Melatonin Regulates the Synthesis of Steroid Hormones on Male Reproduction: A Review

{kind=link}
Abstract
:1. Introduction
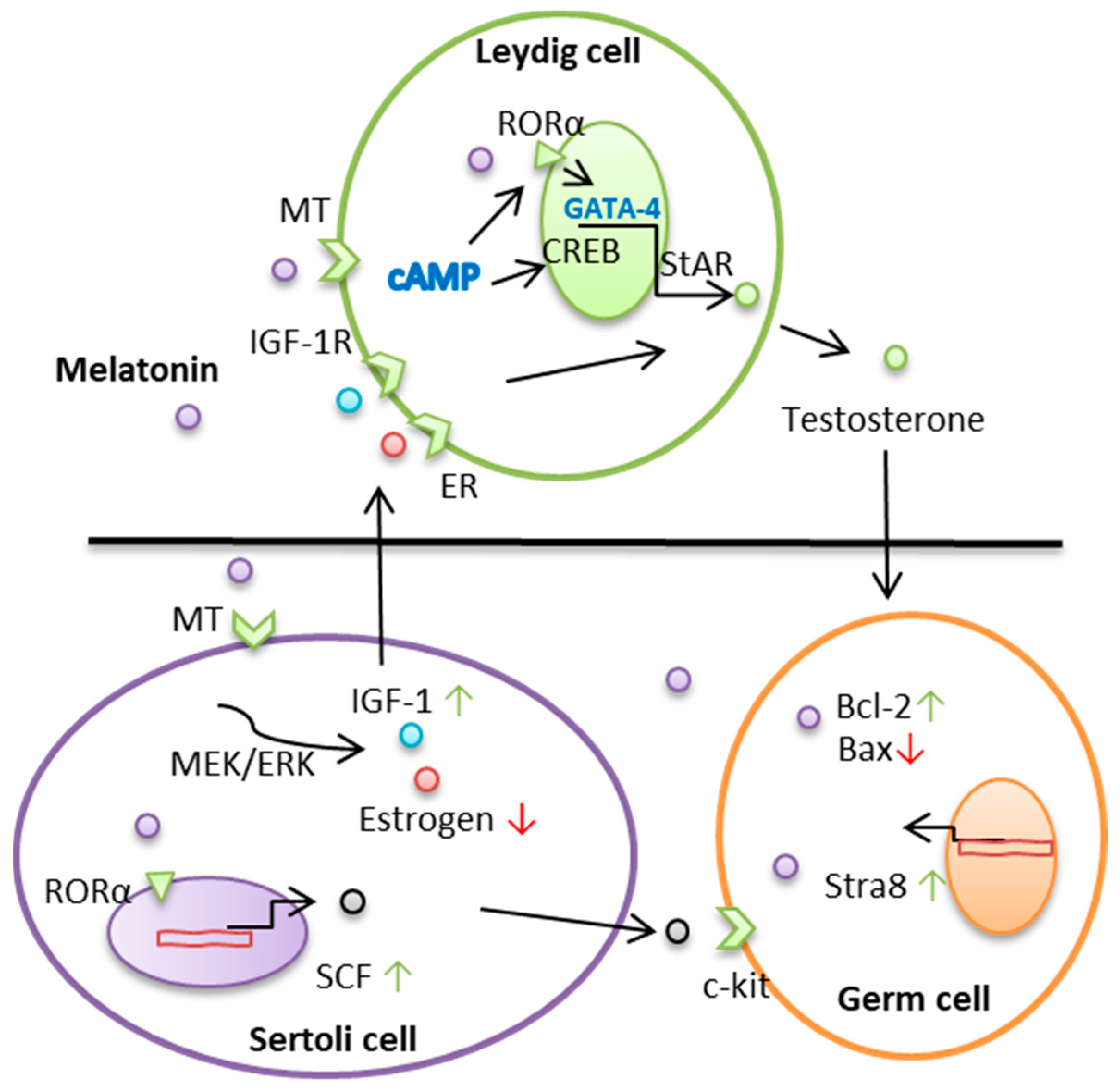
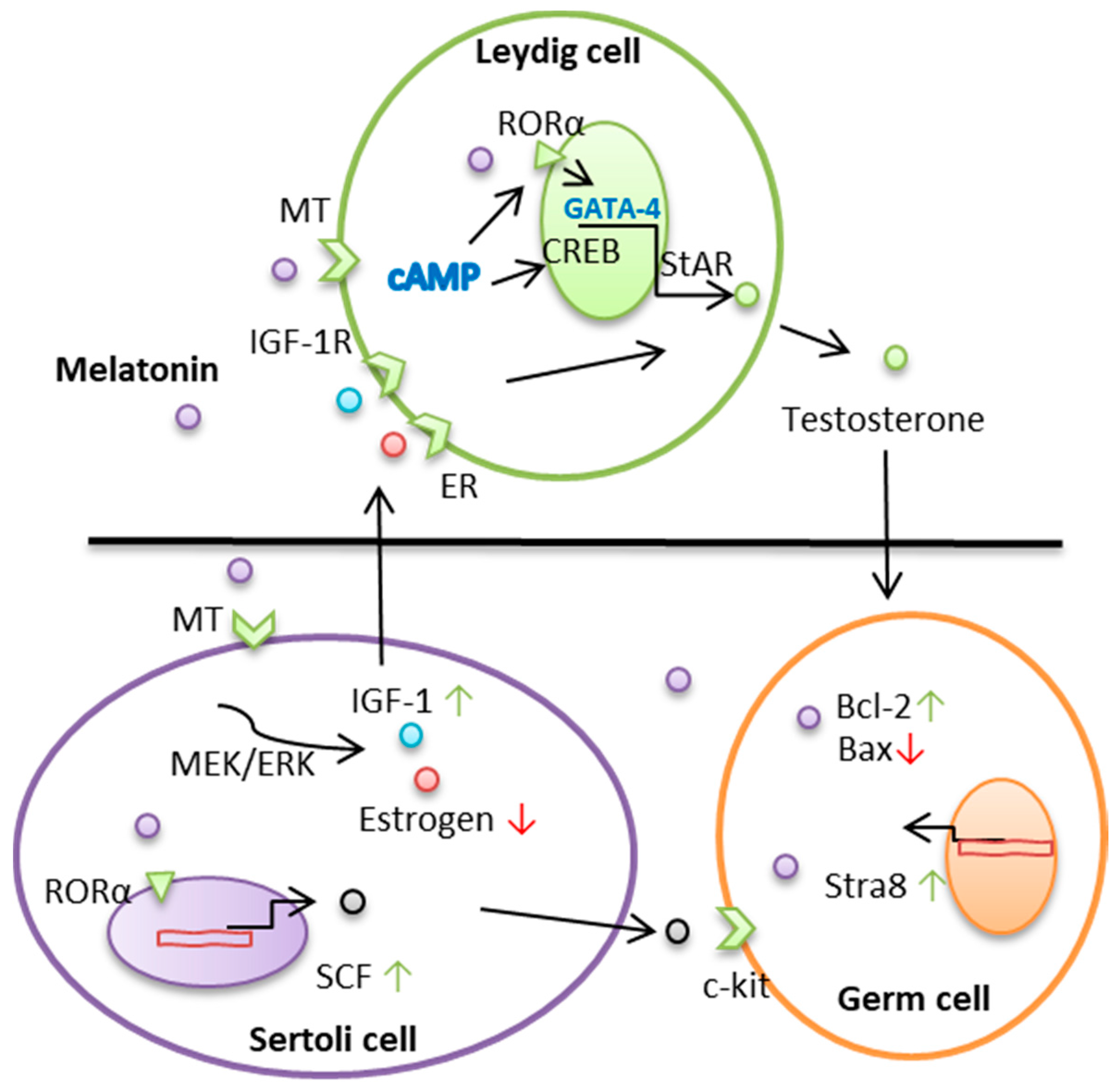
2. Melatonin Regulates Leydig Cell–Testosterone Secretions
3. Melatonin Regulates the Function of Sertoli Cells
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stocco, D.M.; Wang, X.; Jo, Y.; Manna, P.R. Multiple signaling pathways regulating steroidogenesis and steroidogenic acute regulatory protein expression: more complicated than we thought. Mol. Endocrinol. 2005, 19, 2647–5269. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.R.; Liu, Y.X. Regulation of spermatogonial stem cell self-renewal and spermatocyte meiosis by Sertoli cell signaling. J. Soc. Reprod. Fertil. 2015, 149, R159–R167. [Google Scholar] [CrossRef] [PubMed]
- Wen, Q.; Cheng, C.Y.; Liu, Y.X. Development, function and fate of fetal Leydig cells. Semin. Cell Dev. Biol. 2016, 59, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Barcelo, E.J.; Mediavilla, M.D.; Vriend, J.; Reiter, R.J. Constitutive photomorphogenesis protein 1 (COP1) and COP9 signalosome, evolutionarily conserved photomorphogenic proteins as possible targets of melatonin. J. Pineal Res. 2016, 61, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Cipolla-Neto, J.; Amaral, F.G.; Afeche, S.C.; Tan, D.X.; Reiter, R.J. Melatonin, energy metabolism, and obesity: A review. J. Pineal Res. 2014, 56, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Jetten, A.M. RORα is not a receptor for melatonin (response to DOI 10.1002/bies.201600018). Bioessays 2016, 38, 1193–1194. [Google Scholar] [CrossRef] [PubMed]
- Odawara, H.; Iwasaki, T.; Horiguchi, J.; Rokutanda, N.; Hirooka, K.; Miyazaki, W.; Koibuchi, Y.; Shimokawa, N.; Iino, Y.; Takeyoshi, I.; et al. Activation of aromatase expression by retinoic acid receptor-related orphan receptor (ROR) alpha in breast cancer cells: Identification of a novel ROR response element. J Biol. Chem. 2009, 284, 17711–17719. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Rosales-Corral, S.; Manchester, L.C. The universal nature, unequal distribution and antioxidant functions of melatonin and its derivatives. Mini Rev. Med. Chem. 2013, 13, 373–384. [Google Scholar] [PubMed]
- Ahmad, R.; Haldar, C. Effect of intra-testicular melatonin injection on testicular functions, local and general immunity of a tropical rodent Funambulus pennanti. Endocrine 2010, 37, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Zhang, J.; Zan, L.; Guo, W.; Wang, J.; Chen, L.; Cao, Y.; Shen, O.; Tong, J. Inhibitory effect of melatonin on testosterone synthesis is mediated via GATA-4/SF-1 transcription factors. Reprod. Biomed. Online 2015, 31, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Casao, A.; Pérez-Pé, R.; Abecia, J.A.; Forcada, F.; Muiño-Blanco, T.; Cebrián-Pérez, J.Á. The effect of exogenous melatonin during the non-reproductive season on the seminal plasma hormonal profile and the antioxidant defence system of Rasa Aragonesa rams. Anim. Reprod. Sci. 2013, 138, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Mura, M.C.; Luridiana, S.; Bodano, S.; Daga, C.; Cosso, G.; Diaz, M.L.; Bini, P.P.; Carcangiu, V. Influence of melatonin receptor 1A gene polymorphisms on seasonal reproduction in Sarda ewes with different body condition scores and ages. Anim. Reprod. Sci. 2014, 149, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhuo, Z.Y.; Shi, W.Q.; Tan, D.X.; Gao, C.; Tian, X.Z.; Zhang, L.; Zhou, G.B.; Zhu, S.E.; Yun, P.; et al. Melatonin promotes superovulation in sika deer (Cervus nippon). Int. J. Mol. Sci. 2014, 15, 12107–12118. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, M.; Madej, A. Effects of melatonin implants on plasma concentrations of testosterone, thyroxine and prolactin in the male silver fox (Vulpes vulpes). J. Reprod. Fertil. 1990, 89, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Rekik, M.; Taboubi, R.; Ben Salem, I.; Fehri, Y.; Sakly, C.; Lassoued, N.; Hilali, M.E. Melatonin administration enhances the reproductive capacity of young rams under a southern Mediterranean environment. Anim. Sci. J. 2015, 86, 66–672. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-A, M.; Luna, C.; Pérez-Pé, R.; Muiño-Blanco, T.; Cebrián-Pérez, J.A.; Casao, A. New evidence of melatonin receptor contribution to ram sperm functionality. Reprod. Fertil. Dev. 2016, 28, 924–935. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.L.; Chen, S.R.; Wang, Z.P.; Zhang, Y.; Tang, J.X.; Li, J.; Wang, X.X.; Cheng, J.M.; Jin, C.; Li, X.Y.; et al. Melatonin promotes development of haploid germ cells from early developing spermatogenic cells of Suffolk sheep under in vitro condition. J. Pineal Res. 2016, 60, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Viguie, C.; Caraty, A.; Locatelli, A.; Malpaux, B. Regulation of luteinizing hormone-releasing hormone (LHRH) secretion by melatonin in the ewe. II. Changes in N-methyl-D,L-aspartic acid-induced LHRH release during the stimulation of luteinizing hormone secretion by melatonin. Biol. Reprod. 1995, 52, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, K.L.; Yellon, S.M. Delayed puberty in the male Djungarian hamster: Effect of short photoperiod or melatonin treatment on the GnRH neuronal system. Neuroendocrinology 1991, 54, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Belsham, D.D. Melatonin receptor activation regulates GnRH gene expression and secretion in GT1-7 GnRH neurons. Signal transduction mechanisms. J. Biol. Chem. 2002, 277, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Ancel, C.; Bentsen, A.H.; Sébert, M.E.; Tena-Sempere, M.; Mikkelsen, J.D.; Simonneaux, V. Stimulatory effect of RFRP-3 on the gonadotrophic axis in the male Syrian hamster: The exception proves the rule. Endocrinology 2012, 153, 1352–1363. [Google Scholar] [CrossRef] [PubMed]
- Irwig, M.S.; Fraley, G.S.; Smith, J.T.; Acohido, B.V.; Popa, S.M.; Cunningham, M.J.; Gottsch, M.L.; Clifton, D.K.; Steiner, R.A. Kisspeptin activation of gonadotropin releasing hormone neurons and regulation of KiSS-1 mRNA in the male rat. Neuroendocrinology 2004, 80, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Ansel, L.; Bentsen, A.H.; Ancel, C.; Bolborea, M.; Klosen, P.; Mikkelsen, J.D.; Simonneaux, V. Peripheral kisspeptin reverses short photoperiod-induced gonadal regression in Syrian hamsters by promoting GNRH release. Reproduction 2011, 142, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Henningsen, J.B.; Poirel, V.J.; Mikkelsen, J.D.; Tsutsui, K.; Simonneaux, V.; Gauer, F. Sex differences in the photoperiodic regulation of RF-Amide related peptide (RFRP) and its receptor GPR147 in the syrian hamster. J. Comp. Neurol. 2016, 524, 1825–1838. [Google Scholar] [CrossRef] [PubMed]
- Mason, A.O.; Greives, T.J.; Scotti, M.A.; Levine, J.; Frommeyer, S.; Ketterson, E.D.; Demas, G.E.; Kriegsfeld, L.J. Suppression of kisspeptin expression and gonadotropic axis sensitivity following exposure to inhibitory day lengths in female Siberian hamsters. Horm. Behav. 2007, 52, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Revel, F.G.; Saboureau, M.; Masson-Pévet, M.; Pévet, P.; Mikkelsen, J.D.; Simonneaux, V. Kisspeptin mediates the photoperiodic control of reproduction in hamsters. Curr. Biol. 2006, 16, 1730–1735. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, O.; Gioacchini, G.; Maradonna, F.; Olivotto, I.; Migliarini, B. Melatonin induces follicle maturation in Danio rerio. PLoS One. 2011, 6, e19978. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, M.V.; Carrillo, M.; Felip, A. Melatonin-induced changes in kiss/gnrh gene expression patterns in the brain of male sea bass during spermatogenesis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 185, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, P. Expression of EAP1 and CUX1 in the hypothalamus of female rats and relationship with KISS1 and GnRH. Endocr. J. 2016, 63, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, X. Melatonin and male reproduction. Clin. Chim. Acta. 2015, 446, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Baburski, A.Z.; Sokanovic, S.J.; Janjic, M.M.; Stojkov-Mimic, N.J.; Bjelic, M.M.; Andric, S.A.; Kostic, T.S. Melatonin replacement restores the circadian behavior in adult rat Leydig cells after pinealectomy. Mol. Cell Endocrinol. 2015, 413, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Valenti, S.; Thellung, S.; Florio, T.; Giusti, M.; Schettini, G.; Giordano, G. A novel mechanism for the melatonin inhibition of testosterone secretion by rat Leydig cells: reduction of GnRH-induced increase in cytosolic Ca2+. J. Mol. Endocrinol. 1999, 23, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, M.F.; Taniguchi, H.; Viger, R.S. The effect of human GATA4 gene mutations on the activity of target gonadal promoters. J. Mol. Endocrinol. 2009, 42, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Svechnikov, K.; Landreh, L.; Weisser, J.; Izzo, G.; Colón, E.; Svechnikova, I.; Söder, O. Origin, development and regulation of human Leydig cells. Horm. Res. Paediatr. 2010, 73, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.L.; Zhang, Y.; Yu, K.; Wang, X.X.; Chen, S.R.; Han, D.P.; Cheng, C.Y.; Lian, Z.X.; Liu, Y.X. Melatonin up-regulates the expression of the GATA-4 transcription factor and increases testosterone secretion from Leydig cells through RORα signaling in an in vitro goat spermatogonial stem cell differentiation culture system. Oncotarget 2017, 8, 110592–110605. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.K.; Krishna, A. Melatonin affects steroidogenesis and delayed ovulation during winter in vespertilionid bat, Scotophilus heathi. J. Steroid. Biochem. Mol. Biol. 2010, 118, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.S.; Lai, F.P.; Lo, R.K.; Voyno-Yasenetskaya, T.A.; Stanbridge, E.J.; Wong, Y.H. Melatonin mt1 and MT2 receptors stimulate c-Jun N-terminal kinase via pertussis toxin-sensitive and -insensitive G proteins. Cell. Signal. 2002, 14, 249–257. [Google Scholar] [CrossRef]
- Rossi, S.P.; Matzkin, M.E.; Terradas, C.; Ponzio, R.; Puigdomenech, E.; Levalle, O.; Calandra, R.S.; Frungieri, M.B. New insights into melatonin/CRH signaling in hamster Leydig cells. Gen. Comp. Endocrinol. 2012, 178, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.R.; Stocco, D.M. Crosstalk of CREB and Fos/Jun on a single cis-element: transcriptional repression of the steroidogenic acute regulatory protein gene. J. Mol. Endocrinol. 2007, 39, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Bilińska, B.; Schmalz-Fraczek, B.; Kotula, M.; Carreau, S. Photoperiod-dependent capability of androgen aromatization and the role of estrogens in the bank vole testis visualized by means of immunohistochemistry. Mol. Cell. Endocrinol. 2001, 178, 189–198. [Google Scholar] [CrossRef]
- Gould, M.L.; Hurst, P.R.; Nichol, H.D. The effects of oestrogen receptors alpha and beta on testicular cell number and steroidogenesis in mice. Reproduction 2007, 134, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Carreau, S. Germ cells: A new source of estrogens in the male gonad. Mol Cell Endocrinol 2001, 178, 65–72. [Google Scholar] [CrossRef]
- Letellier, K.; Azeddine, B.; Parent, S.; Labelle, H.; Rompré, P.H.; Moreau, A.; Moldovan, F. Estrogen cross-talk with the melatonin signaling pathway in human osteoblasts derived from adolescent idiopathic scoliosis patients. J. Pineal Res. 2008, 45, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Yoshitane, H.; Honma, S.; Imamura, K.; Nakajima, H.; Nishide, S.Y.; Ono, D.; Kiyota, H.; Shinozaki, N.; Matsuki, H.; Wada, N.; et al. JNK regulates the photic response of the mammalian circadian clock. EMBO Rep. 2012, 13, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Vriend, J.; Reiter, R.J. Breast cancer: Modulation by melatonin and the ubiquitin-proteasome system--A review. Mol. Cell. Endocrinol. 2015, 417, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.L.; Wang, Z.P.; Jin, C.; Kang, X.L.; Batool, A.; Zhang, Y.; Li, X.Y.; Wang, X.X.; Chen, S.R.; Chang, C.S.; et al. Melatonin promotes sheep Leydig cell testosterone secretion in a co-culture with Sertoli cells. Theriogenology 2018, 106, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Rocha, C.S.; Martins, A.D.; Rato, L.; Silva, B.M.; Oliveira, P.F.; Alves, M.G. Melatonin alters the glycolytic profile of Sertoli cells: implications for male fertility. Mol. Hum. Reprod. 2014, 20, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, T.; Abe, K.; Mekada, K.; Yoshiki, A.; Kato, T. Genetic variation of melatonin productivity in laboratory mice under domestication. Proc. Natl. Acad. Sci. USA 2010, 107, 6412–6417. [Google Scholar] [CrossRef] [PubMed]
- Frungieri, M.B.; Calandra, R.S.; Rossi, S.P. Local actions of melatonin in somatic cells of the testis. Int. J. Mol. Sci. 2017, 18, 6. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Wang, X.; Wang, Z.; Chen, S.; Wang, Y.; Hao, X.; Sun, T.; Zhang, Y.; Lian, Z.; Liu, Y. In vitro production of functional haploid sperm cells from male germ cells of Saanen dairy goat. Theriogenology 2017, 90, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.L.; Sun, T.C.; Yu, K.; Wang, Z.P.; Zhang, B.L.; Zhang, Y.; Wang, X.X.; Lian, Z.X.; Liu, Y.X. Melatonin reduces oxidative damage and upregulates heat shock protein 90 expression in cryopreserved human semen. Free Radic. Biol. Med. 2017, 113, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Gholami, M.; Saki, G.; Hemadi, M.; Khodadadi, A.; Mohammadi-Asl, J. Melatonin improves spermatogonial stem cells transplantation efficiency in azoospermic mice. Iran J. Basic Med. Sci. 2014, 17, 93–99. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, K.; Deng, S.-L.; Sun, T.-C.; Li, Y.-Y.; Liu, Y.-X. Melatonin Regulates the Synthesis of Steroid Hormones on Male Reproduction: A Review. Molecules 2018, 23, 447. https://doi.org/10.3390/molecules23020447
Yu K, Deng S-L, Sun T-C, Li Y-Y, Liu Y-X. Melatonin Regulates the Synthesis of Steroid Hormones on Male Reproduction: A Review. Molecules. 2018; 23(2):447. https://doi.org/10.3390/molecules23020447
Chicago/Turabian StyleYu, Kun, Shou-Long Deng, Tie-Cheng Sun, Yuan-Yuan Li, and Yi-Xun Liu. 2018. "Melatonin Regulates the Synthesis of Steroid Hormones on Male Reproduction: A Review" Molecules 23, no. 2: 447. https://doi.org/10.3390/molecules23020447
APA StyleYu, K., Deng, S.-L., Sun, T.-C., Li, Y.-Y., & Liu, Y.-X. (2018). Melatonin Regulates the Synthesis of Steroid Hormones on Male Reproduction: A Review. Molecules, 23(2), 447. https://doi.org/10.3390/molecules23020447