
Evaluation of Alkaloids Isolated from Ruta graveolens as Photosynthesis Inhibitors

 , and
, and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Effect of Alkaloids 1–8 on Noncyclic Electron Transport and H+-ATPase Activity
2.2. Uncoupled PSI and PSII Electron-Flow Determination
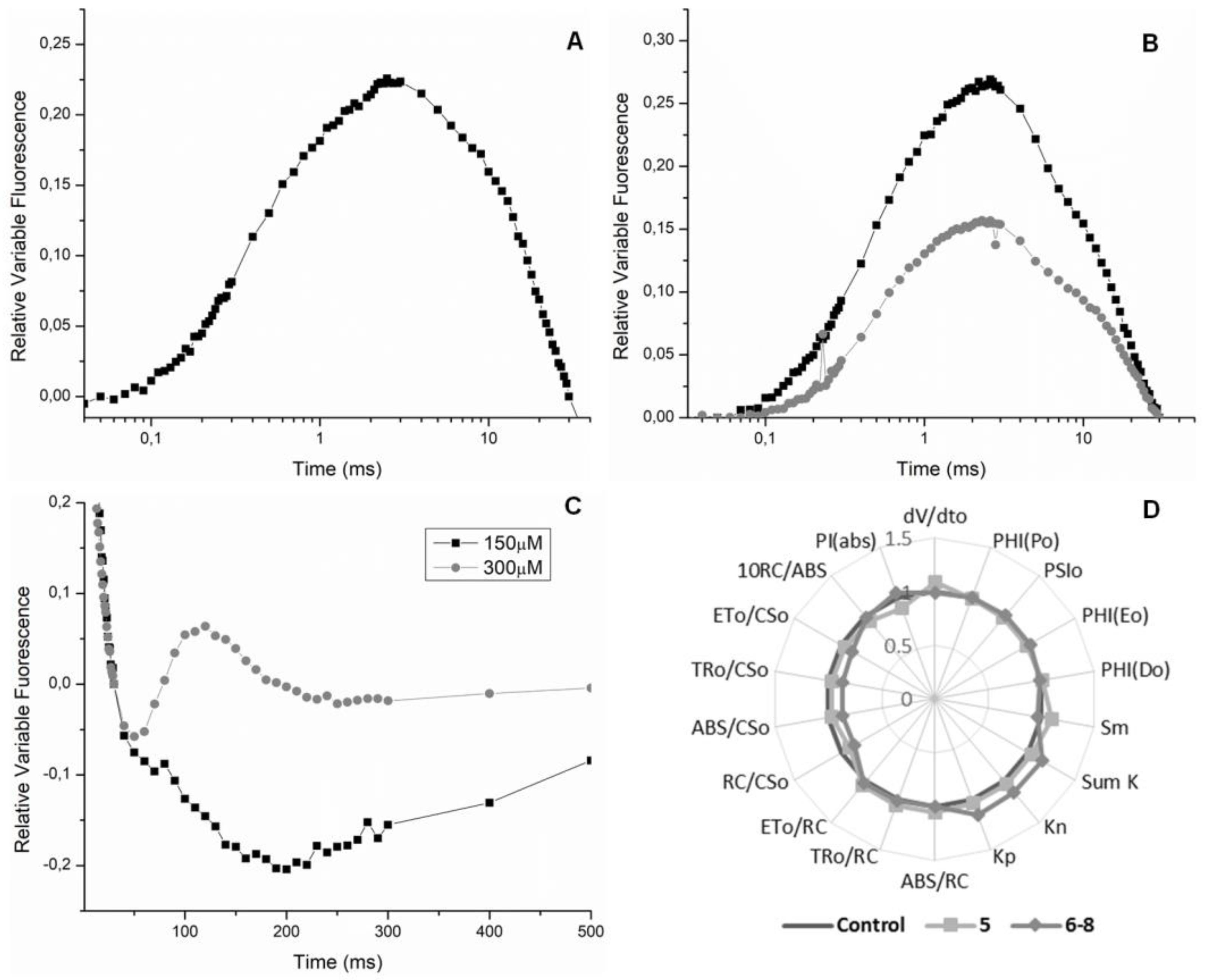
2.3. Chl a Fluorescence Measurements in Spinach Leaf Discs
2.4. In Vivo Assays: Chl a Fluorescence Determination in Intact L. Perenne Leaves
2.5. Dry Biomass Determination
3. Materials and Methods
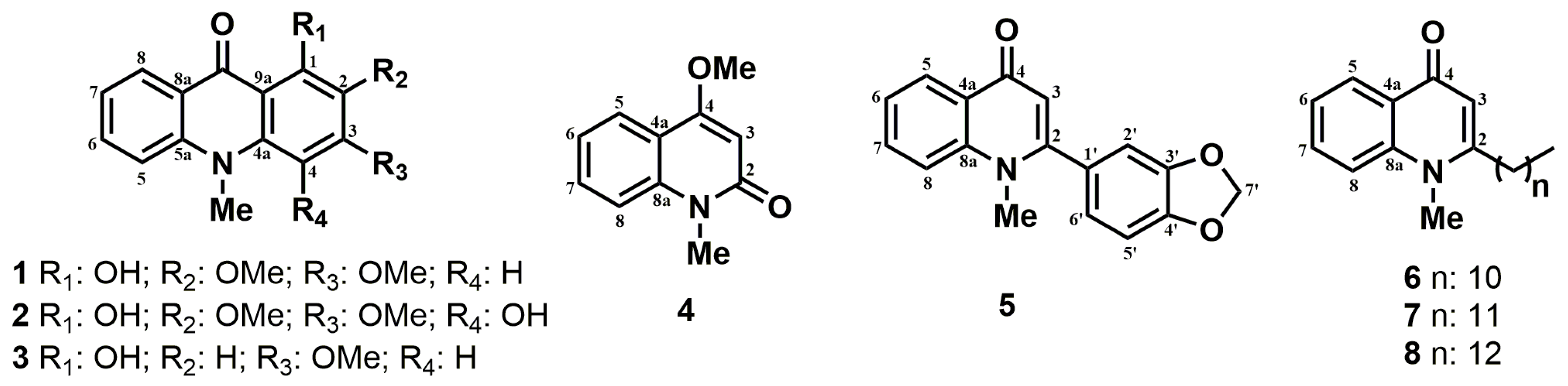
3.1. Alkaloid Isolation from Ruta Graveolens
3.2. Chloroplast Isolation and Chlorophyll Quantitative Determination
3.3. Measurement of Noncyclic Electron Transport Rate
3.4. Uncoupled PSI and PSII Electron-Flow Determination
3.5. H+-ATPase Activity Measurements
3.6. Chlorophyll A Fluorescence Measurements in Spinach-Leaf Discs
3.7. Plant Material for In Vivo Assays
3.8. Chlorophyll a Fluorescence Determination in Intact L. Perenne Leaves and Dry Biomass Determination
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP | adenosine triphosphate |
| Chl | chlorophyll |
| DBMIB | 2,5-dibromo-3-methyl-6-isopropyl-1,4-p-benzoquinone |
| DCMU | 3-(3,4-dichlorophenyl)-1,1-dimethylurea |
| DCPIP | 2,6-dichlorophenolindophenol |
| DMSO | dimethyl sulfoxide |
| MV | methylviologen |
| NADPH | dihydronicotinamide-adenine dinucleotide phosphate |
| Pi | inorganic phospate |
| PSI | photosystem I |
| PSII | photosystem II |
| RC | reaction center |
| SiMo | sodium silicomolybdate |
References
- De Feo, V.; De Simone, F.; Senatore, F. Potential allelochemicals from the essential oil of Ruta graveolens. Phytochemistry 2002, 61, 573–578. [Google Scholar] [CrossRef]
- Hale, A.L.; Meepagala, K.M.; Oliva, A.; Aliotta, G.; Duke, S.O. Phytotoxins from the leaves of Ruta graveolens. J. Agric. Food Chem. 2004, 52, 3345–3349. [Google Scholar] [CrossRef] [PubMed]
- Kuzovkina, I.; Al’terman, I.; Schneider, B. Specific accumulation and revised structures of acridone alkaloid glucosides in the tips of transformed roots of Ruta graveolens. Phytochemistry 2004, 65, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Wansi, J.D.; Wandji, J.; Mbaze Meva’a, L.; Kamdem Waffo, A.F.; Ranjit, R.; Khan, S.N.; Asma, A.; Iqbal, C.M.; Lallemand, M.-C.; Tillequin, F.; Fomum-Tanee, Z. Alpha-glucosidase inhibitory and antioxidant acridone alkaloids from the stem bark of Oriciopsis glaberrima ENGL. (Rutaceae). Chem. Pharm. Bull. 2006, 54, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Michael, J.P. Quinoline, quinazoline and acridone alkaloids. Nat. Prod. Rep. 2007, 24, 223. [Google Scholar] [CrossRef] [PubMed]
- Musiol, R.; Serda, M.; Hensel-Bielowka, S.; Polanski, J. Quinoline-Based Antifungals. Curr. Med. Chem. 2010, 17, 1960–1973. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, D.; Prado, S.; Kamoga, D.; Kasenene, J.; Bodo, B. Structure and in vitro antiparasitic activity of constituents of Citropsis articulata root bark. J. Nat. Prod. 2011, 74, 2286–2289. [Google Scholar] [CrossRef] [PubMed]
- Torres-Romero, D.; King-Díaz, B.; Strasser, R.J.; Jiménez, I.A.; Lotina-Hennsen, B.; Bazz-Cchi, I.L. Friedelane triterpenes from celastrus vulcanicola as photosynthetic inhibitors. J. Agric. Food Chem. 2010, 58, 10847–10854. [Google Scholar] [CrossRef] [PubMed]
- Menezes-De-Oliveira, D.; Aguilar, M.I.; King-Díaz, B.; Vieira-Filho, S.A.; Pains-Duarte, L.; De Fátima Silva, G.D.; Lotina-Hennsen, B. The triterpenes 3β-Lup-20(29)-en-3-ol and 3β-Lup-20(29)-en-3-yl acetate and the carbohydrate 1,2,3,4,5,6-hexa-O-acetyl-dulcitol as photosynthesis light reactions inhibitors. Molecules 2011, 16, 9939–9956. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, O.M.; de Castro Lima, M.M.; Veiga, T.A.M.; King-Díaz, B.; da Silva, M.F.d.G.F.; Lotina-Hennsen, B. Evaluation of antidesmone alkaloid as a photosynthesis inhibitor. Pestic. Biochem. Phys. 2016, 134, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Andreiadis, E.S.; Chavarot-Kerlidou, M.; Fontecave, M.; Artero, V. Artificial photosynthesis: From molecular catalysts for light-driven water splitting to photoelectrochemical cells. Photochem. Photobiol. 2011, 87, 946–964. [Google Scholar] [CrossRef] [PubMed]
- McConnell, I.; Li, G.H.; Brudvig, G.W. Energy Conversion in Natural and Artificial Photosynthesis. Chem. Biol. 2010, 17, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, F.; Yin, C.; Strasser, R.J.; Yang, C.; Qiang, S. Application of fast chlorophyll a fluorescence kinetics to probe action target of 3-acetyl-5-isopropyltetramic acid. Environ. Exp. Bot. 2011, 73, 31–41. [Google Scholar] [CrossRef]
- Strasserf, R.J.; Srivastava, A. Govindjee polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Hasan, C.; Ahmed, N.; Haque, R.; Haque, M.; Begum, S.; Sohrab, M.; Ahsan, M. Secondary metabolites from the stem of Ravenia spectabilis Lindl. Pharmacogn. Mag. 2013, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Spatafora, C.; Tringali, C. Bioactive metabolites from the bark of Fagara macrophylla. Phytochem. Anal. 1997, 8, 139–142. [Google Scholar] [CrossRef]
- Macías-Rubalcava, M.L.; García-Méndez, M.C.; King-Díaz, B.; Macías-Ruvalcaba, N.A. Effect of phytotoxic secondary metabolites and semisynthetic compounds from endophytic fungus Xylaria feejeensis strain SM3e-1b on spinach chloroplast photosynthesis. J. Photochem. Photobiol. B Biol. 2017, 166, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Terrones, M.G.; Aguilar, M.I.; King-Diaz, B.; Lotina-Hennsen, B. Interference of methyl trachyloban-19-oate ester with CF0of spinach chloroplast H+-ATPase. Arch. Biochem. Biophys. 2003, 418, 93–97. [Google Scholar] [CrossRef]
- Xiang, M.; Chen, S.; Wang, L.; Dong, Z.; Huang, J.; Zhang, Y.; Strasser, R.J. Effect of vulculic acid produced by Nimbya alternantherae on the photosynthetic apparatus of Alternanthera philoxeroides. Plant Physiol. Biochem. 2013, 65, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.; Goltsev, V. Effects of different metals on photosynthesis: Cadmium and zinc affect chlorophyll fluorescence in durum wheat. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Marchi, G.; Marchi, E.C.S.; Wang, G.; Mcgiffen, M. Effect of age of a sorghum-sudangrass hybrid on its allelopathic action. Planta Daninha 2008, 26, 707–716. [Google Scholar] [CrossRef]
- King-Díaz, B.; Dos Santos, F.J.L.; Rubinger, M.M.M.; Piló-Veloso, D.; Lotina-Hennsen, B. A diterpene γ-lactone derivative from Pterodon polygalaeflorus Benth. as a photosystem II inhibitor and uncoupler of photosynthesis. Z. Naturforsch. C 2006, 61, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Castelo-Branco, P.; Santos, F.J.L.; Rubinger, M.; Ferreira-Alves, D.; Piló-Veloso, D.; King, B.; Lotina-Hennsen, B. Inhibition and uncoupling of photosynthetic electron transport by diterpene lactone amide derivatives. Z. Naturforsch. C 2008, 63. [Google Scholar] [CrossRef]
- Min, Y.D.; Kwon, H.C.; Yang, M.C.; Lee, K.H.; Choi, S.U.; Lee, K.R. Isolation of limonoids and alkaloids from Phellodendron amurense and their multidrug resistance (MDR) reversal activity. Arch. Pharm. Res. 2007, 30, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Seya, K.; Furukawa, K.-I.; Chiyoya, M.; Yu, Z.; Kikuchi, H.; Daitoku, K.; Motomura, S.; Murakami, M.; Oshima, Y.; Fukuda, I. 1-Methyl-2-undecyl-4(1H)-quinolone, a derivative of quinolone alkaloid evocarpine, attenuates high phosphate-induced calcification of human aortic valve interstitial cells by inhibiting phosphate cotransporter PiT-1. J. Pharmacol. Sci. 2016, 131, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Yruela, I.; Montoya, G.; Alonso, P.; Picorel, R. Identification of the Pheophytin-QA-Fe domain of the reducing side of the photosystem II as the Cu(II)-inhibitory binding site. J. Biol. Chem. 1991, 266, 22847–22850. [Google Scholar] [PubMed]
- Giaquinta, R.T.; Dilley, R.A. A partial reaction in Photosystem II: Reduction of silicomolybdate prior to the site of dichlorophenyldimethylurea inhibition. Bioenergetics 1975, 387, 288–305. [Google Scholar] [CrossRef]
- Garza-Ortiz, A.; King-Díaz, B.; Sosa-Torres, M.E.; Lotina-Hennsen, B. Interference of ruthenium red analogues at photosystem II of spinach thylakoids. J. Photochem. Photobiol. B Biol. 2004, 76, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.D.; Mitchell, P.; Schürmann, P. Modulation of coupling factor ATPase activity in intact chloroplasts: The role of the thioredoxin system. FEBS Lett. 1980, 112, 173–177. [Google Scholar] [CrossRef]
- Aguilar, M.I.; Romero, M.G.; Chávez, M.I.; King-Díaz, B.; Lotina-Hennsen, B. Biflavonoids isolated from Selaginella lepidophylla inhibit photosynthesis in spinach chloroplasts. J. Agric. Food Chem. 2008, 56, 6994–7000. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | (μM) | Pi (%) |
|---|---|---|
| Control | 0 | 100 |
| 1 | 25 | 90 |
| 50 | 78 | |
| 100 | 77 | |
| 4 | 25 | 104 |
| 50 | 108 | |
| 100 | 118 | |
| 6–8 | 100 | 93 |
| 200 | 88 | |
| 300 | 77 |
| (μM) | H2O to DCBQ | H2O to Sodium Silicomolybdate (SiMo) | DPC to DCPIP | |||
|---|---|---|---|---|---|---|
| a | b | A | b | C | b | |
| 0 | 547.5 ± 2.74 | 100 | 511.0 ± 2.56 | 100 | 256.0 ± 1.28 | 100 |
| 50 | - | - | - | - | 283.0 ± 1.42 | 110.6 |
| 100 | 401.5 ± 2.00 | 74 | 328.5 ± 1.64 | 65 | 268.0 ± 1.34 | 104.5 |
| 200 | 292.0 ± 1.46 | 54 | 255.5 ± 1.28 | 50 | 268.0 ± 1.34 | 104.5 |
| 300 | 255.5 ± 1.28 | 47 | 237.3 ± 1.19 | 47 | - | - |
| 400 | 219.0 ± 1.09 | 40 | 219.0 ± 1.09 | 43 | - | - |
| (μM) | DCPIPred a Methylviologen (MV) | |
|---|---|---|
| a | b | |
| 0 | 1467.4 ± 7.34 | 100 |
| 100 | 867.1 ± 4.34 | 59.1 |
| 200 | 733.7 ± 3.67 | 50 |
| 400 | 667.0 ± 3.34 | 46 |
| Treatment | (µM) | Dry Biomass (mg) | Percentage (%) |
|---|---|---|---|
| Control | 0 | 400.0 ± 2.00 | 100 |
| DCMU | 10 | 307.0 ± 1.54 | 77 |
| 5 | 150 | 320.0 ± 1.60 | 80 |
| 300 | 327.0 ± 1.64 | 82 | |
| 6–8 | 150 | 310.0 ± 1.55 | 77 |
| 300 | 350.0 ± 1.75 | 87 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sampaio, O.M.; Vieira, L.C.C.; Bellete, B.S.; King-Diaz, B.; Lotina-Hennsen, B.; Da Silva, M.F.d.G.F.; Veiga, T.A.M. Evaluation of Alkaloids Isolated from Ruta graveolens as Photosynthesis Inhibitors. Molecules 2018, 23, 2693. https://doi.org/10.3390/molecules23102693
Sampaio OM, Vieira LCC, Bellete BS, King-Diaz B, Lotina-Hennsen B, Da Silva MFdGF, Veiga TAM. Evaluation of Alkaloids Isolated from Ruta graveolens as Photosynthesis Inhibitors. Molecules. 2018; 23(10):2693. https://doi.org/10.3390/molecules23102693
Chicago/Turabian StyleSampaio, Olívia Moreira, Lucas Campos Curcino Vieira, Barbara Sayuri Bellete, Beatriz King-Diaz, Blas Lotina-Hennsen, Maria Fátima das Graças Fernandes Da Silva, and Thiago André Moura Veiga. 2018. "Evaluation of Alkaloids Isolated from Ruta graveolens as Photosynthesis Inhibitors" Molecules 23, no. 10: 2693. https://doi.org/10.3390/molecules23102693
APA StyleSampaio, O. M., Vieira, L. C. C., Bellete, B. S., King-Diaz, B., Lotina-Hennsen, B., Da Silva, M. F. d. G. F., & Veiga, T. A. M. (2018). Evaluation of Alkaloids Isolated from Ruta graveolens as Photosynthesis Inhibitors. Molecules, 23(10), 2693. https://doi.org/10.3390/molecules23102693