Bioformation of Volatile and Nonvolatile Metabolites by Saccharomycopsis fibuligera KJJ81 Cultivated under Different Conditions—Carbon Sources and Cultivation Times



Abstract
1. Introduction
2. Results and Discussion
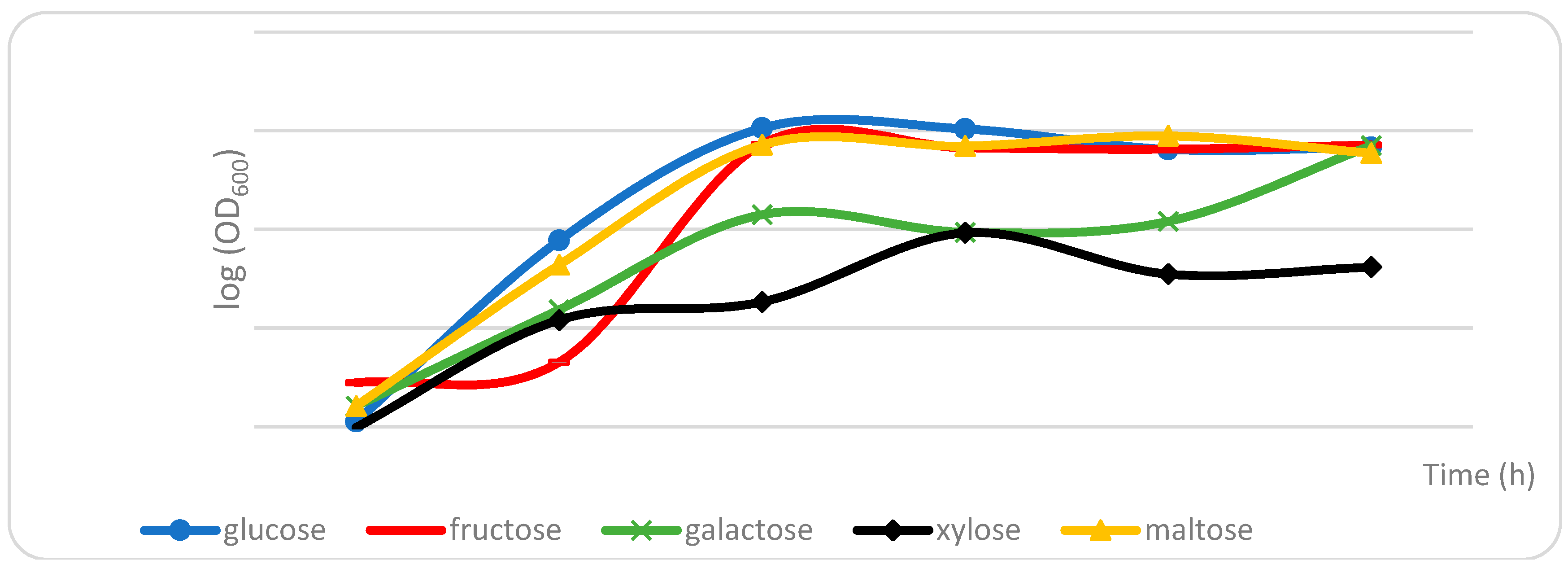
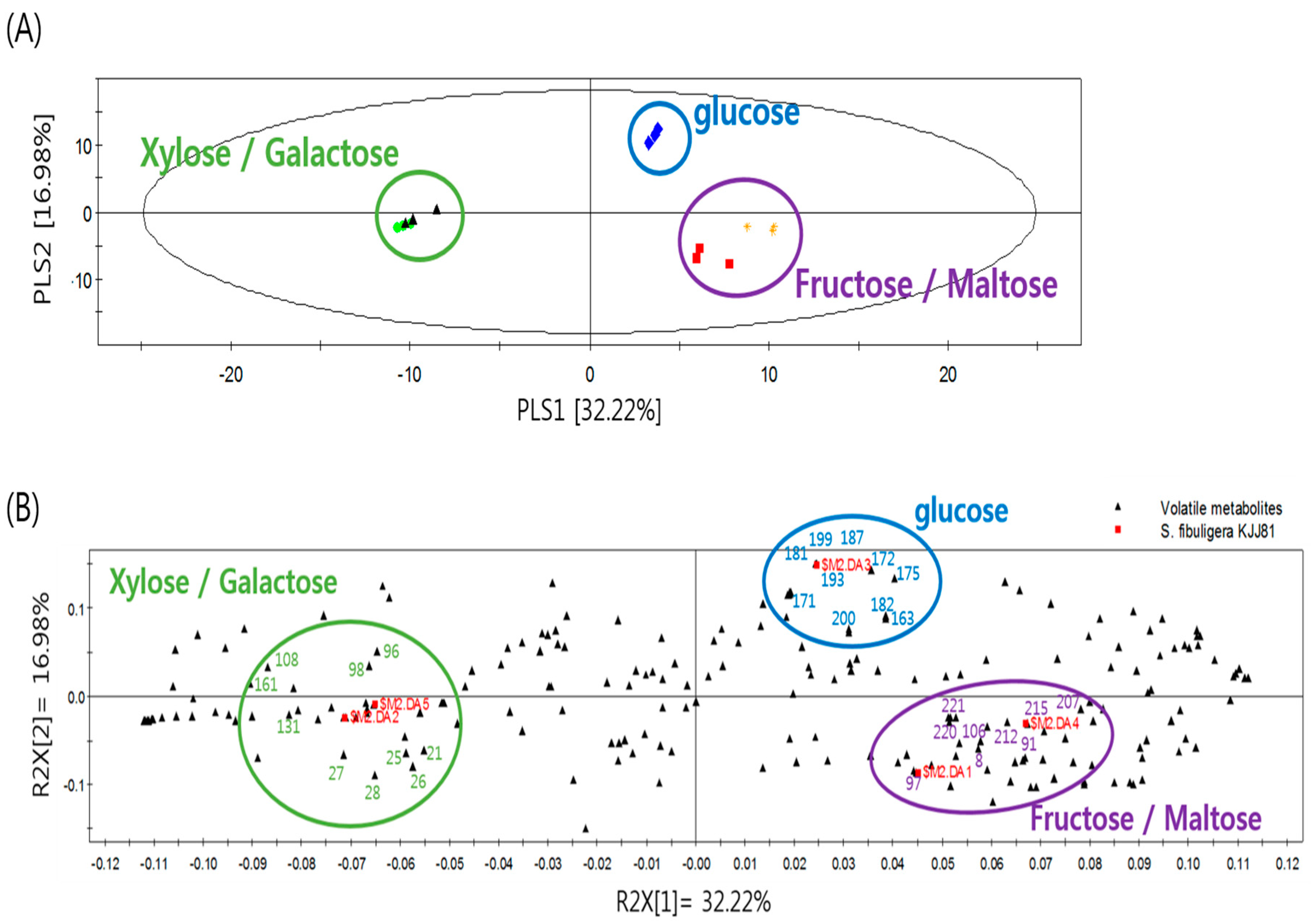
2.1. Effects of Carbon Sources on the Formation of Volatile Metabolites by S. fibuligera KJJ81
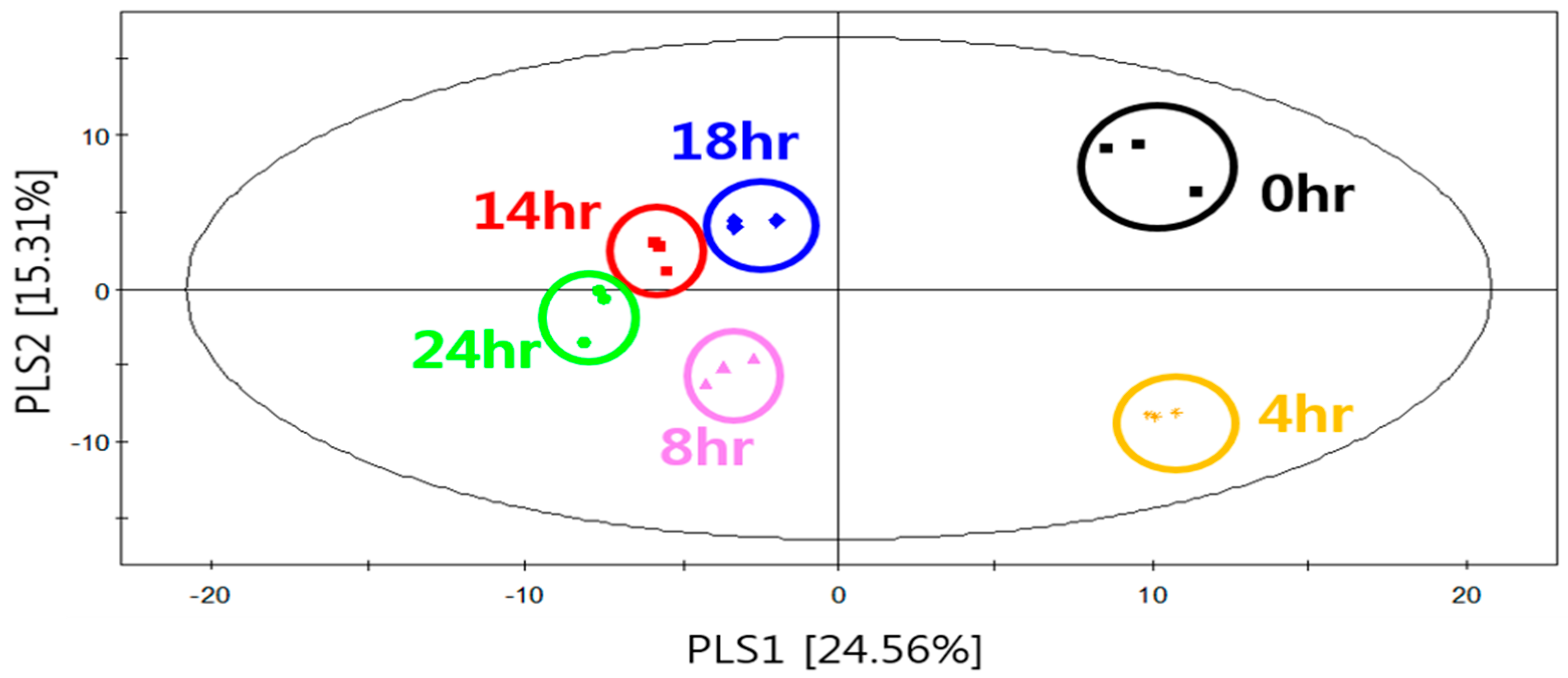
2.2. Cultivation-Time-Dependent Volatile Metabolites of S. fibuligera KJJ81 in YPD Medium
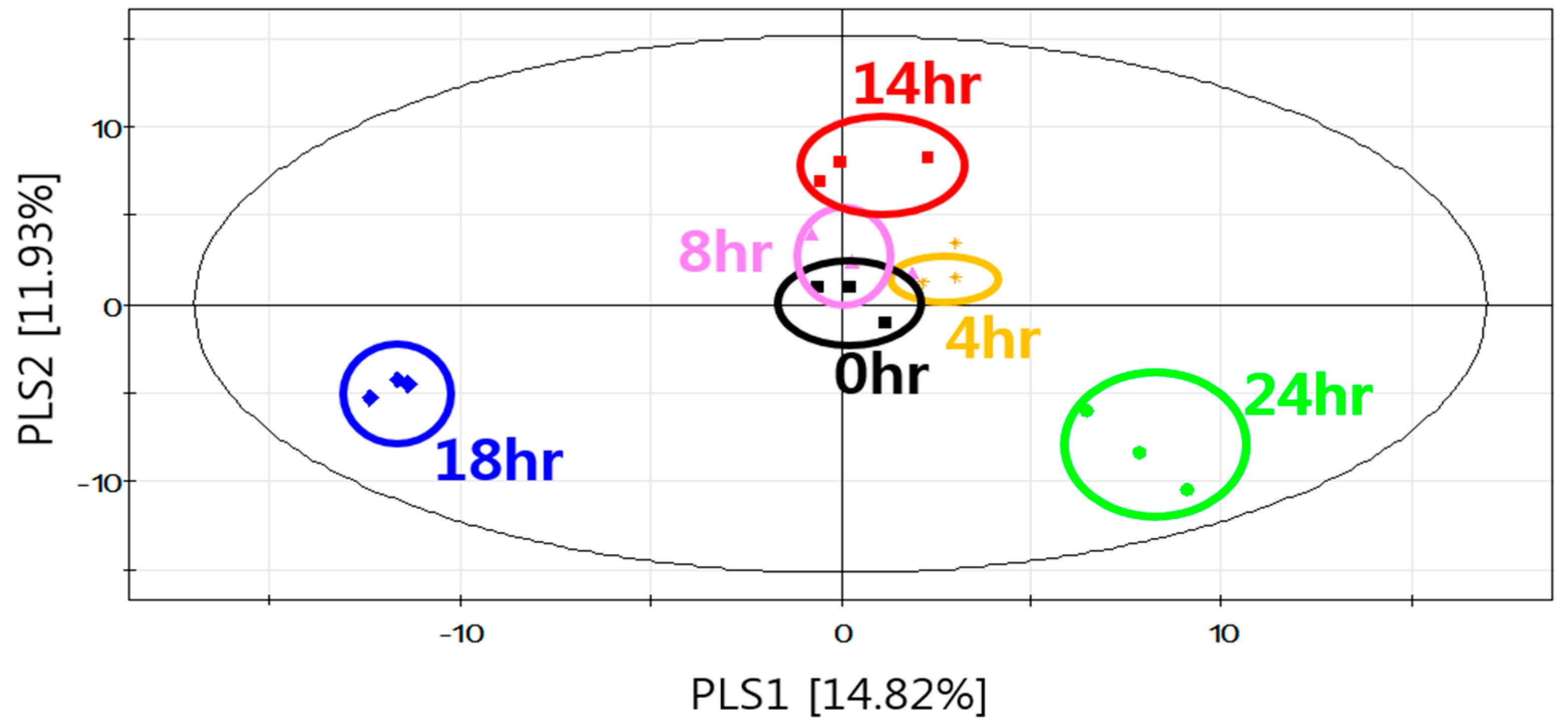
2.3. Cultivation-time-dependent Nonvolatile Metabolites of S. fibuligera KJJ81 in YPD Medium
3. Materials and Methods
3.1. Cultivation of S. fibuligera
3.2. Extraction and Analysis of Volatile Metabolites by GC-MS
3.3. Extraction and Analysis of Nonvolatile Metabolites by GC-TOF/MS
3.4. Mass Spectral Data Processing of Volatile and Nonvolatile Metabolites
3.5. Identification and Quantification of Volatile and Nonvolatile Metabolites
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Carroll, E.; Trinh, T.N.; Son, H.; Lee, Y.W.; Seo, J.A. Comprehensive analysis of fungal diversity and enzyme activity in nuruk, a Korean fermenting starter, for acquiring useful fungi. J. Microbiol. 2017, 55, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Lee, S.J.; Lim, J.H.; Kim, B.K.; Park, K.J. Chemical and sensory profiles of makgeolli, Korean commercial rice wine, from descriptive, chemical, and volatile compound analyses. Food Chem. 2014, 152, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Nam, Y.; Roh, S.W.; Bae, J. Unexpected convergence of fungal and bacterial communities during fermentation of traditional Korean alcoholic beverages inoculated with various natural starters. Food Microbiol. 2012, 30, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Kim, J.H.; Bai, D.H.; Ahn, B.H. Identification and characterization of useful fungi with α-amylase activity from the Korean traditional nuruk. Mycobiology 2011, 39, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, S.M.; Song, S.H.; Kim, Y.S. Characterization of volatile components in Makgeolli, a traditional Korean rice wine, with or without pasteurization, during storage. Molecules 2013, 18, 5317–5325. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.S.; Lee, J.E.; Park, H.J. Electronic tongue-based discrimination of Korean rice wines (makgeolli) including prediction of sensory evaluation and instrumental measurements. Food Chem. 2014, 151, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Son, E.Y.; Lee, S.M.; Kim, M.; Seo, J.A.; Kim, Y.S. Comparison of volatile and non-volatile metabolites in rice wine fermented by Koji inoculated with Saccharomycopsis fibuligera and Aspergillus oryzae. Food Res. Int. 2018, 109, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Park, E.H.; Kim, M.D. Characterization of starch-utilizing yeast Saccharomycopsis fibuligera isolated from Nuruk. J. Microbiol. Biotechnol. 2014, 42, 407–412. [Google Scholar] [CrossRef]
- Chi, Z.; Chi, Z.; Liu, G.; Wang, F.; Ju, L.; Zhang, T. Saccharomycopsis fibuligera and its applications in biotechnology. Biotechnol. Adv. 2009, 27, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Hostinová, E. Amylolytic enzymes produced by the yeast Saccharomycopsis fibuligera. Biologia 2002, 57, 247–252. [Google Scholar]
- Aidoo, K.E.; Nout, M.R.; Sarkar, P.K. Occurrence and function of yeasts in Asian indigenous fermented foods. FEMS Yeast Res. 2006, 6, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Liu, J.; Zhang, W. Trehalose accumulation from soluble starch by Saccharomycopsis fibuligera sdu. Enzyme Microb. Technol. 2001, 28, 240–245. [Google Scholar] [CrossRef]
- Lenz, E.M.; Wilson, I.D. Analytical Strategies in Metabonomics. J. Proteome Res. 2007, 6, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.Z.; Cheng, J.S.; Xiao, W.H. Comparative metabolomic analysis on industrial continuous and batch ethanol fermentation processes by GC–TOF-MS. Metabolomics 2009, 5, 229–238. [Google Scholar] [CrossRef]
- Jonsson, P.; Johansson, A.I.; Gullberg, J. High-throughput data analysis for detecting and identifying differences between samples in GC/MS-based metabolomic analyses. Anal. Chem. 2005, 77, 5635–5642. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, M.; Ugliano, M.; Varela, C.; Siebert, T.; Pretorius, I.S.; Henschke, P.A. Assimilable nitrogen utilisation and production of volatile and non-volatile compounds in chemically defined medium by Saccharomyces cerevisiae wine yeasts. Appl. Microbiol. Biotechnol. 2007, 77, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Soares, E.V.; Vroman, A.; Mortier, J.; Rijsbrack, K.; Mota, M. Carbohydrate carbon sources induce loss of flocculation of an ale-brewing yeast strain. J. Appl. Microbiol. 2004, 96, 1117–1123. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mao, X.; Eksriwong, T.; Chauvatcharin, S.; Zhong, J. Optimization of carbon source and carbon/nitrogen ratio for cordycepin production by submerged cultivation of medicinal mushroom Cordyceps militaris. Process Biochem. 2005, 40, 1667–1672. [Google Scholar] [CrossRef]
- Adams, T.H.; Yu, J.H. Coordinate control of secondary metabolite production and asexual sporulation in Aspergillus nidulans. Curr. Opin. Microbiol. 1998, 1, 674–677. [Google Scholar] [CrossRef]
- Connor, M.R.; Liao, J.C. Engineering of an Escherichia coli strain for the production of 3-methyl-1-butanol. Appl. Environ. Microbiol. 2008, 74, 5769–5775. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Vadlani, P.; Harrison, M.; Bennett, G.; San, K. Aerobic production of isoamyl acetate by overexpression of the yeast alcohol acetyl-transferases AFT1 and AFT2 in Escherichia coli and using low-cost fermentation ingredients. Bioprocess Biosyst. Eng. 2008, 31, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Schoondermark-Stolk, S.A.; Jansen, M.; Veurink, J.H.; Verkleij, A.J.; Verrips, C.T.; Euverink, G.W.; Boonstra, J.; Dijkhuizen, L. Rapid identification of target genes for 3-methyl-1-butanol production in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2006, 70, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Plata, C.; Mauricio, J.C.; Millan, C.; Ortega, J.M. Influence of glucose and oxygen on the production of ethyl acetate and isoamyl acetate by a Saccharomyces cerevisiae strain during alcoholic fermentation. World J. Microbiol. Biotechnol. 2005, 21, 115–121. [Google Scholar] [CrossRef]
- Assaf, S.; Hadar, Y.; Dosoretz, C.G. 1-Octen-3-ol and 13-hydroperoxylinoleate are products of distinct pathways in the oxidative breakdown of linoleic acid by Pleurotus pulmonarius. Enzyme Microb. Technol. 1997, 21, 484–490. [Google Scholar] [CrossRef]
- Pagot, Y.; Le Clainche, A.; Nicaud, J.; Wache, Y.; Belin, J. Peroxisomal β-oxidation activities and γ-decalactone production by the yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 1998, 49, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Romero-Guido, C.; Belo, I.; Ta, T.M.N.; Cao-Hoang, L.; Alchihab, M.; Gomes, N.; Wache, Y. Biochemistry of lactone formation in yeast and fungi and its utilization for the production of flavor and fragrance compounds. Appl. Microbiol. Biotechnol. 2011, 89, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Ravasio, D.; Wendland, J.; Walther, A. Major contribution of the Ehrlich pathway for 2-phenylethanol/rose flavor production in Ashbya gossypii. FEMS Yeast Res. 2014, 14, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Viana, F.; Belloch, C.; Vallés, S.; Manzanares, P. Monitoring a mixed starter of Hanseniaspora vineae–Saccharomyces cerevisiae in natural must: Impact on 2-phenylethyl acetate production. Int. J. Food Microbiol. 2011, 151, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xu, Y. The influence of yeast strains on the volatile flavour compounds of Chinese rice wine. J. Inst. Brew. 2010, 116, 190–196. [Google Scholar] [CrossRef]
- Saerens, S.M.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Xu, E.; Long, J.; Wu, Z.; Li, H.; Wang, F.; Xu, X.; Jiao, A. Characterization of volatile flavor compounds in Chinese rice wine fermented from enzymatic extruded rice. J. Food Sci. 2015, 80, C1476–C1489. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.A.; Pastore, G.M.; Park, Y.K. Microbial transformation of sucrose and glucose to erythritol. Biotechnol. Lett. 1993, 15, 383–388. [Google Scholar] [CrossRef]
- Tomaszewska, L.; Rywińska, A.; Gładkowski, W. Production of erythritol and mannitol by Yarrowia lipolytica yeast in media containing glycerol. J. Ind. Microbiol. Biotechnol. 2012, 39, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- Onishi, H.; Suzuki, T. Production of D-mannitol and glycerol by yeasts. Appl. Microbiol. 1968, 16, 1847–1852. [Google Scholar] [PubMed]
- Lee, D.E.; Lee, S.; Jang, E.S.; Shin, H.W.; Moon, B.S.; Lee, C.H. Metabolomic profiles of Aspergillus oryzae and Bacillus amyloliquefaciens during Rice Koji fermentation. Molecules 2016, 21, 773. [Google Scholar] [CrossRef] [PubMed]
- Lomakin, I.B.; Xiong, Y.; Steitz, T.A. The crystal structure of yeast fatty acid synthase, a cellular machine with eight active sites working together. Cell 2007, 129, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Dai, X.; Zhu, J.; Yu, H. Classification of Chinese Rice wine according to geographic origin and wine age based on chemometric methods and SBSE-TD-GC-MS analysis of volatile compounds. Food Sci. Technol. Res. 2015, 21, 371–380. [Google Scholar] [CrossRef]
- Foster, J.; Nakata, P.A. An oxalyl-CoA synthetase is important for oxalate metabolism in Saccharomyces cerevisiae. FEBS Lett. 2014, 588, 160–166. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Dong, J.; Yin, H.; Zhao, Y.; Chen, R.; Wan, X.; Chen, P.; Hou, X.; Liu, J.; Chen, L. Wort composition and its impact on the flavour-active higher alcohol and ester formation of beer—A review. J. Inst. Brew. 2014, 120, 157–163. [Google Scholar] [CrossRef]
- Swiegers, J.; Bartowsky, E.; Henschke, P.; Pretorius, I. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.M.; van Maris, A.J.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Choo, J.H.; Hong, C.P.; Lim, J.Y.; Seo, J.A.; Kim, Y.S.; Lee, D.W.; Kang, H.A. Whole-genome de novo sequencing, combined with RNA-Seq analysis, reveals unique genome and physiological features of the amylolytic yeast Saccharomycopsis fibuligera and its interspecies hybrid. Biotechnol. Biofuels 2016, 9, 246–268. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, D.Y.; Wohlgemuth, G.; Park, H.S.; Fiehn, O.; Kim, K.H. Evaluation and optimization of metabolome sample preparation methods for Saccharomyces cerevisiae. Anal. Chem. 2013, 85, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RI 1 | Major Volatile Metabolites 2 | VIP 3 | ID 4 |
|---|---|---|---|---|
| Media Containing Xylose and Galactose | ||||
| Alcohols | ||||
| 98 | 1560 | 1-Octanol | 0.80 | A |
| Benzenes & Benzene derivatives | ||||
| 96 | 1429 | 1,3-Bis(2-methyl-2-propanyl)benzene | 1.11 | B |
| Esters | ||||
| 161 | 2261 | (2E,6E)-3,7,11-Trimethyl-2,6,10-dodecatrien-1-yl acetate | 0.79 | B |
| 131 | 1680 | 4-(2-Methyl-2-propanyl)cyclohexyl acetate | 0.74 | B |
| Furans | ||||
| 108 | 2394 | 5-[(2E)-2-Octen-1-yl]dihydro-2(3H)-furanone | 0.74 | B |
| Media Containing Glucose | ||||
| Alcohols | ||||
| 187 | 1910 | 2-Phenylethanol | 1.42 | A |
| 175 | 1709 | (2E)-3,7-Dimethyl-2,6-octadien-1-ol | 1.25 | B |
| Aldehydes | ||||
| 181 | 1750 | 3-Dodecenal | 1.42 | B |
| 199 | 1287 | Octanal | 1.42 | B |
| 172 | 1429 | 2-Octenal | 1.33 | B |
| Esters | ||||
| 183 | 1813 | 2-Phenylethyl acetate | 1.12 | A |
| 182 | 1782 | Ethyl phenylacetate | 0.90 | A |
| 163 | <1100 | Ethyl acetate | 0.87 | A |
| Lactones | ||||
| 193 | 2424 | 6-Heptyltetrahydro-2H-pyran-2-one | 1.43 | A |
| Media Containing Fructose and Maltose | ||||
| Alcohols | ||||
| 97 | 1452 | 1-Octen-3-ol | 1.59 | A |
| 212 | 1659 | 2-Furylmethanol | 1.51 | A |
| 106 | 2003 | Phenol | 1.25 | A |
| 91 | 1218 | 3-Methyl-1-butanol | 0.86 | A |
| Esters | ||||
| 220 | 1128 | 3-Methylbutyl acetate | 1.26 | A |
| Furans | ||||
| 83 | 2375 | 5-Hexyldihydro-2(3H)-furanone | 0.87 | A |
| Sulfur-containing compounds | ||||
| 8 | 1059 | (Methyldisulfanyl)methane | 0.84 | A |
| VIP 1 | Volatile Metabolites 2 | RI 3 | Relative Peak Area (%) 4 | |||||
|---|---|---|---|---|---|---|---|---|
| 0 h 5 | 4 h | 8 h | 14 h | 18 h | 24 h | |||
| Acids | ||||||||
| 1.17 | Butyric acid | 1625 | N.D. 6 a 7 | 0.02 ± 0.01 a | 0.09 ± 0.04 ab | 0.27 ± 0.02 d | 0.14 ± 0.03 bc | 0.24 ± 0.14 cd |
| 1.36 | 2-Methylpropanoic acid | 1568 | N.D. a | N.D. a | 0.01 ± 0.00 a | 0.02 ± 0.00 b | N.D. a | 0.04 ± 0.01 c |
| Alcohols | ||||||||
| 1.10 | 2-Methyl-1-propanol | 1113 | N.D. a | 0.04 ± 0.01 a | 0.43 ± 0.06 b | 0.59 ± 0.07 c | 0.44 ± 0.01 b | 0.46 ± 0.00 b |
| 1.17 | (2E)-3,7-Dimethyl-2,6-octadien-1-ol | 1847 | N.D. a | N.D. a | 0.01 ± 0.01 a | 0.04 ± 0.00 b | 0.03 ± 0.00 b | 0.03 ± 0.01 b |
| 1.17 | Phenol | 1997 | 0.03 ± 0.01 bc | 0.01 ± 0.00 a | N.D. a | 0.03 ± 0.01 bc | 0.02 ± 0.00 b | 0.04 ± 0.01 c |
| 1.19 | 1-Butanol | 1160 | 2.22 ± 0.93 b | 0.91 ± 0.04 a | 0.32 ± 0.03 a | 0.41 ± 0.06 a | 0.25 ± 0.01 a | 0.32 ± 0.02 a |
| 1.27 | 2,4-Bis(2-methyl-2-propanyl)phenol | 2307 | 0.18 ± 0.01 d | 0.13 ± 0.01 ab | 0.14 ± 0.01 bc | 0.15 ± 0.01 bc | 0.12 ± 0.00 a | 0.16 ± 0.02 c |
| 1.29 | 3-Methyl-1-butanol | 1220 | 0.18 ± 0.05 a | 2.24 ± 0.12 b | 4.25 ± 0.21 c | 5.02 ± 0.48 d | 4.67 ± 0.26 cd | 4.85 ± 0.28 d |
| 1.40 | 2-Phenylethanol | 1908 | 0.21 ± 0.04 a | 2.88 ± 0.12 b | 4.63 ± 0.35 c | 5.40 ± 0.40 d | 5.38 ± 0.32 d | 6.18 ± 0.41 e |
| Aldehydes | ||||||||
| 1.23 | 2-Furaldehyde | 1458 | 0.04 ± 0.01 a | 0.01 ± 0.00 a | 0.02 ± 0.00 a | 0.07 ± 0.02 bc | 0.10 ± 0.05 c | 0.05 ± 0.01 ab |
| 1.47 | Octanal | 1287 | N.D. a | 0.01 ± 0.00 b | N.D. a | 0.06 ± 0.01 d | 0.07 ± 0.0037 c | 0.03 ± 0.00 c |
| Benzen and benzene derivatives | ||||||||
| 1.25 | Benzaldehyde | 1518 | 0.16 ± 0.05 b | 0.04 ± 0.01 a | 0.05 ± 0.05 a | 0.03 ± 0.00 a | 0.03 ± 0.00 a | 0.03 ± 0.00 a |
| 1.31 | 1-(2-Aminophenyl)ethanone | 2210 | 0.01 ± 0.00 a | 0.02 ± 0.00 b | 0.02 ± 0.00 b | 0.02 ± 0.00 ab | 0.02 ± 0.00 a | 0.02 ± 0.00 a |
| 1.35 | 3-Methylbenzaldehyde | 1642 | 0.05 ± 0.02 c | N.D. a | N.D. a | 0.06 ± 0.01 cd | 0.03 ± 0.00 b | 0.07 ± 0.01 d |
| Esters | ||||||||
| 1.11 | Ethyl butyrate | <1100 | N.D. a | N.D. a | 0.44 ± 0.07 b | 1.15 ± 0.13 d | 0.91 ± 0.05 c | 0.79 ± 0.08 c |
| 1.14 | (2E)-3,7-Dimethyl-2,6-octadien-1-yl acetate | 1708 | N.D. a | N.D. a | 0.03 ± 0.00 c | 0.03 ± 0.01 c | 0.03 ± 0.00 c | 0.02 ± 0.00 b |
| 1.15 | 3-Methylbutyl acetate | 1130 | 0.08 ± 0.01 a | 0.30 ± 0.01 a | 2.29 ± 0.16 c | 4.14 ± 0.36 e | 3.48 ± 0.17 d | 1.59 ± 0.06 b |
| 1.17 | 2-Phenylethyl acetate | 1812 | 0.05 ± 0.01 a | 0.25 ± 0.01 b | 1.27 ± 0.12 c | 1.63 ± 0.14 d | 1.27 ± 0.08 c | 1.56 ± 0.08 d |
| 1.27 | (2E,6E)-3,7,11-Trimethyl-2,6,10-dodecatrien-1-yl acetate | 2260 | 0.02 ± 0.01 a | 0.02 ± 0.00 a | 0.09 ± 0.01 c | 0.07 ± 0.02 b | 0.09 ± 0.01 c | 0.07 ± 0.00 b |
| 1.37 | Ethyl 3-phenylacrylate | 2125 | N.D. a | 0.16 ± 0.01 b | 0.35 ± 0.01 d | 0.47 ± 0.05 e | 0.22 ± 0.01 c | 0.33 ± 0.04 d |
| 1.45 | Butyl 3-phenylacrylate | 2333 | N.D. a | 0.07 ± 0.01 b | 0.09 ± 0.01 b | 0.09 ± 0.01 b | 0.08 ± 0.01 b | 0.07 ± 0.02 b |
| Hydrocarbons | ||||||||
| 1.18 | Hexadecane | 1598 | 0.02 ± 0.00 ab | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.02 ± 0.00 bc | 0.02 ± 0.01 c | 0.02 ± 0.00 bc |
| Lactones | ||||||||
| 1.21 | 6-Heptyltetrahydro-2H-pyran-2-one | 2423 | N.D. a | 0.02 ± 0.00 b | 0.06 ± 0.00 c | 0.14 ± 0.02 e | 0.09 ± 0.01 d | 0.10 ± 0.01 d |
| 1.22 | 6-Pentyltetrahydro-2H-pyran-2-one | 2189 | N.D. a | 0.02 ± 0.00 b | 0.05 ± 0.00 c | 0.06 ± 0.01 c | 0.06 ± 0.00 c | 0.06 ± 0.00 c |
| 1.25 | 5-Hexyldihydro-2(3H)-furanone | 2139 | 0.05 ± 0.00 a | 0.27 ± 0.03 b | 0.66 ± 0.01 cd | 0.74 ± 0.09 d | 0.61 ± 0.02 c | 0.65 ± 0.06 cd |
| 1.27 | 5-[(2Z)-2-Octen-1-yl]dihydro-2(3H)-furanone | 2393 | N.D. a | 0.03 ± 0.01 b | 0.16 ± 0.01 c | 0.23 ± 0.03 d | 0.21 ± 0.01 d | 0.17 ± 0.02 c |
| 1.41 | 5-Pentyldihydro-2(3H)-furanone | 2023 | 0.01 ± 0.00 a | 0.07 ± 0.00 b | 0.08 ± 0.02 bc | 0.10 ± 0.02 c | 0.08 ± 0.00 b | 0.09 ± 0.01 bc |
| Pyrazines | ||||||||
| 1.16 | 2-Isopropyl-5-methylpyrazine | 1413 | 0.01 ± 0.00 a | 0.02 ± 0.00 b | 0.01 ± 0.01 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a |
| 1.17 | 3-Ethyl-2,5-dimethylpyrazine | 1446 | 0.03 ± 0.01 a | 0.03 ± 0.00 a | 0.04 ± 0.00 b | 0.03 ± 0.01 a | 0.03 ± 0.00 a | 0.04 ± 0.00 b |
| 1.21 | 2-Butyl-3,5-dimethylpyrazine | 1610 | 0.02 ± 0.00 a | 0.02 ± 0.00 a | 0.02 ± 0.00 a | 0.03 ± 0.00 c | 0.03 ± 0.01 c | 0.02 ± 0.00 b |
| 1.22 | 2,5-Dimethylpyrazine | 1320 | 0.08 ± 0.01 ab | 0.11 ± 0.01 c | 0.09 ± 0.01 b | 0.07 ± 0.01 a | 0.07 ± 0.01 a | 0.07 ± 0.01 a |
| 1.25 | 3-Butyl-2,5-dimethylpyrazine | 1658 | N.D. a | N.D. a | 0.01 ± 0.00 a | 0.10 ± 0.05 b | 0.01 ± 0.00 a | 0.01 ± 0.00 a |
| VIP 1 | Non-Volatile Metabolite 2 | MS 3 | Relative Peak Area (%) 4 | |||||
|---|---|---|---|---|---|---|---|---|
| 0 h 5 | 4 h | 8 h | 14 h | 18 h | 24 h | |||
| Carbohydrates | ||||||||
| 1.46 | Mannose | 147 | 0.56 ± 0.03 d 6 | 0.11 ± 0.09 b | 0.01 ± 0.00 a | 0.02 ± 0.00 a | 0.09 ± 0.01 b | 0.48 ± 0.01 c |
| 1.19 | Arabitol | 217 | 0.35 ± 0.31 b | 1.07 ± 0.84 c | 0.24 ± 0.02 a | 0.04 ± 0.00 a | 0.01 ± 0.00 a | 0.28 ± 0.01 a |
| 0.73 | Mannitol | 147 | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.01 ± 0.00 a | N.D. 7 a | 0.03 ± 0.00 b |
| Lipids | ||||||||
| 1.87 | Propionic acid | 147 | 0.22 ± 0.01 c | 0.09 ± 0.01 b | 0.23 ± 0.04 c | 0.01 ± 0.00 a | 0.50 ± 0.04 d | 0.16 ± 0.01 b |
| 1.37 | Palmitic acid | 117 | 3.90 ± 0.22 b | 4.87 ± 0.16 c | 3.75 ± 0.40 b | 3.06 ± 0.34 a | 3.74 ± 0.43 b | 3.03 ± 0.37 a |
| 1.30 | Stearic acid | 117 | 4.19 ± 0.23 b | 5.26 ± 0.14 c | 4.19 ± 0.38 b | 3.38 ± 0.43 a | 4.26 ± 0.50 b | 3.31 ± 0.39 a |
| 1.01 | Myristic acid | 117 | 0.12 ± 0.01 ab | 0.18 ± 0.03 d | 0.14 ± 0.01 bc | 0.14 ± 0.01 bc | 0.10 ± 0.01 a | 0.11 ± 0.02 ab |
| Organic acids | ||||||||
| 1.53 | Oxalic acid1 | 147 | 0.01 ± 0.00 a | 1.19 ± 2.02 b | 1.76 ± 2.99 c | 0.01 ± 0.01 a | N.D. a | 0.01 ± 0.00 a |
| 1.48 | Oxalic acid2 | 147 | 9.95 ± 1.78 b | 5.30 ± 1.70 a | 5.12 ± 1.87 a | 28.11 ± 2.14 d | 10.22 ± 1.66 b | 18.37 ± 1.69 c |
| 0.92 | Succinic acid | 147 | 0.01 ± 0.01 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.02 ± 0.03 a | N.D. a | 0.03 ± 0.06 a |
| Amino acids | ||||||||
| 1.40 | Isoleucine | 158 | 0.01 ± 0.00 a | 0.01 ± 0.001 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.03 ± 0.00 b | 0.15 ± 0.00 c |
| 1.38 | Serine | 204 | 0.01 ± 0.00 b | 0.06 ± 0.00 e | 0.01 ± 0.00 b | 0.03 ± 0.00 c | 0.00 ± 0.00 a | 0.05 ± 0.00 d |
| 1.20 | Alanine | 116 | 0.27 ± 0.01 c | 0.40 ± 0.03 d | 0.17 ± 0.07 b | 0.30 ± 0.03 c | 0.01 ± 0.00 a | 0.26 ± 0.02 c |
| 1.03 | Glutamic acid | 246 | 0.01 ± 0.00 a | 0.10 ± 0.01 b | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a |
| 0.96 | Glycine | 102 | 0.05 ± 0.01 b | 0.15 ± 0.03 c | 0.02 ± 0.02 ab | 0.04 ± 0.01 b | N.D. a | 0.05 ± 0.01 b |
| 0.96 | Proline | 142 | 0.20 ± 0.00 c | 0.04 ± 0.03 ab | 0.08 ± 0.07 b | 0.24 ± 0.01 c | N.D. a | 0.25 ± 0.01 c |
| 0.85 | Phenylalanine | 218 | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.11 ± 0.01 b |
| 0.81 | Threonine | 117 | 0.01 ± 0.00 b | 0.03 ± 0.00 c | 0.01 ± 0.00 ab | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.03 ± 0.00 b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.M.; Jung, J.H.; Seo, J.-A.; Kim, Y.-S. Bioformation of Volatile and Nonvolatile Metabolites by Saccharomycopsis fibuligera KJJ81 Cultivated under Different Conditions—Carbon Sources and Cultivation Times. Molecules 2018, 23, 2762. https://doi.org/10.3390/molecules23112762
Lee SM, Jung JH, Seo J-A, Kim Y-S. Bioformation of Volatile and Nonvolatile Metabolites by Saccharomycopsis fibuligera KJJ81 Cultivated under Different Conditions—Carbon Sources and Cultivation Times. Molecules. 2018; 23(11):2762. https://doi.org/10.3390/molecules23112762
Chicago/Turabian StyleLee, Sang Mi, Ji Hye Jung, Jeong-Ah Seo, and Young-Suk Kim. 2018. "Bioformation of Volatile and Nonvolatile Metabolites by Saccharomycopsis fibuligera KJJ81 Cultivated under Different Conditions—Carbon Sources and Cultivation Times" Molecules 23, no. 11: 2762. https://doi.org/10.3390/molecules23112762
APA StyleLee, S. M., Jung, J. H., Seo, J.-A., & Kim, Y.-S. (2018). Bioformation of Volatile and Nonvolatile Metabolites by Saccharomycopsis fibuligera KJJ81 Cultivated under Different Conditions—Carbon Sources and Cultivation Times. Molecules, 23(11), 2762. https://doi.org/10.3390/molecules23112762