
Anti-Inflammatory and Antioxidant Properties of Peptides Released from β-Lactoglobulin by High Hydrostatic Pressure-Assisted Enzymatic Hydrolysis

Abstract
:1. Introduction
2. Results
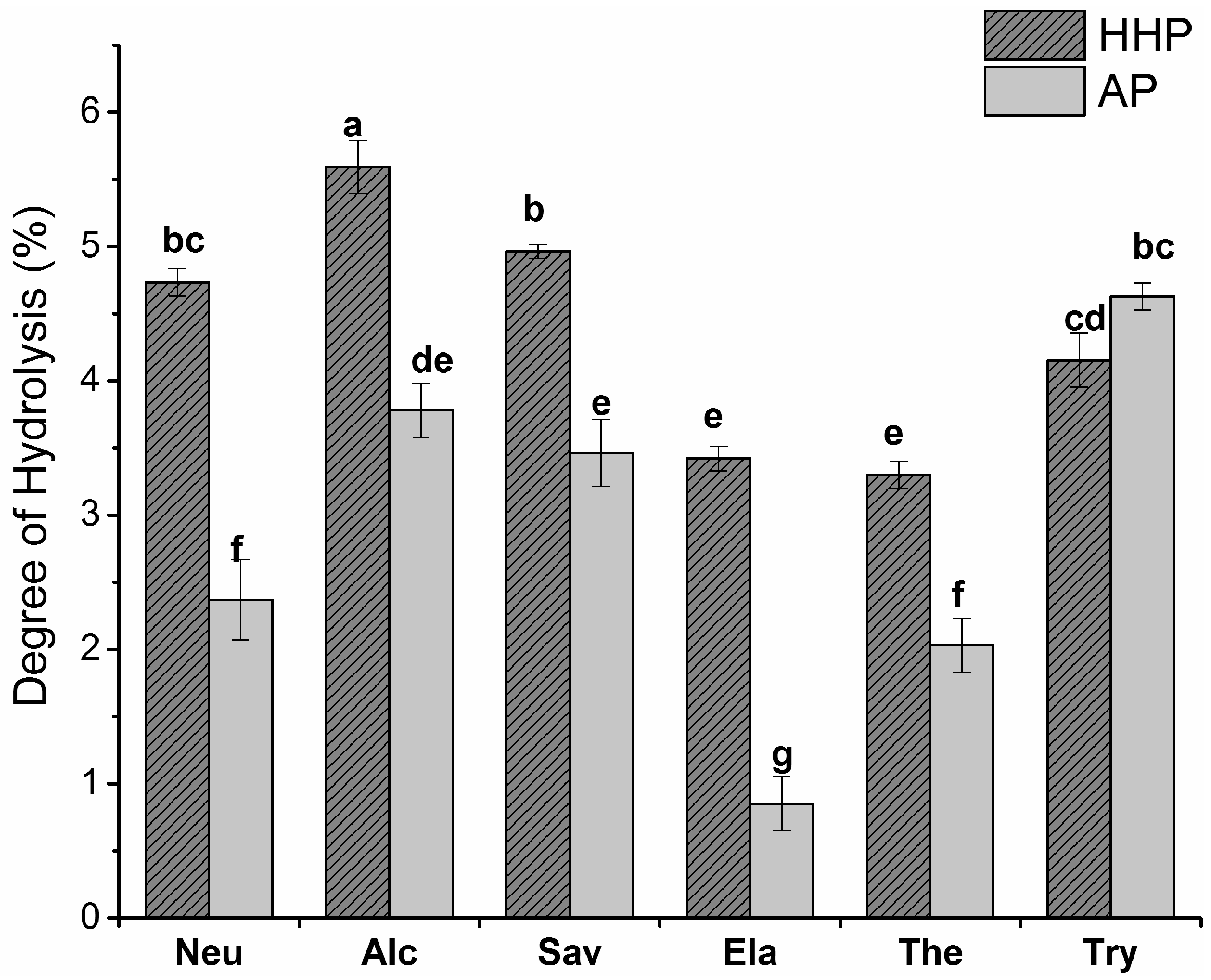
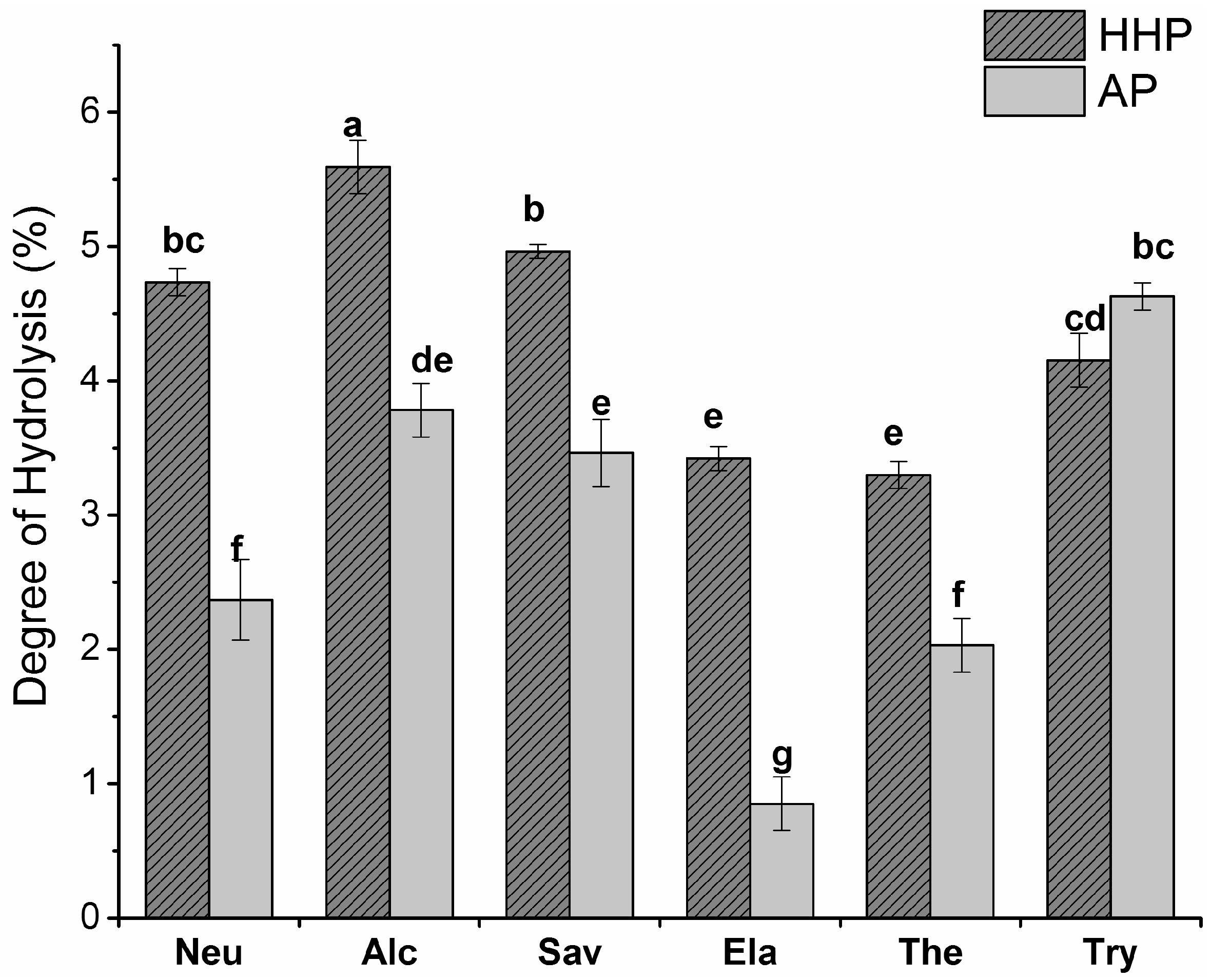
2.1. Degree of Hydrolysis (DH)
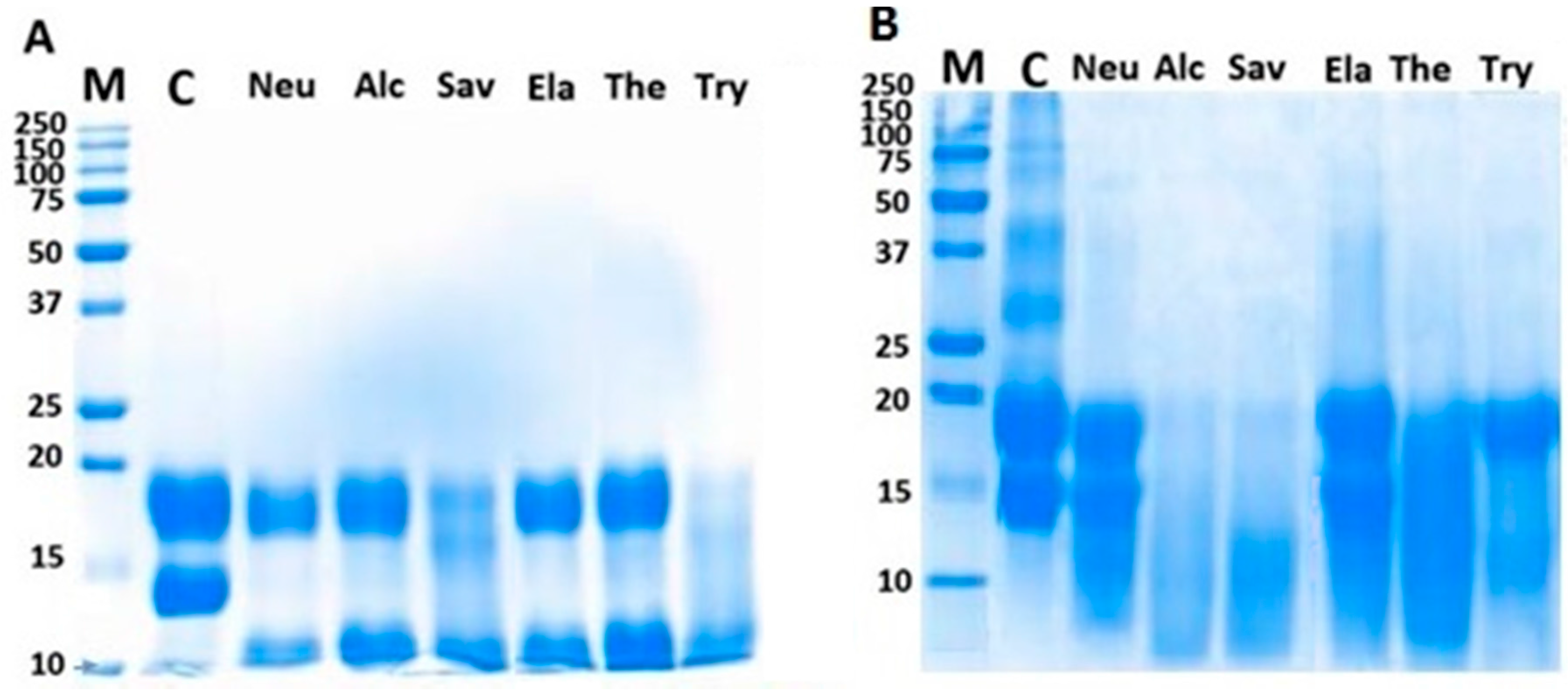
2.2. Sodium Dodecyl Sulfate Gel Electrophoresis (SDS-PAGE)
2.3. Matrix-Assisted Laser Desorption/Ionization Time-Of-Flight (MALDI-TOF) Analysis
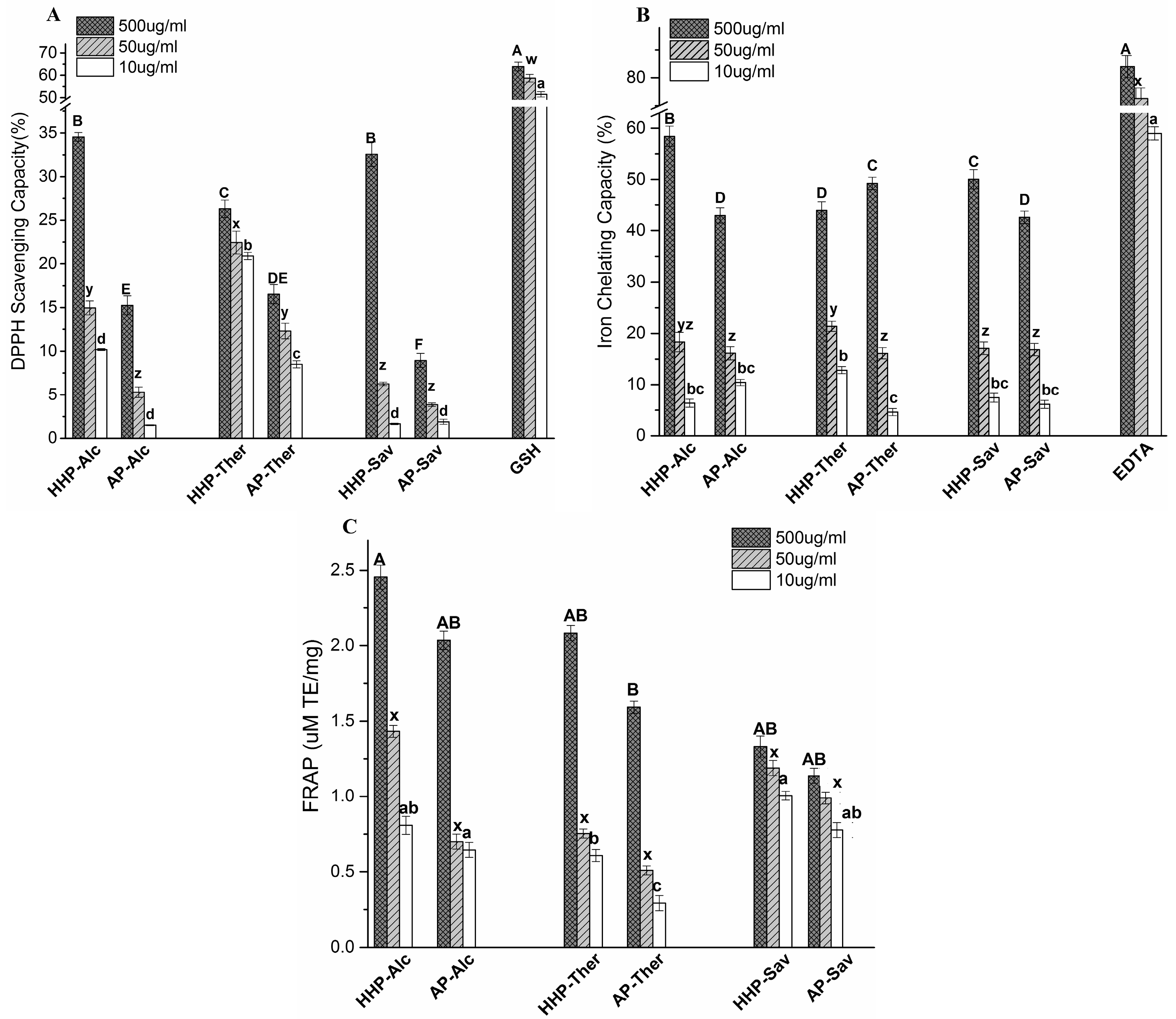
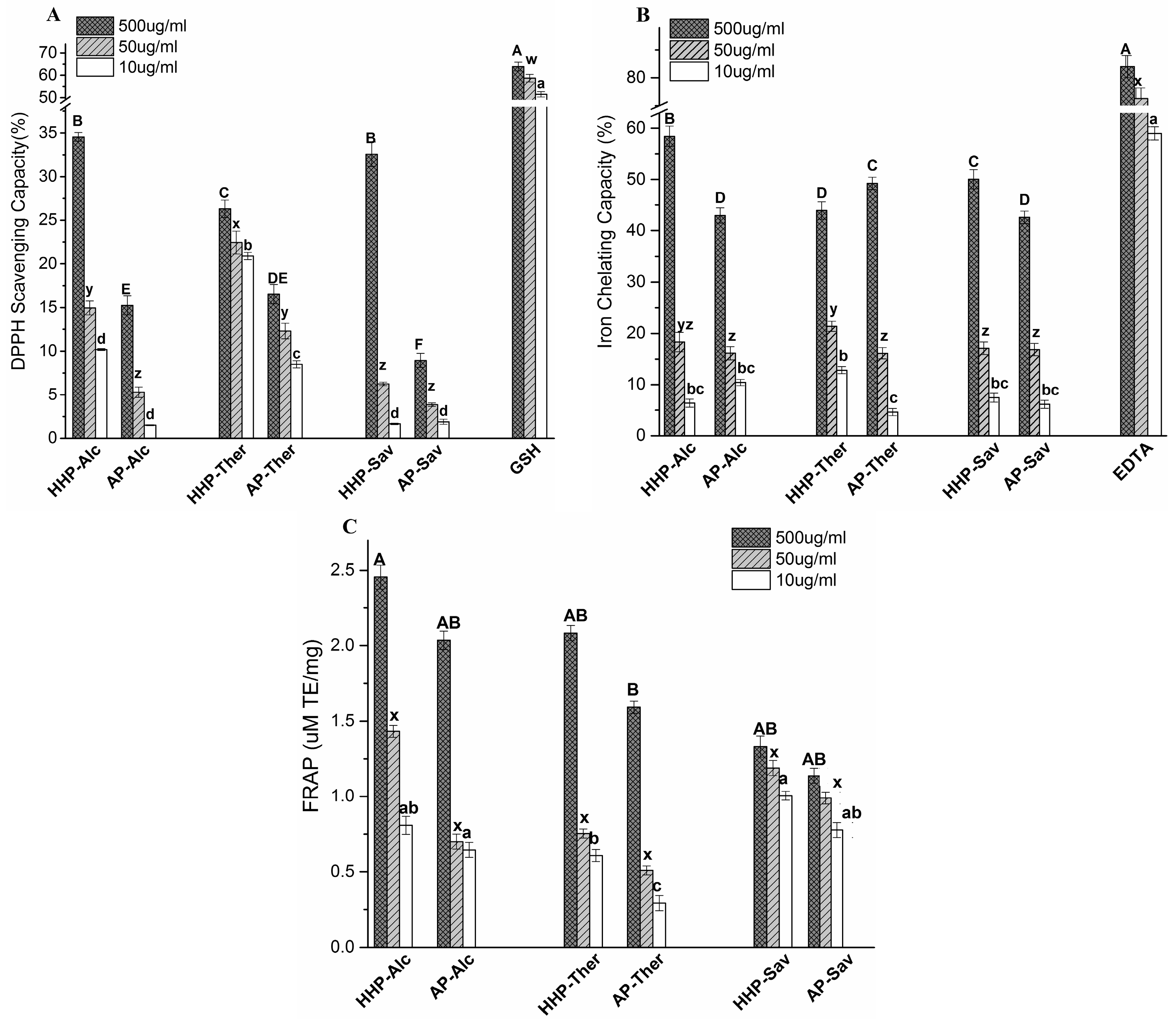
2.4. Antioxidant Capacity of BLG Hydrolysates
2.4.1. 1,1-Diphenyl-2-Picryl Hydrazyl (DPPH) Scavenging Capacity
2.4.2. Iron Chelating Activity
2.4.3. The Ferric Reducing Antioxidant Power (FRAP) Assay
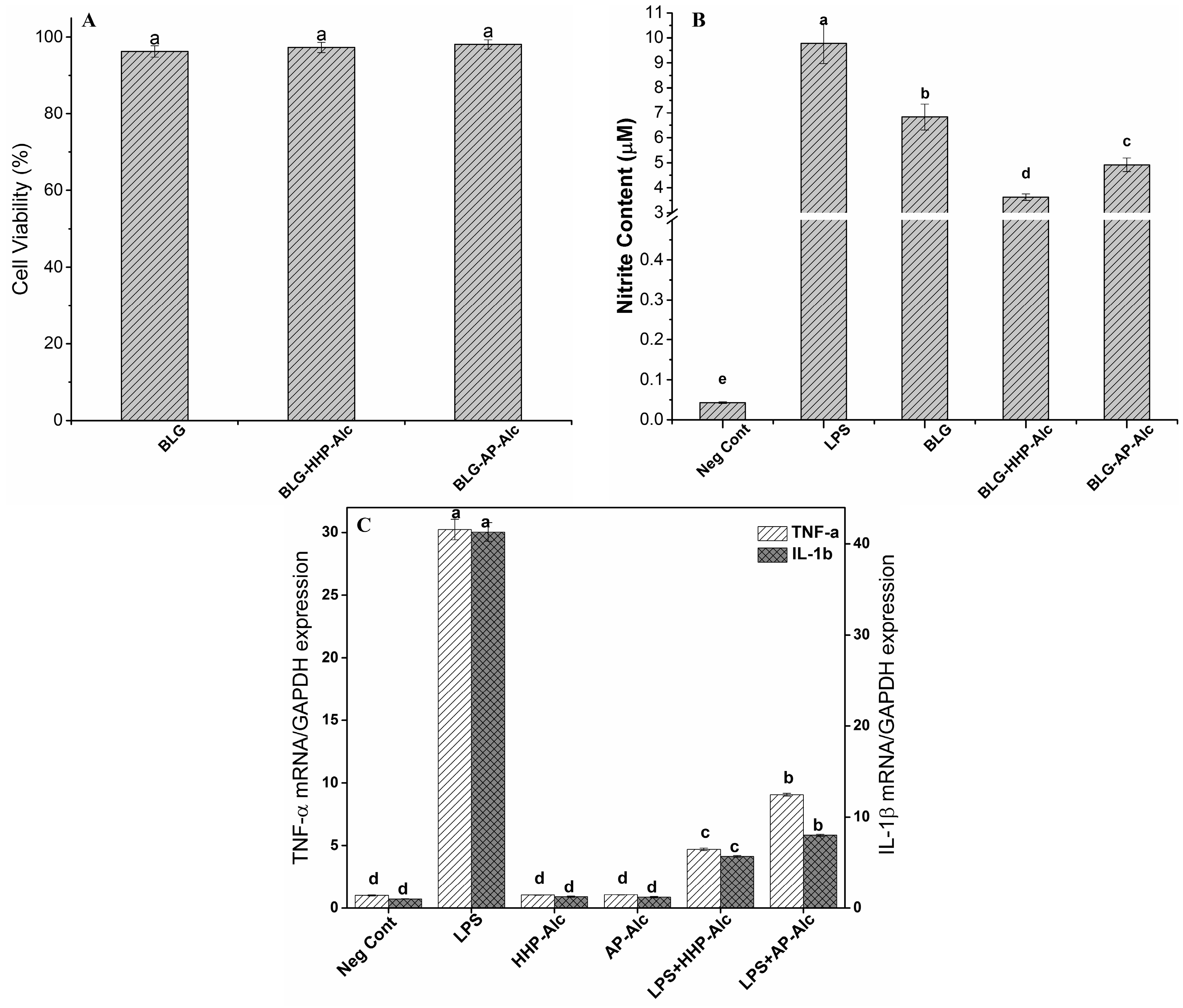
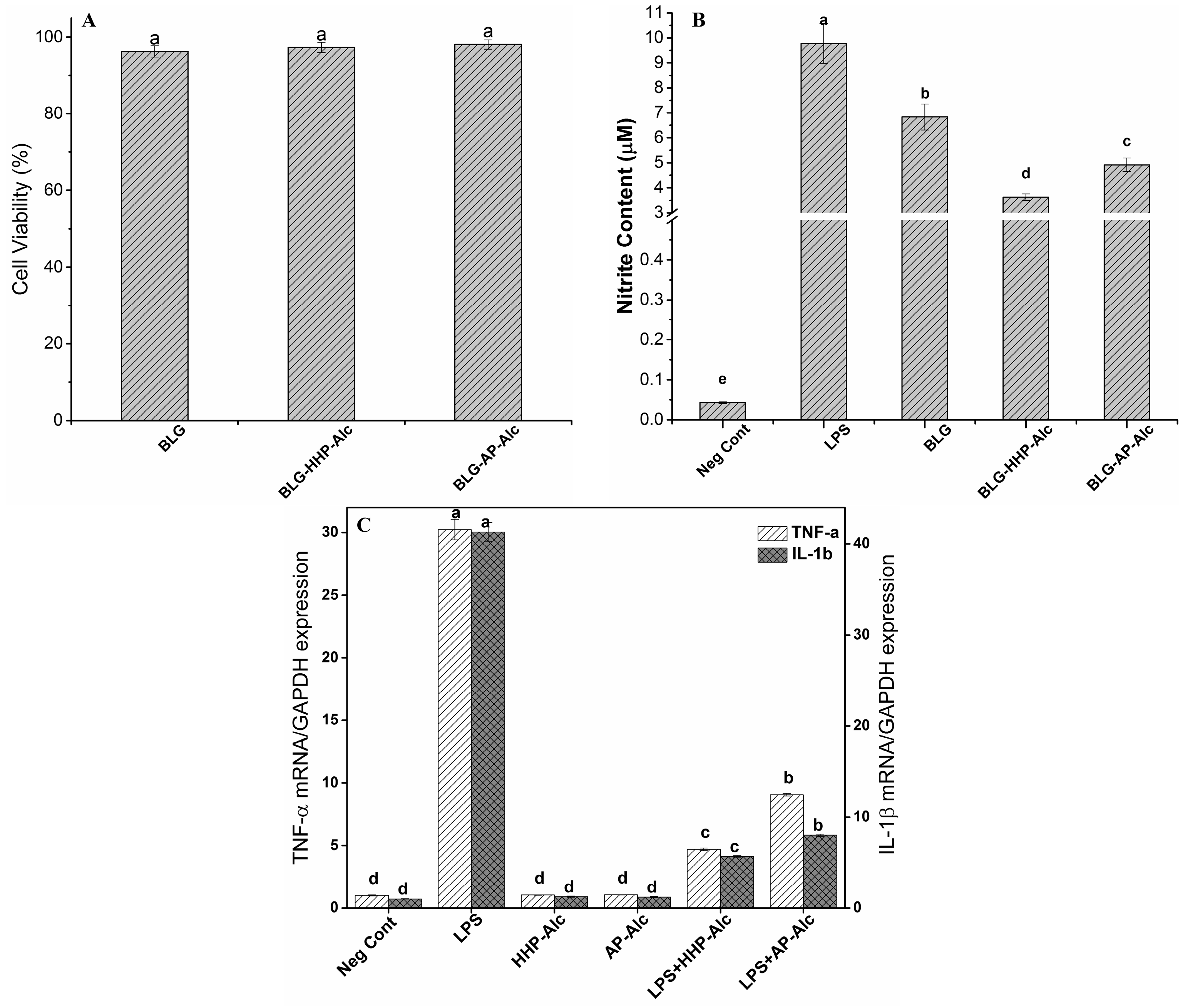
2.5. Anti-Inflammatory Properties of BLG Hydrolysates
2.5.1. Cell Viability Assay
2.5.2. Determination of Nitric Oxide (NO) Production by Macrophage Cells
2.5.3. Gene Expression of Pro-Inflammatory Cytokines in LPS-Stimulated Macrophages
2.6. Peptide Sequencing of the Most Effective BLG Hydrolysate
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Apparatus
4.3. Enzymatic Hydrolysis
4.4. Degree of Hydrolysis
4.5. Sodium Dodecyl Sulfate-Poly Acrylamide Gel Electrophoresis (SDS-PAGE)
4.6. Matrix-Assisted Laser Desorption/Ionization Time-Of-Flight (MALDI-TOF) Analysis
4.7. Antioxidant Capacity Analysis
4.7.1. 1,1-Diphenyl-2-Picryl Hydrazyl (DPPH) Scavenging Assay
4.7.2. Iron Chelating Capacity
4.7.3. The Ferric Reducing Antioxidant Power (FRAP) Assay
4.8. Cell Viability Assay
4.9. Determination of Nitric Oxide (NO) Production
4.10. Real-Time Polymerase Chain Reaction (RT-PCR) Analysis for Cytokine Gene Expression
4.11. Liquid Chromatography-Tandem Mass Spectrometry (LC–MS/MS)
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Khem, S.; Small, D.M.; May, B.K. The behaviour of whey protein isolate in protecting Lactobacillus plantarum. Food Chem. 2016, 190, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Haug, A.; Hostmark, A.T.; Harstad, O.M. Bovine milk in human nutrition—A review. Lipids Health Dis. 2007, 6. [Google Scholar] [CrossRef] [PubMed]
- Jeewanthi, R.K.C.; Lee, N.K.; Paik, H.D. Improved functional characteristics of whey protein hydrolysates in food industry. Korean J. Food Sci. Anim. Resour. 2015, 35, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Chatterton, D.E.W.; Smithers, G.; Roupas, P.; Brodkorb, A. Bioactivity of beta-lactoglobulin and alpha-lactalbumin—Technological implications for processing. Int. Dairy J. 2006, 16, 1229–1240. [Google Scholar] [CrossRef]
- Wada, Y.; Lonnerdal, B. Bioactive peptides released from in vitro digestion of human milk with or without pasteurization. Pediatr. Res. 2015, 77, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Dupont, D.; Boutrou, R.; Menard, O.; Jardin, J.; Tanguy, G.; Schuck, P.; Haab, B.B.; Leonil, J. Heat treatment of milk during powder manufacture increases casein resistance to simulated infant digestion. Food Dig. 2010, 1, 28–39. [Google Scholar] [CrossRef]
- Zeece, M.; Huppertz, T.; Kelly, A. Effect of high-pressure treatment on in vitro digestibility of beta-lactoglobulin. Innov. Food Sci. Emerg. Technol. 2008, 9, 62–69. [Google Scholar] [CrossRef]
- Hernandez-Ledesma, B.; Davalos, A.; Bartolome, B.; Amigo, L. Preparation of antioxidant enzymatic hydrolysates from alpha-lactalbumin and beta-lactoglobulin. Identification of active peptides by HPLC-MS/MS. J. Agric. Food Chem. 2005, 53, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Pena-Ramos, E.A.; Xiong, Y.L.L.; Arteaga, G.E. Fractionation and characterisation for antioxidant activity of hydrolysed whey protein. J. Sci. Food Agric. 2004, 84, 1908–1918. [Google Scholar] [CrossRef]
- Mullally, M.M.; Meisel, H.; Fitzgerald, R.J. Identification of a novel angiotensin-I-converting enzyme inhibitory peptide corresponding to a tryptic fragment of bovine beta-lactoglobulin. FEBS Lett. 1997, 402, 99–101. [Google Scholar] [CrossRef]
- Huang, S.M.; Chen, K.N.; Chen, Y.P.; Hong, W.S.; Chen, M.J. Immunomodulatory properties of the milk whey products obtained by enzymatic and microbial hydrolysis. Int. J. Food Sci. Technol. 2010, 45, 1061–1067. [Google Scholar] [CrossRef]
- Pellegrini, A.; Dettling, C.; Thomas, U.; Hunziker, P. Isolation and characterization of four bactericidal domains in the bovine beta-lactoglobulin. BBA-Gen. Subj. 2001, 1526, 131–140. [Google Scholar] [CrossRef]
- Theolier, J.; Hammami, R.; Labelle, P.; Fliss, I.; Jean, J. Isolation and identification of antimicrobial peptides derived by peptic cleavage of whey protein isolate. J. Funct. Foods 2013, 5, 706–714. [Google Scholar] [CrossRef]
- Antila, P.; Paakkari, I.; Jarvinen, A.; Mattila, M.J.; Laukkanen, M.; Pihlanto-Leppala, A.; Mantsala, P.; Hellman, J. Opioid peptides derived from in vitro proteolysis of bovine whey proteins. Int. Dairy J. 1991, 1, 215–229. [Google Scholar] [CrossRef]
- Tulipano, G.; Faggi, L.; Nardone, A.; Cocchi, D.; Caroli, A.M. Characterization of the potential of -lactoglobulin and -lactalbumin as sources of bioactive peptides affecting incretin function: In silico and in vitro comparative studies. Int. Dairy J. 2015, 48, 66–72. [Google Scholar] [CrossRef]
- Tornvall, U. Pinpointing oxidative modifications in proteins-recent advances in analytical methods. Anal. Methods-UK 2010, 2, 1638–1650. [Google Scholar] [CrossRef]
- Barbe, F.; Menard, O.; Le Gouar, Y.; Buffiere, C.; Famelart, M.H.; Laroche, B.; Le Feunteun, S.; Dupont, D.; Remond, D. The heat treatment and the gelation are strong determinants of the kinetics of milk proteins digestion and of the peripheral availability of amino acids. Food Chem. 2013, 136, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Uluko, H.; Zhang, S.; Liu, L.; Li, H.; Cui, W.; Xue, H.; Zhao, L.; Sun, Y.; Lu, J.; Lv, J. Pilot-scale membrane fractionation of ACE inhibitory and antioxidative peptides from ultrasound pretreated milk protein concentrate hydrolysates. J. Funct. Foods 2014, 7, 350–361. [Google Scholar] [CrossRef]
- Wang, N.; Li, L. Reproducible microwave-assisted acid hydrolysis of proteins using a household microwave oven and its combination with LC-ESI Ms/Ms for mapping protein sequences and modifications. J. Am. Soc. Mass Spectrom. 2010, 21, 1573–1587. [Google Scholar] [CrossRef] [PubMed]
- Datta, N.; Deeth, H.C. High pressure processing of milk and dairy products. Aust. J. Dairy Technol. 1999, 54, 41–48. [Google Scholar]
- Rivalain, N.; Roquain, J.; Demazeau, G. Development of high hydrostatic pressure in biosciences: Pressure effect on biological structures and potential applications in biotechnologies. Biotechnol. Adv. 2010, 28, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Sunwoo, H.H.; Gujral, N.; Huebl, A.C.; Kim, C.T. Application of high hydrostatic pressure and enzymatic hydrolysis for the extraction of ginsenosides from fresh ginseng root (Panax ginseng C.A. Myer). Food Bioprocess Technol. 2014, 7, 1246–1254. [Google Scholar] [CrossRef]
- Garcia-Mora, P.; Penas, E.; Frias, J.; Gomez, R.; Martinez-Villaluenga, C. High-pressure improves enzymatic proteolysis and the release of peptides with angiotensin I converting enzyme inhibitory and antioxidant activities from lentil proteins. Food Chem. 2015, 171, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Bamdad, F.; Wu, J.P.; Chen, L.Y. Effects of enzymatic hydrolysis on molecular structure and antioxidant activity of barley hordein. J. Cereal Sci. 2011, 54, 20–28. [Google Scholar] [CrossRef]
- Zhang, J.H.; Zhang, H.; Wang, L.; Guo, X.N.; Wang, X.G.; Yao, H.Y. Antioxidant activities of the rice endosperm protein hydrolysate: Identification of the active peptide. Eur. Food Res. Technol. 2009, 229, 709–719. [Google Scholar] [CrossRef]
- Iskandar, M.M.; Dauletbaev, N.; Kubow, S.; Mawji, N.; Lands, L.C. Whey protein hydrolysates decrease IL-8 secretion in lipopolysaccharide (LPS)-stimulated respiratory epithelial cells by affecting LPS binding to toll-like receptor 4. Br. J. Nutr. 2013, 110, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Mann, B.; Kumari, A.; Kumar, R.; Sharma, R.; Prajapati, K.; Mahboob, S.; Athira, S. Antioxidant activity of whey protein hydrolysates in milk beverage system. J. Food Sci. Technol. Mysore 2015, 52, 3235–3241. [Google Scholar] [CrossRef] [PubMed]
- Buckow, R.; Weiss, U.; Heinz, V.; Knorr, D. Stability and catalytic activity of alpha-amylase from barley malt at different pressure-temperature conditions. Biotechnol. Bioeng. 2007, 97, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.T.; Gujral, N.; Ganguly, A.; Suh, J.W.; Sunwoo, H.H. Chondroitin sulphate extracted from antler cartilage using high hydrostatic pressure and enzymatic hydrolysis. Biotechnol. Rep. 2014, 4, 14–20. [Google Scholar] [CrossRef]
- Stapelfeldt, H.; Petersen, P.H.; Kristiansen, K.R.; Qvist, K.B.; Skibsted, L.H. Effect of high hydrostatic pressure on the enzymic hydrolysis of -lactoglobulin B by trypsin, thermolysin and pepsin. J. Dairy Res. 1996, 63, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Fandino, R. High pressure-induced changes in milk proteins and possible applications in dairy technology. Int. Dairy J. 2006, 16, 1119–1131. [Google Scholar] [CrossRef]
- Mozhaev, V.V.; Heremans, K.; Frank, J.; Masson, P.; Balny, C. High pressure effects on protein structure and function. Proteins 1996, 24, 81–91. [Google Scholar] [CrossRef]
- Heinrich, M.; Kulozik, U. Study of chymosin hydrolysis of casein micelles under ultra high pressure: Effect on re-association upon pressure release. Int. Dairy J. 2011, 21, 664–669. [Google Scholar] [CrossRef]
- Ludikhuyze, L.; Van Loey, A.; Indrawati; Smout, C.; Hendrickx, M. Effects of combined pressure and temperature on enzymes related to quality of fruits and vegetables: From kinetic information to process engineering aspects. Crit. Rev. Food Sci. 2003, 43, 527–586. [Google Scholar] [CrossRef] [PubMed]
- Eckert, E.; Bamdad, F.; Chen, L.Y. Metal solubility enhancing peptides derived from barley protein. Food Chem. 2014, 159, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Haversen, L.; Ohlsson, B.G.; Hahn-Zoric, M.; Hanson, L.A.; Mattsby-Baltzer, I. Lactoferrin down-regulates the LPS-induced cytokine production in monocytic cells via NF-kappa B. Cell. Immunol. 2002, 220, 83–95. [Google Scholar] [CrossRef]
- Elass-Rochard, E.; Legrand, D.; Salmon, V.; Roseanu, A.; Trif, M.; Tobias, P.S.; Mazurier, J.; Spik, G. Lactoferrin inhibits the endotoxin interaction with CD14 by competition with the lipopolysaccharide-binding protein. Infect. Immun. 1998, 66, 486–491. [Google Scholar] [PubMed]
- Chawla, A.; Nguyen, K.D.; Goh, Y.P.S. Macrophage-mediated inflammation in metabolic disease. Nat. Rev. Immunol. 2011, 11, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.B.G.; Gros, M.; van Neerven, R.J.J.; Faas, M.M.; de Vos, P. Immunomodulating properties of protein hydrolysates for application in cow’s milk allergy. Pediat. Allergy Immunol.-UK 2015, 26, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Mendis, E.; Rajapakse, N.; Byun, H.G.; Kim, S.K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Ukeda, H.; Ochi, H. Isolation and characterization of free radical scavenging activities peptides derived from casein. J. Nutr. Biochem. 2000, 11, 128–131. [Google Scholar] [CrossRef]
- Nan, Y.H.; Park, K.H.; Jeon, Y.J.; Park, Y.; Park, I.S.; Hahm, K.S.; Shin, S.Y. Antimicrobial and anti-inflammatory activities of a Leu/Lys-rich antimicrobial peptide with Phe-peptoid residues. Protein Pept. Lett. 2007, 14, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Bamdad, B.; Shin, S.H.; Suh, J.W.; Nimalaratne, C.; Sunwoo, H. Anti-Inflammatory and antioxidant properties of casein hydrolysate produced using high hydrostatic pressure combined with proteolytic enzymes. Molecules 2017, 22, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Benkhelifa, H.; Bengoa, C.; Larre, C.; Guibal, E.; Popineau, Y.; Legrand, J. Casein hydrolysis by immobilized enzymes in a torus reactor. Process Biochem. 2005, 40, 461–467. [Google Scholar] [CrossRef]
- Schaegger, H. Tricine-SDS-PAGE. Nat. Protoc. 2006, 1, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Fogarasi, A.L.; Kun, S.; Tanko, G.; Stefanovits-Banyai, E.; Hegyesne-Vecseri, B. A comparative assessment of antioxidant properties, total phenolic content of einkorn, wheat, barley and their malts. Food Chem. 2015, 167, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [n-15]-labeled nitrate in biological-fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Chanput, W.; Mes, J.; Vreeburg, R.A.M.; Sayelkoul, H.F.J.; Wichers, H.J. Transcription profiles of LPS-stimulated THP-1 monocytes and macrophages: A tool to study inflammation modulating effects of food-derived compounds. Food Funct. 2010, 1, 254–261. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Sources | Proteolytic Activity a | Optimum Conditions |
|---|---|---|---|
| Neutrase | Protease from Bacillus amyloliquefaciens | ≥0.8 U/g | pH 7; 37 °C |
| Alcalase | Protease from Bacillus licheniformis | ≥2.4 U/g | pH 7; 50 °C |
| Savinase | Protease from Bacillus sp. | ≥16 U/g | pH 7; 55 °C |
| Elastase | Elastase from hog pancreas | ≥4 U/mg | pH 8; 37 °C |
| Thermolysin | Protease from Bacillus thermoproteolyticus | 14 U/mg | pH 7; 50 °C |
| Trypsin | Protease derived from porcine pancreas | 30 U/g | pH 7; 37 °C |
| Samples | Atmospheric Hydrolysis | High Hydrostatic Pressure Hydrolysis | ||||
|---|---|---|---|---|---|---|
| <500 Da | 500–1500 Da | 1500–3000 Da | <500 Da | 500–1500 Da | 1500–3000 Da | |
| Neu | 6.88 | 74.38 | 18.75 | 16.18 | 52.21 | 31.60 |
| Alc | 27.24 | 28.65 | 44.11 | 71.15 | 28.38 | 0.47 |
| Sav | 0.16 | 34.84 | 65.00 | 0.26 | 63.44 | 36.30 |
| Ela | 44.42 | 39.02 | 16.55 | 38.66 | 44.70 | 16.64 |
| Ther | 25.45 | 74.55 | 0.00 | 67.06 | 32.94 | 0.00 |
| Try | 68.47 | 28.20 | 3.33 | 23.59 | 63.76 | 12.65 |
| Sequence | Ion (m/z) | Observed Mass | Calculated Mass | Source (Fragment) |
|---|---|---|---|---|
| GTWYSL | 726.35 | 725.35 | 725.34 | β-lactoglobulin (33–38) |
| LSFNPTQL | 919.49 | 918.48 | 918.48 | β-lactoglobulin (165–172) |
| MAASDISLL | 936.47 | 935.46 | 935.46 | β-lactoglobulin (40–48) |
| AMAASDISLL | 1007.49 | 1006.48 | 1006.50 | β-lactoglobulin (39–48) |
| DTDYKKYLLF | 653.35 (2) | 1304.68 | 1304.66 | β-lactoglobulin (112–121) |
| IIAEKTKIPAVF | 665.41 (2) | 1328.81 | 1328.80 | β-lactoglobulin (87–98) |
| DIQKVAGTWYSL | 690.86 (2) | 1379.72 | 1379.70 | β-lactoglobulin (27–38) |
| ELKPTPEGDLEIL | 727.39 (2) | 1452.77 | 1452.77 | β-lactoglobulin (61–73) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bamdad, F.; Bark, S.; Kwon, C.H.; Suh, J.-W.; Sunwoo, H. Anti-Inflammatory and Antioxidant Properties of Peptides Released from β-Lactoglobulin by High Hydrostatic Pressure-Assisted Enzymatic Hydrolysis. Molecules 2017, 22, 949. https://doi.org/10.3390/molecules22060949
Bamdad F, Bark S, Kwon CH, Suh J-W, Sunwoo H. Anti-Inflammatory and Antioxidant Properties of Peptides Released from β-Lactoglobulin by High Hydrostatic Pressure-Assisted Enzymatic Hydrolysis. Molecules. 2017; 22(6):949. https://doi.org/10.3390/molecules22060949
Chicago/Turabian StyleBamdad, Fatemeh, Seonghee Bark, Chul Hee Kwon, Joo-Won Suh, and Hoon Sunwoo. 2017. "Anti-Inflammatory and Antioxidant Properties of Peptides Released from β-Lactoglobulin by High Hydrostatic Pressure-Assisted Enzymatic Hydrolysis" Molecules 22, no. 6: 949. https://doi.org/10.3390/molecules22060949
APA StyleBamdad, F., Bark, S., Kwon, C. H., Suh, J.-W., & Sunwoo, H. (2017). Anti-Inflammatory and Antioxidant Properties of Peptides Released from β-Lactoglobulin by High Hydrostatic Pressure-Assisted Enzymatic Hydrolysis. Molecules, 22(6), 949. https://doi.org/10.3390/molecules22060949