
Adamantane in Drug Delivery Systems and Surface Recognition


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
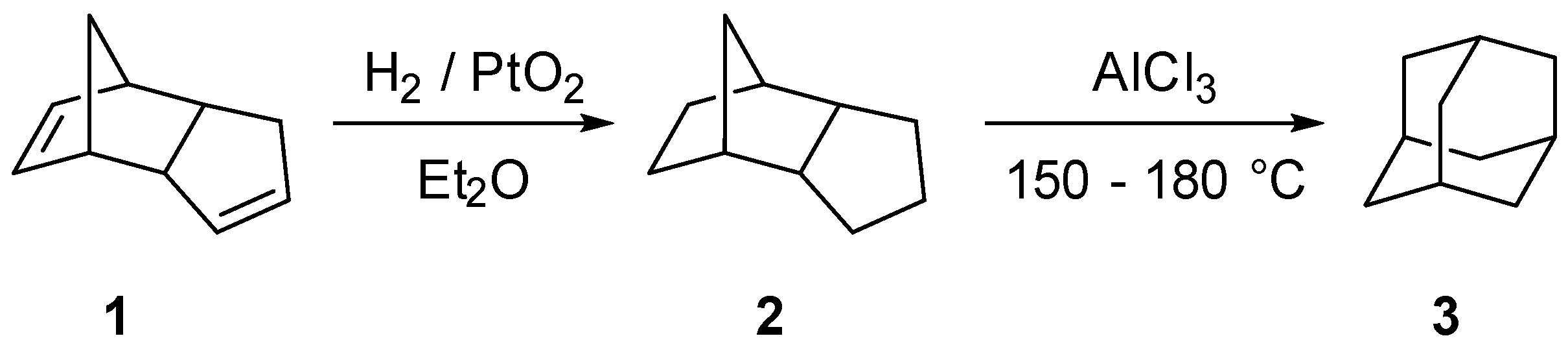
:1. Introduction
2. Adamantane Derivatives in Liposomes
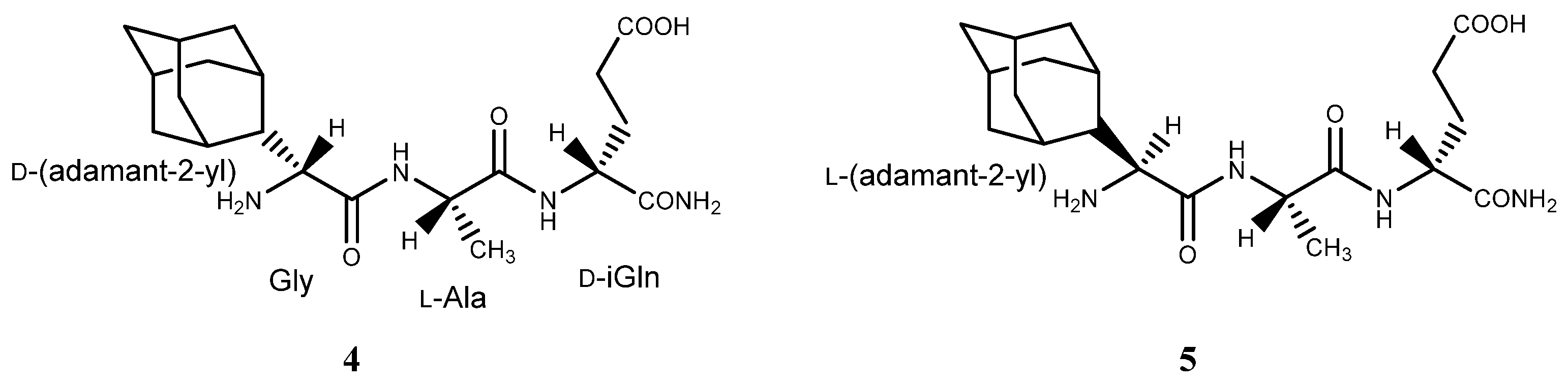
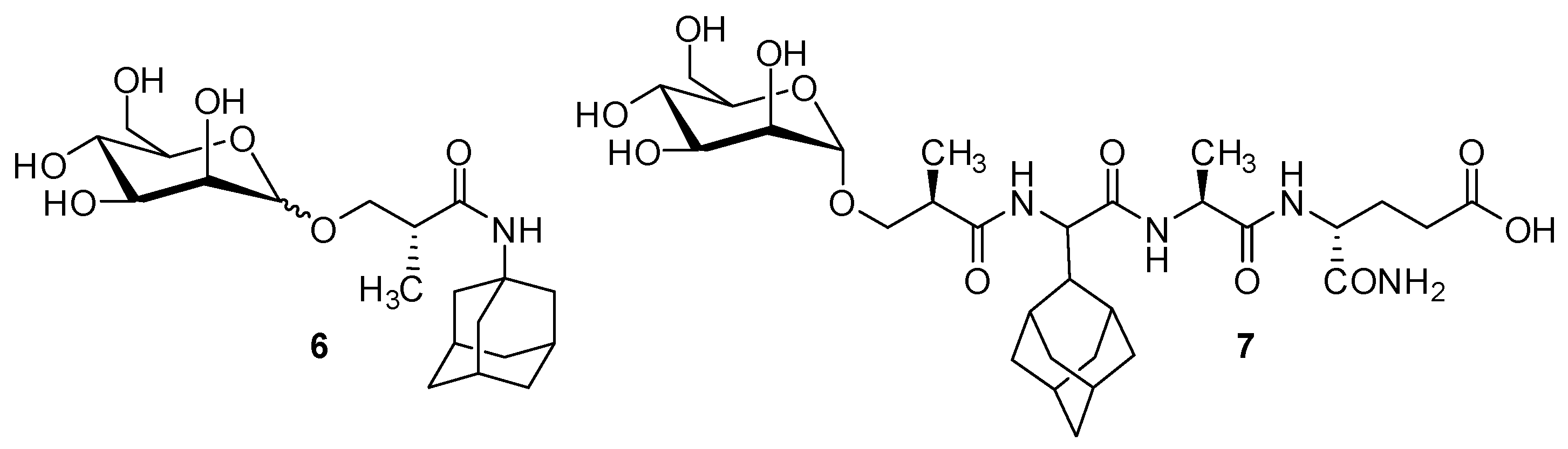
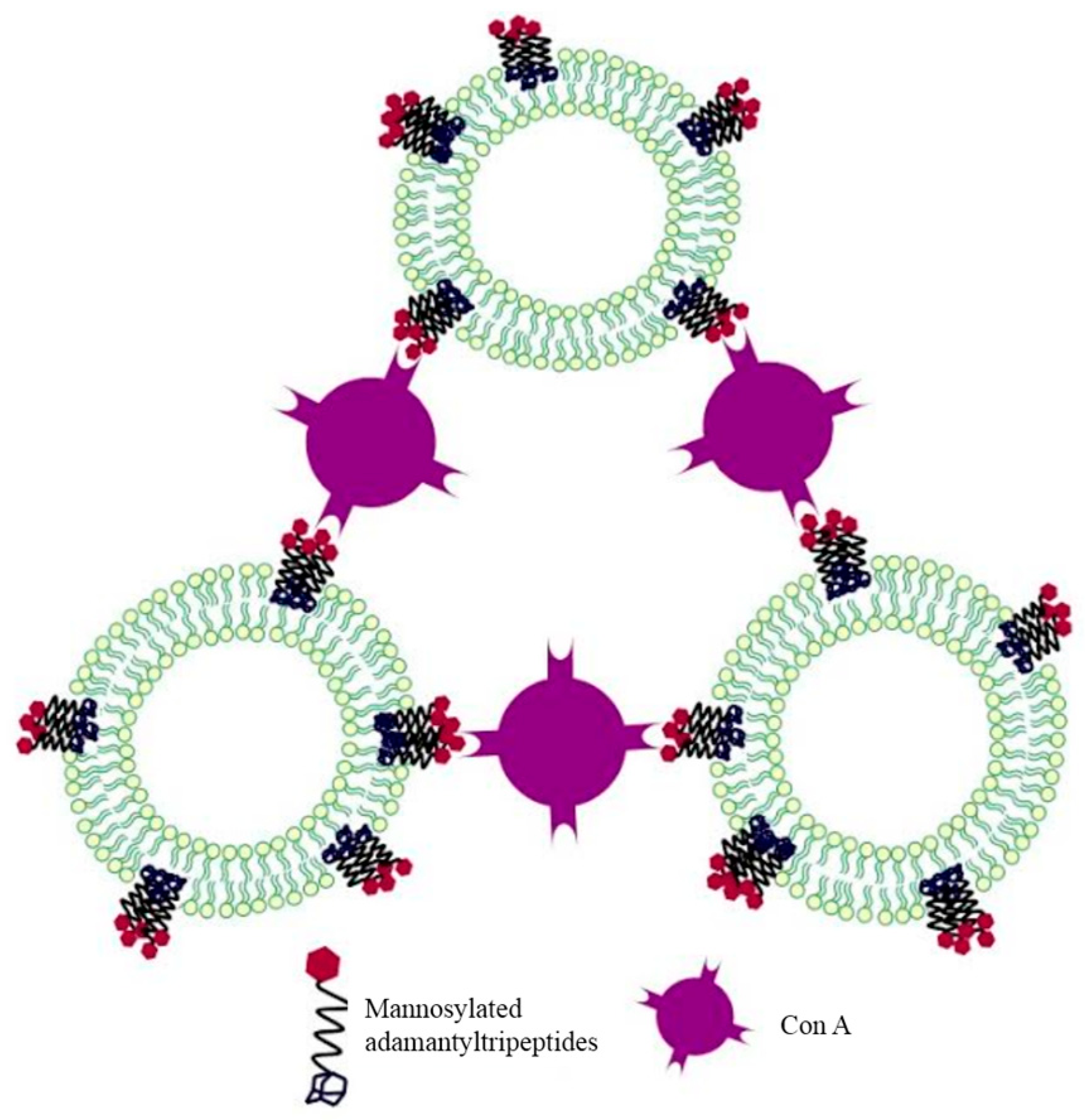
2.1. Adamantyl Peptides and Adamantyl Glycopeptides in Liposomes
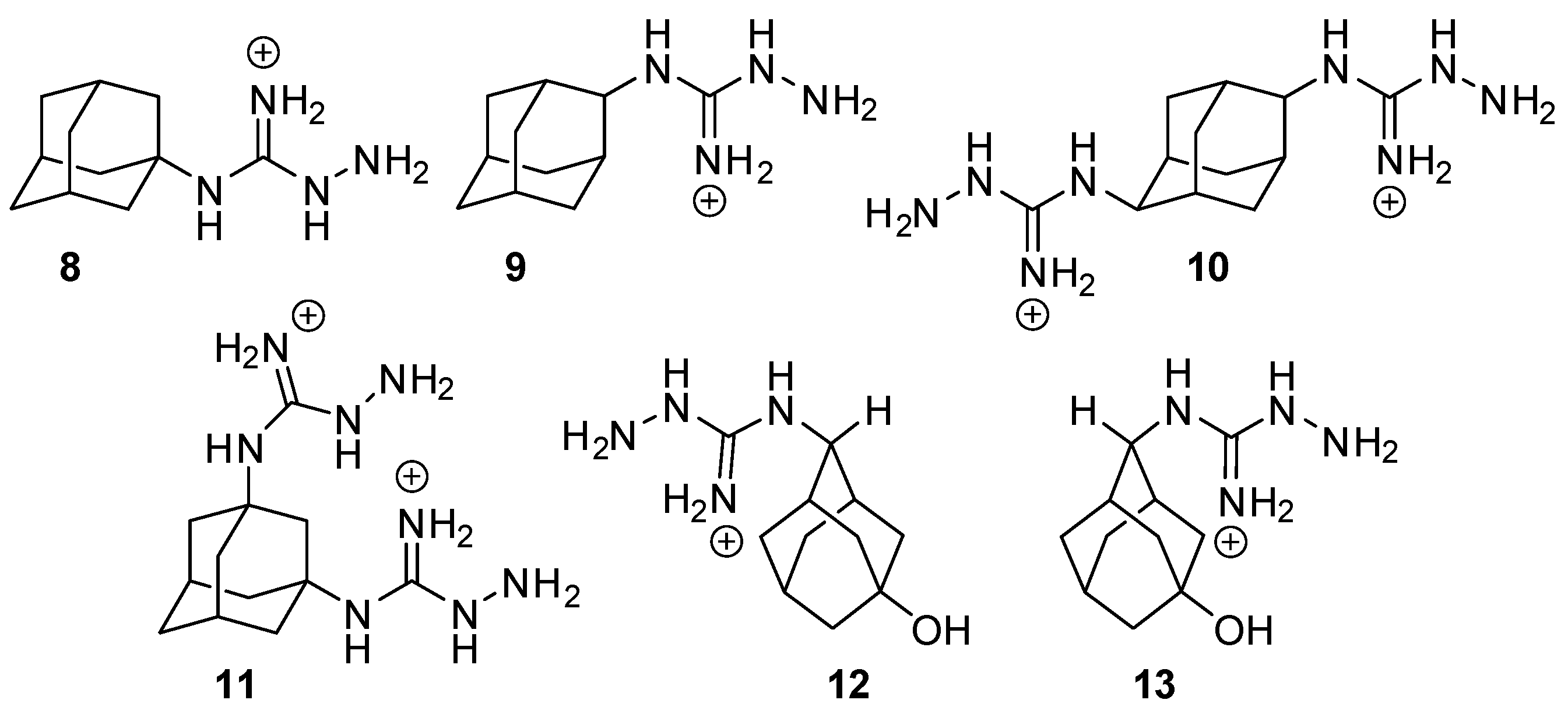
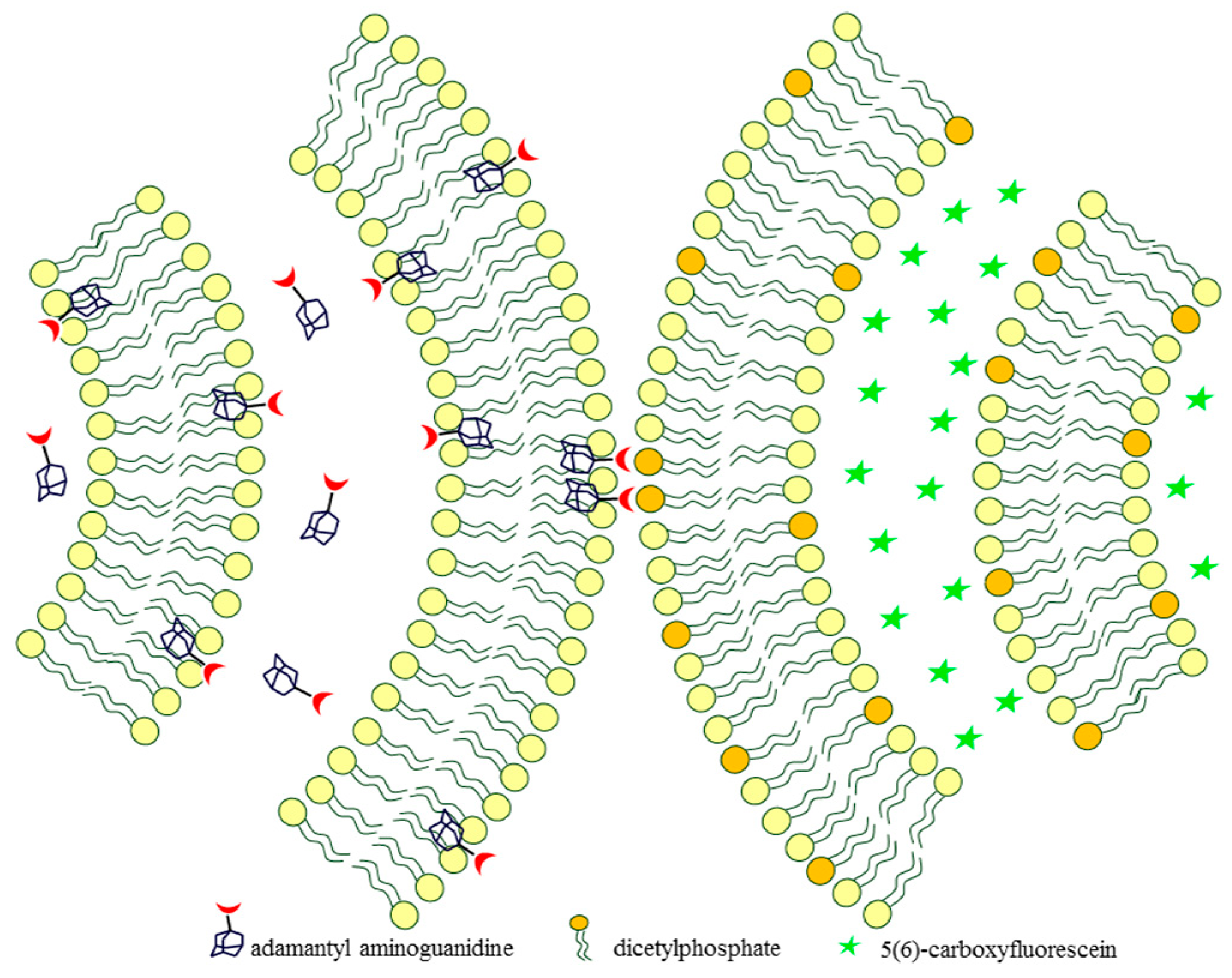
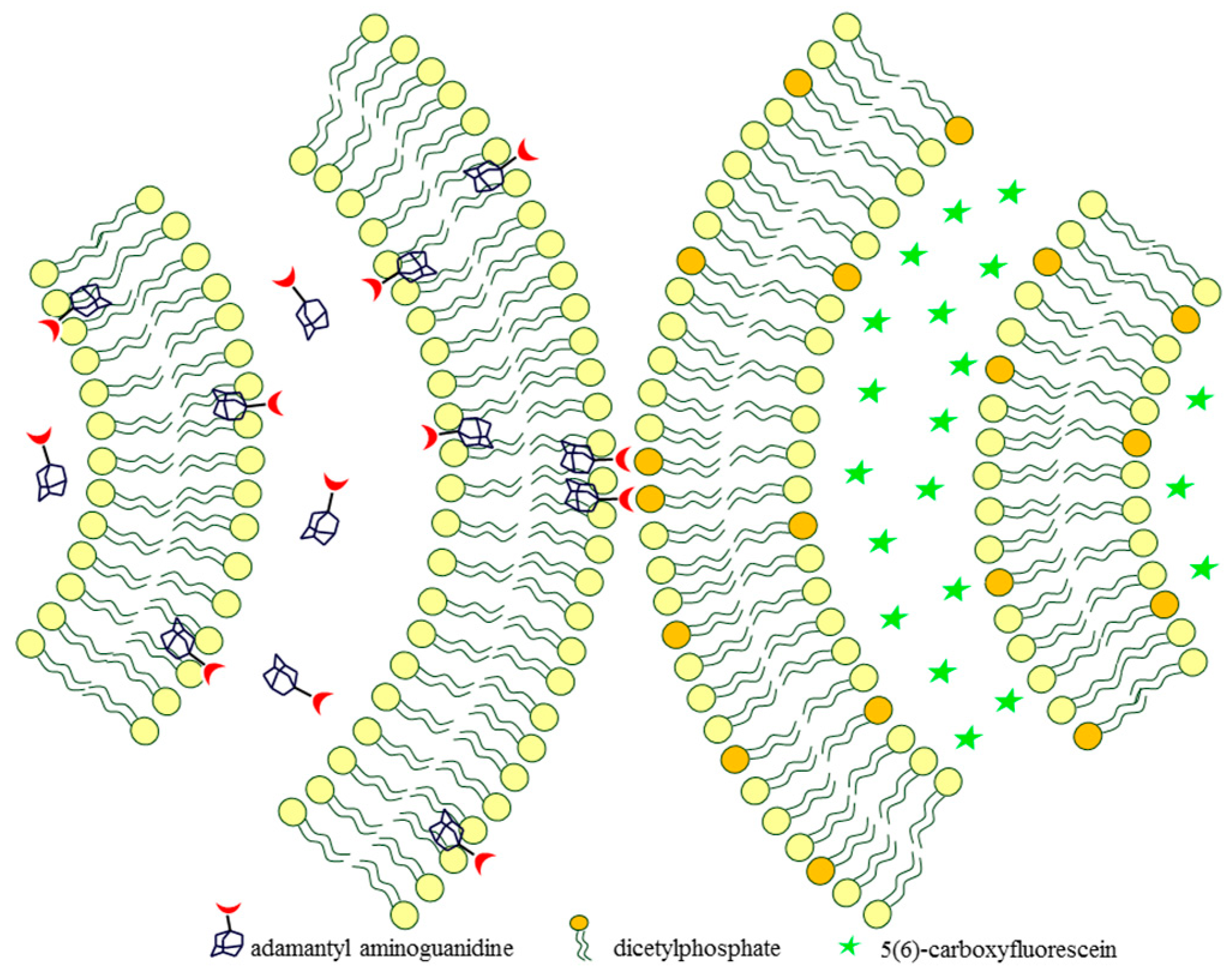
2.2. Adamantyl Aminoguanidines in Liposomes
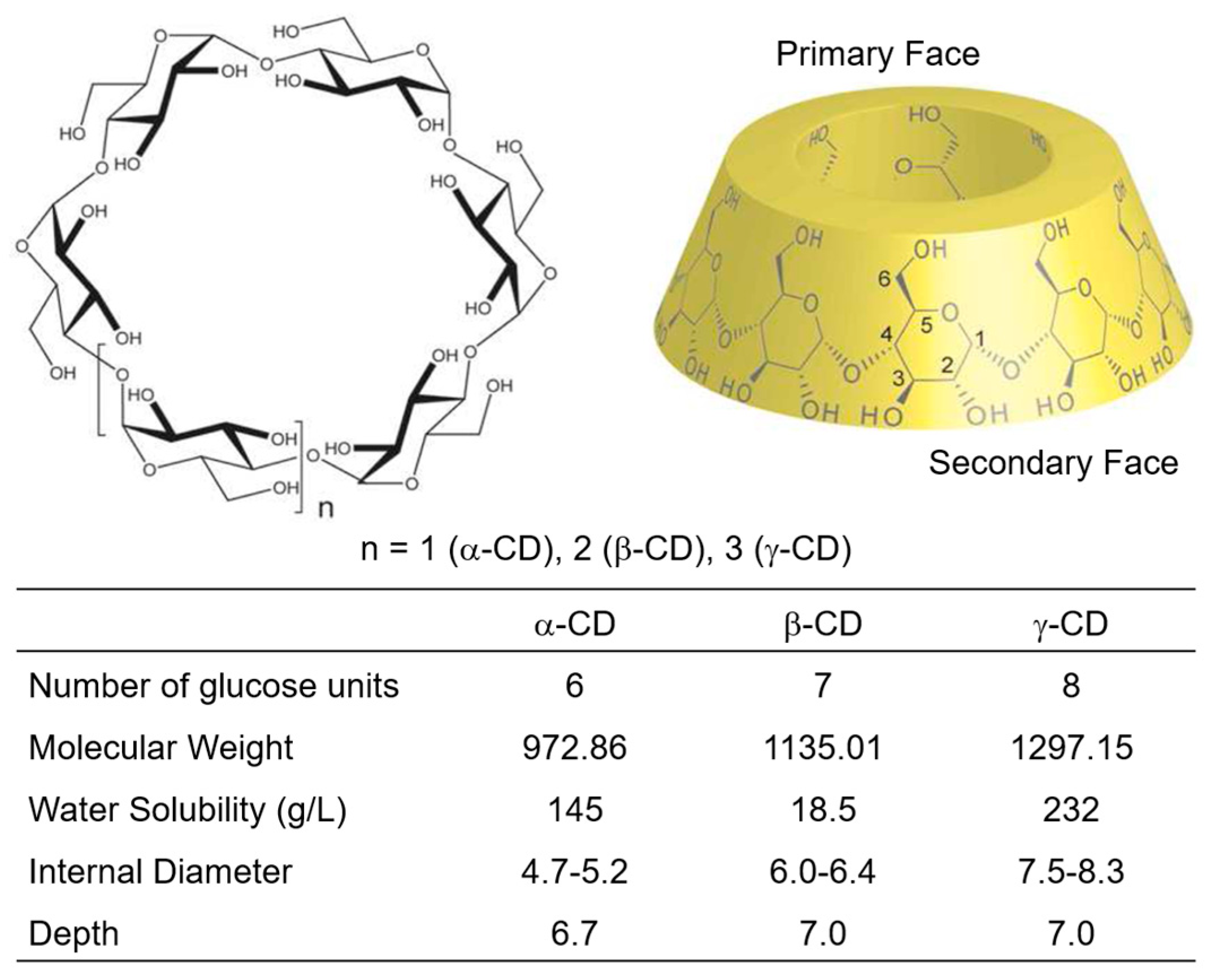
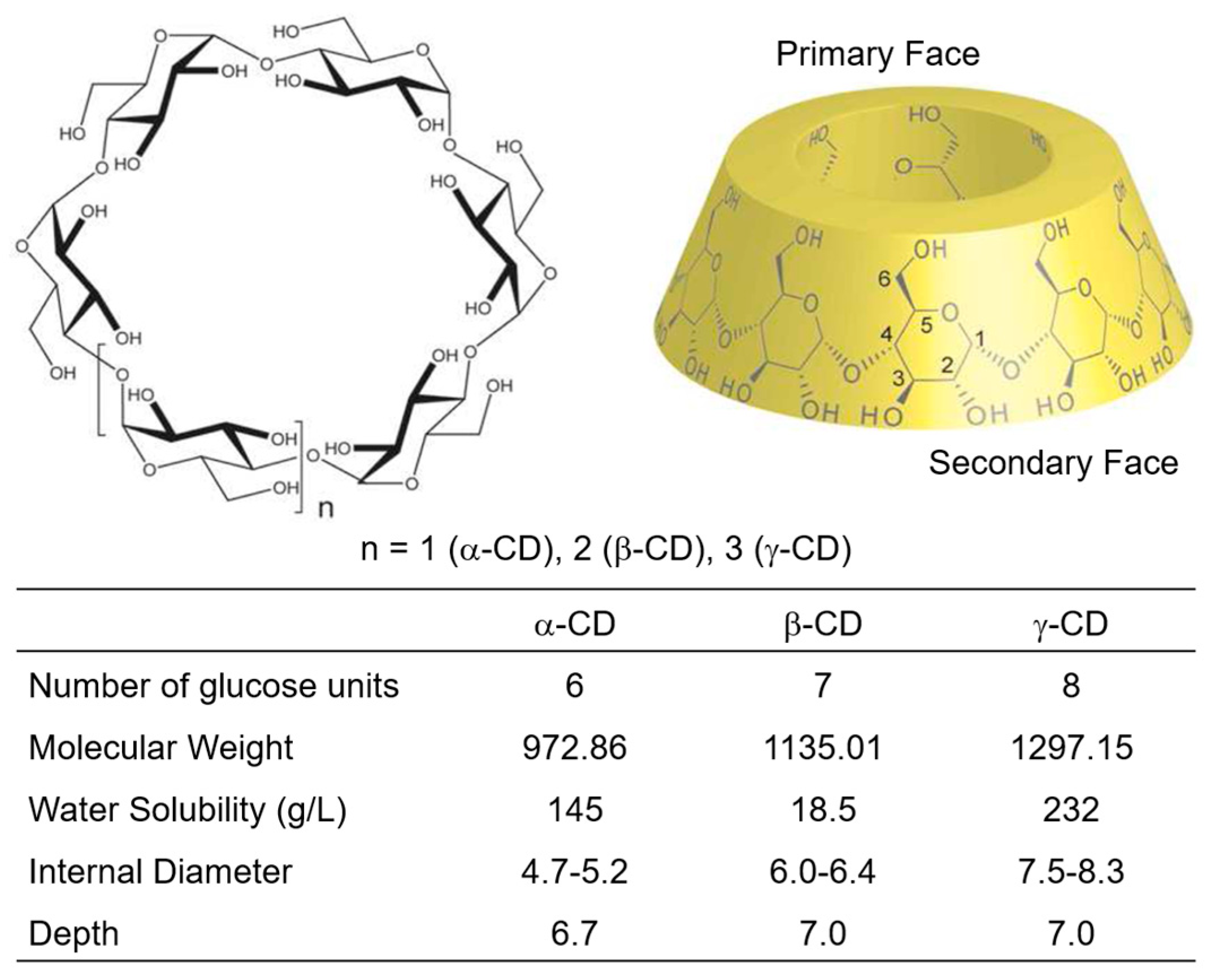
3. Adamantyl Cyclodextrin Complexes
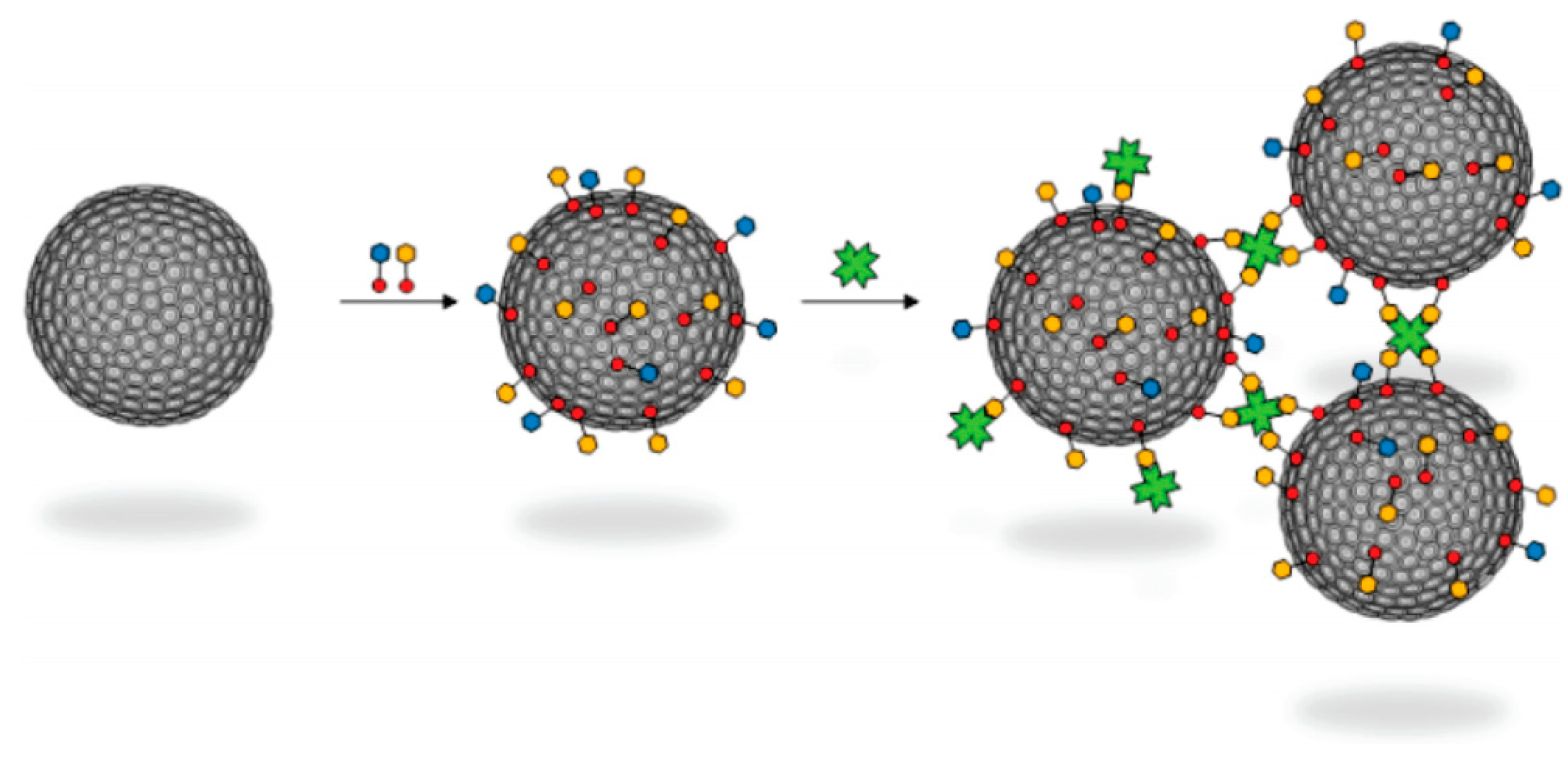
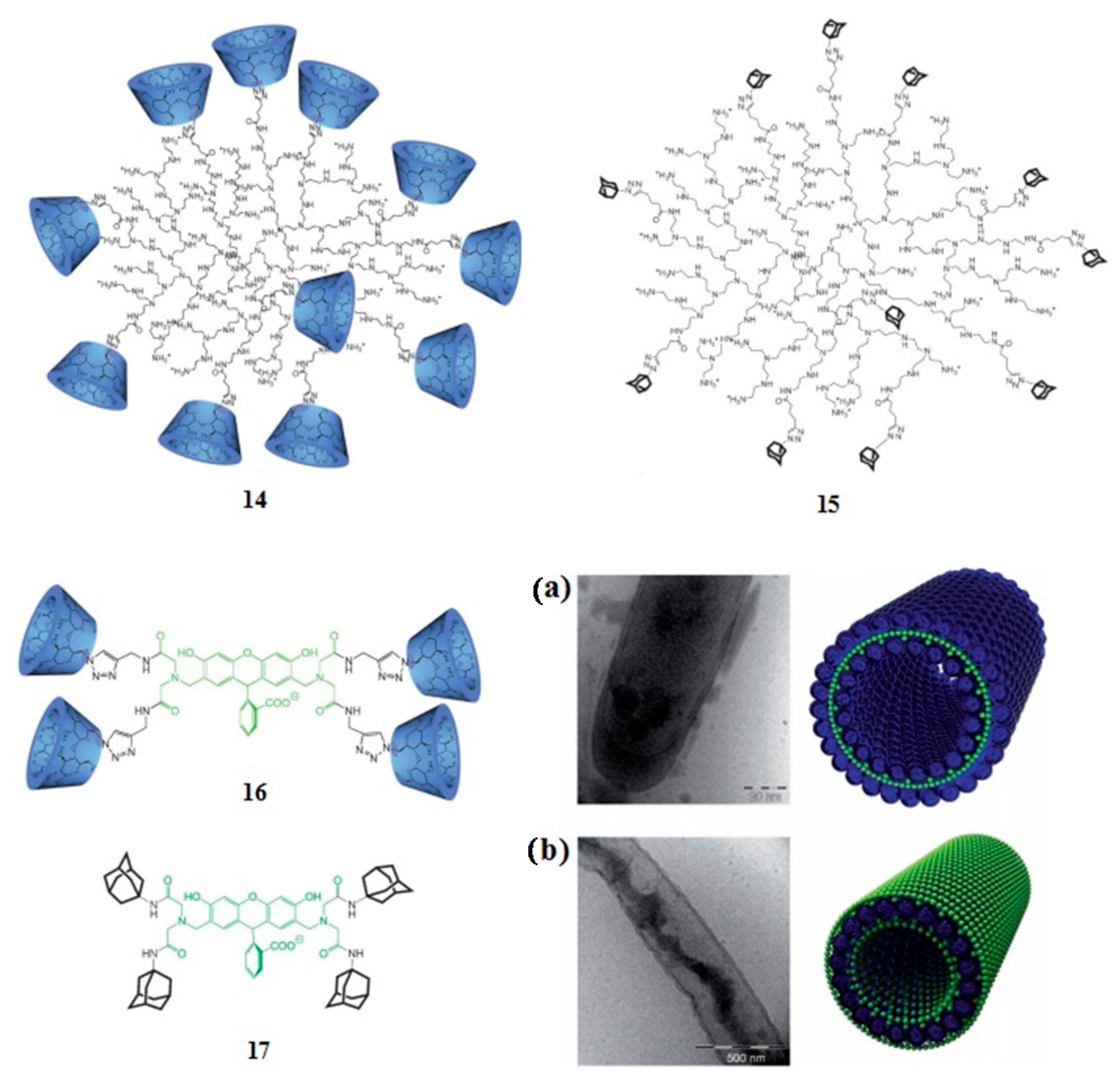
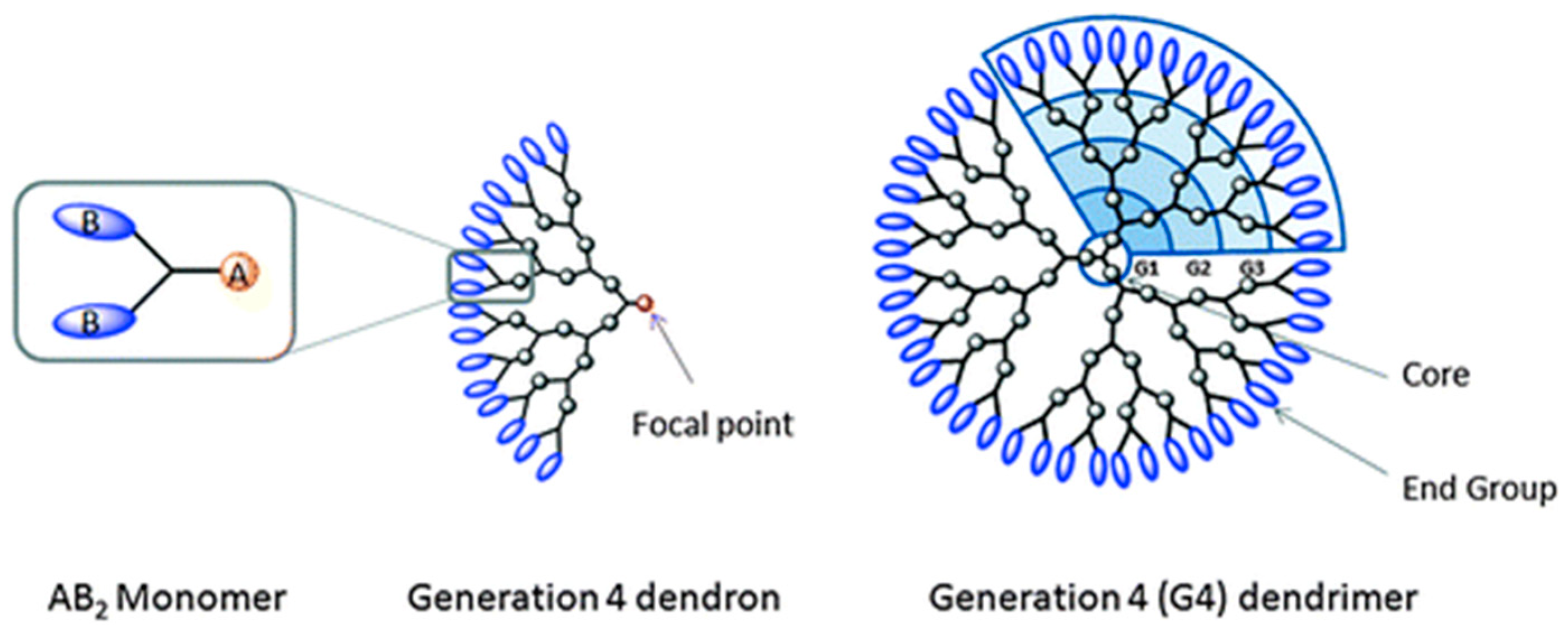
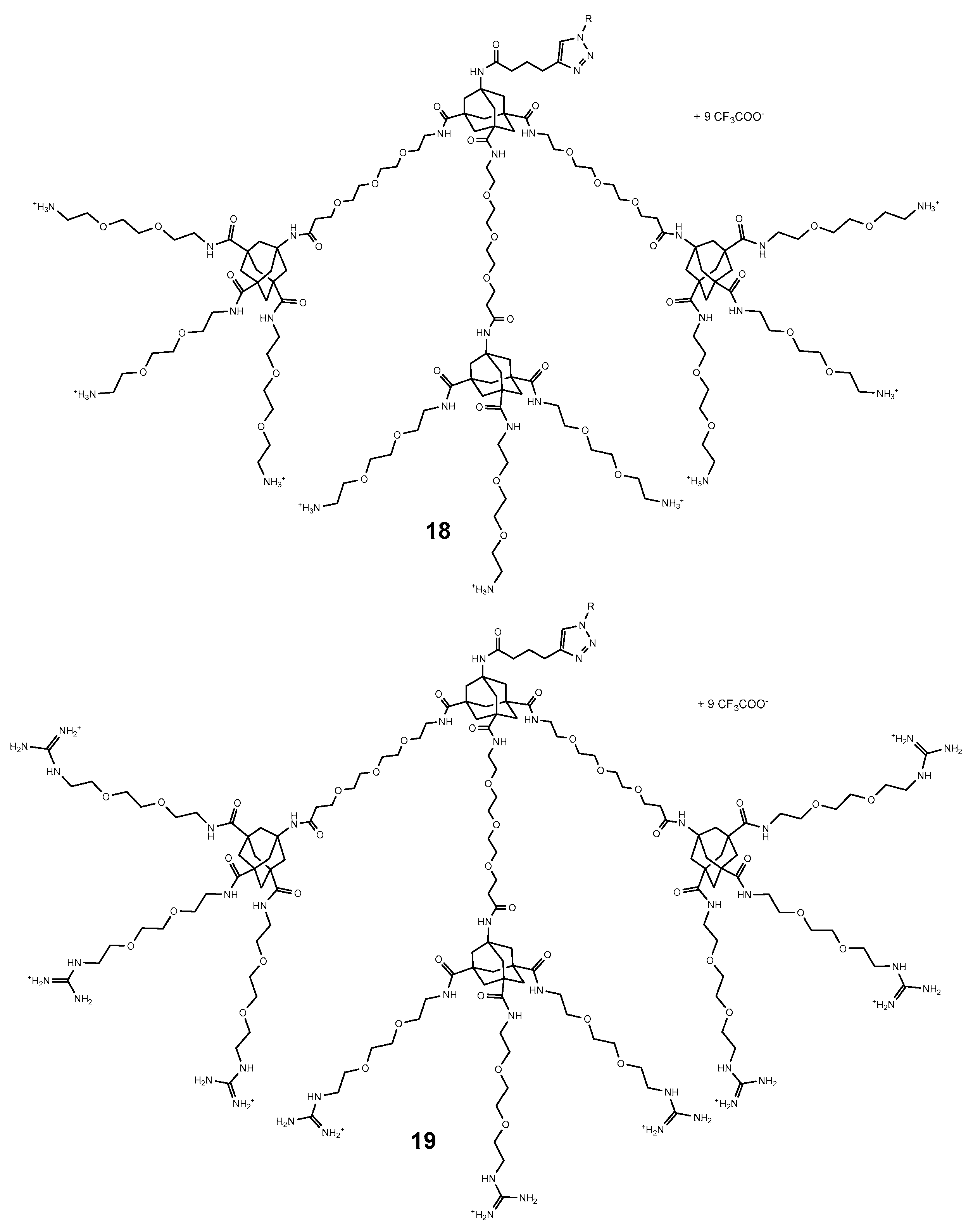
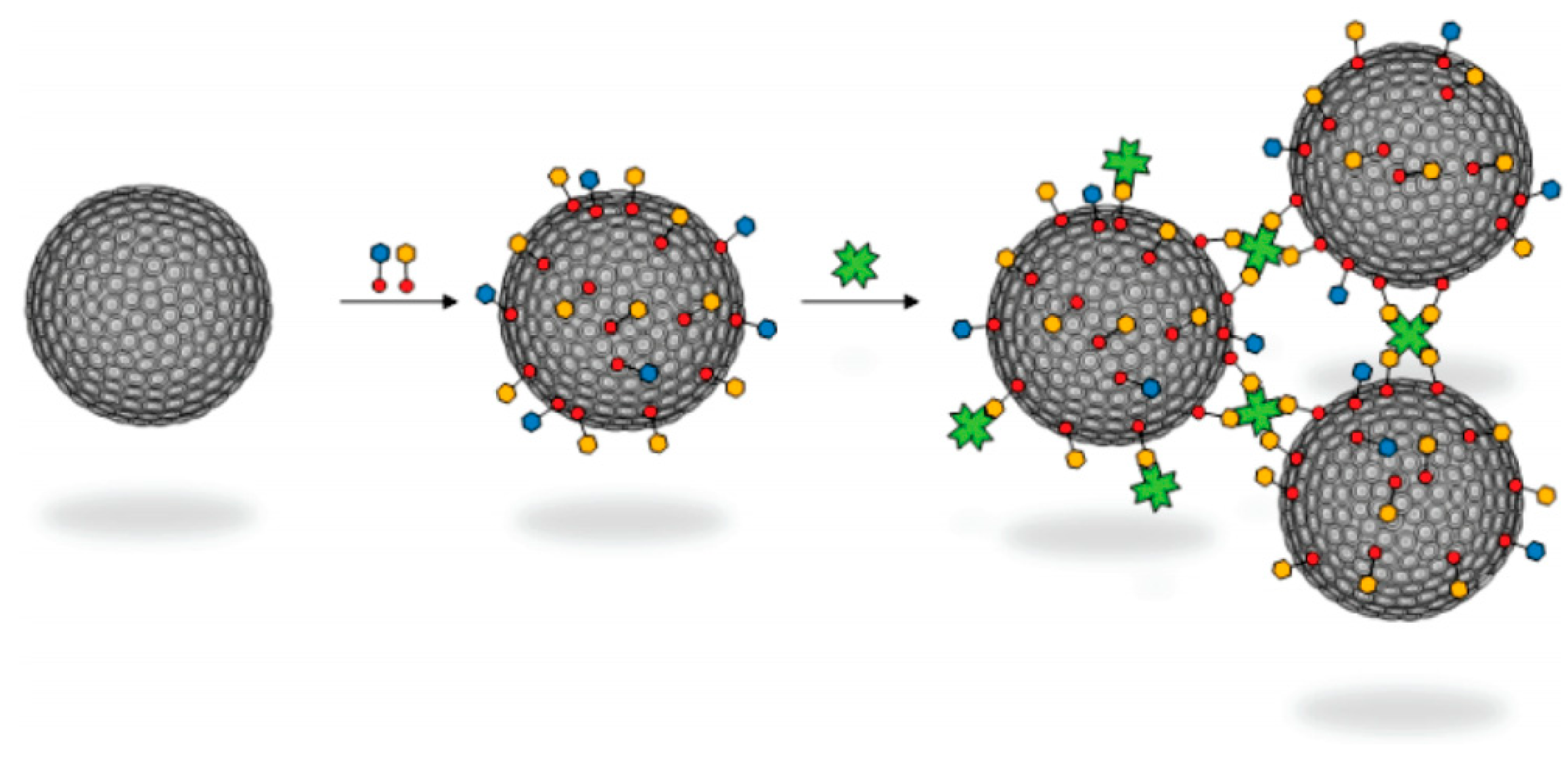
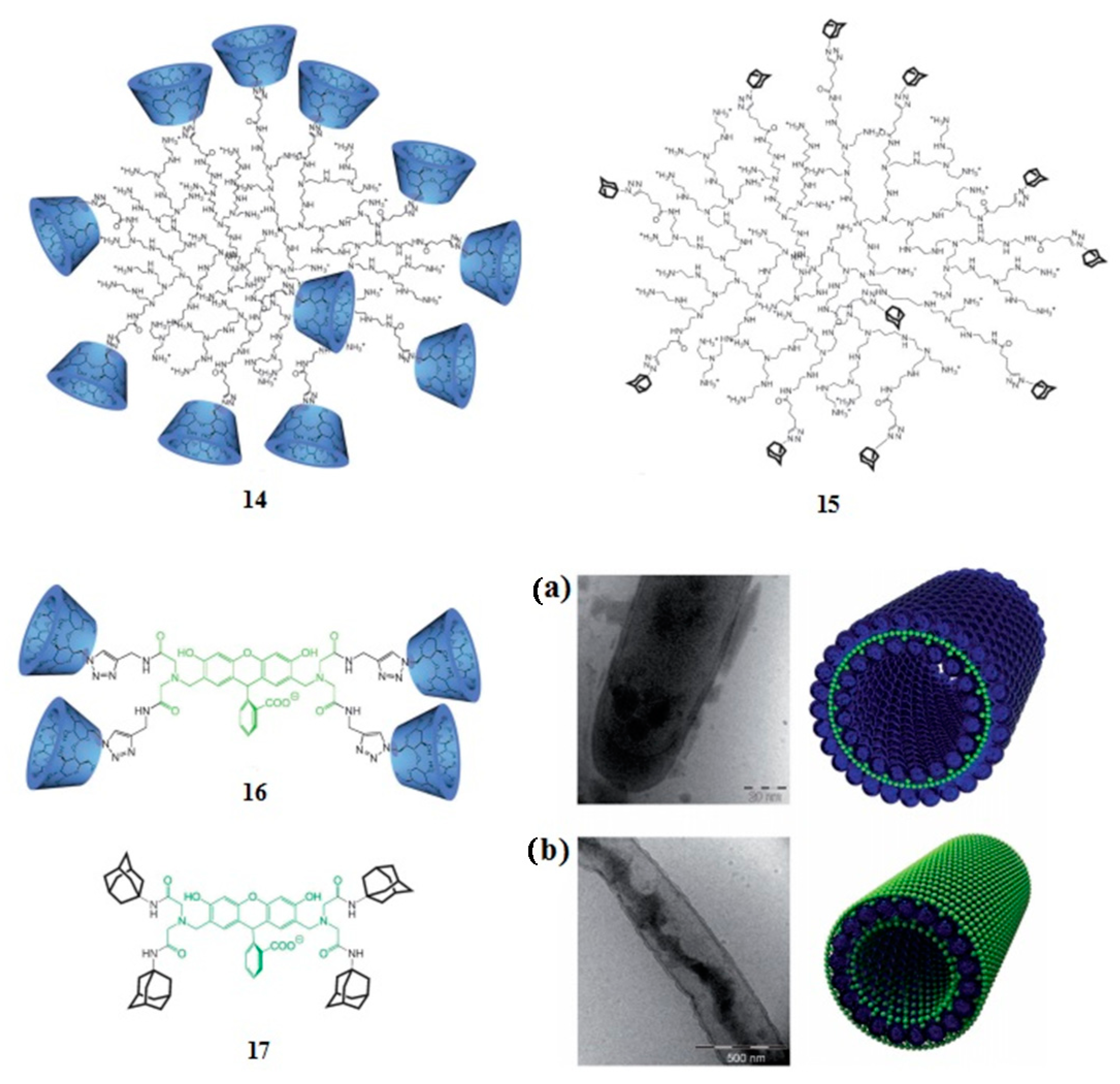
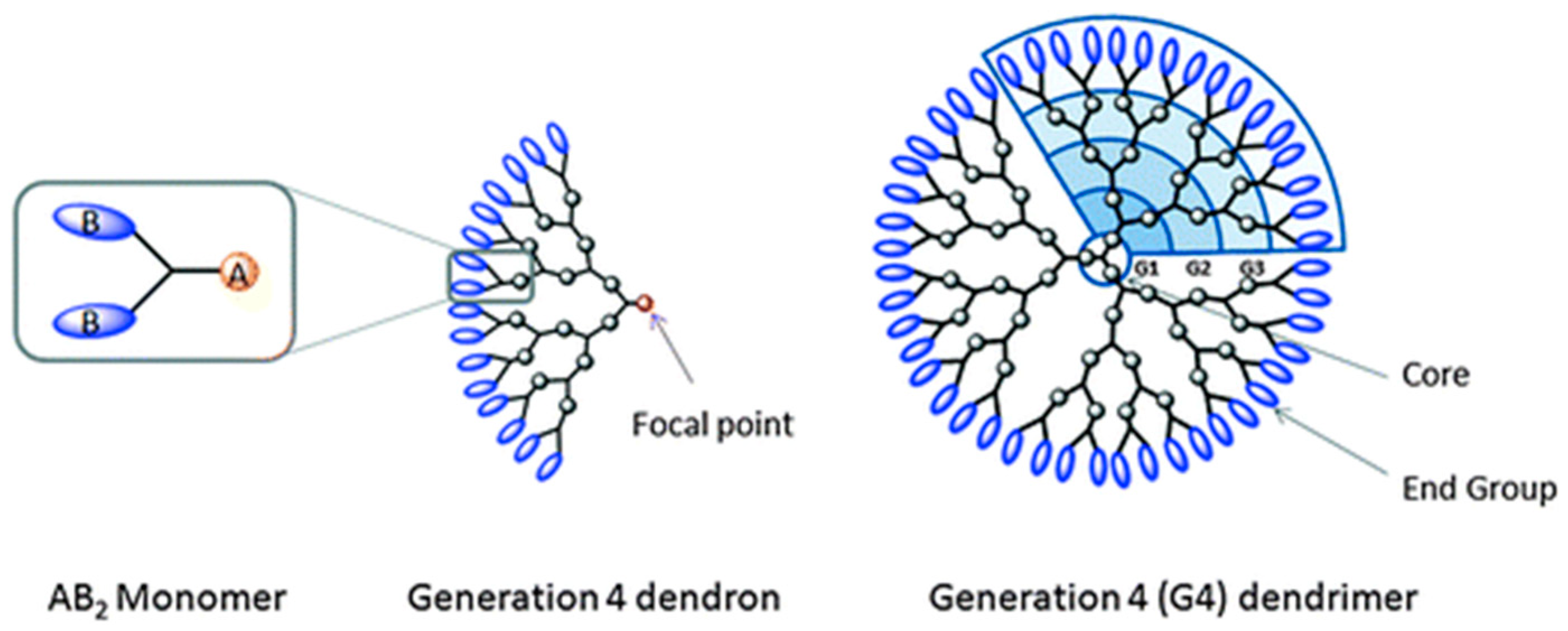
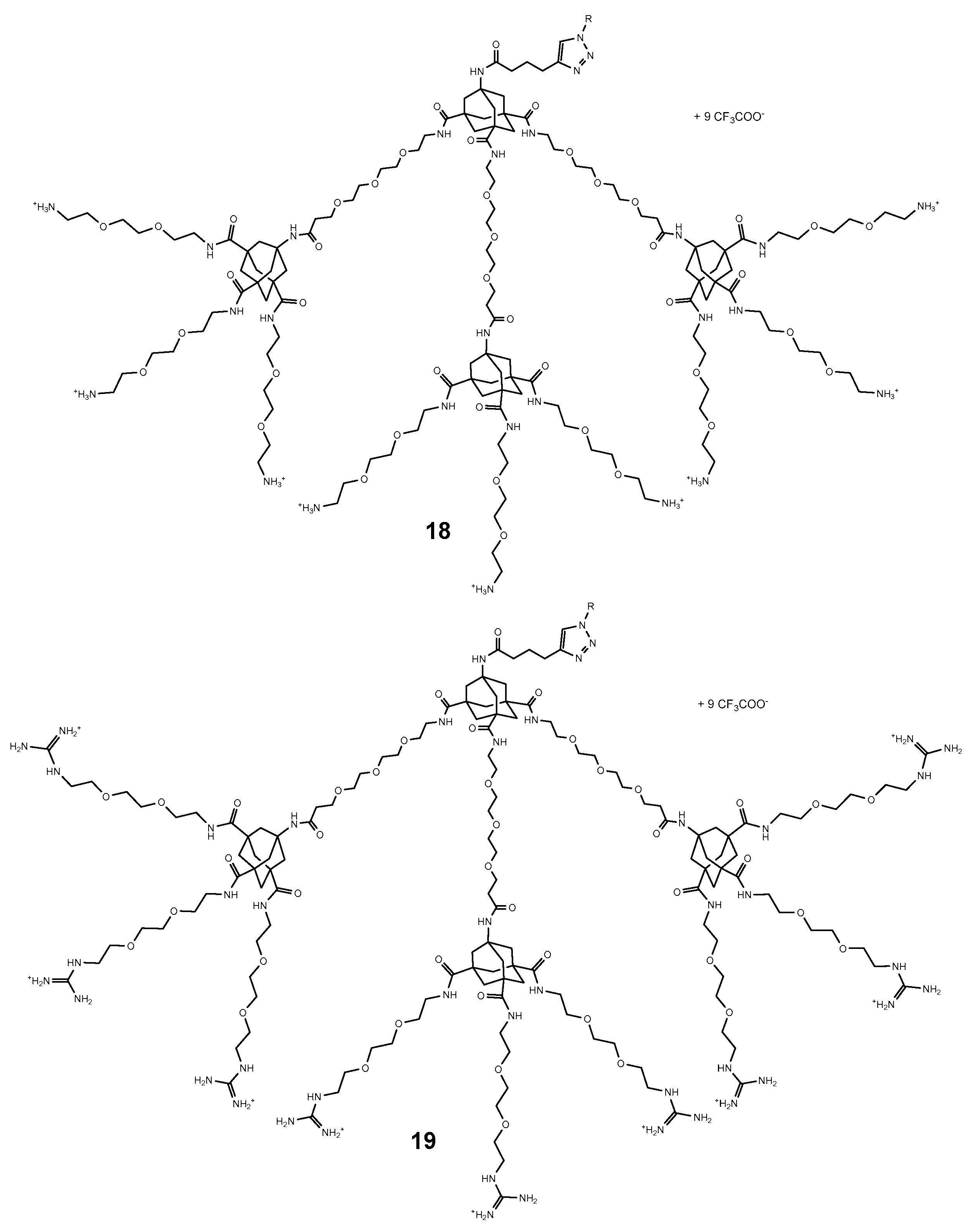
4. Adamantyl Dendrimers
5. Conclusions and Outlook
Acknowledgments
Conflicts of Interest
References
- Fort, R.C. Adamantane: The Chemistry of Diamond Molecules; Marcel Dekker: New York, NY, USA, 1976. [Google Scholar]
- Schwertfeger, H.; Fokin, A.A.; Schreiner, P.R. Diamonds are a chemist’s best friend. Diamondoid chemistry beyond adamantane. Angew. Chem. Int. Ed. 2008, 47, 1022–1036. [Google Scholar] [CrossRef] [PubMed]
- Gunawan, M.A.; Hierso, J.-C.; Poinsot, D.; Fokin, A.A.; Fokina, N.A.; Tkachenko, B.A.; Schreiner, P.R. Diamondoids: Functionalization and subsequent applications of perfectly defined molecular cage hydrocarbons. New J. Chem. 2014, 38, 28–41. [Google Scholar] [CrossRef]
- Landa, S.; Macháček, V. Sur l’adamantane, nouvel hydrocarbure extrait du naphte. Collect. Czechoslov. Chem. Commun. 1933, 5, 1–5. [Google Scholar] [CrossRef]
- Prelog, V.; Seiwerth, R. Über die Synthese des Adamantans. Ber. Dtsch. Chem. Ges. 1941, 74, 1644–1648. [Google Scholar] [CrossRef]
- Schleyer, P.v.R. A Simple Preparation of Adamantane. J. Am. Chem. Soc. 1957, 79, 3292. [Google Scholar] [CrossRef]
- Schleyer, P.v.R.; Donaldson, M.M.; Nicholas, R.D.; Cupas, C. Adamantane. Org. Synth. 1962, 42, 8. [Google Scholar]
- Fort, R.C.; Schleyer, P.v.R. Adamantane: Consequences of the Diamondoid Structure. Chem. Rev. 1964, 64, 277–300. [Google Scholar] [CrossRef]
- Moiseev, I.K.; Makarova, N.V.; Zemtsova, M.N. Reactions of adamantanes in electrophilic media. Russ. Chem. Rev. 1999, 68, 1001–1020. [Google Scholar] [CrossRef]
- Davies, W.L.; Grunert, R.R.; Haff, R.F.; McGahen, J.W.; Neumayer, E.M.; Paulshock, M.; Watts, J.C.; Wood, T.R.; Hermann, E.C.; Hoffmann, C.E. Antiviral Activity of 1-Adamantanamine (Amantadine). Science 1964, 144, 862–863. [Google Scholar] [CrossRef] [PubMed]
- Dolin, R.; Reichman, R.C.; Madore, H.P.; Maynard, R.; Linton, P.N.; Webber-Jones, J. A controlled trial of amantadine and rimantadine in the prophylaxis of influenza A infection. N. Engl. J. Med. 1982, 307, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Obando, D.; Liao, V.; Lifa, T.; Codd, R. The many faces of the adamantyl group in drug design. Eur. J. Med. Chem. 2011, 46, 1949–1963. [Google Scholar] [CrossRef] [PubMed]
- Wanka, L.; Iqbal, K.; Schreiner, P.R. The Lipophilic Bullet Hits the Targets: Medicinal Chemistry of Adamantane Derivatives. Chem. Rev. 2013, 113, 3516–3604. [Google Scholar] [CrossRef] [PubMed]
- Horvat, Š.; Mlinarić-Majerski, K.; Glavaš-Obrovac, L.; Jakas, A.; Veljković, J.; Marczi, S.; Kragol, G.; Roščić, M.; Matković, M.; Milostić-Srb, A. Tumor-Cell-Targeted Methionine-enkephalin Analogues Containing Unnatural Amino Acids: Design, Synthesis, and in Vitro Antitumor Activity. J. Med. Chem. 2006, 49, 3136–3142. [Google Scholar] [CrossRef] [PubMed]
- Roščić, M.; Sabljić, V.; Mlinarić-Majerski, K.; Horvat, Š. In Vitro Enzymatic Stabilities of Methionine-enkephalin Analogues Containing an Adamantane-type Amino Acid. Croat. Chem. Acta 2008, 81, 637–640. [Google Scholar]
- Perl, A.; Gomez-Casado, A.; Thompson, D.; Dam, H.H.; Jonkheijm, P.; Reinhoudt, D.N.; Huskens, J. Gradient-driven motion of multivalent ligand molecules along a surface functionalized with multiple receptors. Nat. Chem. 2011, 3, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.-W.; Zhou, H.; Zhou, S.-T.; Yuan, J.-Y. Macromolecules based on recognition between cyclodextrin and guest molecules: Synthesis, properties and functions. Eur. Polym. J. 2015, 65, 63–81. [Google Scholar] [CrossRef]
- De Clercq, E. Antiviral agents active against Influenza A viruses. Nat. Rev. Drug Discov. 2006, 5, 1015–1025. [Google Scholar] [CrossRef] [PubMed]
- Schnell, J.R.; Chou, J.J. Structure and mechanism of the M2 proton channel of Influenza A virus. Nature 2008, 451, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Cady, S.D.; Luo, W.; Hu, F.; Hong, M. Structure and Function of the Influenza A M2 Proton Channel. Biochemistry 2009, 48, 7356–7364. [Google Scholar] [CrossRef] [PubMed]
- Chew, C.F.; Guy, A.; Biggin, P.C. Distribution and dynamics of adamantanes in a lipid bilayer. Biophys. J. 2008, 95, 5627–5636. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.Y.S.; Rutka, J.T.; Chan, W.C.W. Nanomedicine. N. Engl. J. Med. 2010, 363, 2434–2443. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhao, Y. Biomedical Applications of Supramolecular Systems Based on Host-Guest Interactions. Chem. Rev. 2015, 115, 7794–7839. [Google Scholar] [CrossRef] [PubMed]
- Mashaghi, S.; Jadidi, T.; Koenderink, G.; Mashaghi, A. Lipid Nanotechnology. Int. J. Mol. Sci. 2013, 14, 4242–4282. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.M.; Cullis, P.R. Liposomal drug delivery systems: From concept to clinical applications. Adv. Drug Deliv. Rev. 2013, 65, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Antonietti, M.; Foerster, S. Vesicles and liposomes: A self-assembly principle beyond lipids. Adv. Mater. 2003, 15, 1323–1333. [Google Scholar] [CrossRef]
- Sugrue, R.J.; Hay, A.J. Structural characteristics of the M2 protein of Influenza A viruses: Evidence that it forms a tetrameric channel. Virology 1991, 180, 617–624. [Google Scholar] [CrossRef]
- Duff, K.C.; Cudmore, A.J.; Bradshaw, J.P. The location of amantadine hydrochloride and free base within phospholipid multilayers: A neutron and X-ray diffraction study. Biochim. Biophys. Acta Biomembr. 1993, 1145, 149–156. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Wojas, J.; Pezeshk, V.; Pezeshk, A. Partitioning and Localization of Spin-Labeled Amantadine in Lipid Bilayers: An EPR Study. J. Pharm. Sci. 1998, 87, 1249–1254. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Schnell, J.R.; Chou, J.J. Amantadine partition and localization in phospholipid membrane: A solution NMR study. Biochem. Biophys. Res. Commun. 2004, 324, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Štimac, A.; Šegota, S.; Dutour Sikirić, M.; Ribić, R.; Frkanec, L.; Svetličić, V.; Tomić, S.; Vranešić, B.; Frkanec, R. Surface modified liposomes by mannosylated conjugates anchored via the adamantyl moiety in the lipid bilayer. Biochim. Biophys. Acta Biomembr. 2012, 1818, 2252–2259. [Google Scholar] [CrossRef] [PubMed]
- Šekutor, M.; Štimac, A.; Mlinarić-Majerski, K.; Frkanec, R. Syntheses and characterization of liposome-incorporated adamantyl aminoguanidines. Org. Biomol. Chem. 2014, 12, 6005–6013. [Google Scholar] [CrossRef] [PubMed]
- Frkanec, R.; Vranešić, B.; Tomić, S. Adamantoylated biologically active small peptides and glycopeptides structurally related to the bacterial peptidoglycan. In Methods in Molecular Biology, Peptide Modifications to Increase Metabolic Stability and Activity; Cudic, P., Ed.; Springer Science Business Media: New York, NY, USA, 2013; Volume 1081, pp. 91–106. [Google Scholar]
- Keglević, D.; Ladešić, B.; Tomašić, J.; Valinger, Z.; Naumski, R. Isolation procedure and properties of monomer unit from lysozyme digest of peptidoglycan complex excreted into the medium by penicillin-treated Brevibacterium divaricatum mutant. Biochim. Biophys. Acta 1979, 585, 273–281. [Google Scholar] [CrossRef]
- Ellouz, F.; Adama, A.; Ciorbaru, R.; Lederer, E. Minimal structural requirements for adjuvant activity of bacterial peptidoglycan derivatives. Biochem. Biophys. Res. Commun. 1974, 59, 1317–1325. [Google Scholar] [CrossRef]
- Tomašić, J.; Hršak, I. Peptidoglycan monomer originating from B. Divaricatum: Its metabolism and biological activities in the host. In Surface Structures of Microorganisms and Their Interaction with the Mammalian Host; Schrinner, E., Richmond, M.H., Seibert, G., Schwartz, U., Eds.; VCH: Weinheim, Germany, 1987; pp. 113–121. [Google Scholar]
- Ljevaković, Đ.; Tomašić, J.; Šporec, V.; Halassy Špoljar, B.; Hanzl Dujmović, I. Synthesis of novel adamantylacetyl derivative of peptidoglycan monomer—Biological evaluation of immunomodulatory peptidoglycan monomer and respective derivatives with lipophilic substituents on amino group. Bioorg. Med. Chem. 2000, 8, 2441–2449. [Google Scholar] [CrossRef]
- Vranešić, B.; Tomašić, J.; Smerdel, S.; Kantoci, D.; Benedetti, F. Synthesis and Antiviral Activity of Novel Adamantylpeptides. Helv. Chim. Acta 1993, 76, 1752–1758. [Google Scholar] [CrossRef]
- Vranešić, B.; Tomašić, J.; Ljevaković, Đ.; Hršak, I. Biological activity of novel adamantyltripeptides with emphasis on their immunorestorative effect. In Immunotherapy of Infection; Masihi, K.N., Ed.; Marcel Dekker: New York, NY, USA, 1994; pp. 241–248. [Google Scholar]
- Frkanec, R.; Noethig-Laslo, V.; Vranešić, B.; Mirosavljević, K.; Tomašić, J. A spin labelling study of immunomodulating peptidoglycan monomer and adamantyltripeptides entrapped into liposomes. Biochim. Biophys. Acta Biomembr. 2003, 1611, 187–196. [Google Scholar] [CrossRef]
- Killion, J.J.; Fidler, I.J. Therapy of Cancer Metastasis by Tumoricidal Activation of Tissue Macrophages Using Liposome-Encapsulated Immunomodulators. Pharmacol. Ther. 1998, 78, 141–154. [Google Scholar] [CrossRef]
- Turánek, J.; Záluská, D.; Vacek, A.; Borkovcová, P.; Thurnvaldová, J.; Bláha, L.; Masek, L. Stimulation of nonspecific immunity, haemopoiesis and protection of mice against radiation injury by 1-adamantylamide-l-alanyl-d-isoglutamine incorporated in liposomes. Int. Immunopharmacol. 2001, 1, 167–175. [Google Scholar] [CrossRef]
- Ribić, R.; Habjanec, L.; Vranešić, B.; Frkanec, R.; Tomić, S. Synthesis and Biological Evaluation of New Mannose Derived Immunomodulating Adamantyltripeptides. Croat. Chem. Acta 2011, 84, 233–244. [Google Scholar] [CrossRef]
- Ribić, R.; Habjanec, L.; Vranešić, B.; Frkanec, R.; Tomić, S. Synthesis and immunostimulating properties of novel adamant-1-yl tripeptides. Chem. Biodivers. 2012, 9, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Ribić, R.; Habjanec, L.; Vranešić, B.; Frkanec, R.; Tomić, S. Influence of Mannosylation on Immunostimulating Activity of Adamant-1-yl Tripeptide. Chem. Biodivers. 2012, 9, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Lättig-Tunnemann, G.; Prinz, M.; Hofmann, D.; Behlke, J.; Palm-Apergi, C.; Morano, I.; Herce, H.D.; Cardoso, M.C. Backbone rigidity and static presentation of guanidinium groups increases cellular uptake of arginine-rich cell-penetrating peptides. Nat. Commun. 2011, 2, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Wheaten, S.A.; Ablan, F.D.O.; Spaller, B.L.; Trieu, J.M.; Almeida, P.F. Translocation of Cationic Amphipathic Peptides across the Membranes of Pure Phospholipid Giant Vesicles. J. Am. Chem. Soc. 2013, 135, 16517–16525. [Google Scholar] [CrossRef] [PubMed]
- Rozas, I.; Sánchez-Sanz, G.; Alkorta, I.; Elguero, J. Solvent effects on guanidinium-anion interactions and the problem of guanidinium Y-aromaticity. J. Phys. Org. Chem. 2013, 26, 378–385. [Google Scholar] [CrossRef]
- Šekutor, M.; Mlinarić-Majerski, K. Adamantyl aminoguanidines as receptors for oxo-anions. Tetrahedron Lett. 2014, 55, 6665–6670. [Google Scholar] [CrossRef]
- Tiwari, G.; Tiwari, R.; Rai, A.K. Cyclodextrins in delivery systems: Applications. J. Pharm. Bioallied Sci. 2010, 2, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, H.J.; Schollmeyer, E.J. Application of cyclodextrins in cosmetics products. Cosmet. Sci. 2002, 53, 185–191. [Google Scholar]
- Shimpi, S.; Chauhan, B.; Shimpi, P. Cyclodextrins: Application in different routes of drug administration. Acta Pharm. 2005, 55, 139–156. [Google Scholar] [PubMed]
- Szejtli, J.; Osa, T. Comprehensive Cyclodextrin Chemistry; Pergmon: Oxford, UK, 1996; Volume 3. [Google Scholar]
- Biwer, A.; Antranikian, G.; Heinzle, E. Enzymatic production of cyclodextrins. Appl. Microbiol. Biotechnol. 2002, 59, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Harada, A.; Hashidzume, A.; Takashima, Y. Cyclodextrin-based supramolecular polymers. Adv. Polym. Sci. 2006, 201, 1–43. [Google Scholar]
- Li, J.; Loh, X.J. Cyclodextrin-based supramolecular architectures: Syntheses, structures and applications for drug and gene delivery. Adv. Drug Deliv. Rev. 2008, 60, 1000–1017. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.R.; Forgo, P.; Stine, K.J.; D’Souza, V.T. Methods for selective modifications of cyclodextrins. Chem. Rev. 1998, 98, 1977–1996. [Google Scholar] [CrossRef] [PubMed]
- Coleman, A.W.; Kasselouri, A. Supramolecular assemblies based on amphiphilic cyclodextrins. Supramol. Chem. 1993, 1, 155–161. [Google Scholar] [CrossRef]
- Ravoo, B.J.; Darcy, R. Cyclodextrin bilayer vesicles. Angew. Chem. Int. Ed. 2000, 39, 4324–4326. [Google Scholar] [CrossRef]
- Messner, M.; Kurkov, S.V.; Jansook, P.; Loftsson, T. Self-assembled cyclodextrin aggregates and nanoparticles. Int. J. Pharm. 2010, 387, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Figueiras, A.; Sarraguca, J.M.G.; Carvalho, R.A.; Pais, A.A.C.C.; Veiga, F.J.B. Interaction of Omeprazole with a Methylated Derivative of β-Cyclodextrin: Phase Solubility, NMR Spectroscopy and Molecular Simulation. Pharm. Res. 2007, 24, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rio, L.; Godoy, A. Use of spectra resolution methodology to investigate surfactant/β-cyclodextrin mixed systems. J. Phys. Chem. B 2007, 111, 6400–6409. [Google Scholar] [CrossRef] [PubMed]
- Denter, U.; Schollmeyer, E. Surface modification of synthetic and natural fibres by fixation of cyclodextrin derivatives. J. Inclusion Phenom. Mol. Recognit. Chem. 1996, 25, 197–202. [Google Scholar] [CrossRef]
- Eftink, M.R.; Andy, M.L.; Bystrom, K.; Perlmytter, H.D.; Kristol, S.J. Cyclodextrin Inclusion Complexes: Studies of the Variation in the Size of Alicyclic Guests. J. Am. Chem. Soc. 1989, 111, 6765–6772. [Google Scholar] [CrossRef]
- Falvey, P.; Lim, C.W.; Darcy, R.; Revermann, T.; Karst, U.; Giesbers, M.; Marcelis, A.T.M.; Lazar, A.; Coleman, A.W.; Reinhoudt, D.N.; et al. Bilayer vesicles of amphiphilic cyclodextrins: Host membranes that recognize guest molecules. Chem. Eur. J. 2005, 11, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- Ogoshi, T.; Harada, A. Chemical Sensors Based on Cyclodextrin Derivatives. Sensors 2008, 8, 4961–4982. [Google Scholar] [CrossRef] [PubMed]
- Park, I.K.; von Recum, H.A.; Jiang, S.; Pun, S.H. Supramolecular assembly of cyclodextrin-based nanoparticles on solid surfaces for gene delivery. Langmuir 2006, 22, 8478–8484. [Google Scholar] [CrossRef] [PubMed]
- Stricker, L.; Fritz, E.C.; Peterlechner, M.; Doltsinis, N.L.; Ravoo, B.J. Arylazopyrazoles as light-responsive molecular switches in cyclodextrin-based supramolecular systems. J. Am. Chem. Soc. 2016, 138, 4547–4554. [Google Scholar] [CrossRef] [PubMed]
- Holzinger, M.; Bouffier, L.; Villalonga, R.; Cosnier, S. Adamantane/beta-cyclodextrin affinity biosensors based on single-walled carbon nanotubes. Biosens. Bioelectron. 2009, 24, 1128–1134. [Google Scholar] [CrossRef] [PubMed]
- Binkowski, C.; Lequart, V.; Hapiot, F.; Tilloy, S.; Cecchelli, R.; Monflier, E.; Martin, P. Adamantoylated monosaccharides: New compounds for modification of the properties of cyclodextrin-containing materials. Carbohydr. Res. 2005, 340, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Voskuhl, J.; Stuart, M.C.A.; Ravoo, B.J. Sugar-decorated sugar vesicles: Lectin-carbohydrate recognition at the surface of cyclodextrin vesicles. Chem. Eur. J. 2010, 16, 2790–2796. [Google Scholar] [CrossRef] [PubMed]
- Vico, R.V.; Voskuhl, J.; Ravoo, B.J. Multivalent Interaction of Cyclodextrin Vesicles, Carbohydrate Guests, and Lectins: A Kinetic Investigation. Langmuir 2011, 27, 1391–1397. [Google Scholar] [CrossRef] [PubMed]
- Kauscher, U.; Stuart, M.C.A.; Drucker, P.; Galla, H.J.; Ravoo, B.J. Incorporation of amphiphilic cyclodextrins into liposomes as artificial receptor units. Langmuir 2013, 29, 7377–7383. [Google Scholar] [CrossRef] [PubMed]
- Bohm, I.; Isenbugel, K.; Ritter, H.; Branscheid, R.; Kolb, U. Cyclodextrin and Adamantane Host–Guest Interactions of Modified Hyperbranched Poly(ethylene imine) as Mimetics for Biological Membranes. Angew. Chem. Int. Ed. 2011, 50, 7896–7899. [Google Scholar] [CrossRef] [PubMed]
- Grayson, S.M.; Fréchet, J.M.J. Convergent Dendrons and Dendrimers: From Synthesis to Applications. Chem. Rev. 2001, 101, 3819–3868. [Google Scholar] [CrossRef] [PubMed]
- Walter, M.V.; Malkoch, M. Simplifying the synthesis of dendrimers: Accelerated approaches. Chem. Soc. Rev. 2012, 41, 4593–4609. [Google Scholar] [CrossRef] [PubMed]
- Esfanda, R.; Tomalia, D.A. Poly(amidoamine) (PAMAM) dendrimers: From biomimicry to drug delivery and biomedical applications. Drug Discov. Today 2001, 6, 427–436. [Google Scholar] [CrossRef]
- Sharma, A.; Gautam, S.; Gupta, A.K. Surface modified dendrimers: Synthesis and characterization for cancer targeted drug delivery. Bioorg. Med. Chem. 2011, 19, 3341–3346. [Google Scholar] [CrossRef] [PubMed]
- Kircheis, R.; Wightman, L.; Wagner, E. Design and gene delivery activity of modified Polyethylenimines. Adv. Drug Deliv. Rev. 2001, 53, 341–358. [Google Scholar] [CrossRef]
- Duncan, R.; Izzo, L. Dendrimer biocompatibility and toxicity. Adv. Drug Deliv. Rev. 2005, 57, 2215–2237. [Google Scholar] [CrossRef] [PubMed]
- Lamanna, G.; Russier, J.; Menard-Moyon, C.; Bianco, A. Hydramers: Design, synthesis and characterization of different generation novel Hydra-like dendrons based on multifunctionalized adamantine. Chem. Commun. 2011, 47, 8955–8957. [Google Scholar] [CrossRef] [PubMed]
- Grillaud, M.; Russier, J.; Bianco, A. Polycationic Adamantane-Based Dendrons of Different Generations Display High Cellular Uptake without Triggering Cytotoxicity. J. Am. Chem. Soc. 2014, 136, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Grillaud, M.; Perez Ruiz de Garibay, A.; Bianco, A. Polycationic adamantane-based dendrons form nanorods in complex with plasmid DNA. RSC Adv. 2016, 6, 42933–42942. [Google Scholar] [CrossRef]
- Paolino, M.; Ennen, F.; Lamponi, S.; Cernescu, M.; Voit, B.; Cappelli, A.; Appelhans, D.; Komber, H. Cyclodextrin-Adamantane Host–Guest Interactions on the Surface of Biocompatible Adamantyl-Modified Glycodendrimers. Macromolecules 2013, 46, 3215–3227. [Google Scholar] [CrossRef]
- Grillaud, M.; Bianco, A. Multifunctional adamantane derivatives as new scaffolds for the multipresentation of bioactive peptides. J. Pept. Sci. 2015, 21, 330–345. [Google Scholar] [CrossRef] [PubMed]
- Lamanna, G.; Grillaud, M.; Macri, C.; Chaloin, O. Adamantane-based dendrons for trimerization of the therapeutic P140 peptide. Biomaterials 2014, 35, 1–9. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štimac, A.; Šekutor, M.; Mlinarić-Majerski, K.; Frkanec, L.; Frkanec, R. Adamantane in Drug Delivery Systems and Surface Recognition. Molecules 2017, 22, 297. https://doi.org/10.3390/molecules22020297
Štimac A, Šekutor M, Mlinarić-Majerski K, Frkanec L, Frkanec R. Adamantane in Drug Delivery Systems and Surface Recognition. Molecules. 2017; 22(2):297. https://doi.org/10.3390/molecules22020297
Chicago/Turabian StyleŠtimac, Adela, Marina Šekutor, Kata Mlinarić-Majerski, Leo Frkanec, and Ruža Frkanec. 2017. "Adamantane in Drug Delivery Systems and Surface Recognition" Molecules 22, no. 2: 297. https://doi.org/10.3390/molecules22020297
APA StyleŠtimac, A., Šekutor, M., Mlinarić-Majerski, K., Frkanec, L., & Frkanec, R. (2017). Adamantane in Drug Delivery Systems and Surface Recognition. Molecules, 22(2), 297. https://doi.org/10.3390/molecules22020297