Evaluating Marine Cyanobacteria as a Source for CNS Receptor Ligands


Abstract
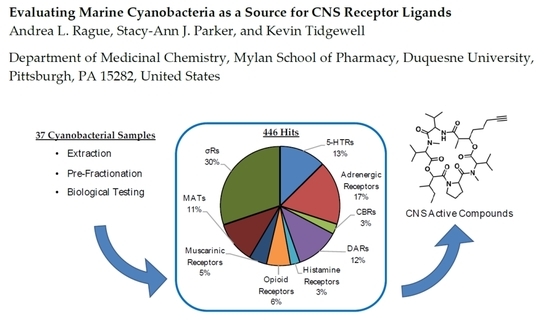
1. Introduction
2. Cyanobacterial Library Screening
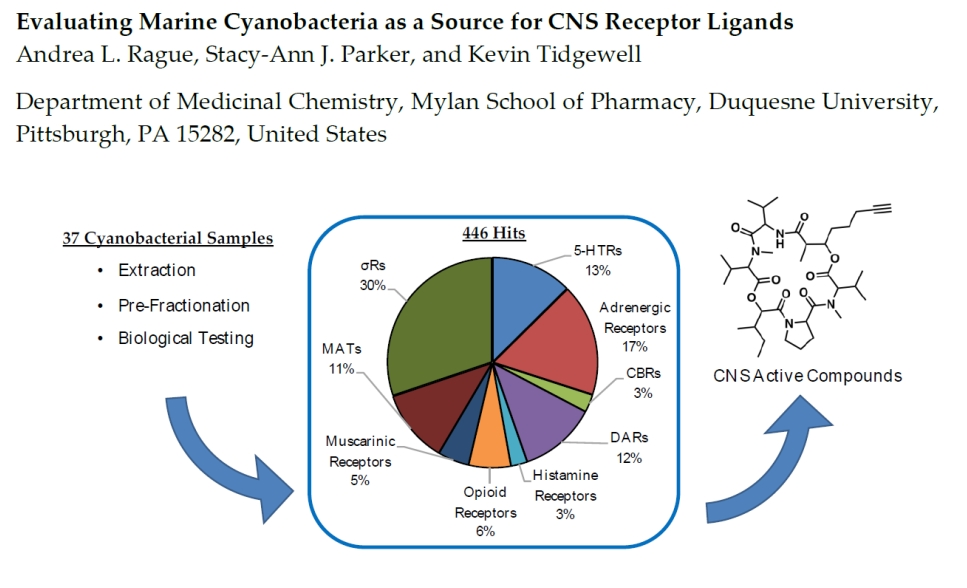
2.1. Radiolabaled Ligand Screening of Marine Cyanobacterial Extracts and Fractions
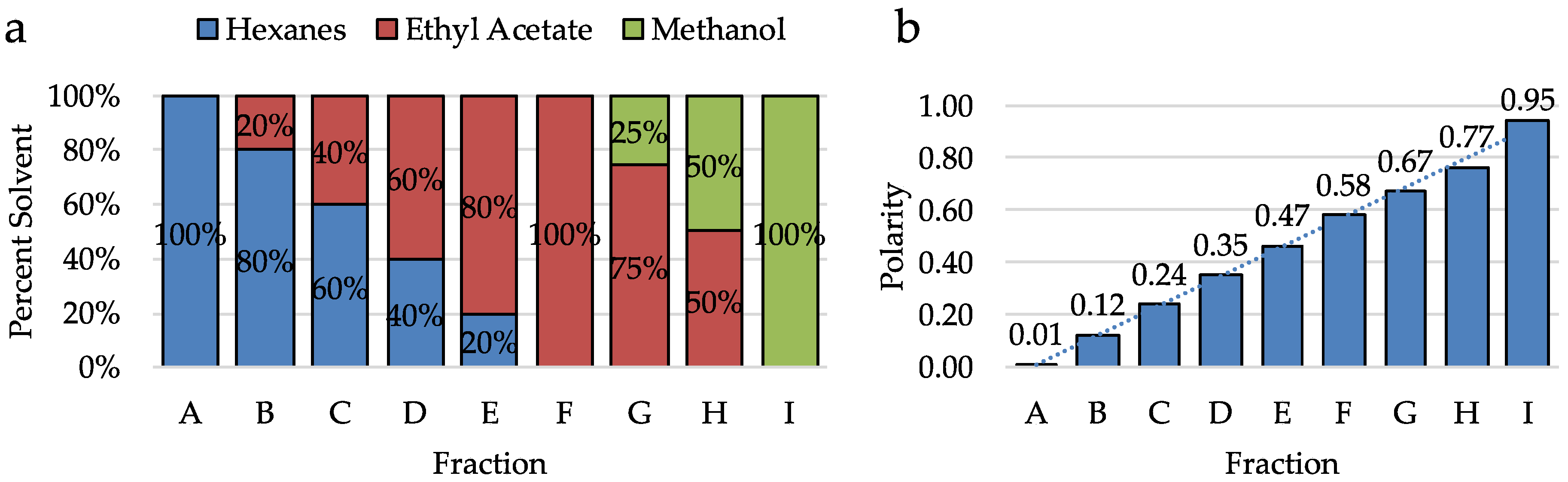
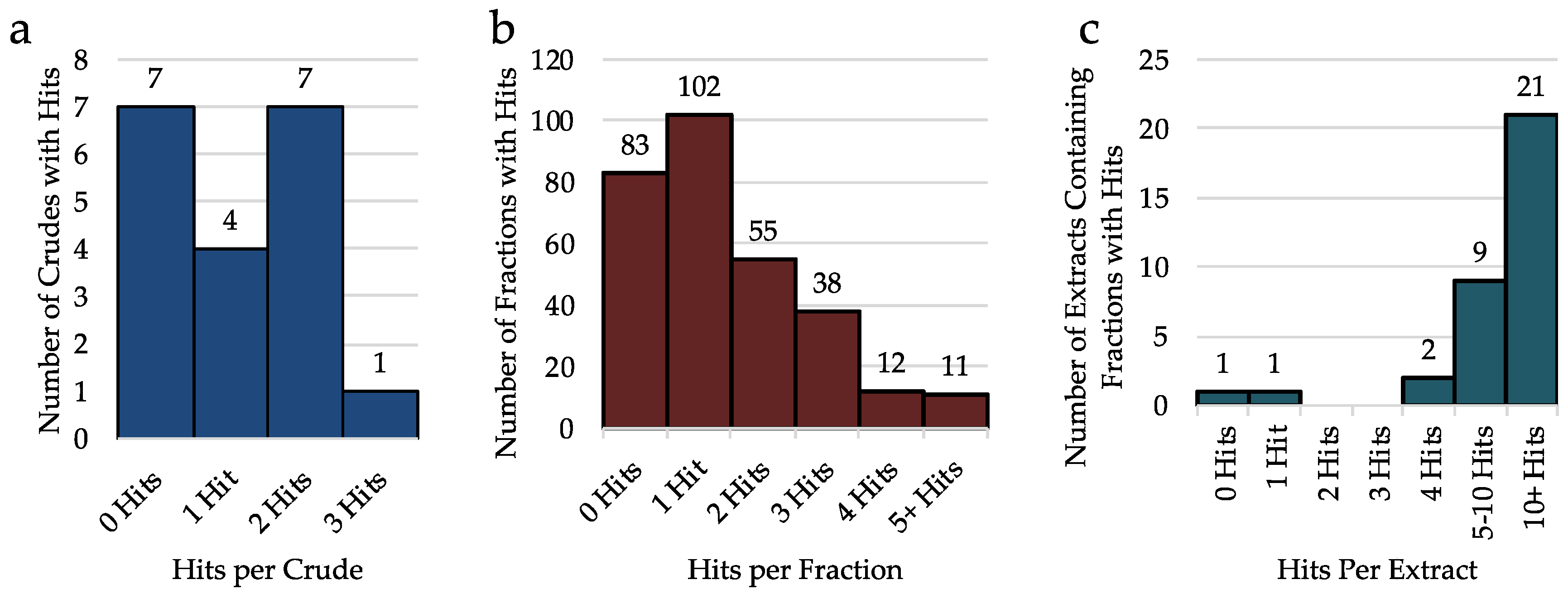
2.2. Analysis of Crude Extracts
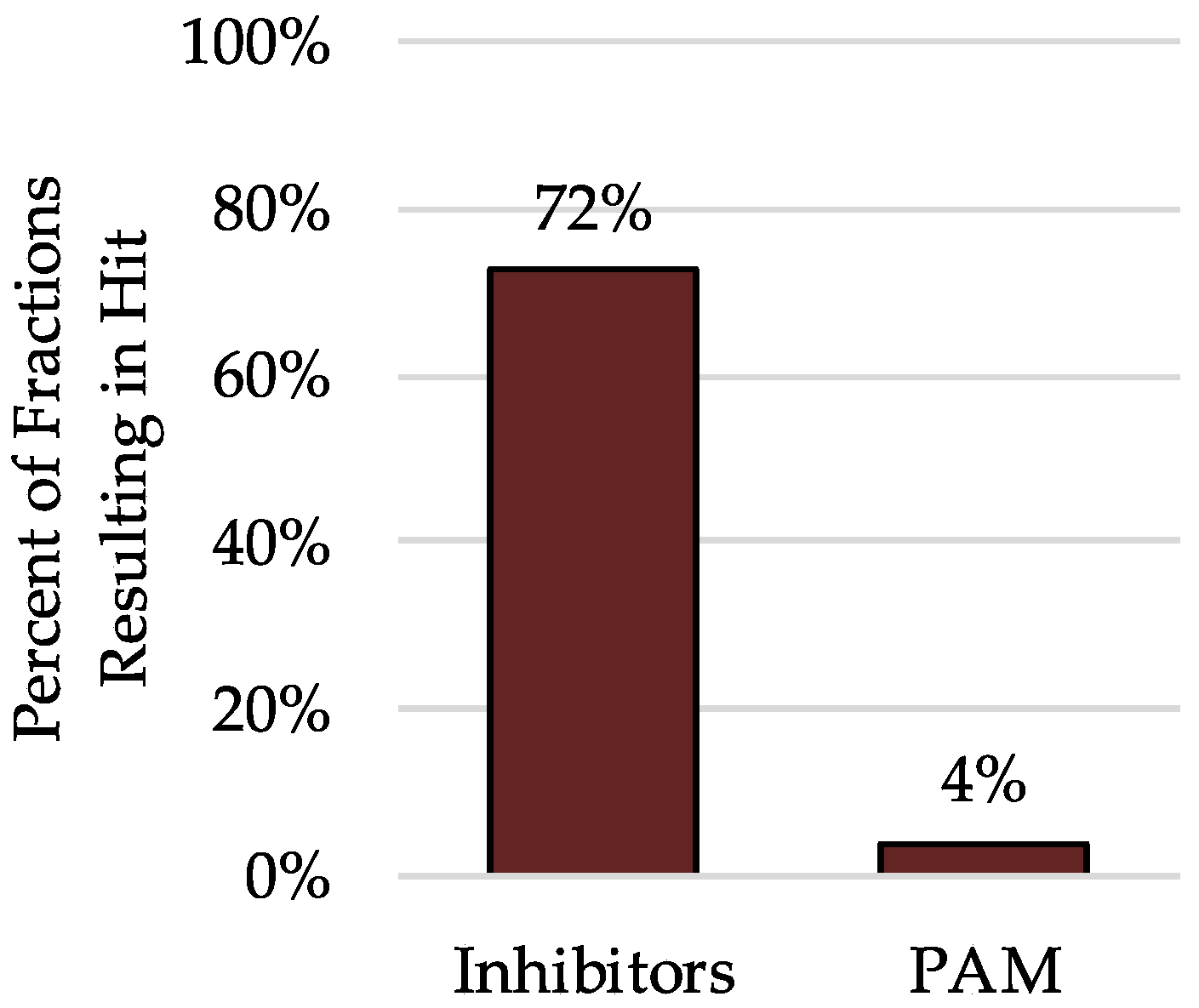
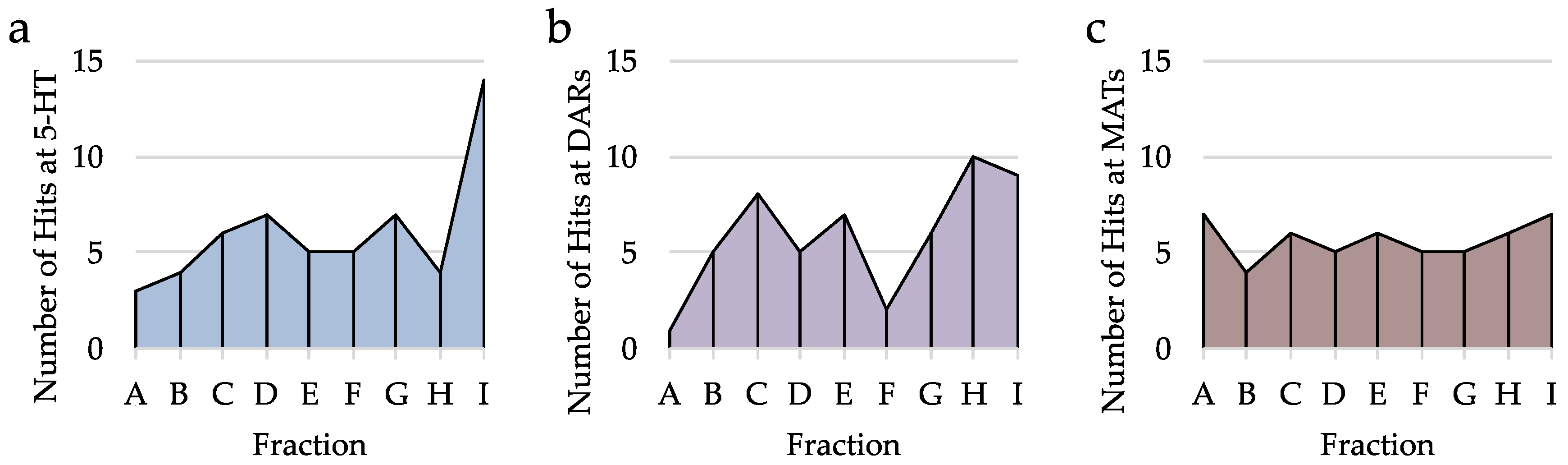
2.3. Analysis of Hits from Individual Fractions
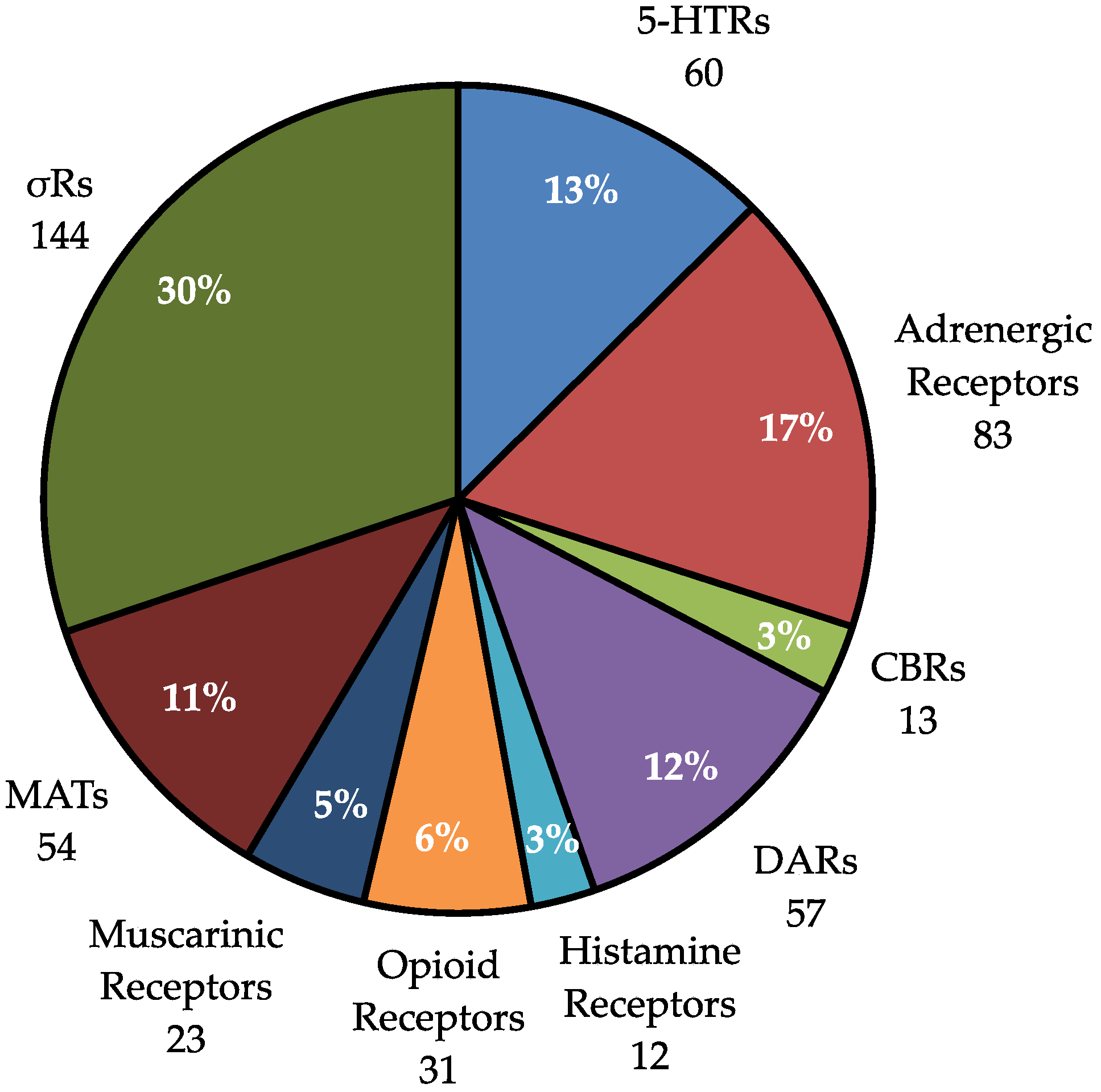
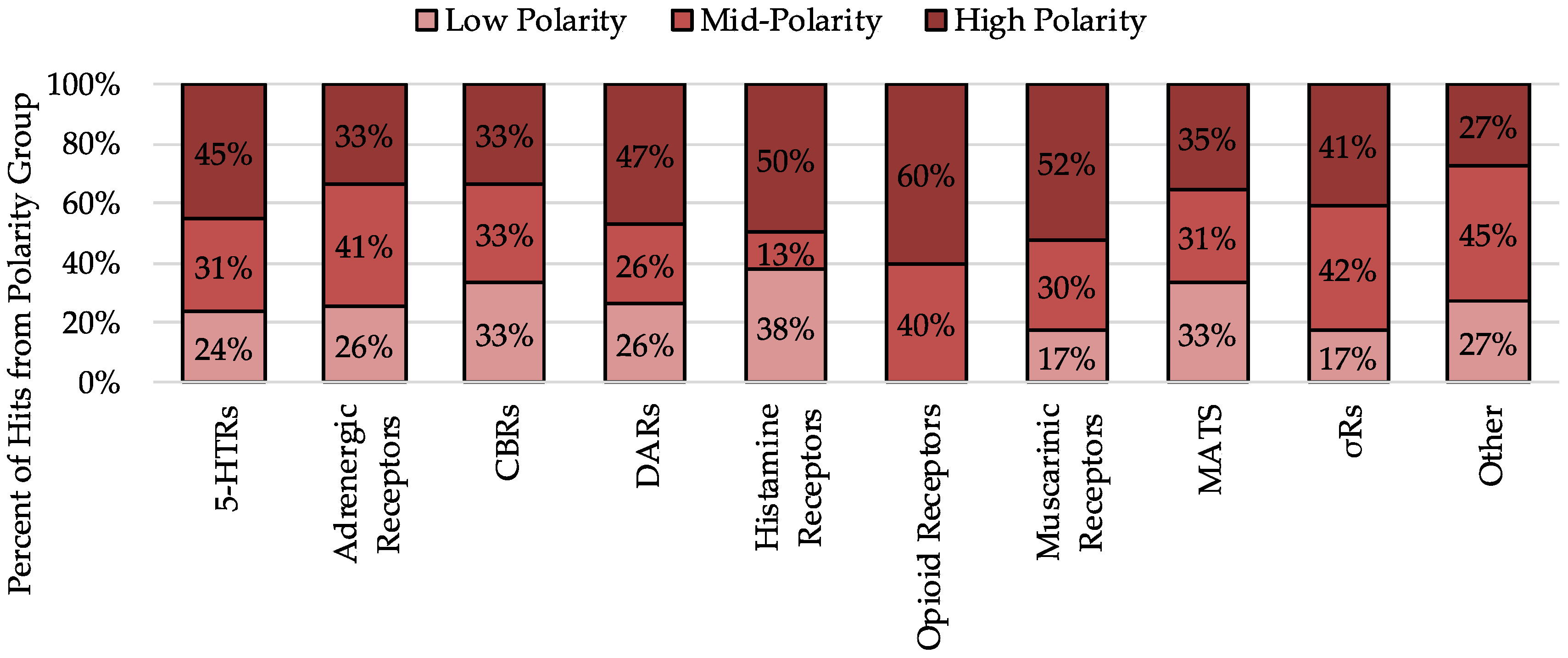
2.4. Analysis of Hits by Receptor Group
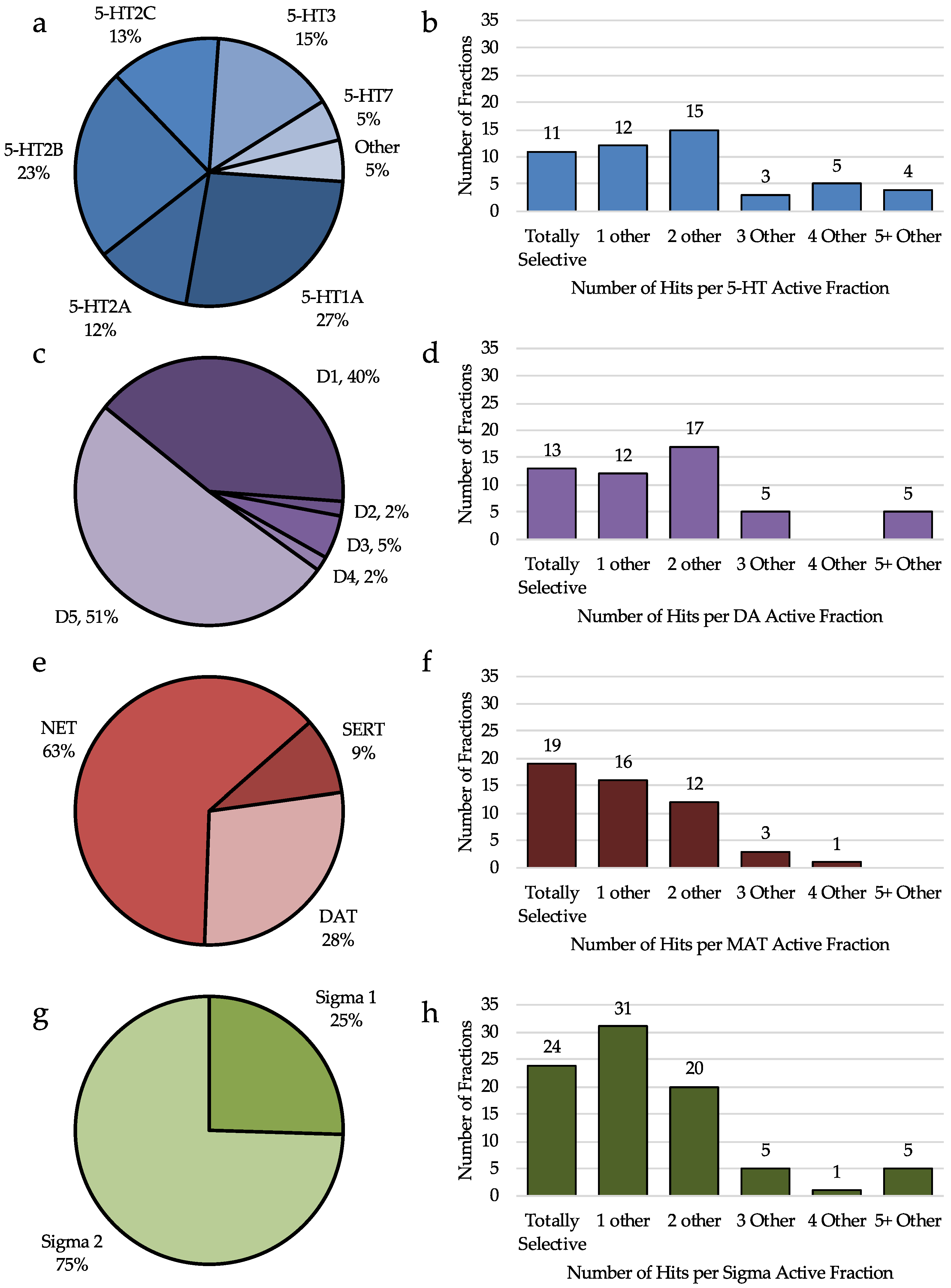
2.4.1. Analysis of Serotonin Receptor Hits
2.4.2. Analysis of Dopamine Receptor Hits
2.4.3. Analysis of Monoamine Transporter Hits
2.4.4. Analysis of Sigma Receptor Hits
2.5. Analysis of Hits by Fraction
3. Case Studies from Panamanian Cyanobacterial Extract DUQ0008
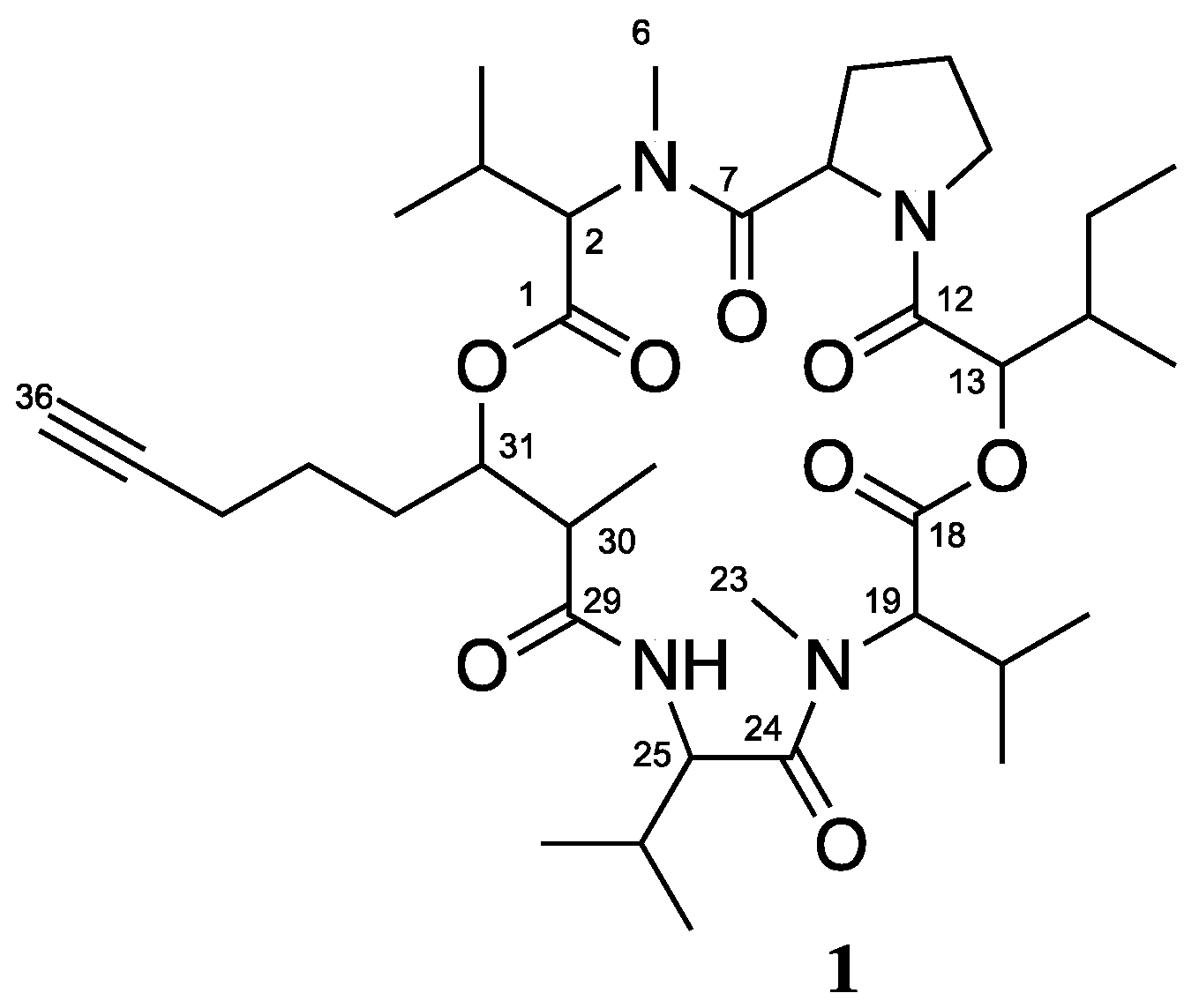
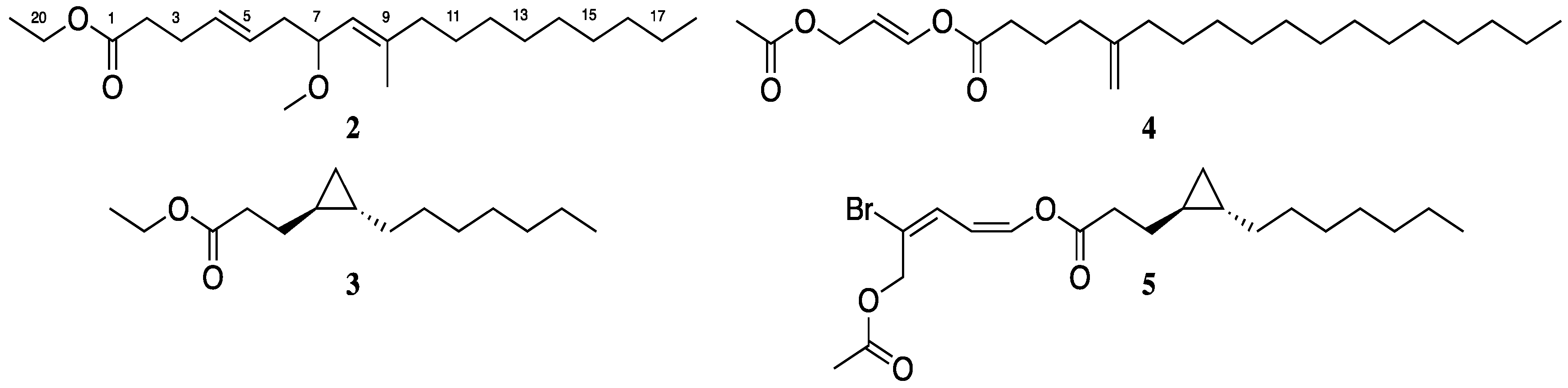
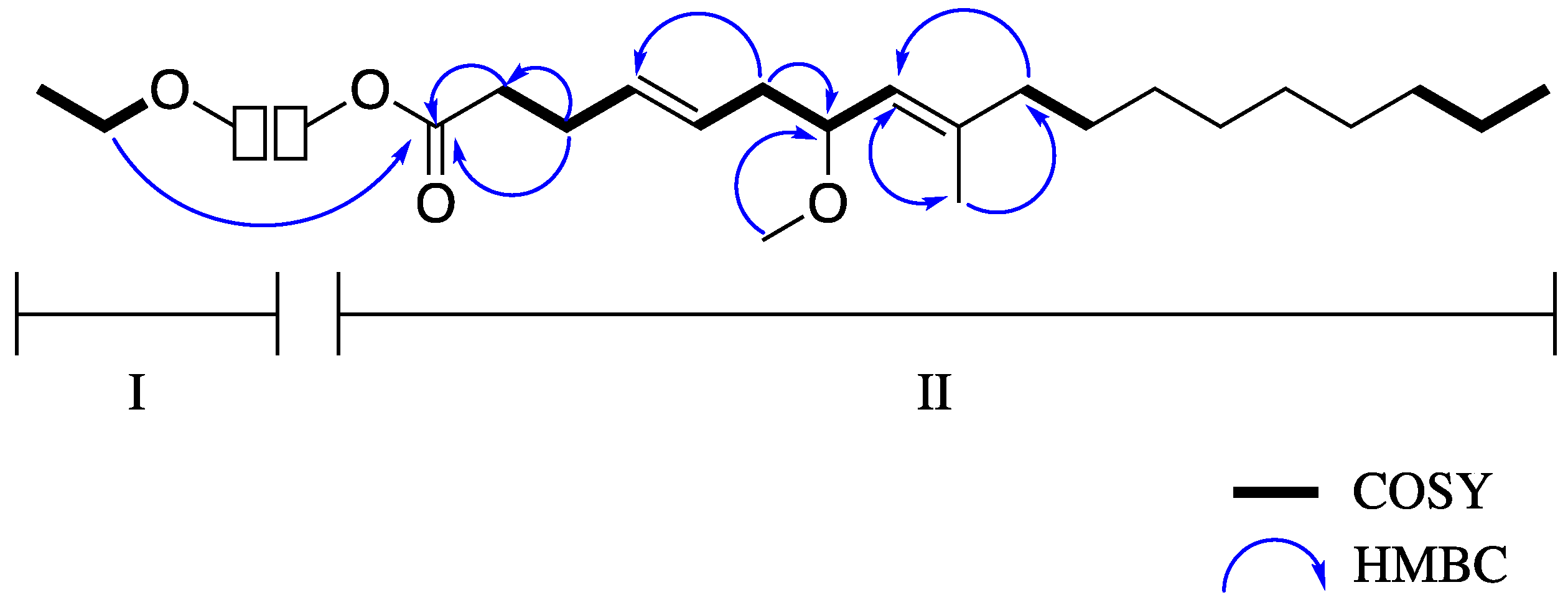
3.1. Isolation and Biological Testing of Veraguamide C
3.2. Isolation and Biological Testing of Fatty Acid Derivatives
4. Conclusions
5. Materials and Methods
5.1. General Experimental Procedures
5.2. Sample Material
5.3. Extraction and Isolation
5.4. Receptor Binding Profile
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marner, F.J.; Moore, R.E.; Hirotsu, K.; Clardy, J. Majusculamides A and B, two epimeric lipodipeptides from Lyngbya majuscula Gomont. J. Org. Chem. 1977, 42, 2815–2819. [Google Scholar] [CrossRef]
- Mynderse, J.S.; Moore, R.E.; Kashiwagi, M.; Norton, T.R. Antileukemia activity in the Osillatoriaceae: Isolation of Debromoaplysiatoxin from Lyngbya. Science 1977, 196, 538–540. [Google Scholar] [CrossRef] [PubMed]
- Medina, R.A.; Goeger, D.E.; Hills, P.; Mooberry, S.L.; Huang, N.; Romero, L.I.; Ortega-Barria, E.; Gerwick, W.H.; McPhail, K.L. Coibamide A, a potent antiproliferative cyclic depsipeptide from the Panamanian marine cyanobacterium Leptolyngbya sp. J. Am. Chem. Soc. 2008, 130, 6324–6325. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, M.; Tidgewell, K.; Capson, T.L.; Engene, N.; Almanza, A.; Schemies, J.; Jung, M.; Gerwick, W.H. Malyngolide Dimer, a Bioactive Symmetric Cyclodepside from the Panamanian Marine Cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 709–711. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Chan, K.P.; Chen, D.Y.-K.; Tan, L.T. Lagunamide C, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 2011, 72, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Malloy, K.L.; Villa, F.A.; Engene, N.; Matainaho, T.; Gerwick, L.; Gerwick, W.H. Malyngamide 2, an oxidized lipopeptide with nitric oxide inhibiting activity from a Papua New Guinea marine cyanobacterium. J. Nat. Prod. 2011, 74, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.T.; Lax, N.; Tidgewell, K. Central nervous system modulators from the ocean. In Marine Biomedicine from Beach to Bedside, 1st ed.; Baker, B.J., Ed.; CRC Press: Boca Raton, FL, USA, 2015; ISBN 9781466582125-CAT# K18918. [Google Scholar]
- Wernicke, J.F.; Iyengar, S.; Dolores Ferrer-Garcia, M. Treatment of Chronic Pain with Drugs that Modulate Central Nervous System Serotonin and Norepinephrine. Curr. Drug Ther. 2007, 2, 161–167. [Google Scholar] [CrossRef]
- Taylor, A.M.W.; Becker, S.; Schweinhardt, P.; Cahill, C. Mesolimbic dopamine signaling in acute and chronic pain: Implications for motivation, analgesia, and addiction. Pain 2016, 157, 1194–1198. [Google Scholar] [CrossRef] [PubMed]
- Belujon, P.; Grace, A.A. Dopamine System Dysregulation in Major Depressive Disorders. Int. J. Neuropsychopharmacol. 2017, 20, 1036–1046. [Google Scholar] [CrossRef] [PubMed]
- Lax, N.C.; Ahmed, K.T.; Ignatz, C.M.; Spadafora, C.; Kolber, B.J.; Tidgewell, K.J. Marine cyanobacteria-derived serotonin receptor 2C active fraction induces psychoactive behavioral effects in mice. Pharm. Biol. 2016, 54, 2723–2731. [Google Scholar] [CrossRef] [PubMed]
- Lax, N.C.; Parker, S.A.J.; Hilton, E.J.; Seliman, Y.; Tidgewell, K.J.; Kolber, B.J. Cyanobacterial extract with serotonin receptor subtype 7 (5-HT7R) affinity modulates depression and anxiety-like behavior in mice. Synapse 2018, 72, e22059. [Google Scholar] [CrossRef] [PubMed]
- Sahn, J.J.; Mejia, G.L.; Ray, P.R.; Martin, S.F.; Price, T.J. Sigma 2 Receptor/Tmem97 Agonists Produce Long Lasting Antineuropathic Pain Effects in Mice. ACS Chem. Neurosci. 2017, 8, 1801–1811. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 68, 954–979. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural Product Biosynthetic Diversity and Comparative Genomics of the Cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Besnard, J.; Ruda, G.F.; Setola, V.; Abecassis, K.; Rodriguiz, R.M.; Huang, X.P.; Norval, S.; Sassano, M.F.; Shin, A.I.; Webster, L.A.; et al. Automated design of ligands to polypharmacological profiles. Nature 2012, 492, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Al Dulayymi, J.R.; Baird, M.S.; Jones, K. The absolute stereochemistry of grenadamide. Tetrahedron 2004, 60, 341–345. [Google Scholar] [CrossRef]
- Gutiérrez, M.; Pereira, A.R.; Debonsi, H.M.; Ligresti, A.; Di Marzo, V.; Gerwick, W.H. Cannabinomimetic Lipid from a Marine Cyanobacterium. J. Nat. Prod. 2011, 74, 2313–2317. [Google Scholar] [CrossRef] [PubMed]
- Sitachitta, N.; Gerwick, W.H. Grenadadiene and grenadamide, cyclopropyl-containing fatty acid metabolites from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 1998, 61, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Mevers, E.; Liu, W.T.; Engene, N.; Mohimani, H.; Byrum, T.; Pevzner, P.A.; Dorrestein, P.C.; Spadafora, C.; Gerwick, W.H. Cytotoxic Veraguamides, Alkynyl Bromide-Containing Cyclic Depsipeptides from the Marine Cyanobacterium cf. Oscillatoria margaritifera. J. Nat. Prod. 2011, 74, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Macmillan, J.B.; Molinski, T.F. Majusculoic acid, a brominated cyclopropyl fatty acid from a marine cyanobacterial mat assemblage. J. Nat. Prod. 2005, 68, 604–606. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
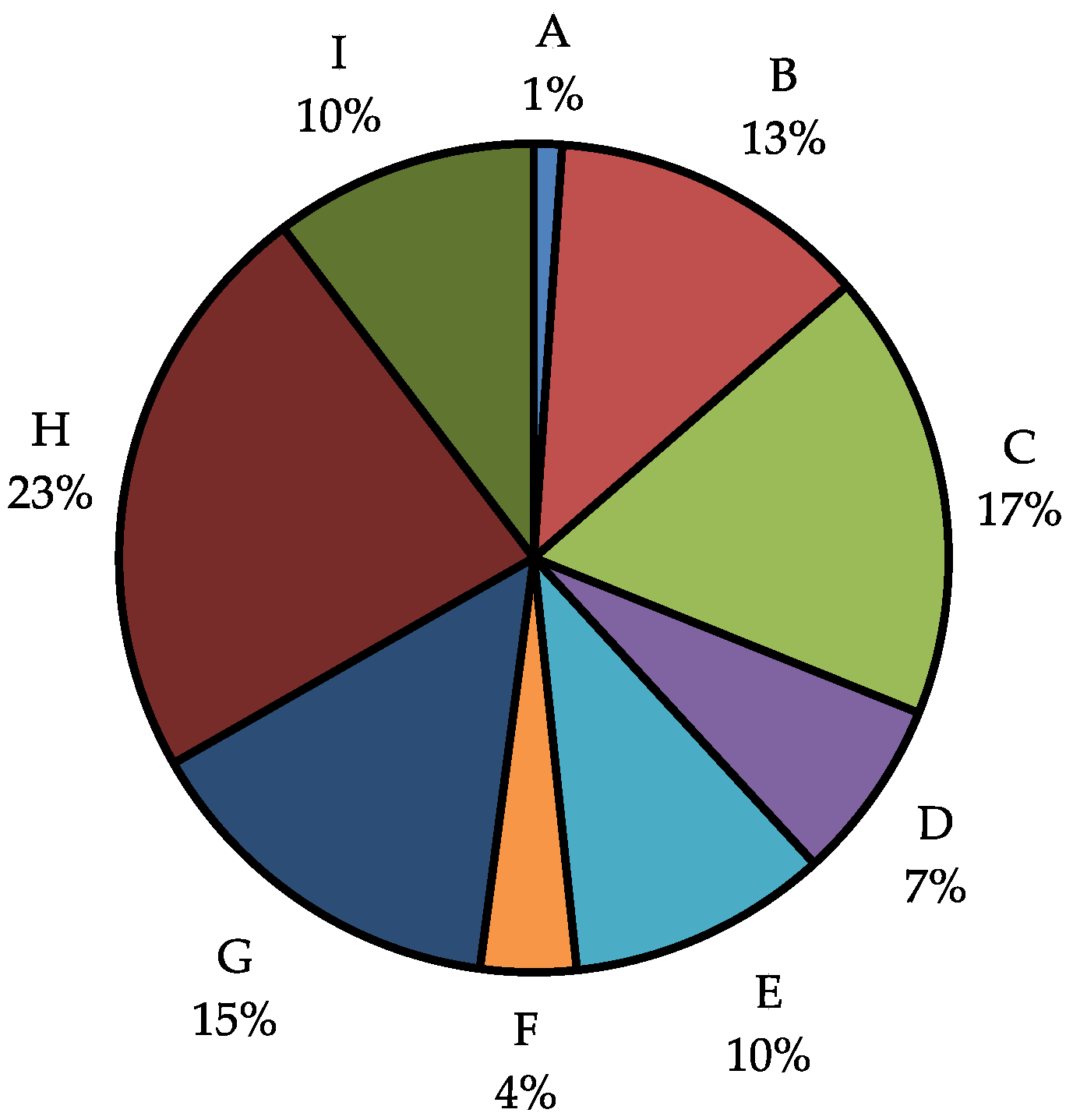{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 5-HT1A (IC50) | 5-HT2A (IC50) | 5-HT2C (IC50) | 5-HT7 (IC50) | Alpha2B (IC50) | D1 (IC50) | D5 (IC50) | DOR (IC50) | σ2R (IC50) | |
|---|---|---|---|---|---|---|---|---|---|
| Crude | - | - | - | - | - | - | - | - | 722.0 ± 0.8 |
| A | - | - | - | - | - | - | - | - | - |
| B | - | - | - | - | - | - | - | - | - |
| C | - | - | 191.0 ± 0.8 | - | - | 5083.0 ± 0.9 | - | - | - |
| D | - | >10,000 | - | - | 1999.7 ± 0.8 | - | - | 1776.0 ± 0.9 | - |
| E | - | - | - | - | - | - | - | - | 1502.0 ± 0.8 |
| F | - | - | - | - | - | - | - | - | - |
| G | - | 1854.0 ± 0.9 | 98.0 ± 0.8 | - | 2030.0 ± 0.8 | 1526.0 ± 0.9 | 1531.0 ± 0.9 | 1618.0 ± 0.9 | 1085.0 ± 0.8 |
| H | - | - | - | - | - | - | - | - | 1874.0 ± 0.8 |
| I | 2441.0 ± 0.8 | - | - | 1223.00 ± 0.8 | - | - | - | - | 424.0 ± 0.8 |
| Pos. | δC, Type | δH Multi. (J in Hz) | HMBC a |
|---|---|---|---|
| 1 | 173.6, C | ||
| 2 | 34.4, CH2 | 2.34 m | 1, 3 |
| 3 | 28.0, CH2 | 2.32 m | 1 |
| 4 | 127.4, CH | 5.47 m | 6 |
| 5 | 130.4, CH | 5.45 m | 6 |
| 6 | 38.9, CH2 | 2.28 m, 2.13 m | 4, 5, 7 |
| 7 | 77.2, CH | 3.88 m | 20 |
| 8 | 125.2, CH | 4.99 d (8.9) | 10, 19 |
| 9 | 140.2, C | ||
| 10 | 39.9, CH2 | 2.01 t (7.0) | 8, 9, 11, 19 |
| 11 | 27.8, CH2 | 1.41 m | |
| 12 | 29.4, CH2 | 1.27 m | |
| 13 | 29.4, CH2 | 1.27 m | |
| 14 | 29.4, CH2 | 1.27 m | |
| 15 | 29.4, CH2 | 1.27 m | |
| 16 | 31.9, CH2 | 1.27 m | |
| 17 | 22.5, CH2 | 1.29 m b | |
| 18 | 14.4, CH3 | 0.88 t (7.1) | 16, 17 |
| 19 | 16.7, CH3 | 1.64 s | 8, 9, 10 |
| 20 | 55.5, OCH3 | 3.23 s | 7 |
| 21 | 60.2, OCH2 | 4.13 q (8.0, 7.9) | 1, 22 |
| 22 | 14.5, CH3 | 1.26 t b | 21 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rague, A.L.; Parker, S.-A.J.; Tidgewell, K.J. Evaluating Marine Cyanobacteria as a Source for CNS Receptor Ligands. Molecules 2018, 23, 2665. https://doi.org/10.3390/molecules23102665
Rague AL, Parker S-AJ, Tidgewell KJ. Evaluating Marine Cyanobacteria as a Source for CNS Receptor Ligands. Molecules. 2018; 23(10):2665. https://doi.org/10.3390/molecules23102665
Chicago/Turabian StyleRague, Andrea L., Stacy-Ann J. Parker, and Kevin J. Tidgewell. 2018. "Evaluating Marine Cyanobacteria as a Source for CNS Receptor Ligands" Molecules 23, no. 10: 2665. https://doi.org/10.3390/molecules23102665
APA StyleRague, A. L., Parker, S.-A. J., & Tidgewell, K. J. (2018). Evaluating Marine Cyanobacteria as a Source for CNS Receptor Ligands. Molecules, 23(10), 2665. https://doi.org/10.3390/molecules23102665