Int. J. Plant Biol. 2017, 8, 6895. [Google Scholar] [CrossRef]
- Marrelli, M.; Cristaldi, B.; Menichini, F.; Conforti, F. Inhibitory effects of wild dietary plants on lipid peroxidation and on the proliferation of human cancer cells. Food Chem. Toxicol. 2015, 86, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Archundia, M.; Garcia-Vazquez, J.B.; Colin-Astudillo, B.; Bello, M.; Prestegui-Martel, B.; Chavez-Blanco, A.; Duenas-Gonzalez, A.; Fragoso-Vazquez, M.J.; Mendieta-Wejebe, J.; Abarca-Rojano, E.; et al. Computational study of the binding modes of diverse DPN analogues on estrogen receptors (ER) and the biological evaluation of a new potential antiestrogenic ligand. Anticancer Agents Med. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yugandhar, P.; Kumar, K.K.; Neeraja, P.; Savithramma, N. Isolation, characterization and in silico docking studies of synergistic estrogen receptor a anticancer polyphenols from Syzygium alternifolium (Wt.) Walp. J. Intercult. Ethnopharmacol. 2017, 6, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Muchtaridi, M.; Syahidah, H.N.; Subarnas, A.; Yusuf, M.; Bryant, S.D.; Langer, T. Molecular docking and 3D-pharmacophore modeling to study the interactions of chalcone derivatives with estrogen receptor alpha. Pharmaceuticals 2017, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Manas, E.S.; Unwalla, R.J.; Xu, Z.B.; Malamas, M.S.; Miller, C.P.; Harris, H.A.; Hsiao, C.; Akopian, T.; Hum, W.T.; Malakian, K.; et al. Structure-based design of estrogen receptor-beta selective ligands. J. Am. Chem. Soc. 2004, 126, 15106–15119. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Manas, E.S.; Xu, Z.B.; Unwalla, R.J.; Somers, W.S. Understanding the selectivity of genistein for human estrogen receptor-beta using X-ray crystallography and computational methods. Structure 2004, 12, 2197–2207. [Google Scholar] [CrossRef] [PubMed]
- Koide, A.; Abbatiello, S.; Rothgery, L.; Koide, S. Probing protein conformational changes in living cells by using designer binding proteins: application to the estrogen receptor. Proc. Natl. Acad. Sci. USA 2002, 99, 1253–1258. [Google Scholar] [CrossRef] [PubMed]
- Pike, A.C.; Brzozowski, A.M.; Hubbard, R.E.; Bonn, T.; Thorsell, A.G.; Engstrom, O.; Ljunggren, J.; Gustafsson, J.A.; Carlquist, M. Structure of the ligand-binding domain of oestrogen receptor beta in the presence of a partial agonist and a full antagonist. Embo J. 1999, 18, 4608–4618. [Google Scholar] [CrossRef] [PubMed]
- Nettles, K.W.; Bruning, J.B.; Gil, G.; Nowak, J.; Sharma, S.K.; Hahm, J.B.; Kulp, K.; Hochberg, R.B.; Zhou, H.; Katzenellenbogen, J.A.; et al. NFkB selectivity of estrogen receptor ligands revealed by comparative crystallographic analyses. Nat. Chem. Biol. 2008, 4, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Tanenbaum, D.M.; Wang, Y.; Williams, S.P.; Sigler, P.B. Crystallographic comparison of the estrogen and progesterone receptor's ligand binding domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5998–6003. [Google Scholar] [CrossRef] [PubMed]
- Warnmark, A.; Treuter, E.; Gustafsson, J.A.; Hubbard, R.E.; Brzozowski, A.M.; Pike, A.C. Interaction of transcriptional intermediary factor 2 nuclear receptor box peptides with the coactivator binding site of estrogen receptor alpha. J. Biol. Chem. 2002, 277, 21862–21868. [Google Scholar] [CrossRef] [PubMed]
- Souza, P.C.T.; Textor, L.C.; Melo, D.C.; Nascimento, A.S.; Skaf, M.S.; Polikarpov, I. An alternative conformation of ERbeta bound to estradiol reveals H12 in a stable antagonist position. Sci. Rep. 2017, 7, 3509. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Labbè, C.M.; Pencheva, T.; Jereva, D.; Desvillechabrol, D.; Becot, J.; Villoutreix, B.O.; Pajeva, I.; Miteva, M.A. AMMOS2: A web server for protein-ligand-water complexes refinement via molecular mechanics. Nucleic Acids Res. 2017, 45, W350–W355. [Google Scholar] [CrossRef] [PubMed]
- Summa, C.M.; Levitt, M. Near-native structure refinement using in vacuo energy minimization. Proc. Natl. Acad. Sci. USA 2007, 104, 3177–3182. [Google Scholar] [CrossRef] [PubMed]
- Rizzuti, B.; Daggett, V. Using simulations to provide the framework for experimental protein folding studies. Arch. Biochem. Biophys. 2013, 531, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein-ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef] [PubMed]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Rappé, A.K.; Casewit, C.J.; Colwell, K.S.; Goddard, W.A. UFF, a full periodic table force field for molecular mechanics and molecular dynamics simulations. J. Am. Chem. Soc. 1992, 114, 10024–10035. [Google Scholar] [CrossRef]
- Weiner, S.J.; Kollman, P.A.; Nguyen, D.T.; Case, D.A. An all atom force field for simulations of proteins and nucleic acids. J. Comput. Chem. 1986, 7, 230–252. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Data Bank (PDB) Entry | Receptor | Ligand | Binding Energy (kcal/mol) | |
|---|---|---|---|---|
| Score-Only | Volume-Search | |||
| 1A52 | ERα | E | −9.7/−9.8 | −9.5/−9.8 |
| 1GWR | ERα | E | −9.7/−10.0 | −10.3/−10.1 |
| 2OCF | ERα | E | −9.4 | −9.7 |
| 2QA8 | ERα | G | −8.6/−9.6 | −8.7/−8.7 |
| 1X7R | ERα | G | −7.8 | −8.5 |
| 5TOA | ERβ | E | −9.7/−9.9 | −9.5/−9.5 |
| 1X7J | ERβ | G | −9.2/−8.6 | −8.6/−8.8 |
| 1QKM | ERβ | G | −9.5 | −9.4 |
| PDB Entry | Receptor | Binding Energy (kcal/mol) | |||
|---|---|---|---|---|---|
| (R)-N1 | (S)-N1 | (R)-N2 | (S)-N2 | ||
| 1A52 | ERα | −8.4/−8.5 | −8.2/−8.2 | −8.2/−8.9 | −9.2/−9.3 |
| 1GWR | ERα | −8.4/−8.6 | −8.3/−8.5 | −8.6/−8.6 | −8.9/−9.2 |
| 2OCF | ERα | −8.6 | −8.9 | −9.4 | −8.6 |
| 2QA8 | ERα | −8.8/−9.1 | −8.9/−9.5 | −8.8/−9.0 | −8.0/−8.3 |
| 1X7R | ERα | −9.1 | −8.4 | −8.1 | −8.5 |
| 5TOA | ERβ | −8.4/−8.6 | −8.2/−8.4 | −8.2/−8.7 | −7.2/−8.7 |
| 1X7J | ERβ | −9.2/−9.2 | −8.4/−8.9 | −8.5/−8.7 | −8.8/−9.0 |
| 1QKM | ERβ | −9.2 | −9.0 | −8.8 | −8.8 |
| Interacting Residues | ||||||
|---|---|---|---|---|---|---|
| Ligand | ERα | ERβ | ||||
| HI | HB | π-st. | HI | HB | π-st. | |
| E | Leu349 Ala350 Leu384 Leu387 Leu391 | Leu387 Arg394 His524 | Phe404 | Leu298 Ile373 Ile376 | Glu305 Arg346 His475 | Phe356 |
| G | Leu349 Leu387 Leu391 Ile424 | Ala350 Leu387 Arg394 His524 | Phe404 | Leu339 Ile373 Leu476 | Glu305 Arg346 His475 | Phe356 |
| N1 | Leu349 (S) Ala350 (S) Leu384 Ile 424 Leu525 (*1) (R) | Leu387 Arg394 His524 | Phe404 | Leu301 (*2) (R) Leu339 Leu343 (*3) Ile373 His475 (*4) (S) | Leu298 Leu339 (*5) Leu343 (*3) Arg346 His475 | Phe356 |
| N2 | Leu349 (S) Leu387 Leu391 Leu525 (*1) (R) | Thr347 Leu387 Arg394 His524 | Phe404 | Leu301 (*2) Leu339 | Arg346 Phe356 (*6) His475 | Phe356 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grande, F.; Rizzuti, B.; Occhiuzzi, M.A.; Ioele, G.; Casacchia, T.; Gelmini, F.; Guzzi, R.; Garofalo, A.; Statti, G. Identification by Molecular Docking of Homoisoflavones from Leopoldia comosa as Ligands of Estrogen Receptors. Molecules 2018, 23, 894. https://doi.org/10.3390/molecules23040894
Grande F, Rizzuti B, Occhiuzzi MA, Ioele G, Casacchia T, Gelmini F, Guzzi R, Garofalo A, Statti G. Identification by Molecular Docking of Homoisoflavones from Leopoldia comosa as Ligands of Estrogen Receptors. Molecules. 2018; 23(4):894. https://doi.org/10.3390/molecules23040894
Chicago/Turabian StyleGrande, Fedora, Bruno Rizzuti, Maria A. Occhiuzzi, Giuseppina Ioele, Teresa Casacchia, Fabrizio Gelmini, Rita Guzzi, Antonio Garofalo, and Giancarlo Statti. 2018. "Identification by Molecular Docking of Homoisoflavones from Leopoldia comosa as Ligands of Estrogen Receptors" Molecules 23, no. 4: 894. https://doi.org/10.3390/molecules23040894
APA StyleGrande, F., Rizzuti, B., Occhiuzzi, M. A., Ioele, G., Casacchia, T., Gelmini, F., Guzzi, R., Garofalo, A., & Statti, G. (2018). Identification by Molecular Docking of Homoisoflavones from Leopoldia comosa as Ligands of Estrogen Receptors. Molecules, 23(4), 894. https://doi.org/10.3390/molecules23040894