Lavandula angustifolia Essential Oil and Linalool Counteract Social Aversion Induced by Social Defeat

 ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Effect of LA Essential Oil and Linalool on Anxiety and Social Behaviors
2.1.1. Experiment 1: Acute Effects of Linalool and LA Essential Oil
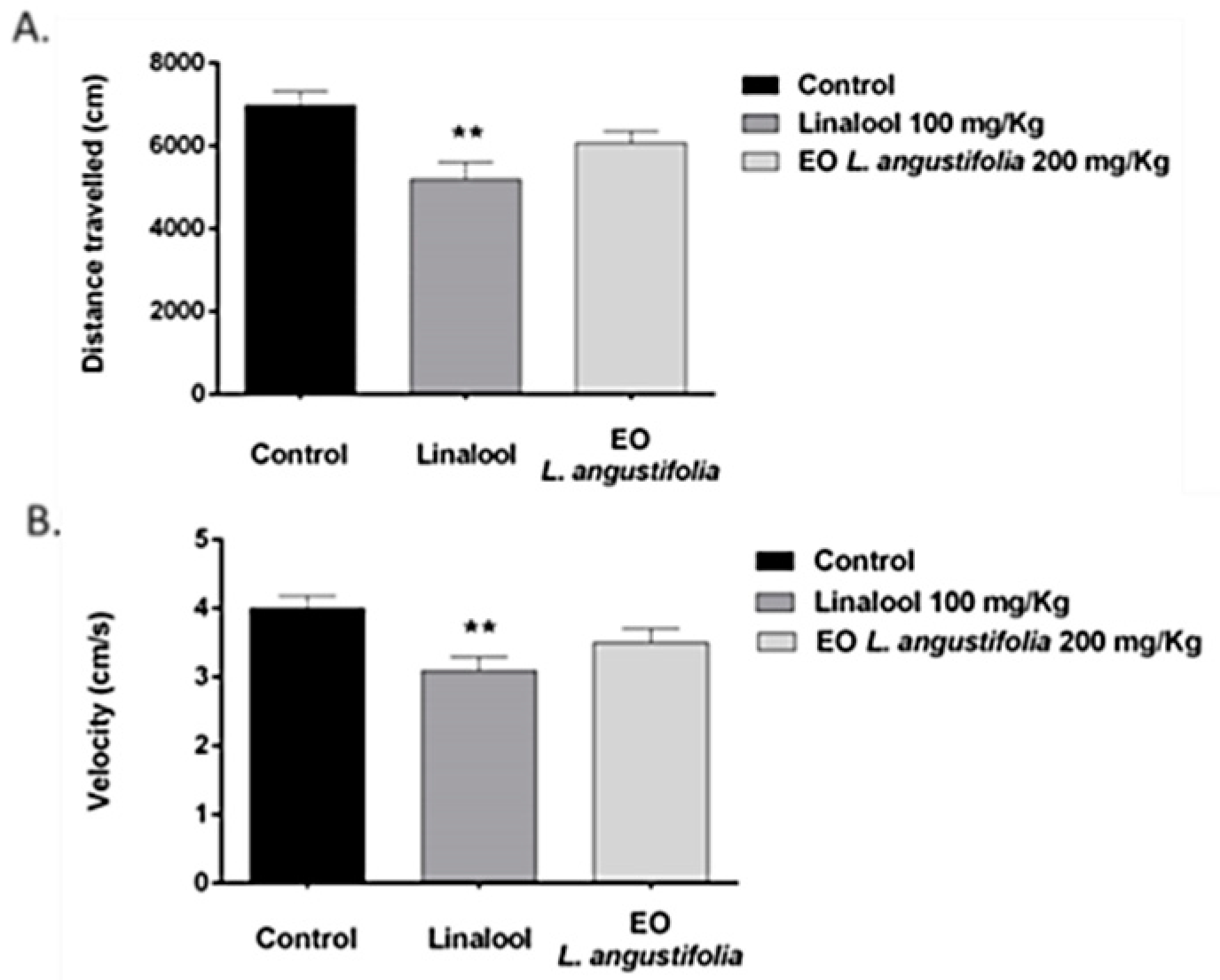
Open Field
Elevated Plus Maze
Social Interaction Test
2.1.2. Experiment 2: Linalool and LA Essential Oil Effects When Administered after Social Stress
Elevated Plus Maze
Social Interaction Test
2.1.3. Experiment 3: Effects of Acute or Chronic Administration of Linalool and LA Essential Oil before Social Defeat in Mice
Social Defeat Encounters
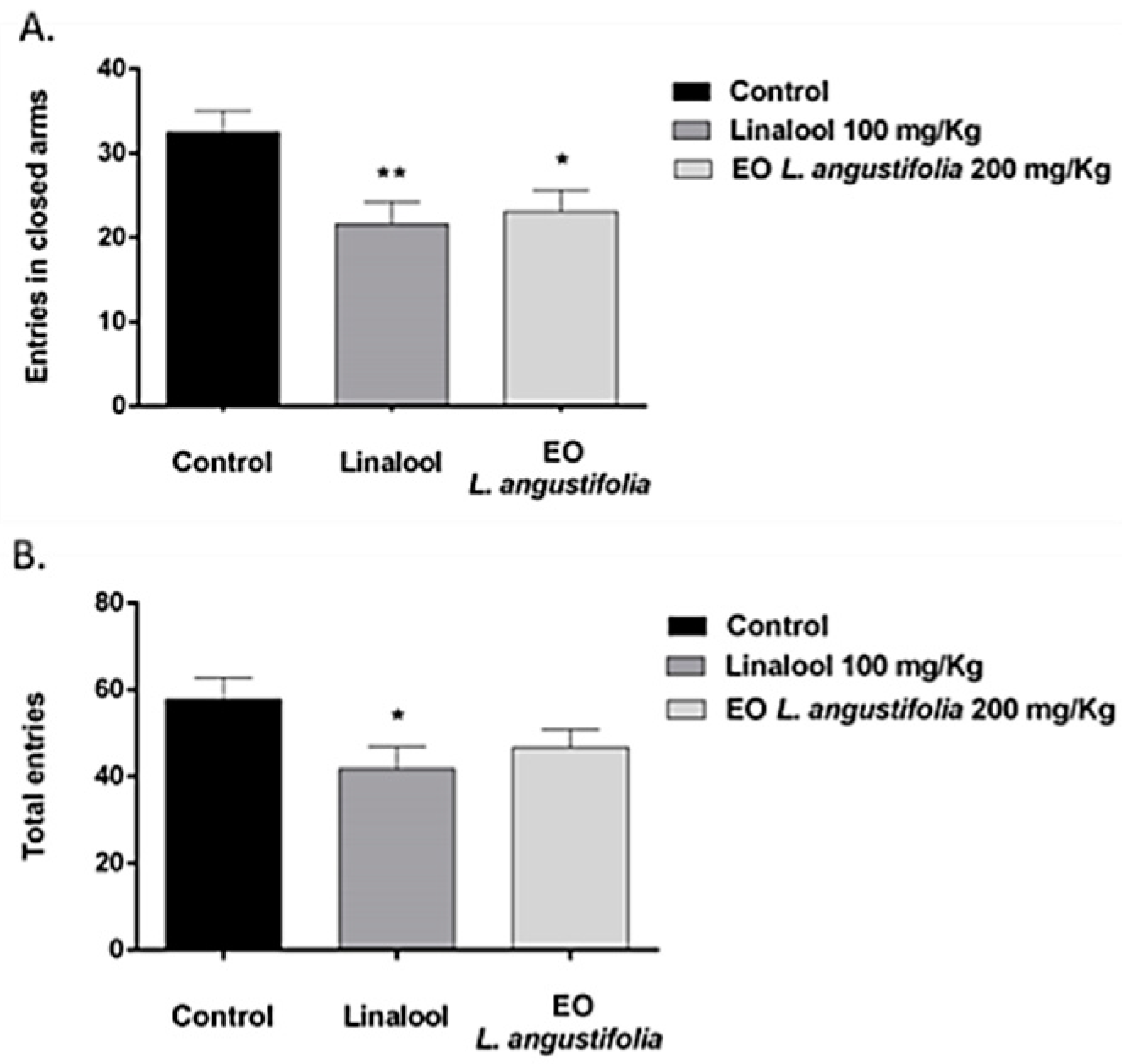
Elevated Plus Maze
Social Interaction Test
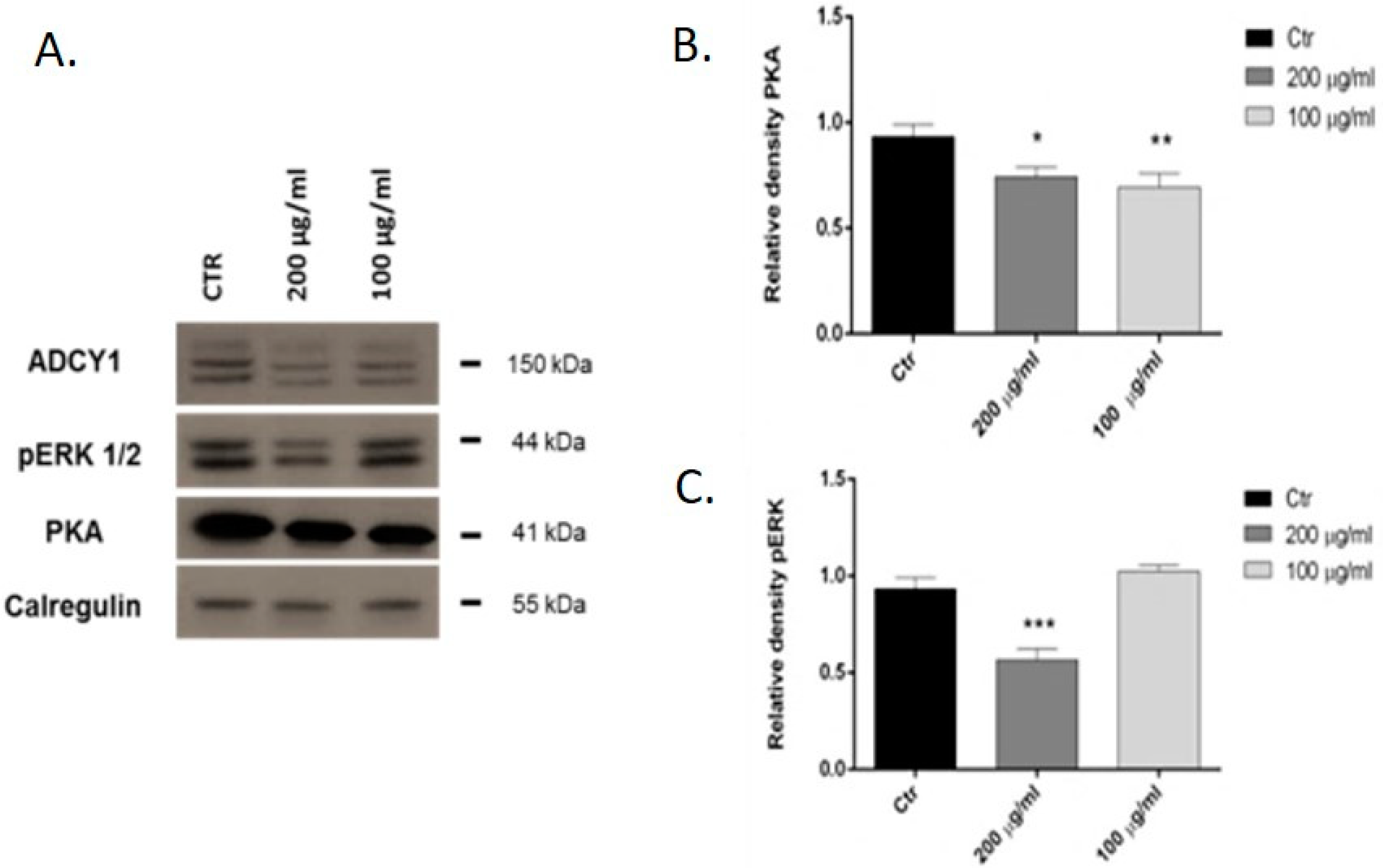
2.2. Western Blot Analysis: pERK and PKA
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drug Treatment
4.3. Experimental Design
4.4. Open-Field Test
4.5. Elevated Plus Maze-EPM
4.6. Social Encounters
4.7. Procedure of Social Defeat
4.8. Human Neuroblastoma Cell Cultures
4.9. Extraction of Proteins and Western Blotting
4.10. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Weismann, M.; Yousry, I.; Heuberger, E.; Nolte, A.; Ilmberger, J.; Kobal, G.; Naidich, T.P. Functional magnetic resonance imaging of human olfaction. Neuroimaging Clin. N. Am. 2001, 11, 237–250. [Google Scholar] [PubMed]
- Lee, Y.L.; Wu, Y.; Tsang, H.W.; Leung, A.Y.; Cheung, W.M. A systematic review on the anxiolytic effects of aromatherapy in people with anxiety symptoms. J. Altern. Complement. Med. 2011, 17, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Dobetsberger, C.; Buchbauer, G. Actions of essential oils on the central nervous system: An updated review. Flavour Frag. J. 2011, 26, 300–316. [Google Scholar] [CrossRef]
- World Health Organization. Depression and Other Common Mental Disorders: Global Health Estimates; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Linck, V.; da Silva, A.; Figueiró, M.; Caramão, E.B.; Moreno, P.R.H.; Elisabetsky, E. Effects of inhaled Linalool in anxiety, social interaction and aggressive behavior in mice. Phytomedicine 2010, 17, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Woelk, H.; Schläfke, S. A multi-center, double-blind, randomised study of the Lavender oil preparation Silexan in comparison to Lorazepam for generalized anxiety disorder. Phytomedicine 2010, 17, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Koulivand, P.H.; Ghadiri, M.K.; Gorji, A. Lavender and the nervous system. Evid.-Based Complement. Altern. Med. 2013, 2012, 681304. [Google Scholar] [CrossRef] [PubMed]
- Wotman, M.; Levinger, J.; Leung, L.; Kallush, A.; Mauer, E.; Kacker, A. The Efficacy of Lavender Aromatherapy in Reducing Preoperative Anxiety in Ambulatory Surgery Patients Undergoing Procedures in General Otolaryngology. Laryngoscope Investig. Otolaryngol. 2017, 2, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Farshbaf-Khalili, A.; Kamalifard, M.; Namadian, M. Comparison of the effect of lavender and bitter orange on anxiety in postmenopausal women: A triple-blind, randomized, controlled clinical trial. Complement. Ther. Clin. Pract. 2018, 31, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Qadeer, S.; Emad, S.; Perveen, T.; Yousuf, S.; Sheikh, S.; Sarfaraz, Y.; Sadaf, S.; Haider, S. Role of ibuprofen and lavender oil to alter the stress induced psychological disorders: A comparative study. Pak. J. Pharm. Sci. 2018, 31, 1603–1608. [Google Scholar] [PubMed]
- Giovannini, D.; Gismondi, A.; Basso, A.; Canuti, L.; Braglia, R.; Canini, A.; Mariani, F.; Cappelli, G. Lavandula angustifolia Mill. Essential Oil Exerts Antibacterial and Anti-Inflammatory Effect in Macrophage Mediated Immune Response to Staphylococcus aureus. Immunol. Investing. 2016, 45, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, K.; Lu, C.; Dong, L.; Gao, L.; Yan, M.; Liu, X. Protective effect of lavender oil on scopolamine induced cognitive deficits in mice and H2O2 induced cytotoxicity in PC12 cells. J. Ethnopharmacol. 2016, 193, 408–415. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency (EMA); Committee on Herbal Medicinal Products (HPMC). Community Herbal Monograph on Lavandula angustifolia Mill, aetheroleum; EMA/HMPC/143181/2010; European Medicines Agency: London, UK, 2012. [Google Scholar]
- López, V.; Nielsen, B.; Solas, M.; Ramírez, M.J.; Jäger, A.K. Exploring pharmacological mechanisms of lavender (Lavandula angustifolia) essential oil on central nervous system targets. Front. Pharmacol. 2017, 8, 280. [Google Scholar] [CrossRef] [PubMed]
- Elisabetsky, E.; Marschner, J.; Souza, D.O. Effects of linalool on glutamatergic system in the rat cerebral cortex. Neurochem. Res. 1995, 20, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Elisabetsky, E.; Silva Brum, L.F.; Souza, D.O. Anticonvulsant properties of linalool in glutamate-related seizure models. Phytomedicine 1999, 6, 107–113. [Google Scholar] [CrossRef]
- Anjos, P.J.; Lima, A.O.; Cunha, P.S.; De Sousa, D.P.; Onofre, A.S.; Ribeiro, T.P.; Medeiros, I.A.; Antoniolli, A.R.; Quintans-Júnior, L.J.; Santosa, M.R. Cardiovascular effects induced by linalool in normotensive and hypertensive rats. Z. Naturforsch. C 2013, 68, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Huo, M.; Cui, X.; Xue, J.; Chi, G.; Gao, R.; Deng, X.; Guan, S.; Wei, J.; Soromou, L.W.; Feng, H.; et al. Anti-inflammatory effects of linalool in RAW 264.7 macrophages and lipopolysaccharide-induced lung injury model. J. Surg. Res. 2013, 180, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Seol, G.H.; Ryu, S.; Choi, I.Y. Neuroprotective effects of (−)-linalool against oxygen-glucose deprivation-induced neuronal injury. Arch. Pharm. Res. 2016, 39, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Sabogal-Guáqueta, A.M.; Osorio, E.; Cardona-Gómez, G.P. Linalool reverses neuropathological and behavioral impairments in old triple transgenic Alzheimer’s mice. Neuropharmacology 2016, 102, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Coelho, V.; Mazzardo-Martins, L.; Martins, D.F.; Santos, A.R.S.; da Silva Brum, L.F.; Picada, J.N.; Pereira, P. Neurobehavioral and genotoxic evaluation of (−)-linalool in mice. J. Nat. Med. 2013, 67, 876–880. [Google Scholar] [CrossRef] [PubMed]
- Caputo, L.; Souza, L.F.; Alloisio, S.; Cornara, L.; De Feo, V. Coriandrum sativum and Lavandula angustifolia Essential Oils: Chemical Composition and Activity on Central Nervous System. Int. J. Mol. Sci. 2016, 17, 1999. [Google Scholar] [CrossRef] [PubMed]
- Caputo, L.; Nazzaro, F.; Souza, L.F.; Aliberti, L.; De Martino, L.; Fratianni, F.; De Feo, V. Laurus nobilis: Composition of Essential Oil and Its Biological Activities. Molecules 2017, 22, 930. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, G.D.; Storm, D.R. Why calcium-stimulated adenylyl cyclases? Physiology 2004, 19, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Covington, H.E., 3rd; Nikulina, E.M., Jr.; Hammer, R.P. Aggression and defeat: Persistent effects on cocaine self-administration and gene expression in peptidergic and aminergicmesocorticolimbic circuits. Neurosci. Biobehav. Rev. 2004, 27, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Yap, J.J.; Covington, H.E., 3rd. Social stress, therapeutics and drug abuse: Preclinical models of escalated and depressed intake. Pharmacol. Therapeut. 2008, 120, 102–128. [Google Scholar] [CrossRef] [PubMed]
- Montagud-Romero, S.; Reguilon, M.D.; Roger-Sanchez, C.; Pascual, M.; Aguilar, M.A.; Guerri, C.; Miñarro, J.; Rodríguez-Arias, M. Role of dopamine neurotransmission in the long-term effects of repeated social defeat on the conditioned rewarding effects of cocaine. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 71, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Montagud-Romero, S.; Nuñez, C.; Blanco-Gandia, M.C.; Martínez-Laorden, E.; Aguilar, M.A.; Navarro-Zaragoza, J.; Almela, P.; Milanés, M.V.; Laorden, M.L.; Miñarro, J.; et al. Repeated social defeat and the rewarding effects of cocaine in adult and adolescent mice: Dopamine transcription factors, proBDNF signaling pathways, and the TrkB receptor in the mesolimbic system. Psychopharmacology 2017, 234, 2063–2075. [Google Scholar] [CrossRef] [PubMed]
- Stein, D.J.; Vasconcelos, M.F.; Albrechet-Souza, L.; Ceresér, K.M.M.; de Almeida, R.M.M. Microglial Over-Activation by Social Defeat Stress Contributes to Anxiety- and Depressive-Like Behaviors. Front. Behav. Neurosci. 2017, 11, 207. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Flanigan, M.E.; McEwen, B.S.; Russo, S.J. Aggression, Social Stress, and the Immune System in Humans and Animal Models. Front. Behav. Neurosci. 2018, 12, 56. [Google Scholar] [CrossRef] [PubMed]
- Malcolm, B.J.; Tallian, K. Essential oil of lavender in anxiety disorders: Ready for prime time? MHC 2018, 7, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Toda, M.; Morimoto, K. Effect of lavender aroma on salivary endocrinological stress markers. Arch. Oral. Biol. 2008, 53, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Ayuob, N.N.; Firgany, A.E.L.; El-Mansy, A.A.; Ali, S. Can Ocimumbasilicum relieve chronic unpredictable mild stress-induced depression in mice? Exp. Mol. Pathol. 2017, 103, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Ayuob, N.N.; El Wahab, M.G.A.; Ali, S.S.; Abdel-Tawab, H.S. Ocimumbasilicum improve chronic stress-induced neurodegenerative changes in mice hippocampus. Metab. Brain Dis. 2018, 33, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, C.; Duman, R.S. Stress, depression, and neuroplasticity: A convergence of mechanisms. Neuropsychopharmacology 2008, 33, 88–109. [Google Scholar] [CrossRef] [PubMed]
- Higashida, S.; Nagai, H.; Nakayama, K.; Shinohara, R.; Taniguchi, M.; Nagai, M.; Hikida, T.; Yawata, S.; Ago, Y.; Kitaoka, S.; et al. Repeated social defeat stress impairs attentional set shifting irrespective of social avoidance and increases female preference associated with heightened anxiety. Sci. Rep. 2018, 8, 10454. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.; Annett, J.M.; Doherty, B.; Leslie, J.C. Anxiolytic effects of lavender oil inhalation on open-field behavior in rats. Phytomedicine 2007, 14, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Chioca, L.R.; Ferro, M.M.; Baretta, I.P.; Oliveira, S.M.; Silva, C.R.; Ferreira, J.; Losso, E.M.; Andreatini, R. Anxiolytic-like effect of lavender essential oil inhalation in mice: Participation of serotonergic but not GABAA/benzodiazepine neurotransmission. J. Ethnopharmacol. 2013, 147, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V. Characterization of anxiolytic and neuropharmacological activities of Silexan. Wien Med. Wochenschr. 2013, 163, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Linck, V.; da Silva, A.L.; Figueiró, M.; Piato, A.L.; Herrmann, A.P.; Birck, F.D.; Elisabetsky, E. Inhaled linalool-induced sedation in mice. Phytomedicine 2009, 16, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Kingston, R.C.; Smith, M.; Lacey, T.; Edwards, M.; Best, J.N.; Markham, C.M. Voluntary exercise increases resilience to social defeat stress in Syrian hamsters. Physiol. Behav. 2018, 188, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Macedo, G.C.; Morita, G.M.; Domingues, L.P.; Favoretto, C.A.; Suchecki, D.; Quadros, I.M.H. Consequences of continuous social defeat stress on anxiety- and depressive-like behaviors and ethanol reward in mice. Horm. Behav. 2018, 97, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Szyszkowicz, J.K.; Wong, A.; Anisman, H.; Merali, Z.; Audet, M.C. Implications of the gut microbiota in vulnerability to the social avoidance effects of chronic social defeat in male mice. Brain Behav. Immun. 2017, 66, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Chamero, P.; Marton, T.F.; Logan, D.W.; Flanagan, K.; Cruz, J.R.; Saghatelian, A.; Cravatt, B.F.; Stowers, L. Identification of protein pheromones that promote aggressive behaviour. Nature 2007, 450, 899–902. [Google Scholar] [CrossRef] [PubMed]
- Mugford, R.A.; Nowell, N.W. Pheromones and their effect on aggression in mice. Nature 1970, 226, 967–968. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.D. Pheromones and signature mixtures: Defining species-wide signals and variable cues for identity in both invertebrates and vertebrates. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2010, 196, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Karlson, P.; Luscher, M. Pheromones’: A new term for a class of biologically active substances. Nature 1959, 183, 55–56. [Google Scholar] [CrossRef] [PubMed]
- Stowers, L.; Kuo, T.H. Mammalian pheromones: Emerging properties and mechanisms of detection. Curr. Opin. Neurobiol. 2015, 34, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.B.; Stettler, D.D.; Kallman, B.R.; Bhaskar, S.T.; Fleischmann, A.; Axel, R. Driving opposing behaviors with ensembles of piriform neurons. Cell 2011, 146, 1004–1015. [Google Scholar] [CrossRef] [PubMed]
- Grewal, S.S.; York, R.D.; Stork, P.J. Extracellular-signal-regulated kinase signalling in neurons. Curr. Opin. Neurobiol. 1999, 9, 544–553. [Google Scholar] [CrossRef]
- Martin, K.C.; Michael, D.; Rose, J.C.; Barad, M.; Casadio, A.; Zhu, H.; Kandel, E.R. MAP kinase translocates into the nucleus of the presynaptic cell and is required for long-term facilitation in aplysia. Neuron 1997, 18, 899–912. [Google Scholar] [CrossRef]
- Villalba, M.; Bockaert, J.; Journot, L. Pituitary adenylate cyclase activating polypeptide (PACAP-38) protects cerebellar granule neurons from apoptosis by activating the mitogen-activated protein kinase (MAP kinase) pathway. J. Neurosci. 1997, 17, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.D.; Cruz, F. The ERK 1 and 2 pathway in the nervous system: From basic aspects to possible clinical applications in pain and visceral dysfunction. Curr. Neuropharmacol. 2007, 5, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Baldinger, P.; Hoflich, A.S.; Mitterhauser, M.; Hahn, A.; Rami-Mark, C.; Spies, M.; Wadsak, W.; Lanzenberger, R.; Kasper, S. Effects of Silexan on the serotonin-1A receptor and microstructure of the human brain: A randomized, placebo-controlled, double-blind, cross-over study with molecular and structural neuroimaging. Int. J. Neuropsychopharmacol. 2015, 18, 63. [Google Scholar] [CrossRef] [PubMed]
- Spindelegger, C.; Lanzenberger, R.; Wadsak, W.; Mien, L.K.; Stein, P.; Mitterhauser, M.; Moser, U.; Holik, A.; Pezawas, L.; Kletter, K.; et al. Influence of escitalopram treatment on 5-HT 1A receptor binding in limbic regions in patients with anxiety disorders. Mol. Psychiatry 2009, 14, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Kasper, S.; Gastpar, M.; Muller, W.E.; Volz, H.P.; Moller, H.J.; Schlafke, S.; Dienel, A. Lavender oil preparation Silexan is effective in generalized anxiety disorder—A randomized, double-blind comparison to placebo and paroxetine. Int. J. Neuropsychopharmacol. 2014, 17, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Schuwald, A.M.; Noldner, M.; Wilmes, T.; Klugbauer, N.; Leuner, K.; Muller, W.E. Lavender oil-potent anxiolytic properties via modulating voltage dependent calcium channels. PLoS ONE 2013, 8, e59998. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, M.; Kim, H.; Kim, K. Effect of lavender oil on motor function and dopamine receptor expression in the olfactory bulb of mice. J. Ethnopharmacol. 2009, 125, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Daza-Losada, M.; Rodríguez-Arias, M.; Maldonado, C.; Aguilar, M.A.; Guerri, C.; Miñarro, J. Acute behavioural and neurotoxic effects of MDMA plus cocaine in adolescent mice. Neurotoxicol. Teratol. 2009, 31, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Smoothy, R.; Brain, P.F.; Berry, M.S.; Haug, M. Alcohol and social behaviour in group housed female mice. Physiol. Behav. 1986, 37, 689–694. [Google Scholar] [CrossRef]
- Brain, P.F.; Benton, D.; Childs, G.; Parmigiani, S. The effect of the type of opponent in test of murine aggression. Behav. Process. 1981, 6, 319–327. [Google Scholar] [CrossRef]
- Rodriguez-Arias, M.; Miñarro, J.; Aguilar, M.A.; Pinazo, J.; Simon, V.M. Effects of risperidone and SCH 23390 on isolation-induced aggression in male mice. Eur. Neuropsychopharmacol. 1998, 8, 95–103. [Google Scholar] [CrossRef]
- Tornatzky, W.; Miczek, K.A. Long-term impairment of autonomic circadian rhythms after brief intermittent social stress. Physiol. Behav. 1993, 53, 983–993. [Google Scholar] [CrossRef]
- Covington, H.E.; Miczek, K.A. Repeated social-defeat stress, cocaine or morphine. Effects on behavioral sensitization and intravenous cocaine self-administration ‘binges’. Psychopharmacology 2001, 158, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Yoshimura, H. Disruption of primate social behavior by d-amphetamine and cocaine: Differential antagonism by antipsychotics. Psychopharmacology 1982, 76, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Festa, M.; Del Valle, L.; Khalili, K.; Franco, R.; Scognamiglio, G.; Graziano, V.; Rosati, A. BAG3 protein is overexpressed in human glioblastoma and is a potential target for therapy. Am. J. Pathol. 2011, 178, 2504–2512. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of linalool and Lavandula angustifolia essential oil are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Control | Linalool | L. angustifolia EO | |
|---|---|---|---|
| Non-social exploration | 534 ± 5 | 494 ± 9 ** | 491 ± 12 ** |
| Exploration from a distance | 3 ± 0.3 | 2 ± 0.3 | 2 ± 0.4 |
| Social investigation | 50 ± 6 | 83 ± 8 * | 84 ± 10 * |
| Unit of social investigation | 1 ± 0.2 | 3 ± 0.2 *** | 2 ± 0.3 ** |
| Latency of social investigation | 20 ± 3 | 13 ± 2 | 12 ± 2 |
| Control (Explora) | SD Saline | SD Linalool | SD L. angustifolia EO | |
|---|---|---|---|---|
| Time in open arms | 52 ± 7 | 27 ± 4 * | 21 ± 6 ** | 34 ± 7 |
| % Time in open arms | 22 ± 2 | 13 ± 3 | 10 ± 3 * | 14 ± 4 |
| Time in central platform | 38 ± 4 | 50 ± 9 | 37 ± 12 | 40 ± 10 |
| Time in closed arms | 169 ± 14 | 213 ± 13 | 244 ± 18 ** | 225 ± 16 * |
| Entries in open arms | 24 ± 4 | 26 ± 6 | 9 ± 3 * | 15 ± 2 |
| % Open entries | 32 ± 3 | 35 ± 5 | 29 ± 7 | 31 ± 4 |
| Entries in closed arms | 49 ± 5 | 42 ± 3 | 22 ± 6 *** | 33 ± 4 |
| Total entries | 73 ± 7 | 68 ± 8 | 28 ± 8 *** | 48 ± 6 * |
| Control (Explora) | SD Saline | SD Linalool | SD L. angustifolia EO | |
|---|---|---|---|---|
| Non-social exploration | 448 ± 14 | 492 ± 8 * | 433 ± 13 ## | 447 ± 12 # |
| Exploration from a distance | 3 ± 0.5 | 3 ± 1 | 3 ± 0.4 | 3 ± 0.4 |
| Social investigation | 121 ± 12 | 77 ± 6 ** | 132 ± 9 ## | 129 ± 12 ## |
| Unit of social investigation | 4 ± 1 | 3 ± 0.1 | 4 ± 0.4 | 4 ± 0.4 |
| Latency of social investigation | 12 ± 3 | 17 ± 3 | 15 ± 2 | 15 ± 2 |
| Social Defeat | Control (Exploration) | Acute Linalool | Chronic Linalool | Acute L. angustifolia EO | Chronic L. angustifolia EO |
|---|---|---|---|---|---|
| Intruder mice | |||||
| Avoidance | 82 ± 9 | 66 ± 6 | 57 ± 9 | 57 ± 13 | 37 ± 4 ** |
| Latency avoidance | 12 ± 5 | 6 ± 2 | 7 ± 2 | 19 ± 9 | 16 ± 4 |
| Defense/Submissive | 48 ± 9 | 51 ± 11 | 43 ± 5 | 34 ± 12 | 10 ± 4 * |
| Latency Defense/Submissive | 27 ± 9 | 11 ± 3 | 20 ± 7 | 111 ± 43 * | 21 ± 6 |
| Resident mice | |||||
| Threat | 3 ± 1 | 2 ± 0.3 | 3 ± 1 | 1 ± 0.3 | 22 ± 1 **** |
| Latency threat | 18 ± 6 | 17 ± 7 | 53 ± 30 | 28 ± 9 | 88 ± 37 |
| Attack | 33 ± 5 | 18 ± 2 ** | 25 ± 3 | 10 ± 2 **** | 6 ± 2 **** |
| Latency attack | 9 ± 5 | 15 ± 7 | 5 ± 2 | 37 ± 26 | 95 ± 36 * |
| Control (Explora) | SD Saline | SD Acute Linalool | SD Chronic Linalool | SD Acute L. angustifolia EO | SD Chronic L. angustifolia EO | |
|---|---|---|---|---|---|---|
| Time in open arms | 57 ± 6 | 21 ± 3 *** | 29 ± 6 * | 25 ± 6 ** | 45 ± 7 | 28 ± 5 * |
| % Time in open arms | 21 ± 2 | 8 ± 2 * | 12 ± 2 | 12 ± 4 | 18 ± 4 | 14 ± 2 |
| Time in central platform | 27 ± 6 | 23 ± 9 | 39 ± 12 | 50 ± 13 | 40 ±11 | 74 ± 18 * |
| Time in closed arms | 229 ± 11 | 243 ± 14 | 231 ± 15 | 237 ± 19 | 213 ±17 | 208 ± 20 |
| Entries in open arms | 22 ± 4 | 17 ± 2 | 16 ± 3 | 13 ± 3 | 23 ± 4 | 18 ± 3 |
| % Open entries | 36 ± 9 | 40 ± 4 | 36 ± 5 | 35 ± 5 | 43 ± 3 | 36 ± 4 |
| Entries in closed arms | 39 ± 6 | 31 ± 6 | 25 ± 5 | 20 ± 4 * | 31 ± 4 | 28 ± 3 |
| Total entries | 61 ± 9 | 49 ± 7 | 41 ± 8 | 33 ± 7 * | 54 ± 7 | 46 ± 5 |
| Control (Exploration) | SD Saline | SD Acute Linalool | SD Chronic Linalool | SD Acute L. angustifolia EO | SD Acute L. angustifolia EO | |
|---|---|---|---|---|---|---|
| Non-social exploration | 454 ± 11 | 511 ± 8 ** | 447 ± 12 ### | 459 ± 11 ## | 469 ± 13 # | 446 ± 12 ### |
| Explore from a distance | 2 ± 0.3 | 3 ± 0.4 | 2 ± 0.3 | 2 ± 0.3 | 2 ± 0.4 | 2 ± 1 |
| Social investigation | 110 ± 11 | 63 ± 8 ** | 126 ± 12 ### | 115 ± 10 ## | 96 ± 6 | 114 ± 11 ## |
| Unit of social investigation | 4 ± 0.4 | 2 ± 0.2 | 3 ± 0.3 | 4 ± 1 | 3 ± 0.6 | 4 ± 0.4 |
| Latency of social investigation | 8 ± 2 | 9 ± 3 | 7 ± 2 | 20 ± 5 *,# | 8 ± 2 | 15 ± 0.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caputo, L.; Reguilon, M.D.; Mińarro, J.; De Feo, V.; Rodriguez-Arias, M. Lavandula angustifolia Essential Oil and Linalool Counteract Social Aversion Induced by Social Defeat. Molecules 2018, 23, 2694. https://doi.org/10.3390/molecules23102694
Caputo L, Reguilon MD, Mińarro J, De Feo V, Rodriguez-Arias M. Lavandula angustifolia Essential Oil and Linalool Counteract Social Aversion Induced by Social Defeat. Molecules. 2018; 23(10):2694. https://doi.org/10.3390/molecules23102694
Chicago/Turabian StyleCaputo, Lucia, Marina D. Reguilon, José Mińarro, Vincenzo De Feo, and Marta Rodriguez-Arias. 2018. "Lavandula angustifolia Essential Oil and Linalool Counteract Social Aversion Induced by Social Defeat" Molecules 23, no. 10: 2694. https://doi.org/10.3390/molecules23102694
APA StyleCaputo, L., Reguilon, M. D., Mińarro, J., De Feo, V., & Rodriguez-Arias, M. (2018). Lavandula angustifolia Essential Oil and Linalool Counteract Social Aversion Induced by Social Defeat. Molecules, 23(10), 2694. https://doi.org/10.3390/molecules23102694