The HMGB1/RAGE Pro-Inflammatory Axis in the Human Placenta: Modulating Effect of Low Molecular Weight Heparin


 ,
, 
Abstract
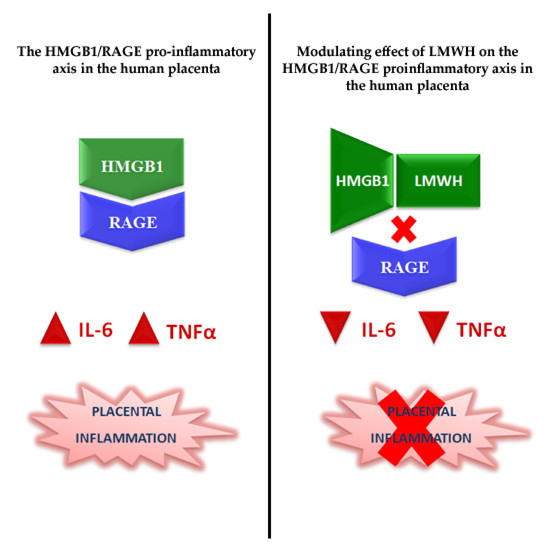
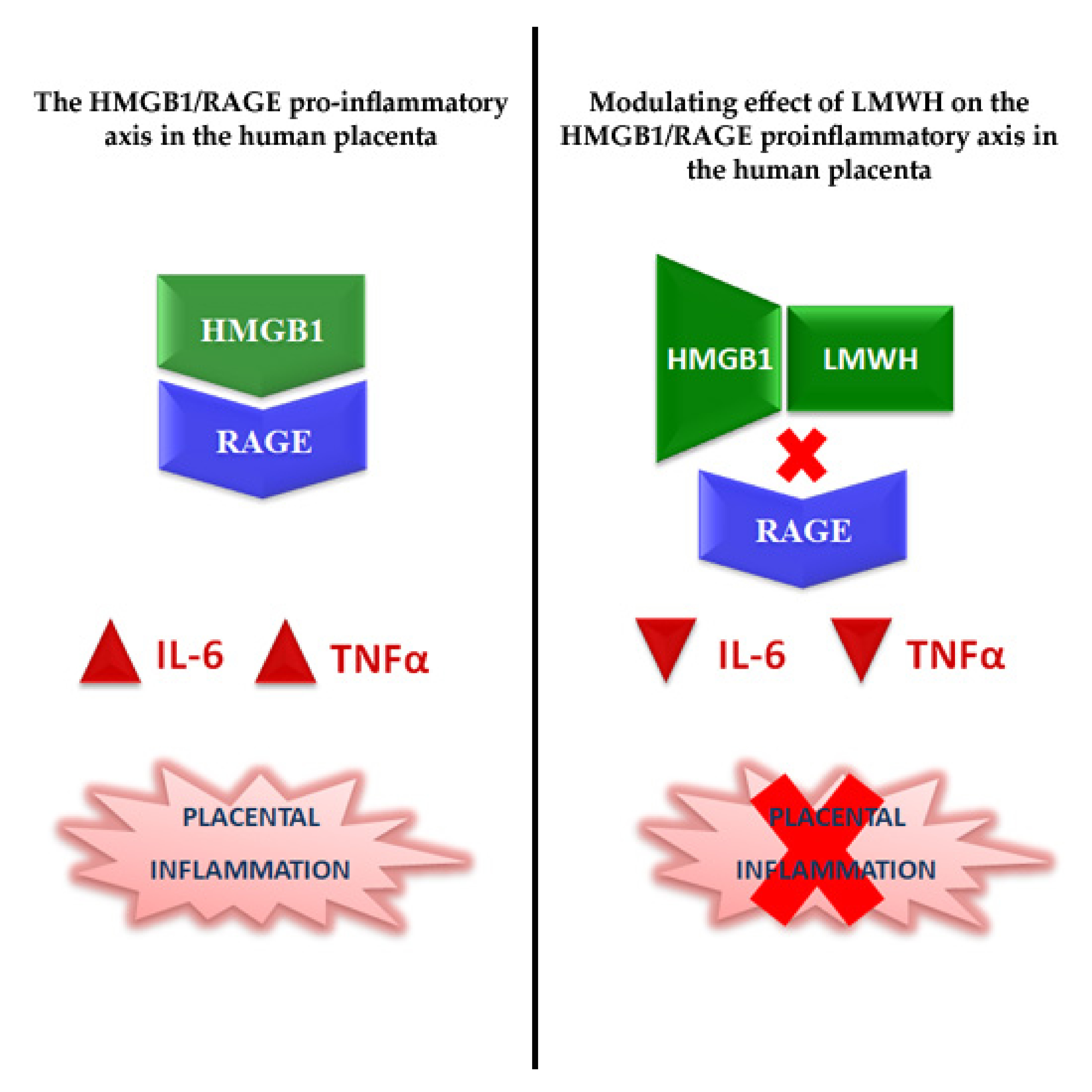
1. Introduction
2. Results
2.1. Study Population
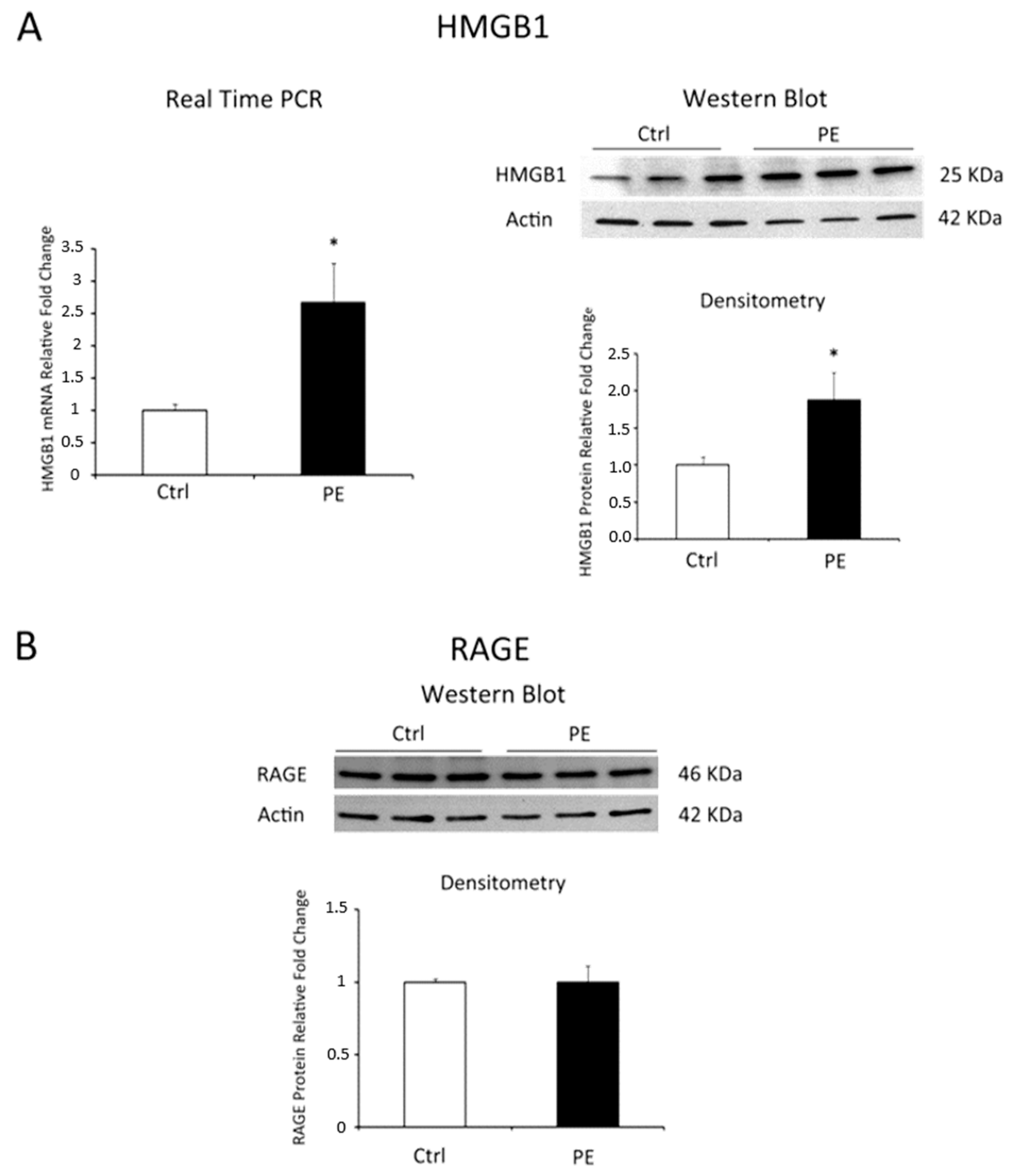
2.2. HMGB1 and RAGE Expressions in Physiological and Pre-Eclamptic Placentae
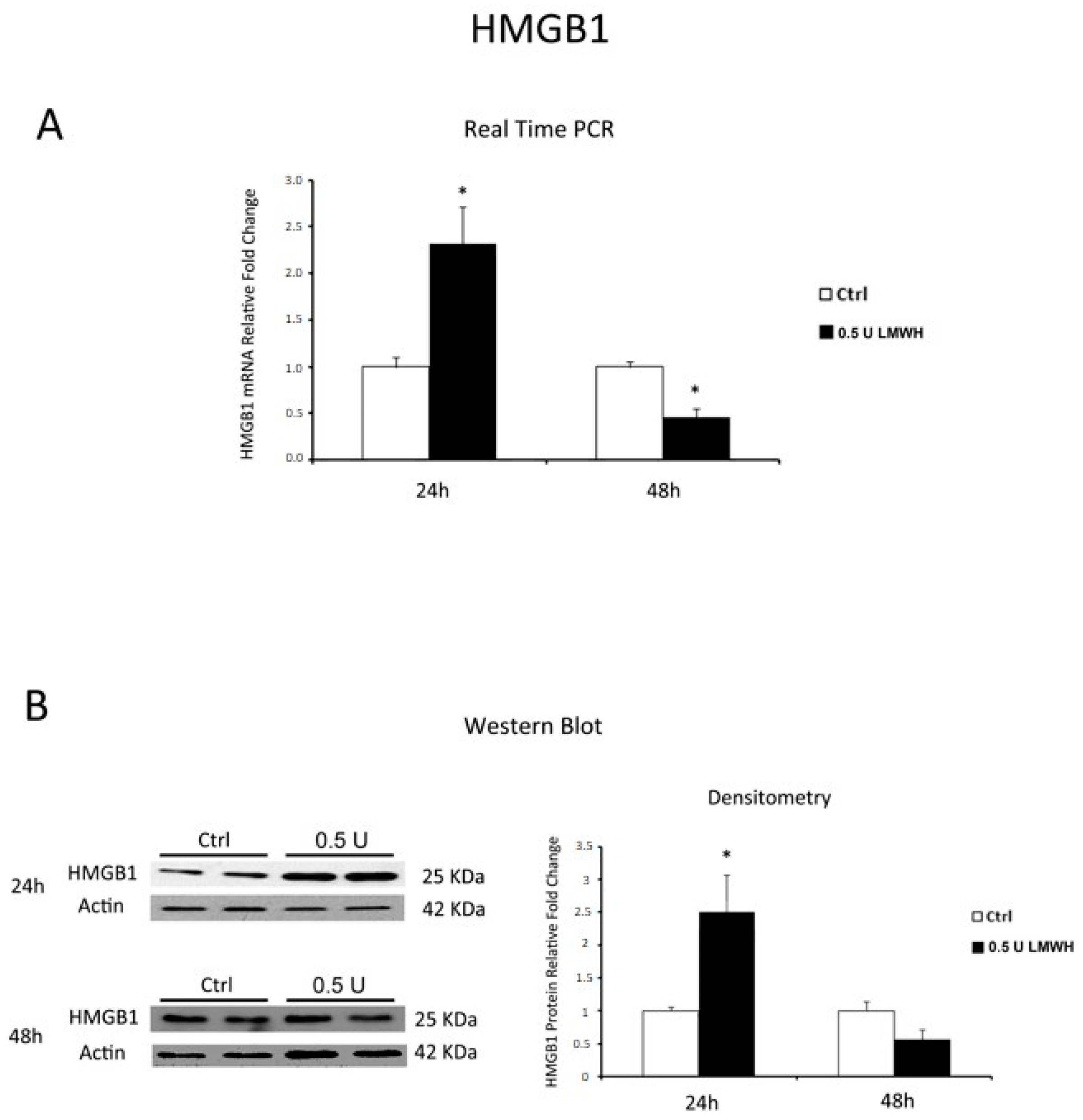
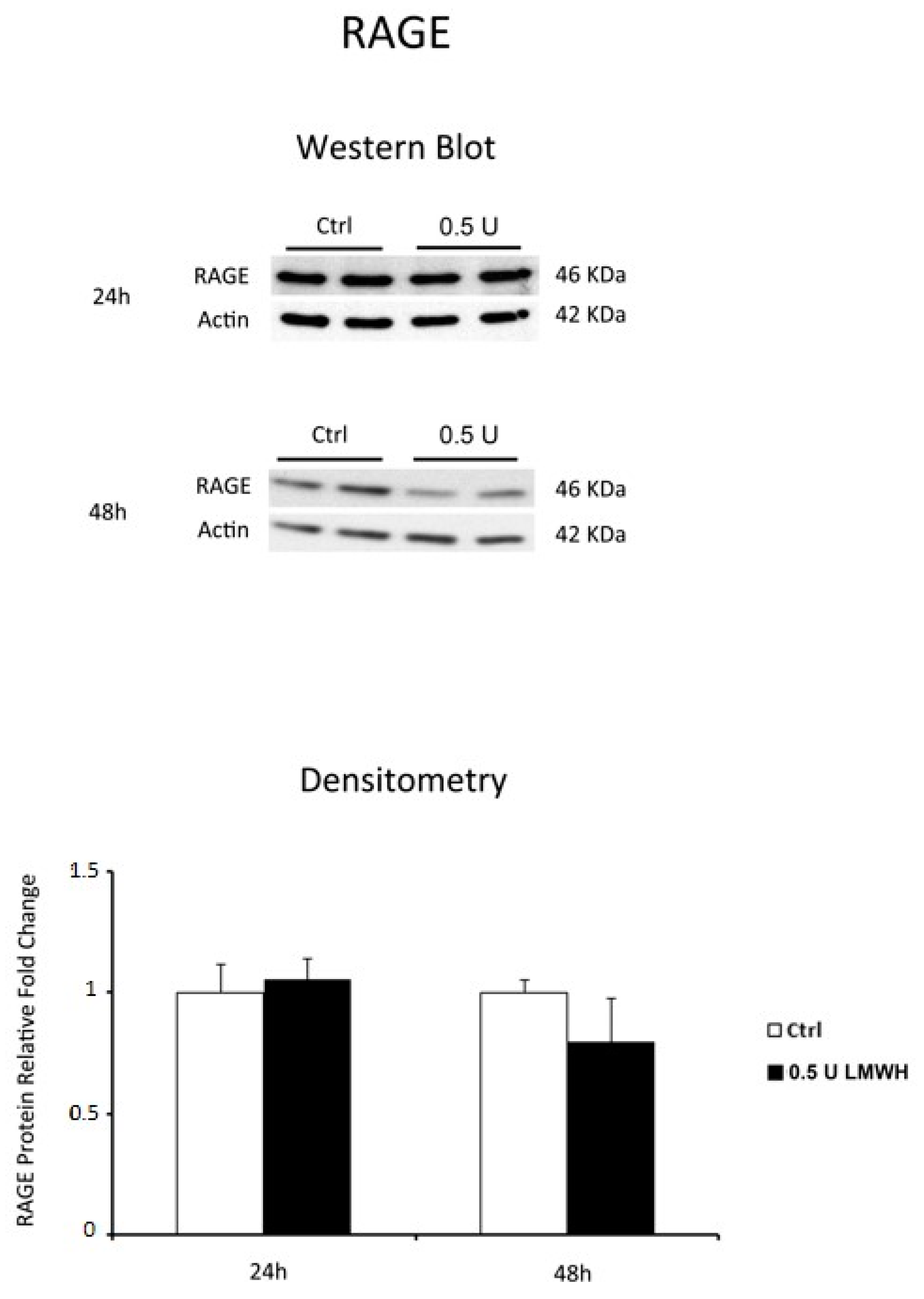
2.3. LMWH Effect on HMGB1 and RAGE Expression in Physiological Human Villous Explants
2.4. 0.5 U LMWH Treatment Did Not Affect Physiological Term Placental Villous Explants Viability
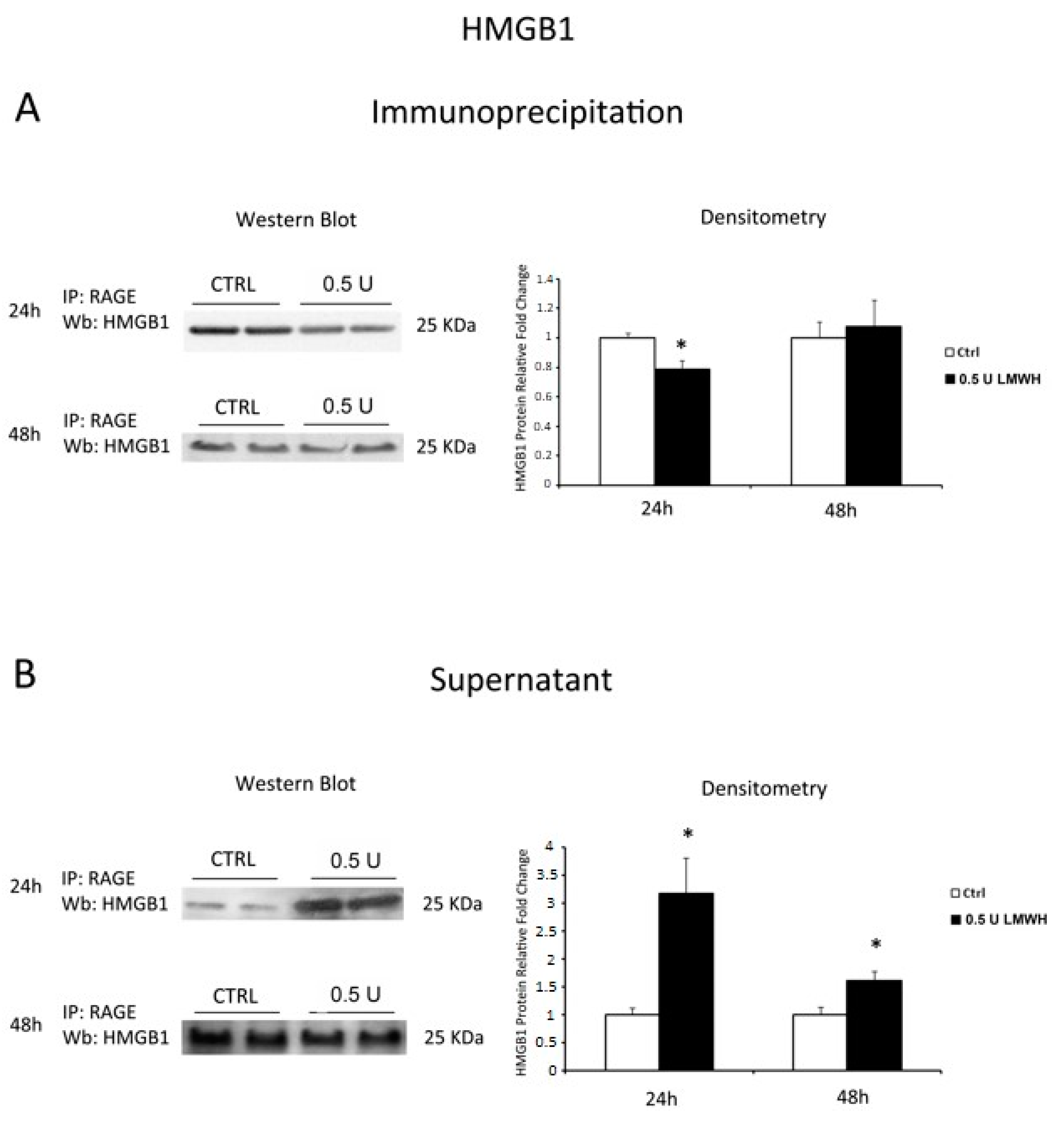
2.5. LMWH Inhibits HMGB1/RAGE Interaction
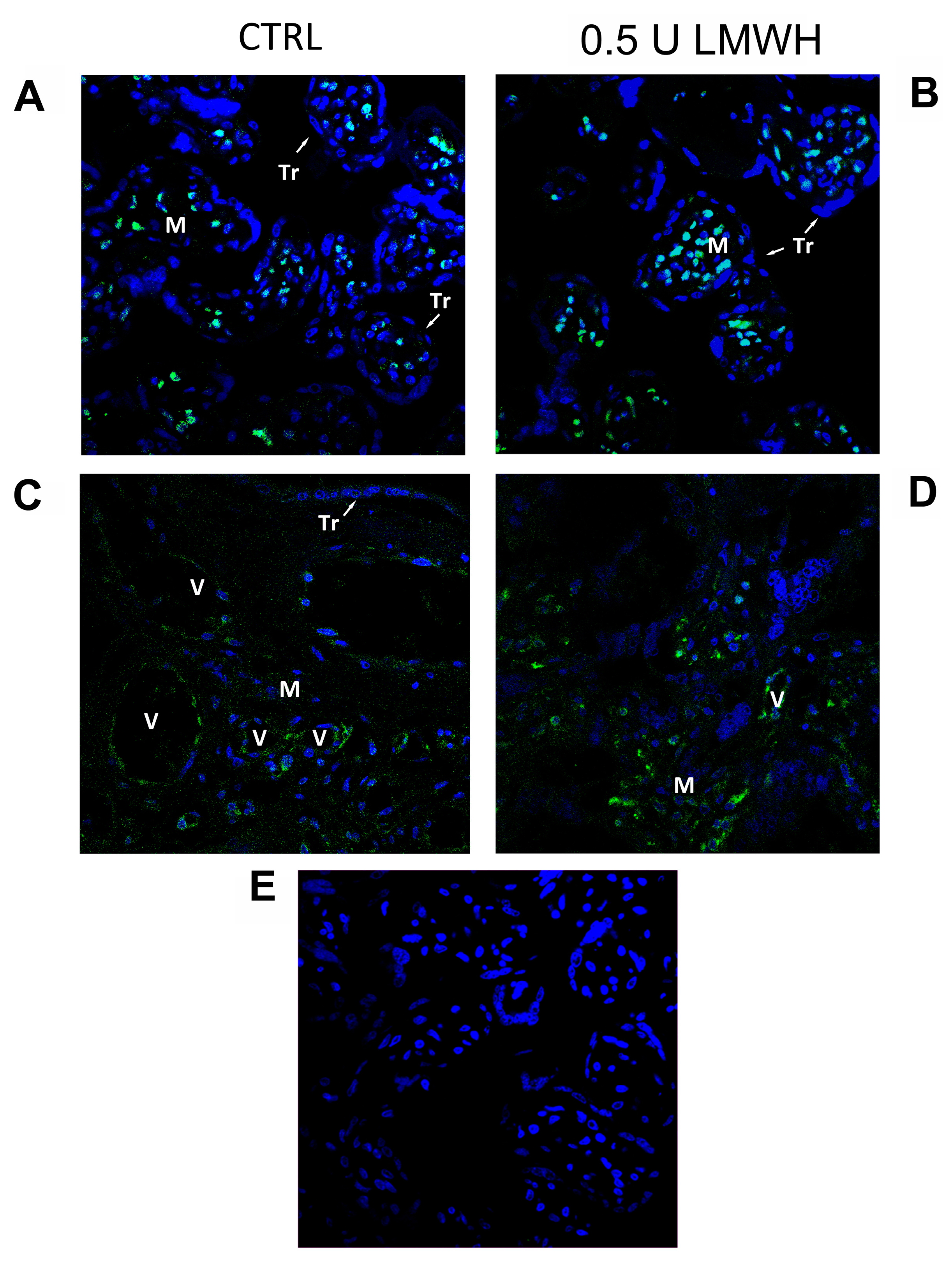
2.6. HMGB1 Immunofluorescent Staining in Physiological Placental Villous Explants
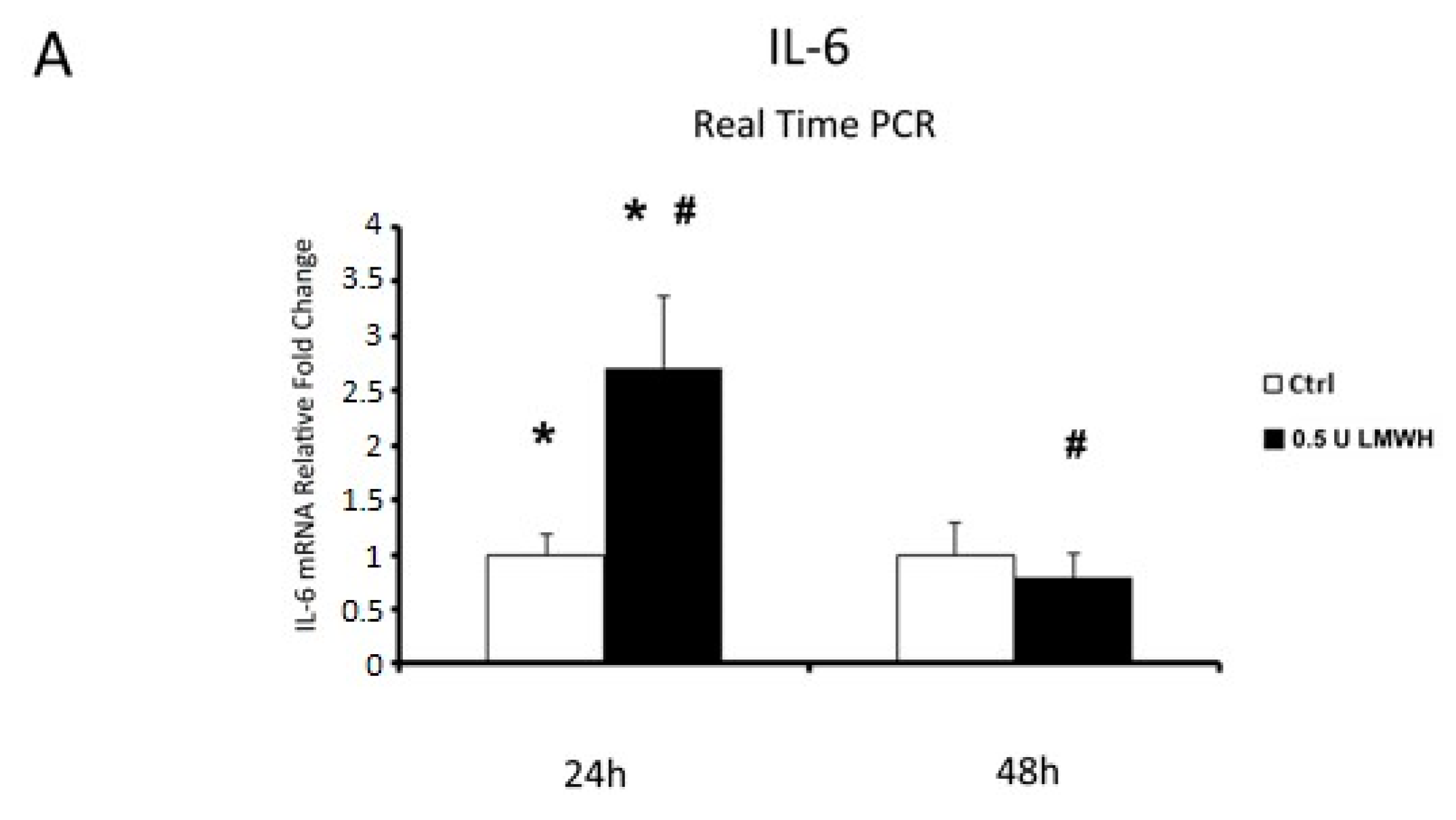
2.7. IL-6 and TNFα Gene Expression in Physiological Placental Villous Explants
3. Discussion
4. Materials and Methods
4.1. Ethics Statement and Place of Recruitment
4.2. Study Population and Tissues Collection
4.3. Human Chorionic Villous Explants Cultures and LMWH Treatment
4.4. Lactate Dehydrogenase (LDH) Cytotoxicity Assay
4.5. RNA Isolation and Real Time PCR
4.6. Western Blot Analysis
4.7. HMGB1 Immunofluorescence (IF) Staining
4.8. Immunoprecipitation (IP) Assay
4.9. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- ACOG Committee on Obstetric Practice. ACOG Practice Bulletin. Diagnosis and management of preeclampsia and eclampsia. Number 33, January 2002. American College of Obstetricians and Gynecologists. Int. J. Gynaecol. Obstet. Off. Organ Int. Fed. Gynaecol. Obstet. 2002, 77, 67–75. [Google Scholar]
- Redman, C.W.; Sargent, I.L. Latest advances in understanding preeclampsia. Science 2005, 308, 1592–1594. [Google Scholar] [CrossRef] [PubMed]
- Myatt, L.; Webster, R.P. Is vascular biology in preeclampsia better? J. Thromb. Haemost. 2009, 7, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Aranguren, L.C.; Prada, C.E.; Riano-Medina, C.E.; Lopez, M. Endothelial dysfunction and preeclampsia: Role of oxidative stress. Front. Physiol. 2014, 5, 372. [Google Scholar] [CrossRef] [PubMed]
- Smarason, A.K.; Sargent, I.L.; Starkey, P.M.; Redman, C.W. The effect of placental syncytiotrophoblast microvillous membranes from normal and pre-eclamptic women on the growth of endothelial cells in vitro. Br. J. Obstet. Gynaecol. 1993, 100, 943–949. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E. Damps, pamps and alarmins: All we need to know about danger. J. Leukoc. Biol. 2007, 81, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.N.; Hay, D.P. A clear and present danger: Inflammasomes damping down disorders of pregnancy. Hum. Reprod. Update 2014. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, G.H.; Sanders, C.; Johns, E.W. A new group of chromatin-associated proteins with a high content of acidic and basic amino acids. Eur. J. Biochem.—FEBS 1973, 38, 14–19. [Google Scholar] [CrossRef]
- Scaffidi, P.; Misteli, T.; Bianchi, M.E. Release of chromatin protein hmgb1 by necrotic cells triggers inflammation. Nature 2002, 418, 191–195. [Google Scholar] [CrossRef] [PubMed]
- O'Connor, K.A.; Hansen, M.K.; Rachal Pugh, C.; Deak, M.M.; Biedenkapp, J.C.; Milligan, E.D.; Johnson, J.D.; Wang, H.; Maier, S.F.; Tracey, K.J.; et al. Further characterization of high mobility group box 1 (hmgb1) as a proinflammatory cytokine: Central nervous system effects. Cytokine 2003, 24, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Campana, L.; Bosurgi, L.; Rovere-Querini, P. Hmgb1: A two-headed signal regulating tumor progression and immunity. Curr. Opin. Immunol. 2008, 20, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E.; Manfredi, A.A. High-mobility group box 1 (hmgb1) protein at the crossroads between innate and adaptive immunity. Immunol. Rev. 2007, 220, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Hori, O.; Brett, J.; Slattery, T.; Cao, R.; Zhang, J.; Chen, J.X.; Nagashima, M.; Lundh, E.R.; Vijay, S.; Nitecki, D.; et al. The receptor for advanced glycation end products (rage) is a cellular binding site for amphoterin. Mediation of neurite outgrowth and co-expression of rage and amphoterin in the developing nervous system. J. Biol. Chem. 1995, 270, 25752–25761. [Google Scholar] [CrossRef] [PubMed]
- Andersson, U.; Wang, H.; Palmblad, K.; Aveberger, A.C.; Bloom, O.; Erlandsson-Harris, H.; Janson, A.; Kokkola, R.; Zhang, M.; Yang, H.; et al. High mobility group 1 protein (hmg-1) stimulates proinflammatory cytokine synthesis in human monocytes. J. Exp. Med. 2000, 192, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bloom, O.; Zhang, M.; Vishnubhakat, J.M.; Ombrellino, M.; Che, J.; Frazier, A.; Yang, H.; Ivanova, S.; Borovikova, L.; et al. Hmg-1 as a late mediator of endotoxin lethality in mice. Science 1999, 285, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Mori, S.; Takahashi, H.K.; Tomono, Y.; Wake, H.; Kanke, T.; Sato, Y.; Hiraga, N.; Adachi, N.; Yoshino, T.; et al. Anti-high mobility group box 1 monoclonal antibody ameliorates brain infarction induced by transient ischemia in rats. FASEB J. 2007, 21, 3904–3916. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Torikai, M.; Kuwazuru, A.; Tanaka, M.; Horai, N.; Fukuda, T.; Yamada, S.; Nagayama, S.; Hashiguchi, K.; Sunahara, N.; et al. Extracellular high mobility group box chromosomal protein 1 is a coupling factor for hypoxia and inflammation in arthritis. Arthritis Rheumatol. 2008, 58, 2675–2685. [Google Scholar] [CrossRef] [PubMed]
- Sawa, H.; Ueda, T.; Takeyama, Y.; Yasuda, T.; Shinzeki, M.; Nakajima, T.; Kuroda, Y. Blockade of high mobility group box-1 protein attenuates experimental severe acute pancreatitis. World J. Gastroenterol. 2006, 12, 7666–7670. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.; Arcaroli, J.; Carmody, A.; Wang, H.; Tracey, K.J. Hmg-1 as a mediator of acute lung inflammation. J. Immunol. 2000, 165, 2950–2954. [Google Scholar] [CrossRef] [PubMed]
- Holmlund, U.; Wahamaa, H.; Bachmayer, N.; Bremme, K.; Sverremark-Ekstrom, E.; Palmblad, K. The novel inflammatory cytokine high mobility group box protein 1 (hmgb1) is expressed by human term placenta. Immunology 2007, 122, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Koga, K.; Osuga, Y.; Hirata, T.; Saito, A.; Yoshino, O.; Hirota, Y.; Harada, M.; Takemura, Y.; Fujii, T.; et al. High mobility group box 1 (hmgb1) levels in the placenta and in serum in preeclampsia. Am. J. Reprod. Immunol. 2011, 66, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Cooke, C.L.; Brockelsby, J.C.; Baker, P.N.; Davidge, S.T. The receptor for advanced glycation end products (rage) is elevated in women with preeclampsia. Pregnancy Hypertens. 2003, 22, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, I.; Mutlu, M.F.; Biri, A.; Bulut, B.; Erdem, M.; Erdem, A. Effects of anticoagulant therapy on pregnancy outcomes in patients with thrombophilia and previous poor obstetric history. Blood Coagul. Fibrinolysis 2014. [Google Scholar] [CrossRef] [PubMed]
- Buyse, F.G.; Wormgoor, B.H.; Bernard, J.T.; Koudstaal, J. Anticoagulant therapy of patients with repeated placental infarction. Obstet. Gynecol. 1974, 43, 844–848. [Google Scholar] [PubMed]
- Kingdom, J.C.; Drewlo, S. Is heparin a placental anticoagulant in high-risk pregnancies? Blood 2011, 118, 4780–4788. [Google Scholar] [CrossRef] [PubMed]
- D'Souza, R.; Keating, S.; Walker, M.; Drewlo, S.; Kingdom, J. Unfractionated heparin and placental pathology in high-risk pregnancies: Secondary analysis of a pilot randomized controlled trial. Placenta 2014, 35, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Rey, E.; Garneau, P.; David, M.; Gauthier, R.; Leduc, L.; Michon, N.; Morin, F.; Demers, C.; Kahn, S.R.; Magee, L.A.; et al. Dalteparin for the prevention of recurrence of placental-mediated complications of pregnancy in women without thrombophilia: A pilot randomized controlled trial. J. Thromb. Haemost. 2009, 7, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Rodger, M.A.; Carrier, M.; Le Gal, G.; Martinelli, I.; Perna, A.; Rey, E.; de Vries, J.I.P.; Gris, J.-C.; Low-Molecular-Weight Heparin for Placenta-Mediated Pregnancy Complications Study Group. Meta-analysis of low-molecular-weight heparin to prevent recurrent placenta-mediated pregnancy complications. Blood 2014, 123, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, V., Jr.; Pacheco, R.G.; Esposito, C.C.; Buongusto, F.; Lima Castelo-Branco, M.T.; Madi, K.; Belmiro, C.R.; Goncalves Pavao, M.S.; Pereira de Souza, H.S.; Schanaider, A. Ascidian (chordate-tunicate) and mammalian heparin enemas attenuate experimental diversion colitis. Surgery 2014, 155, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Babazada, H.; Yamashita, F.; Hashida, M. Suppression of experimental arthritis with self-assembling glycol-split heparin nanoparticles via inhibition of TLR4-NF-κB signaling. J. Control. Release 2014, 194, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Xie, H.; Tian, R.; Yu, K.; Wang, R. Efficacy of low molecular weight heparin in patients with acute exacerbation of chronic obstructive pulmonary disease receiving ventilatory support. COPD J. Chronic Obstruct. Pulm. Dis. 2014, 11, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Poli, M.; Asperti, M.; Ruzzenenti, P.; Mandelli, L.; Campostrini, N.; Martini, G.; Di Somma, M.; Maccarinelli, F.; Girelli, D.; Naggi, A.; et al. Oversulfated heparins with low anticoagulant activity are strong and fast inhibitors of hepcidin expression in vitro and in vivo. Biochem. Pharmacol. 2014, 92, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Mori, S.; Wake, H.; Zhang, J.; Liu, K.; Izushi, Y.; Takahashi, H.K.; Peng, B.; Nishibori, M. Establishment of in vitro binding assay of high mobility group box-1 and s100a12 to receptor for advanced glycation endproducts: Heparin's effect on binding. Acta Med. Okayama 2009, 63, 203–211. [Google Scholar] [PubMed]
- Ling, Y.; Yang, Z.Y.; Yin, T.; Li, L.; Yuan, W.W.; Wu, H.S.; Wang, C.Y. Heparin changes the conformation of high-mobility group protein 1 and decreases its affinity toward receptor for advanced glycation endproducts in vitro. Int. Immunopharmacol. 2011, 11, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Merenmies, J.; Pihlaskari, R.; Laitinen, J.; Wartiovaara, J.; Rauvala, H. 30-kda heparin-binding protein of brain (amphoterin) involved in neurite outgrowth. Amino acid sequence and localization in the filopodia of the advancing plasma membrane. J. Biol. Chem. 1991, 266, 16722–16729. [Google Scholar] [PubMed]
- Parazzini, F.; Cortinovis, I.; Bortolus, R.; Fedele, L. Standards of birth weight in Italy. Ann. Ostet. Ginecol. Med. Perinat. 1991, 112, 203–246. [Google Scholar] [PubMed]
- Bertino, E.; Di Battista, E.; Bossi, A.; Pagliano, M.; Fabris, C.; Aicardi, G.; Milani, S. Fetal growth velocity: Kinetic, clinical, and biological aspects. Arch. Dis. Child. Fetal Neonatal Ed. 1996, 74, F10–F15. [Google Scholar] [CrossRef] [PubMed]
- Lappas, M.; Rice, G.E. The role and regulation of the nuclear factor kappa b signalling pathway in human labour. Placenta 2007, 28, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Cornelli, U.; Fareed, J. Human pharmacokinetics of low molecular weight heparins. Semin. Thromb. Hemost. 1999, 25 (Suppl. 3), 57–61. [Google Scholar] [PubMed]
- Sobel, M.L.; Kingdom, J.; Drewlo, S. Angiogenic response of placental villi to heparin. Obstet. Gynecol. 2011, 117, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Drewlo, S.; Levytska, K.; Sobel, M.; Baczyk, D.; Lye, S.J.; Kingdom, J.C. Heparin promotes soluble vegf receptor expression in human placental villi to impair endothelial vegf signaling. J. Thromb. Haemost. 2011, 9, 2486–2497. [Google Scholar] [CrossRef] [PubMed]
- Germanova, A.; Koucky, M.; Hajek, Z.; Parizek, A.; Zima, T.; Kalousova, M. Soluble receptor for advanced glycation end products in physiological and pathological pregnancy. Clin. Biochem. 2010, 43, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Pradervand, P.A.; Clerc, S.; Frantz, J.; Rotaru, C.; Bardy, D.; Waeber, B.; Liaudet, L.; Vial, Y.; Feihl, F. High mobility group box 1 protein (hmgb-1): A pathogenic role in preeclampsia? Placenta 2014, 35, 784–786. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yin, Y.X.; Wei, J.; Tong, M.; Shen, F.; Zhao, M.; Chamley, L. Increased expression of high mobility group box 1 (hmgb1) in the cytoplasm of placental syncytiotrophoblast from preeclamptic placentae. Cytokine 2016, 85, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Giacobbe, A.; Grasso, R.; Imbesi, G.; Salpietro, C.D.; Grasso, L.; Lagana, A.S.; Triolo, O.; Di Benedetto, A. High mobility group protein b1: A new biomarker of obesity in pregnant women? Gynecol. Endocrinol. 2015, 31, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Miranda, J.; Chaiworapongsa, T.; Korzeniewski, S.J.; Chaemsaithong, P.; Gotsch, F.; Dong, Z.; Ahmed, A.I.; Yoon, B.H.; Hassan, S.S.; et al. Prevalence and clinical significance of sterile intra-amniotic inflammation in patients with preterm labor and intact membranes. Am. J. Reprod. Immunol. 2014, 72, 458–474. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Cai, J.; Zhu, Z.; Chen, D.; Wang, J.; Wang, Q.; Teng, Y.; Huang, Y.; Tao, M.; Xia, A.; et al. Hypoxic trophoblast hmgb1 induces endothelial cell hyperpermeability via the trl-4/caveolin-1 pathway. J. Immunol. 2014, 193, 5000–5012. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sargent, I.L. Immunology of pre-eclampsia. Am. J. Reprod. Immunol. 2010, 63, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Tannetta, D.S.; Dragovic, R.A.; Gardiner, C.; Southcombe, J.H.; Collett, G.P.; Sargent, I.L. Review: Does size matter? Placental debris and the pathophysiology of pre-eclampsia. Placenta 2012, 33 (Suppl.), S48–S54. [Google Scholar] [CrossRef] [PubMed]
- Chekir, C.; Nakatsuka, M.; Noguchi, S.; Konishi, H.; Kamada, Y.; Sasaki, A.; Hao, L.; Hiramatsu, Y. Accumulation of advanced glycation end products in women with preeclampsia: Possible involvement of placental oxidative and nitrative stress. Placenta 2006, 27, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Bose, P.; Black, S.; Kadyrov, M.; Weissenborn, U.; Neuten, J.; Regan, L.; Huppertz, B. Heparin and aspirin attenuate placental apoptosis in vitro: Implications for early pregnancy failure. A. J. Obstet. Gynecol. 2005, 192, 23–30. [Google Scholar] [CrossRef] [PubMed]
- D'Ippolito, S.; Di Nicuolo, F.; Marana, R.; Castellani, R.; Stinson, J.; Tersigni, C.; Scambia, G.; Di Simone, N. Emerging nonanticoagulant role of low molecular weight heparins on extravillous trophoblast functions and on heparin binding-epidermal growth factor and cystein-rich angiogenic inducer 61 expression. Fertil. Steril. 2012, 98, 1028–1036. [Google Scholar] [CrossRef] [PubMed]
- Bierhaus, A.; Humpert, P.M.; Morcos, M.; Wendt, T.; Chavakis, T.; Arnold, B.; Stern, D.M.; Nawroth, P.P. Understanding rage, the receptor for advanced glycation end products. J. Mol. Med. (Berl.) 2005, 83, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.; Covacu, R.; Sunnemark, D.; Danilov, A.I.; Dal Bianco, A.; Khademi, M.; Wallstrom, E.; Lobell, A.; Brundin, L.; Lassmann, H.; et al. Pivotal advance: Hmgb1 expression in active lesions of human and experimental multiple sclerosis. J. Leukoc. Biol. 2008, 84, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Huebener, P.; Pradere, J.P.; Hernandez, C.; Gwak, G.Y.; Caviglia, J.M.; Mu, X.; Loike, J.D.; Jenkins, R.E.; Antoine, D.J.; Schwabe, R.F. The hmgb1/rage axis triggers neutrophil-mediated injury amplification following necrosis. J. Clin. Investig. 2015, 125, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Bredeson, S.; Papaconstantinou, J.; Deford, J.H.; Kechichian, T.; Syed, T.A.; Saade, G.R.; Menon, R. Hmgb1 promotes a p38mapk associated non-infectious inflammatory response pathway in human fetal membranes. PLoS ONE 2014, 9, e113799. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Gamboni-Robertson, F.; He, Q.; Svetkauskaite, D.; Kim, J.Y.; Strassheim, D.; Sohn, J.W.; Yamada, S.; Maruyama, I.; Banerjee, A.; et al. High mobility group box 1 protein interacts with multiple toll-like receptors. Am. J. Physiol. Cell Physiol. 2006, 290, C917–C924. [Google Scholar] [CrossRef] [PubMed]
- Musumeci, D.; Roviello, G.N.; Montesarchio, D. An overview on hmgb1 inhibitors as potential therapeutic agents in hmgb1-related pathologies. Pharmacol. Ther. 2014, 141, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Wu, J.; Patel, V.; Sitapara, R.; Rao, N.V.; Kennedy, T.P.; Mantell, L.L. Partially-desulfated heparin improves survival in pseudomonas pneumonia by enhancing bacterial clearance and ameliorating lung injury. J. Immunotoxicol. 2014, 11, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Mitola, S.; Belleri, M.; Urbinati, C.; Coltrini, D.; Sparatore, B.; Pedrazzi, M.; Melloni, E.; Presta, M. Cutting edge: Extracellular high mobility group box-1 protein is a proangiogenic cytokine. J. Immunol. 2006, 176, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Schlueter, C.; Weber, H.; Meyer, B.; Rogalla, P.; Roser, K.; Hauke, S.; Bullerdiek, J. Angiogenetic signaling through hypoxia: Hmgb1: An angiogenetic switch molecule. Am. J. Pathol. 2005, 166, 1259–1263. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, Z.; Zhang, L.; Shi, Y.; Qi, J.; Chang, A.; Gao, J.; Feng, Y.; Yang, X. Hmgb1-rage signaling pathway in severe preeclampsia. Placenta 2015, 36, 1148–1152. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, N.; Agrotis, A.; Antropova, Y.; DiVitto, G.; Kanellakis, P.; Kostolias, G.; Ilyinskaya, O.; Tararak, E.; Bobik, A. Increased expression of the DNA-binding cytokine hmgb1 in human atherosclerotic lesions: Role of activated macrophages and cytokines. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2320–2325. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Vishnubhakat, J.M.; Bloom, O.; Zhang, M.; Ombrellino, M.; Sama, A.; Tracey, K.J. Proinflammatory cytokines (tumor necrosis factor and interleukin 1) stimulate release of high mobility group protein-1 by pituicytes. Surgery 1999, 126, 389–392. [Google Scholar] [CrossRef]
- Xiong, X.X.; Gu, L.J.; Shen, J.; Kang, X.H.; Zheng, Y.Y.; Yue, S.B.; Zhu, S.M. Probenecid protects against transient focal cerebral ischemic injury by inhibiting hmgb1 release and attenuating aqp4 expression in mice. Neurochem. Res. 2014, 39, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Lu, J.; Dai, W.; Wang, F.; Xu, L.; Chen, K.; He, L.; Cheng, P.; Zhang, Y.; Wang, C.; et al. Ethyl pyruvate ameliorates hepatic ischemia-reperfusion injury by inhibiting intrinsic pathway of apoptosis and autophagy. Mediat. Inflamm. 2013, 2013, 461536. [Google Scholar] [CrossRef] [PubMed]
- Entezari, M.; Javdan, M.; Antoine, D.J.; Morrow, D.M.; Sitapara, R.A.; Patel, V.; Wang, M.; Sharma, L.; Gorasiya, S.; Zur, M.; et al. Inhibition of extracellular hmgb1 attenuates hyperoxia-induced inflammatory acute lung injury. Redox Biol. 2014, 2, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Raymond, D.; Peterson, E. A critical review of early-onset and late-onset preeclampsia. Obstet. Gynecol. Surv. 2011, 66, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological (n = 19) | Preeclampsia (n = 18) | p Value | |
|---|---|---|---|
| Nulliparae (%) | 47.36 | 50 | n.s. |
| Gestational age at delivery (weeks) | 39.4 (38–41) | 32.8 (28–37) | <0.001 |
| Maternal age at delivery (years) | 34 (23–39) | 34 (20–41) | n.s. |
| Ethnicity (%) | |||
| Caucasian | 89.47 | 86.33 | n.s. |
| Prenatal medications (%) | |||
| Albumin | - | 11.11 | n.s. |
| Antibiotics | 42.1 | 16.67 | n.s. |
| Antidepressant | - | - | - |
| Diuretics | - | 16.67 | n.s. |
| Eutirox | 5.26 | - | n.s. |
| Folin | 84.21 | 94.44 | n.s. |
| Heparin | 5.26 | 16.66 | n.s. |
| Iron | 15.79 | 5.55 | n.s. |
| Proton Pump Inhibitors (PPIs) | 5.26 | 11.11 | n.s. |
| Ventolin | - | - | - |
| Smokers (%) | 15.78 | 16.67 | n.s. |
| Alcohol (%) | 10.52 | 16.67 | n.s. |
| Previous prenatal admission (%) | 5.26 | 16.67 | n.s. |
| Systolic Blood pressure (mm Hg) | 120 (90–140) | 162.5 (130–180) | <0.001 |
| Diastolic Blood pressure (mm Hg) | 75 (60–90) | 100 (85–117) | <0.001 |
| Proteinuria (g/24 h) | Absent | 1.2 (0.33–8.28) | <0.001 |
| A/REDF (%) | 0 | 50 | <0.001 |
| Pathological Uterine Doppler (%) | 0 | 66.67 | <0.001 |
| Labor (%) | 63 | 17 | p = 0.032 |
| Antibiotics in labor (%) | 31.58 | 83.33 | p = 0.04 |
| Delivery to processing (range in hours) | 0–3 | 0–3 | |
| Caesarean section (%) | 37 | 83 | p = 0.011 |
| Anesthesia (%) | 63.15 | 83.33 | n.s. |
| Local | 25 | - | p = 0.044 |
| Spinal | 58.33 | 93.33 | |
| Epidural | 16.67 | - | |
| General | - | 6.67 | |
| Maternal oxygen given at delivery? (%) | - | 16.67 | n.s. |
| Birth weight (g) | 3550 (2920–4020) | AGA (n = 5): 2150 (1110–3180) FGR (n = 13): 1250 (610–1880) | <0.001 |
| Placental weight (g) | 580 (450–845) | 310 (184–650) | <0.001 |
| Fetal sex (%) | p = 0.061 | ||
| Male | 57.9 | 22.2 | |
| Female | 42.1 | 77.8 | |
| Magnesium sulfate (%) | - | 27.78 | p = 0.02 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zenerino, C.; Nuzzo, A.M.; Giuffrida, D.; Biolcati, M.; Zicari, A.; Todros, T.; Rolfo, A. The HMGB1/RAGE Pro-Inflammatory Axis in the Human Placenta: Modulating Effect of Low Molecular Weight Heparin. Molecules 2017, 22, 1997. https://doi.org/10.3390/molecules22111997
Zenerino C, Nuzzo AM, Giuffrida D, Biolcati M, Zicari A, Todros T, Rolfo A. The HMGB1/RAGE Pro-Inflammatory Axis in the Human Placenta: Modulating Effect of Low Molecular Weight Heparin. Molecules. 2017; 22(11):1997. https://doi.org/10.3390/molecules22111997
Chicago/Turabian StyleZenerino, Cristian, Anna Maria Nuzzo, Domenica Giuffrida, Marilisa Biolcati, Alessandra Zicari, Tullia Todros, and Alessandro Rolfo. 2017. "The HMGB1/RAGE Pro-Inflammatory Axis in the Human Placenta: Modulating Effect of Low Molecular Weight Heparin" Molecules 22, no. 11: 1997. https://doi.org/10.3390/molecules22111997
APA StyleZenerino, C., Nuzzo, A. M., Giuffrida, D., Biolcati, M., Zicari, A., Todros, T., & Rolfo, A. (2017). The HMGB1/RAGE Pro-Inflammatory Axis in the Human Placenta: Modulating Effect of Low Molecular Weight Heparin. Molecules, 22(11), 1997. https://doi.org/10.3390/molecules22111997