A Review of the Biomedical Applications of Zerumbone and the Techniques for Its Extraction from Ginger Rhizomes


Abstract
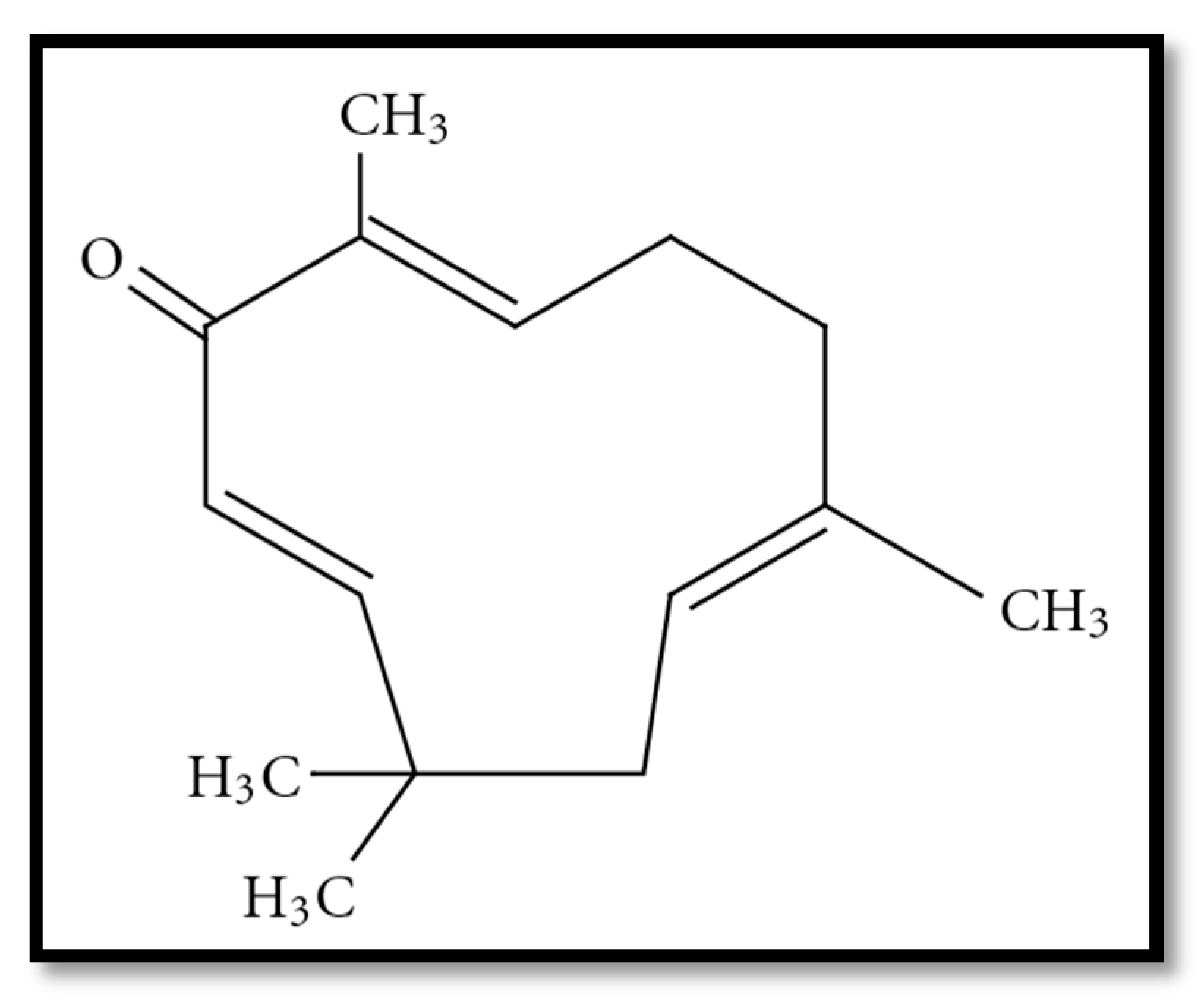
1. Introduction
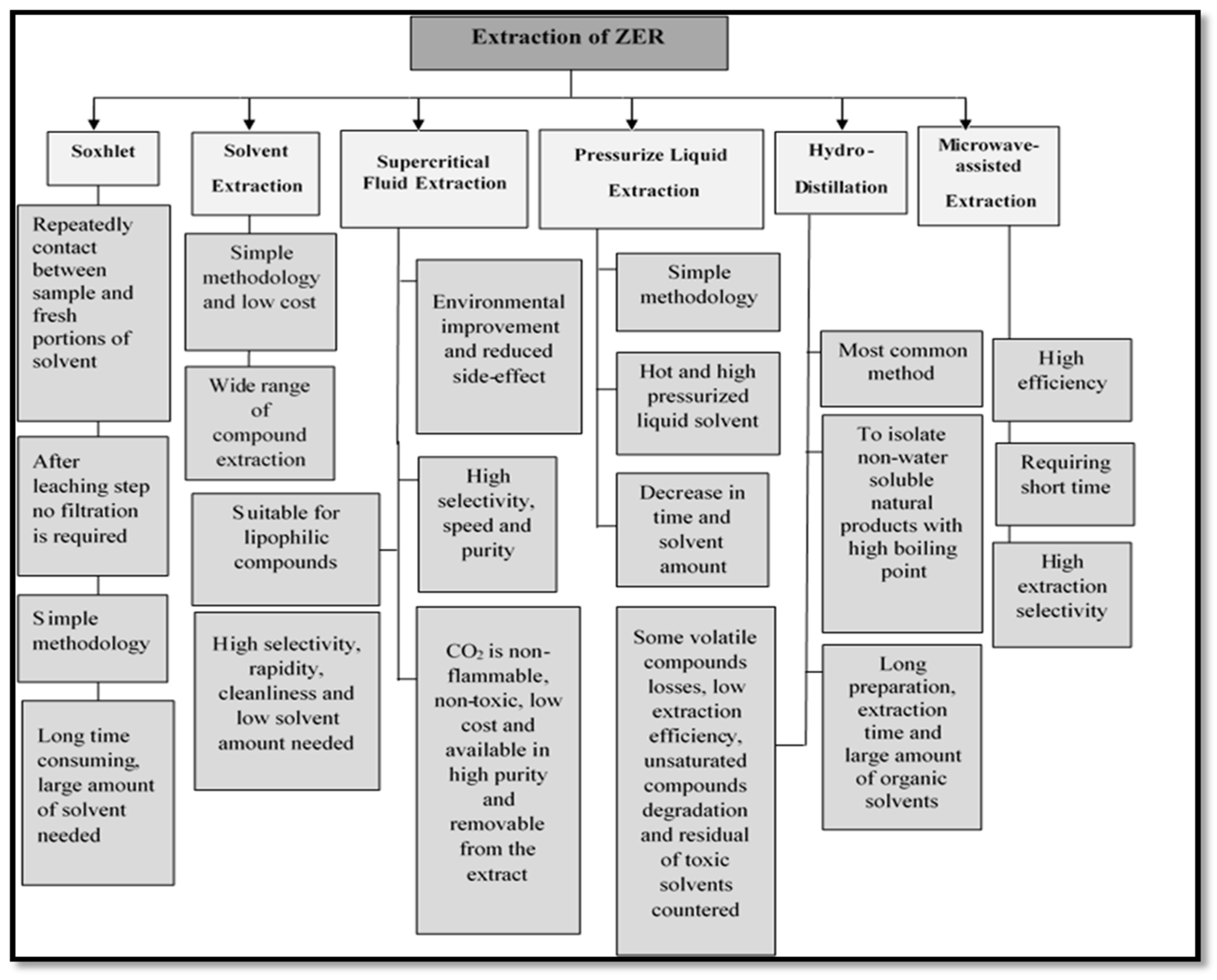
2. Different Extraction Methods of ZER
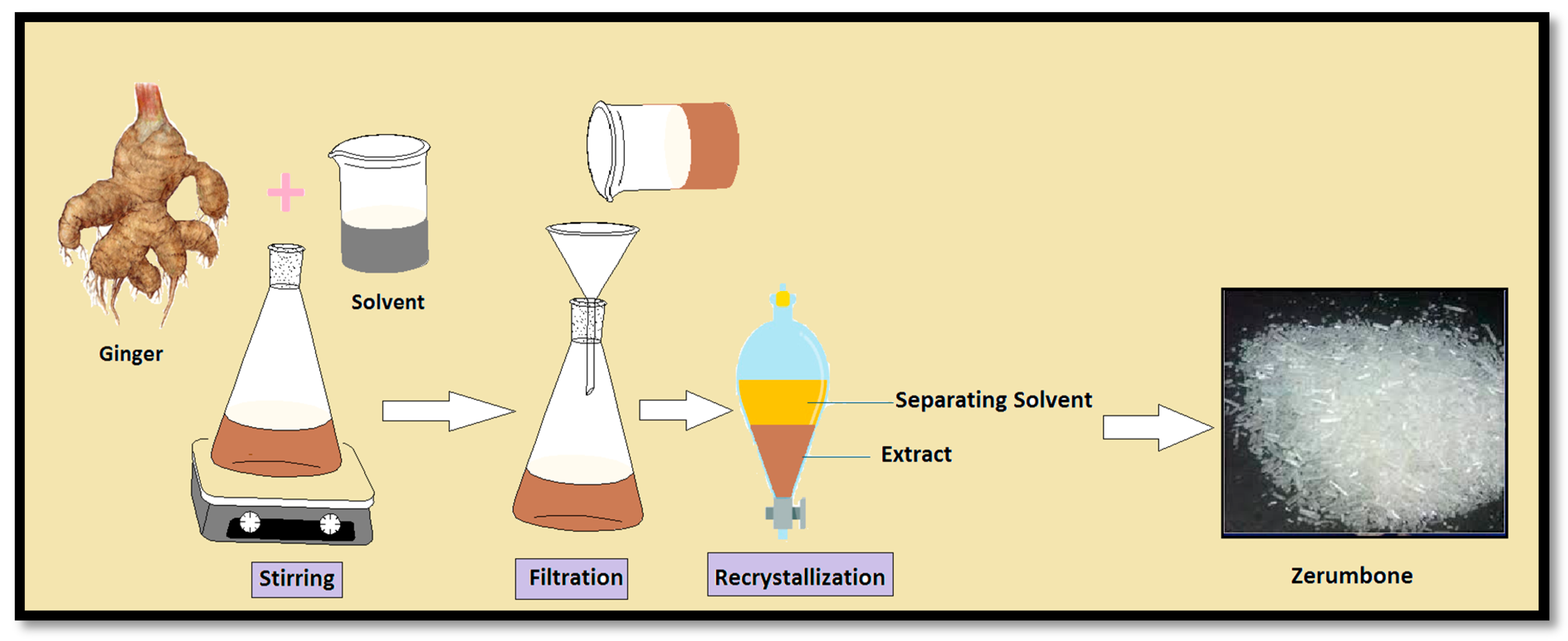
2.1. Solvent Extraction
2.2. Hydro Distillation
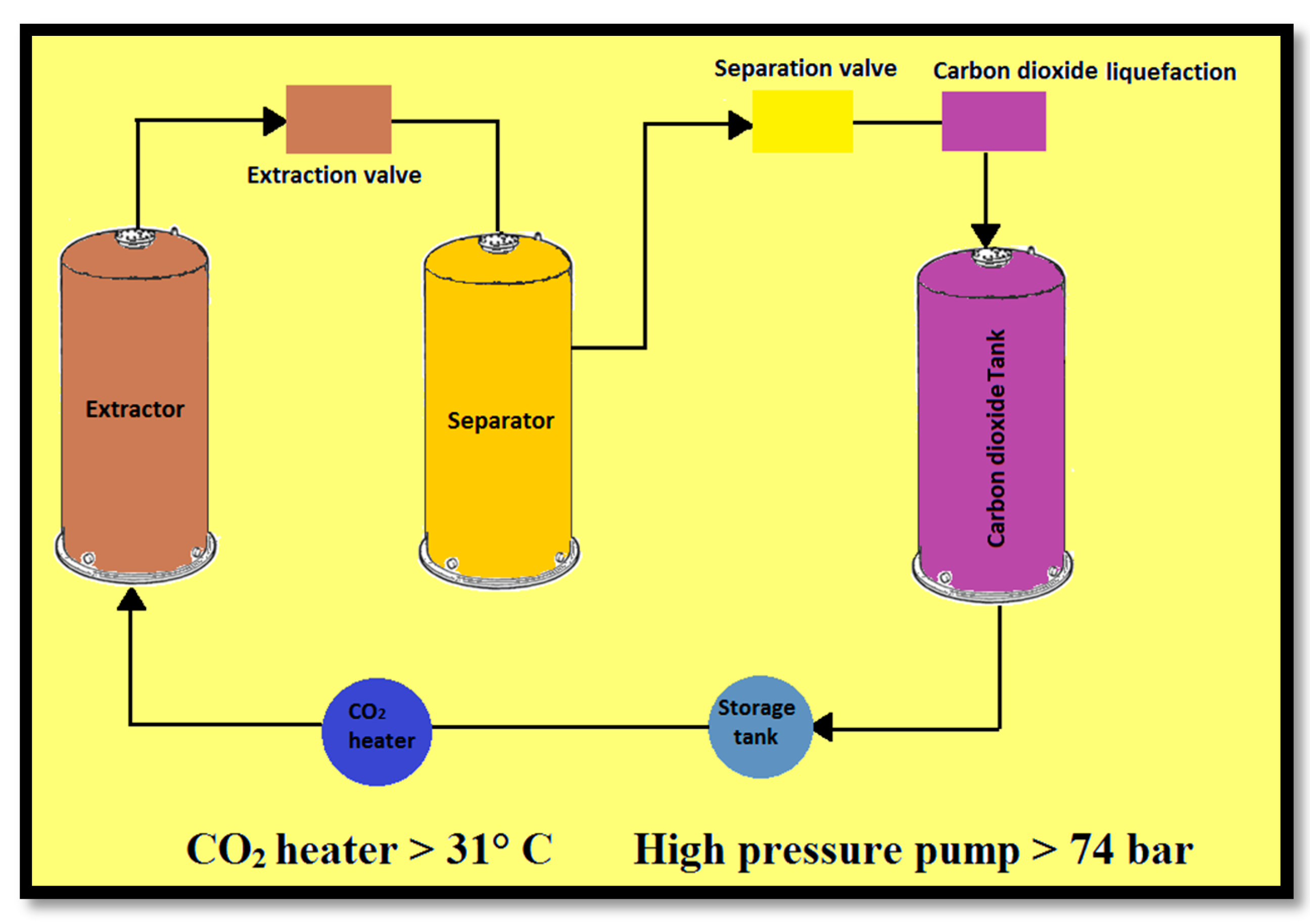
2.3. Supercritical Fluid Extraction (SFE)
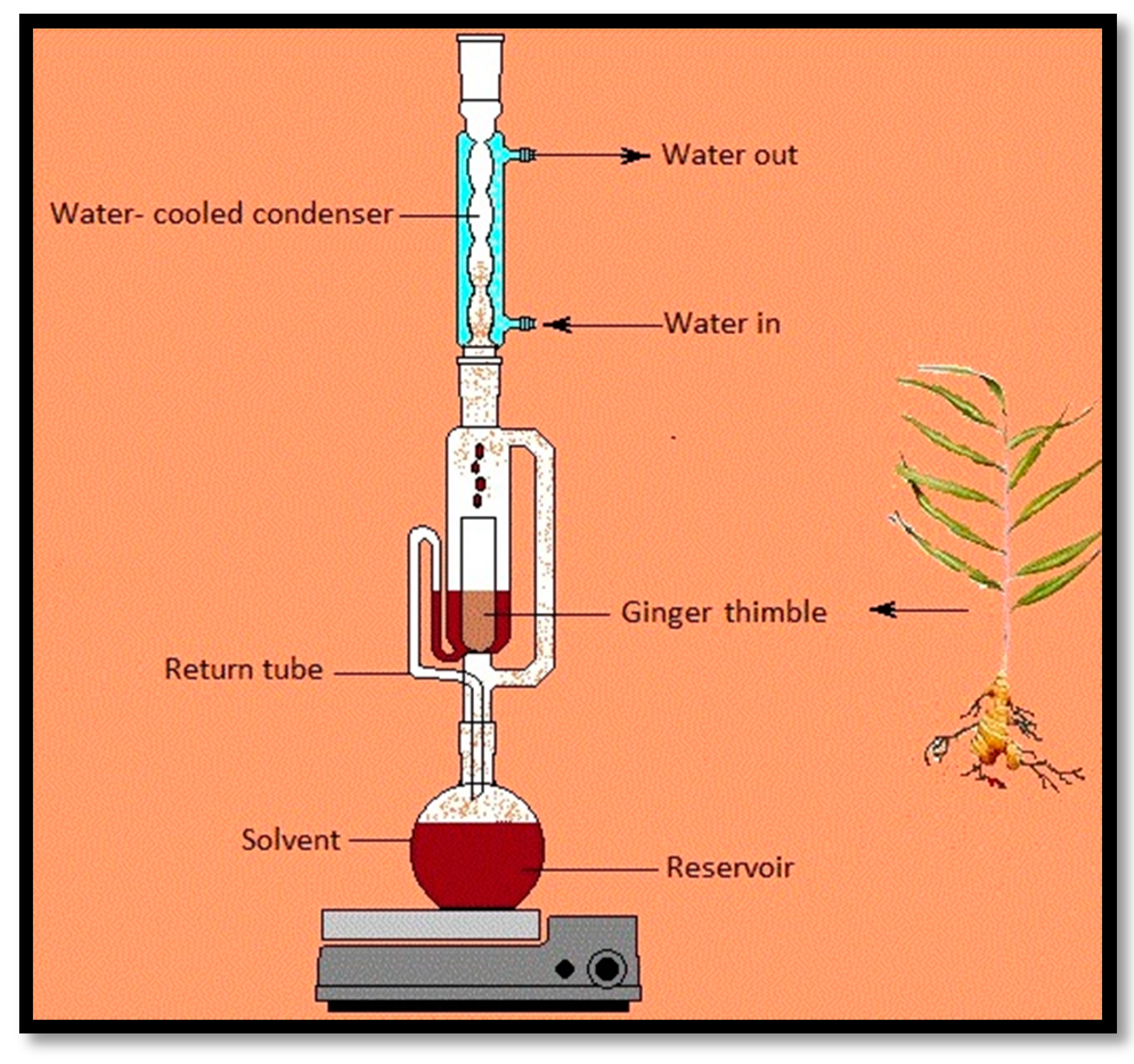
2.4. Soxhlet Extraction
2.5. Pressurized Liquid Extraction (PLE)
2.6. Microwave-Assisted Extraction
3. Biomedical Applications of ZER
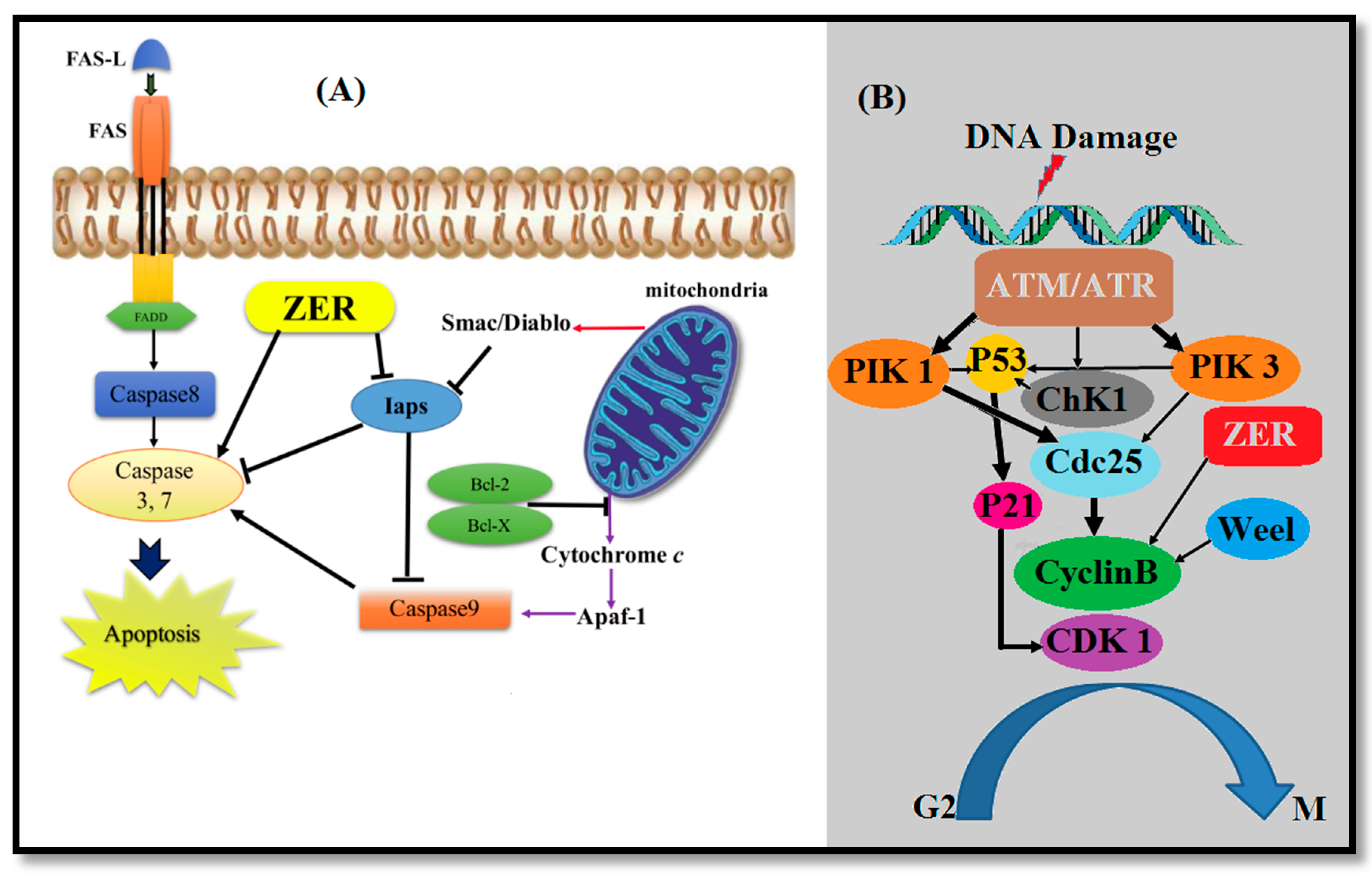
3.1. Anticancer Properties of ZER
3.1.1. Blood Cancer (Leukemia)
3.1.2. Breast Cancer
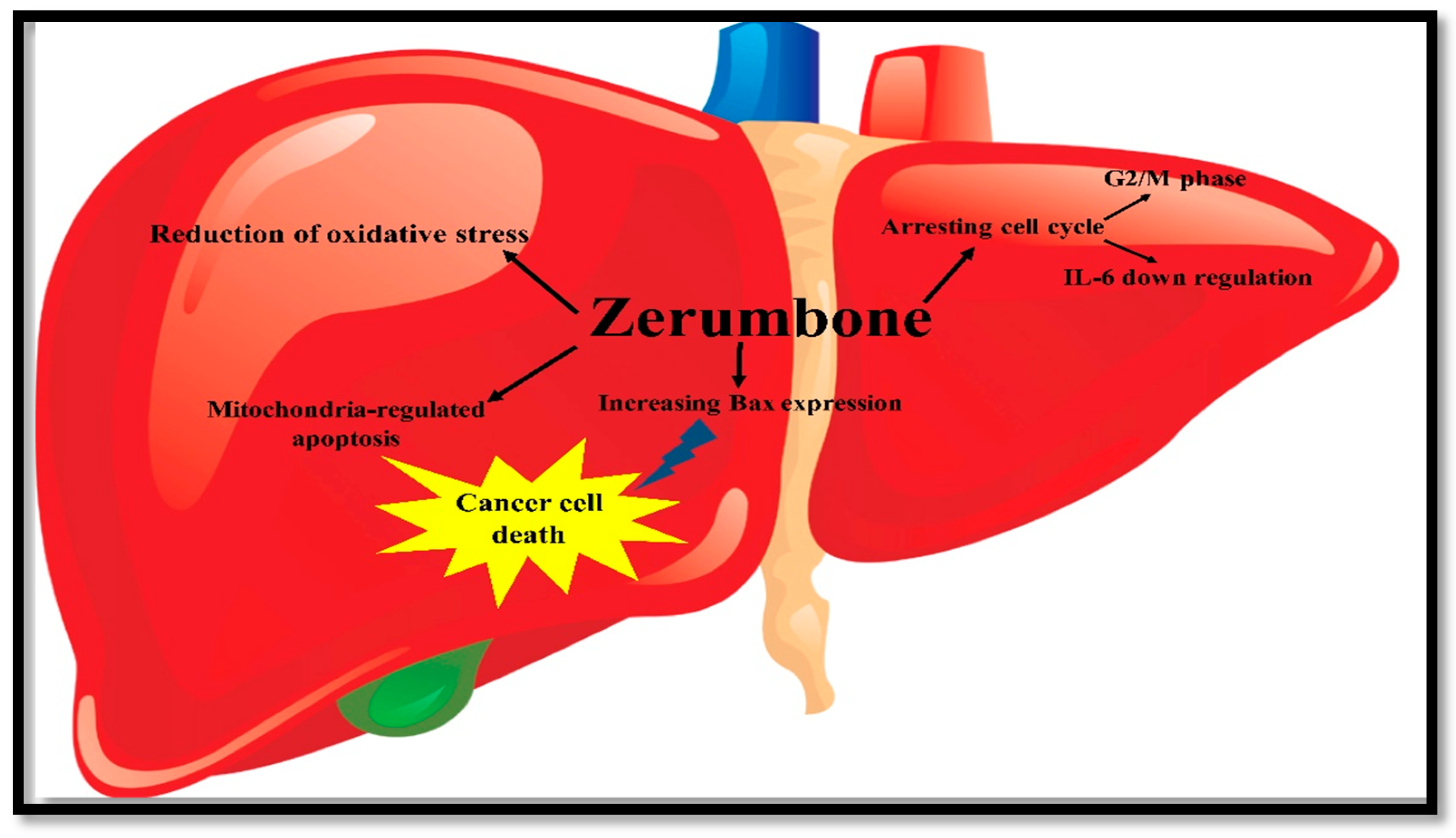
3.1.3. Liver Cancer
3.1.4. Colon Cancer
3.1.5. Lung Cancer
3.1.6. Pancreatic Cancer
3.1.7. Gastric Cancer
3.2. Anti-Inflammatory Activity
3.3. Antioxidant Activity
3.4. Immunomodulatory Activity
3.5. Anti Gastric Ulcer Activity
3.6. Antimicrobial Activity
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Singh, G.; Kapoor, I.; Singh, P.; de Heluani, C.S.; de Lampasona, M.P.; Catalan, C.A. Chemistry, antioxidant and antimicrobial investigations on essential oil and oleoresins of Zingiber officinale. Food Chem. Toxicol. 2008, 46, 3295–3302. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.V.; Rios, R. Inhibition of fungal growth on bread by volatile components from spices and herbs, and the possible application in active packaging, with special emphasis on mustard essential oil. Int. J. Food Microbiol. 2000, 60, 219–229. [Google Scholar] [CrossRef]
- Suhaj, M. Spice antioxidants isolation and their antiradical activity: A review. J. Food Compos. Anal. 2006, 19, 531–537. [Google Scholar] [CrossRef]
- Chuanmao, W. Extraction study on natural plant ingredients as preservative. Food Sci. 2000, 9, 006. [Google Scholar]
- Compton, M.E.; Koch, J.M. Influence of plant preservative mixture (ppm) tm on adventitious organogenesis in melon, petunia, and tobacco. In Vitro Cell. Dev. Biol.-Plant 2001, 37, 259–261. [Google Scholar] [CrossRef]
- Haraguchi, H. Antioxidative Plant Constituents; Taylor & Francis: London, UK, 2001; pp. 337–378. [Google Scholar]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.-P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Nostro, A.; Germano, M.; D’angelo, V.; Marino, A.; Cannatelli, M. Extraction methods and bioautography for evaluation of medicinal plant antimicrobial activity. Lett. Appl. Microbiol. 2000, 30, 379–384. [Google Scholar] [CrossRef] [PubMed]
- González-Lamothe, R.; Mitchell, G.; Gattuso, M.; Diarra, M.S.; Malouin, F.; Bouarab, K. Plant antimicrobial agents and their effects on plant and human pathogens. Int. J. Mol. Sci. 2009, 10, 3400–3419. [Google Scholar] [CrossRef] [PubMed]
- Chrystal, K.; Allan, S.; Forgeson, G.; Isaacs, R. The use of complementary/alternative medicine by cancer patients in a new zealand regional cancer treatment centre. Age 2003, 30, 31–50. [Google Scholar]
- Lampe, J.W. Health effects of vegetables and fruit: Assessing mechanisms of action in human experimental studies. Am. J. Clin. Nutr. 1999, 70, 475s–490s. [Google Scholar] [PubMed]
- Surh, Y.-J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Jikihara, H.; Qi, G.; Nozoe, K.; Hirokawa, M.; Sato, H.; Sugihara, Y.; Shimamoto, F. Aged garlic extract inhibits 1,2-dimethylhydrazine-induced colon tumor development by suppressing cell proliferation. Oncol. Rep. 2015, 33, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Kubatka, P.; Kapinová, A.; Kello, M.; Kruzliak, P.; Kajo, K.; Výbohová, D.; Mahmood, S.; Murin, R.; Viera, T.; Mojžiš, J. Fruit peel polyphenols demonstrate substantial anti-tumour effects in the model of breast cancer. Eur. J. Nutr. 2016, 55, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Kubatka, P.; Kello, M.; Kajo, K.; Kruzliak, P.; Výbohová, D.; Mojžiš, J.; Adamkov, M.; Fialová, S.; Veizerová, L.; Zulli, A. Oregano demonstrates distinct tumour-suppressive effects in the breast carcinoma model. Eur. J. Nutr. 2017, 56, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Montbriand, M.J. Alternative therapies. Health professionals’ attitudes. Can. Nurse 2000, 96, 22–26. [Google Scholar] [PubMed]
- Langner, E.; Greifenberg, S.; Gruenwald, J. Ginger: History and use. Adv. Ther. 1997, 15, 25–44. [Google Scholar]
- Da Rocha, A.B.; Lopes, R.M.; Schwartsmann, G. Natural products in anticancer therapy. Curr. Opin. Pharmacol. 2001, 1, 364–369. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.P.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Ramar, P.S.; Looi, C.Y. Cancer prevention and therapy through the modulation of transcription factors by bioactive natural compounds. In Seminars in Cancer Biology; Elsevier: Kidlington, UK, 2016; pp. 35–47. [Google Scholar]
- Larsen, K.; Ibrahim, H.; Khaw, S.; Saw, L. Gingers of Peninsular Malaysia and Singapore; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 1999. [Google Scholar]
- Rahman, H.S.; Rasedee, A.; Yeap, S.K.; Othman, H.H.; Chartrand, M.S.; Namvar, F.; Abdul, A.B.; How, C.W. Biomedical properties of a natural dietary plant metabolite, zerumbone, in cancer therapy and chemoprevention trials. BioMed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Yob, N.J.; Jofrry, S.M.; Affandi, M.; Teh, L.; Salleh, M.; Zakaria, Z. Zingiber zerumbet (L.) Smith: A review of its ethnomedicinal, chemical, and pharmacological uses. Evid. Based Complement. Altern. Med. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Grant, K.L.; Lutz, R.B. Alternative Therapies: Ginger. Am. J. Health-Syst. Pharm. 2000, 57, 945–947. [Google Scholar] [PubMed]
- Bordia, A.; Verma, S.; Srivastava, K. Effect of ginger (Zingiber officinale Rosc.) and fenugreek (Trigonella foenumgraecum L.) on blood lipids, blood sugar and platelet aggregation in patients with coronary artery disease. Prostaglandins Leukot. Essent. Fat. Acids 1997, 56, 379–384. [Google Scholar] [CrossRef]
- Shukla, Y.; Singh, M. Cancer preventive properties of ginger: A brief review. Food Chem. Toxicol. 2007, 45, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Dorai, T.; Aggarwal, B.B. Role of chemopreventive agents in cancer therapy. Cancer Lett. 2004, 215, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, M.N.I.; Chowdhury, J.U.; Begum, J. Chemical investigation of the leaf and rhizome essential oils of Zingiber zerumbet (L.) Smith from bangladesh. Bangladesh J. Pharmacol. 2008, 4, 9–12. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yoshida, C.; Murakami, A.; Ohigashi, H.; Osawa, T.; Uchida, K. Zerumbone, a tropical ginger sesquiterpene, activates phase ii drug metabolizing enzymes. FEBS Lett. 2004, 572, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, T.; Okamoto, T.; Hill, R.K.; Kawai, Y.; Takahashi, S.; Yonemori, S.; Yamamoto, Y.; Ohe, K.; Uemura, S.; Sawada, S. Chemistry of zerumbone. 1. Simplified isolation, conjugate addition reactions, and a unique ring contracting transannular reaction of its dibromide. J. Org. Chem. 1999, 64, 2667–2672. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, T.; Yamamoto, K.; Utsumi, R.; Takatani, M.; Hill, R.K.; Kawai, Y.; Sawada, S.; Okamoto, T. Chemistry of zerumbone. 2. Regulation of ring bond cleavage and unique antibacterial activities of zerumbone derivatives. Biosci. Biotechnol. Biochem. 2001, 65, 2193–2199. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, T.; Yokoi, T.; Kawai, Y.; Hill, R.K.; Morita, M.; Okamoto, T.; Yamamoto, Y.; Fokin, V.V.; Sharpless, K.B.; Sawada, S. The chemistry of zerumbone. Part 5: Structural transformation of the dimethylamine derivatives. Tetrahedron 2003, 59, 4857–4866. [Google Scholar] [CrossRef]
- Duve, R. Highlights of the chemistry and pharmacology of wild ginger Zingiber zerumbet Smith. Fiji Agric. J. 1980, 42, 41–43. [Google Scholar]
- Haque, M.A.; Jantan, I.; Arshad, L.; Bukhari, S.N.A. Exploring the immunomodulatory and anticancer properties of zerumbone. Food Funct. 2017. [Google Scholar] [CrossRef] [PubMed]
- Abdelwahab, S.I.; Abdul, A.B.; Zain, Z.N.M.; Hadi, A.H.A. Zerumbone inhibits interleukin-6 and induces apoptosis and cell cycle arrest in ovarian and cervical cancer cells. Int. Immunopharmacol. 2012, 12, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Prasannan, R.; Kalesh, K.A.; Shanmugam, M.K.; Nachiyappan, A.; Ramachandran, L.; Nguyen, A.H.; Kumar, A.P.; Lakshmanan, M.; Ahn, K.S.; Sethi, G. Key cell signaling pathways modulated by zerumbone: Role in the prevention and treatment of cancer. Biochem. Pharmacol. 2012, 84, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Abdelwahab, S.I.; Abdul, A.B.; Mohan, S.; Taha, M.M.E.; Syam, S.; Ibrahim, M.Y.; Mariod, A.A. Zerumbone induces apoptosis in T-acute lymphoblastic leukemia cells. Leuk. Res. 2011, 35, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Abdelwahab, S.I.; Abdul, A.B.; Devi, N.; Taha, M.M.E.; Al-Zubairi, A.S.; Mohan, S.; Mariod, A.A. Regression of cervical intraepithelial neoplasia by zerumbone in female Balb/c mice prenatally exposed to diethylstilboestrol: Involvement of mitochondria-regulated apoptosis. Exp. Toxicol. Pathol. 2010, 62, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Kirana, C.; McIntosh, G.H.; Record, I.R.; Jones, G.P. Antitumor activity of extract of Zingiber aromaticum and its bioactive sesquiterpenoid zerumbone. Nutr. Cancer 2003, 45, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, N.E.; Abu, N.; Rahman, H.S.; Ky, H.; Ho, W.Y.; Lim, K.L.; How, C.W.; Rasedee, A.; Alitheen, N.B.; Yeap, S.K. Nanostructured lipid carrier improved in vivo anti-tumor and immunomodulatory effect of zerumbone in 4t1 challenged mice. RSC Adv. 2015, 5, 22066–22074. [Google Scholar] [CrossRef]
- Jegannathan, S.D.; Arul, S.; Dayalan, H. Zerumbone, a sesquiterpene, controls proliferation and induces cell cycle arrest in human laryngeal carcinoma cell line Hep-2. Nutr. Cancer 2016, 68, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, Y.; Ohnishi, K.; Irie, K.; Murakami, A. Possible contribution of zerumbone-induced proteo-stress to its anti-inflammatory functions via the activation of heat shock factor 1. PLoS ONE 2016, 11, e0161282. [Google Scholar] [CrossRef] [PubMed]
- Azam, F.; Amer, A.M.; Abulifa, A.R.; Elzwawi, M.M. Ginger components as new leads for the design and development of novel multi-targeted anti-alzheimer’s drugs: A computational investigation. Drug Des. Dev. Ther. 2014, 8, 2045. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.B.; Seo, W.-D.; Lee, Y.-J.; Lee, Y.-S.; Lee, H.-J. Toxicological evaluation of zerumbone on antitumor effects in mice. Afr. J. Pharm. Pharmacol. 2013, 7, 466–473. [Google Scholar] [CrossRef]
- Ohnishi, K.; Nakahata, E.; Irie, K.; Murakami, A. Zerumbone, an electrophilic sesquiterpene, induces cellular proteo-stress leading to activation of ubiquitin–proteasome system and autophagy. Biochem. Biophys. Res. Commun. 2013, 430, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Takahashi, M.; Jiwajinda, S.; Koshimizu, K.; Ohigashi, H. Identification of zerumbone in Zingiber zerumbet smith as a potent inhibitor of 12-O-tetradecanoylphorbol-13-acetate-induced epstein-barr virus activation. Biosci. Biotechnol. Biochem. 1999, 63, 1811–1812. [Google Scholar] [CrossRef] [PubMed]
- Tzheng, T.-F.; Liou, S.-S.; Ju, C.; Liu, I.-M. The ethanol extract of Zingiber zerumbet attenuates streptozotocin-induced diabetic nephropathy in rats. Evid.-Based Complement. Altern. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Amer, N.; Khuder, M.H.; Yacoub, S.A.; Baker, N.N. Histological effects of excessive consumption of Zingeber officinale on liver and spleen of the mice. J. Al Nahrain Univ. 2013, 16, 151–156. [Google Scholar]
- Sriphanaa, U.; Pitchuanchoma, S.; Kongsaereeb, P.; Yenjaia, C. Antimalarial activity and cytotoxicity of zerumbone derivatives. ScienceAsia 2013, 39, 95–99. [Google Scholar] [CrossRef]
- Hong, T.-Y.; Tzeng, T.-F.; Liou, S.-S.; Liu, I.-M. The ethanol extract of Zingiber zerumbet rhizomes mitigates vascular lesions in the diabetic retina. Vasc. Pharmacol. 2015, 76, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Setiyono, A. Indonesian wild ginger (Zingiber sp.) extract: Antibacterial activity against mycoplasma gallisepticum. IOSR J. Pharm. 2015, 5, 59–64. [Google Scholar]
- Murakami, A.; Hayashi, R.; Takana, T.; Kwon, K.H.; Ohigashi, H.; Safitri, R. Suppression of dextran sodium sulfate-induced colitis in mice by zerumbone, a subtropical ginger sesquiterpene, and nimesulide: Separately and in combination. Biochem. Pharmacol. 2003, 66, 1253–1261. [Google Scholar] [CrossRef]
- Huang, G.-C.; Chien, T.-Y.; Chen, L.-G.; Wang, C.-C. Antitumor effects of zerumbone from Zingiber zerumbet in p-388d1 cells in vitro and in vivo. Plant. Med. 2005, 71, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Chien, T.; Chen, L.; Lee, C.; Lee, F.; Wang, C. Anti-inflammatory constituents of Zingiber zerumbet. Food Chem. 2008, 110, 584–589. [Google Scholar] [CrossRef]
- Shieh, Y.-H.; Huang, H.-M.; Wang, C.-C.; Lee, C.-C.; Fan, C.-K.; Lee, Y.-L. Zerumbone enhances the Th1 response and ameliorates ovalbumin-induced th2 responses and airway inflammation in mice. Int. Immunopharmacol. 2015, 24, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Norulaini, N.N.; Anuar, O.; Omar, A.; AlKarkhi, A.F.; Setianto, W.B.; Fatehah, M.; Sahena, F.; Zaidul, I. Optimization of SC–CO2 extraction of zerumbone from Zingiber zerumbet (L.) Smith. Food Chem. 2009, 114, 702–705. [Google Scholar] [CrossRef]
- Ibrahim, M.Y.; Abdul, A.B.; Ibrahim, T.A.T.; Wahab, S.A.; Elhassan, M.M.; Mohan, S. Attenuation of cisplatin-induced nephrotoxicity in rats using zerumbone. Afr. J. Biotechnol. 2012, 9, 4434–4441. [Google Scholar]
- Sivasothy, Y.; Awang, K.; Ibrahim, H.; Thong, K.; Fitrah, N.; Koh, X.; Tan, L. Chemical composition and antibacterial activities of essential oils from Zingiber spectabile griff. J. Essent. Oil Res. 2012, 24, 305–313. [Google Scholar] [CrossRef]
- Das, A.; Kasoju, N.; Bora, U.; Rangan, L. Chemico-biological investigation of rhizome essential oil of Zingiber moran—Native to northeast India. Med. Chem. Res. 2013, 22, 4308–4315. [Google Scholar] [CrossRef]
- Srivastava, A.; Srivastava, S.; Shah, N. Essential oil composition of Zingiber zerumbet (L.) sm. From India. J. Essent. Oil Res. 2000, 12, 595–597. [Google Scholar] [CrossRef]
- Kishore, N.; Dwivedi, R. Zerumbone: A potential fungitoxic agent isolated from Zingiber cassumunar roxb. Mycopathologia 1992, 120, 155–159. [Google Scholar] [CrossRef]
- Omar, M.N.; Razman, S.; Nor, M.; Nazreen, M.M.; Zuberdi, A.M. Supercritical fluid extraction (Sfe) of malaysian wild ginger Zingiber puberulum inflorescence. Orient. J. Chem. 2013, 29, 89–92. [Google Scholar] [CrossRef]
- Yonei, Y.; Ōhinata, H.; Yoshida, R.; Shimizu, Y.; Yokoyama, C. Extraction of ginger flavor with liquid or supercritical carbon dioxide. J. Supercrit. Fluids 1995, 8, 156–161. [Google Scholar] [CrossRef]
- De Castro, M.L.; Garcıa-Ayuso, L. Soxhlet extraction of solid materials: An outdated technique with a promising innovative future. Anal. Chim. Acta 1998, 369, 1–10. [Google Scholar] [CrossRef]
- Ahmed Hamdi, O.A.; Syed Abdul Rahman, S.N.; Awang, K.; Abdul Wahab, N.; Looi, C.Y.; Thomas, N.F.; Abd Malek, S.N. Cytotoxic constituents from the rhizomes of Curcuma zedoaria. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Jalil, M.; Annuar, M.S.M.; Tan, B.C.; Khalid, N. Effects of selected physicochemical parameters on zerumbone production of Zingiber zerumbet smith cell suspension culture. Evid. Based Complement. Altern. Med. 2015, 21, 426–437. [Google Scholar]
- Thang, T.D.; Hung, N.H.; Dung, D.M.; Dai, D.N.; Ogunwande, I.A. Volatile terpenes from essential oils of selected medicinal plants grown in vietnam forest reserves. J. Herbs Spices Med. Plants 2015, 21, 426–437. [Google Scholar] [CrossRef]
- Norfazlina, M.; Farida Zuraina, M.; Rajab, N.; Mohd Nazip, S.; Rumiza, A.; Suziana Zaila, C. In vitro cytotoxicity effects of single and combination nigella sativa and zingiber zerumbet extracts on human myeloid leukemia (Hl60) cells and its mode of cell death. J. Appl. Pharm. Sci. 2014, 4, 51–55. [Google Scholar]
- Chen, L.; Jin, H.; Ding, L.; Zhang, H.; Li, J.; Qu, C.; Zhang, H. Dynamic microwave-assisted extraction of flavonoids from Herba Epimedii. Sep. Purif. Technol. 2008, 59, 50–57. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z.; Rahmat, A.; Swamy, M.K. Optimization of microwave-assisted extraction of zerumbone from Zingiber zerumbet L. Rhizome and evaluation of antiproliferative activity of optimized extracts. Chem. Cent. J. 2017, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Shafi, M.T.M.P. Rhizome essential oil composition of costus speciosus and its antimicrobial properties. Int. J. 2015, 4, 28–32. [Google Scholar]
- Park, J.-H.; Park, G.M.; Kim, J.-K. Zerumbone, sesquiterpene photochemical from ginger, inhibits angiogenesis. Korean J. Physiol. Pharmacol. 2015, 19, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, S. The rapidly emerging role of zerumbone in attenuating tumor growth in systemic malignancies. Mol. Nutr. Food Res. 2012, 56, 1487. [Google Scholar] [CrossRef] [PubMed]
- Deorukhkar, A.; Ahuja, N.; Mercado, A.; Diagaradjane, P.; Mohindra, P.; Guha, S.; Aggarwal, B.; Krishnan, S. Zerumbone, a sesquiterpene from Southeast Asian edible ginger sensitizes colorectal cancer cells to radiation therapy. Int. J. Radiat. Oncol. Biol. Phys. 2010, 78, S654. [Google Scholar] [CrossRef]
- Weng, H.-Y.; Hsu, M.-J.; Wang, C.-C.; Chen, B.-C.; Hong, C.-Y.; Chen, M.-C.; Chiu, W.-T.; Lin, C.-H. Zerumbone suppresses ikkα, akt, and foxo1 activation, resulting in apoptosis of gbm 8401 cells. J. Biomed. Sci. 2012, 19, 86. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, A.; Spetner, L.; Burke, M. Redox-regulated mechanism may account for zerumbone’s ability to suppress cancer-cell proliferation. Carcinogenesis 2002, 23, 1961–1962. [Google Scholar] [CrossRef] [PubMed]
- Samad, N.A.; Abdul, A.B.; Abdullah, R.; Ibrahim, T.A.T.; Rahman, H.; Keong, Y.S. Zerumbone (zer) induces apoptosis in Hepg2 cells via mitochondrial pathway. Int. J. Pharm. Pharm. Sci. 2015, 7, 599–604. [Google Scholar]
- Wang, C.; Zou, S.; Cui, Z.; Guo, P.; Meng, Q.; Shi, X.; Gao, Y.; Yang, G.; Han, Z. Zerumbone protects ins-1 rat pancreatic beta cells from high glucose-induced apoptosis through generation of reactive oxygen species. Biochem. Biophys. Res. Commun. 2015, 460, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Coffman, R.; Jorvig, J. Zerumbone, a phytochemical from asian ginger is a novel inhibitor of Jak2/Stat3 inhibits promigratory gene expression, growth and migration of pancreatic cancer cells. Pancreatology 2013, 13, e18–e19. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Q.; Liu, Y.; Qiao, H.; Liu, Y. Zerumbone, a southeast asian ginger sesquiterpene, induced apoptosis of pancreatic carcinoma cells through p53 signaling pathway. Evid. Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Shamoto, T.; Matsuo, Y.; Shibata, T.; Tsuboi, K.; Nagasaki, T.; Takahashi, H.; Funahashi, H.; Okada, Y.; Takeyama, H. Zerumbone inhibits angiogenesis by blocking nf-κb activity in pancreatic cancer. Pancreas 2014, 43, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Van Truong, V.; Nam, T.D.; Hung, T.N.; Nga, N.T.; Quan, P.M.; Van Chinh, L.; Jung, S.-H. Synthesis and anti-proliferative activity of novel azazerumbone conjugates with chalcones. Bioorg. Med. Chem. Lett. 2015, 25, 5182–5185. [Google Scholar] [CrossRef] [PubMed]
- Hseu, Y.-C.; Huang, Y.-C.; Korivi, M.; Wu, J.-J.; Way, T.-D.; Ou, T.-T.; Chiu, L.-W.; Lee, C.-C.; Lin, M.-L.; Yang, H.-L. Zerumbone attenuates Tgf-β1-mediated epithelial–mesenchymal transition via upregulated E-cadherin expression and downregulated smad2 signalling pathways in non-small cell lung cancer (A549) cells. J. Funct. Foods 2015, 18, 58–72. [Google Scholar] [CrossRef]
- Pitchuanchom, S.; Songsiang, U.; Weerapreeyakul, N.; Yenjai, C. Anticancer activity of the bioreductive and non-bioreductive zerumbone derivatives. Lett. Drug Des. Discov. 2011, 8, 536–543. [Google Scholar] [CrossRef]
- Hu, Z.; Zeng, Q.; Zhang, B.; Liu, H.; Wang, W. Promotion of p53 expression and reactive oxidative stress production is involved in zerumbone-induced cisplatin sensitization of non-small cell lung cancer cells. Biochimie 2014, 107, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, K.; Irie, K.; Murakami, A. Modulation of protein quality control systems as novel mechanisms underlying functionality of food phytochemicals. Funct. Foods Health Dis. 2013, 3, 400–415. [Google Scholar]
- Sehrawat, A.; Sakao, K.; Singh, S.V. Notch2 activation is protective against anticancer effects of zerumbone in human breast cancer cells. Breast Cancer Res. Treat. 2014, 146, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Fatima, A.; Abdul, A.B.H.; Abdullah, R.; Karjiban, R.A.; Lee, V.S. Binding mode analysis of zerumbone to key signal proteins in the tumor necrosis factor pathway. Int. J. Mol. Sci. 2015, 16, 2747–2766. [Google Scholar] [CrossRef] [PubMed]
- Hosseinpour, M.; Abdul, A.B.; Rahman, H.S.; Rasedee, A.; Yeap, S.K.; Ahmadi, N.; Othman, H.H.; Chartrand, M.S. Comparison of apoptotic inducing effect of zerumbone and zerumbone-loaded nanostructured lipid carrier on human mammary adenocarcinoma Mda-mb-231 cell line. J. Nanomater. 2014, 2014. [Google Scholar] [CrossRef]
- Han, J.; Bae, S.Y.; Oh, S.J.; Lee, J.; Lee, J.H.; Lee, H.C.; Lee, S.K.; Kil, W.H.; Kim, S.W.; Nam, S.J. Zerumbone suppresses il-1β-induced cell migration and invasion by inhibiting Il-8 and Mmp-3 expression in human triple-negative breast cancer cells. Phytother. Res. 2014, 28, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- Sehrawat, A.; Arlotti, J.A.; Murakami, A.; Singh, S.V. Zerumbone causes bax-and bak-mediated apoptosis in human breast cancer cells and inhibits orthotopic xenograft growth in vivo. Breast Cancer Res. Treat. 2012, 136, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kil, W.H.; Lee, J.; Oh, S.-J.; Han, J.; Jeon, M.; Jung, T.; Lee, S.K.; Bae, S.Y.; Lee, H.C. Zerumbone suppresses Egf-induced Cd44 expression through the inhibition of stat3 in breast cancer cells. Oncol. Rep. 2014, 32, 2666–2672. [Google Scholar] [CrossRef] [PubMed]
- Rajan, I.; Jayasree, P.; Kumar, P.M. Zerumbone induces mitochondria-mediated apoptosis via increased calcium, generation of reactive oxygen species and upregulation of soluble histone H2ax in k562 chronic myelogenous leukemia cells. Tumor Biol. 2015, 36, 8479–8489. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.S.; Rasedee, A.; How, C.W.; Zeenathul, N.A.; Chartrand, M.S.; Yeap, S.K.; Abdul, A.B.; Tan, S.W.; Othman, H.H.; Ajdari, Z. Antileukemic effect of zerumbone-loaded nanostructured lipid carrier in Wehi-3b cell-induced murine leukemia model. Int. J. Nanomed. 2015, 10, 1649. [Google Scholar] [CrossRef] [PubMed]
- Norfazlina, M.; Zuraina, M.F.; Rajab, N.; Nazip, S.M.; Rumiza, A.; Zaila, C.S.; Mun, L.L.; Nurshahirah, N.; Florinsiah, L. Cytotoxicity study of nigella sativa and Zingiber zerumbet extracts, thymoquinone and zerumbone isolated on human myeloid leukemia (Hl60) cell. Open Conf. Proc. J. 2014, 4, 99–107. [Google Scholar] [CrossRef]
- Al-Zubairi, A.S.; Abdul, A.B.; Syam, M. Evaluation of the genotoxicity of zerumbone in cultured human peripheral blood lymphocytes. Toxicol. In Vitro 2010, 24, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Keong, Y.S.; Alitheen, N.B.; Mustafa, S.; Aziz, S.A.; Rahman, M.A.; Ali, A.M. Immunomodulatory effects of zerumbone isolated from roots of zingiber zerumbet. Pak. J. Pharm. Sci. 2010, 23, 75–82. [Google Scholar] [PubMed]
- Al-Zubairi, A.S.; Abdul, A.B.; Yousif, M.; Abdelwahab, S.I.; Elhassan, M.M.; Mohan, S. In vivo and in vitro genotoxic effects of zerumbone. Caryologia 2010, 63, 11–17. [Google Scholar] [CrossRef]
- Deorukhkar, A.; Ahuja, N.; Mercado, A.L.; Diagaradjane, P.; Raju, U.; Patel, N.; Mohindra, P.; Diep, N.; Guha, S.; Krishnan, S. Zerumbone increases oxidative stress in a thiol-dependent Ros-independent manner to increase DNA damage and sensitize colorectal cancer cells to radiation. Cancer Med. 2015, 4, 278–292. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.-L.; Liang, J.-W.; Hsu, L.-C.; Chang, W.-L.; Lee, S.-S.; Guh, J.-H. Zerumbone, a ginger sesquiterpene, induces apoptosis and autophagy in human hormone-refractory prostate cancers through tubulin binding and crosstalk between endoplasmic reticulum stress and mitochondrial insult. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 1223–1236. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Miller, R.; Lee, P.; Holden, C.A.; Lindhorst, S.M.; Jaboin, J.; Vandergrift, W.A.; Banik, N.L.; Giglio, P.; Varma, A.K. A novel component from citrus, ginger, and mushroom family exhibits antitumor activity on human meningioma cells through suppressing the Wnt/β-catenin signaling pathway. Tumor Biol. 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Rajendran, P.; Li, F.; Kim, C.; Sikka, S.; Siveen, K.S.; Kumar, A.P.; Ahn, K.S.; Sethi, G. Abrogation of Stat3 signaling cascade by zerumbone inhibits proliferation and induces apoptosis in renal cell carcinoma xenograft mouse model. Mol. Carcinog. 2014, 10, 971–985. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, M.R.; Perimal, E.K.; Akhtar, M.N.; Mohamad, A.; Khalid, M.H.; Tasrip, N.A.; Mokhtar, F.; Zakaria, Z.A.; Lajis, N.; Israf, D. Anti-inflammatory effect of zerumbone on acute and chronic inflammation models in mice. Fitoterapia 2010, 81, 855–858. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, T.-F.; Liou, S.-S.; Chang, C.J.; Liu, I.-M. Zerumbone, a natural cyclic sesquiterpene of Zingiber zerumbet smith, attenuates nonalcoholic fatty liver disease in hamsters fed on high-fat diet. Evid. Based Complement. Altern. Med. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-W.; Ohnishi, K.; Murakami, A.; Lee, J.-S.; Kundu, J.K.; Na, H.-K.; Ohigashi, H.; Surh, Y.-J. Zerumbone induces heme oxygenase-1 expression in mouse skin and cultured murine epidermal cells through activation of Nrf2. Cancer Prev. Res. 2011, 4, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.M.E.; Abdul, A.B.; Abdullah, R.; Ibrahim, T.A.T.; Abdelwahab, S.I.; Mohan, S. Potential chemoprevention of diethylnitrosamine-initiated and 2-acetylaminofluorene-promoted hepatocarcinogenesis by zerumbone from the rhizomes of the subtropical ginger (Zingiber zerumbet). Chem. Biol. Interact. 2010, 186, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-Y.; Lin, D.P.-C.; Su, K.-C.; Chen, Y.-L.; Wu, C.-Y.; Teng, M.-C.; Tsai, Y.-T.; Sun, C.-Y.; Wang, S.-R.; Chang, H.-H. Dietary zerumbone prevents against ultraviolet b-induced cataractogenesis in the mouse. Mol. Vis. 2011, 17, 723–730. [Google Scholar] [PubMed]
- Xian, M.; Ito, K.; Nakazato, T.; Shimizu, T.; Chen, C.K.; Yamato, K.; Murakami, A.; Ohigashi, H.; Ikeda, Y.; Kizaki, M. Zerumbone, a bioactive sesquiterpene, induces g2/M cell cycle arrest and apoptosis in leukemia cells via a fas-and mitochondria-mediated pathway. Cancer Sci. 2007, 98, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Anasamy, T.; Abdul, A.B.; Sukari, M.A.; Abdelwahab, S.I.; Mohan, S.; Kamalidehghan, B.; Azid, M.Z.; Muhammad Nadzri, N.; Andas, A.; Kuan Beng, N. A phenylbutenoid dimer, cis-3-(3′,4′-dimethoxyphenyl)-4-[(e)-3′′′,4′′′-dimethoxystyryl] cyclohex-1-ene, exhibits apoptogenic properties in T-acute lymphoblastic leukemia cells via induction of p53-independent mitochondrial signalling pathway. Evid. Based Complement. Altern. Med. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.S.; Rasedee, A.; How, C.W.; Abdul, A.B.; Zeenathul, N.A.; Othman, H.H.; Saeed, M.I.; Yeap, S.K. Zerumbone-loaded nanostructured lipid carriers: Preparation, characterization, and antileukemic effect. Int. J. Nanomed. 2013, 8, 2769. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.; Gogineni, V.; Saeian, K. Epidemiology of primary and secondary liver cancers. In Seminars in Interventional Radiology; Thieme Medical Publishers: Stuttgart, Germany, 2006; p. 47. [Google Scholar]
- Al-Saffar, F. Effects on renal histology, renal metabolizing enzymes and oxidative stress biomarkers in rats administered zerumbone against osteoarthritis. Asian J. Cell Biol. 2015, 10, 66–79. [Google Scholar]
- Murakami, A.; Miyamoto, M.; Ohigashi, H. Zerumbone, an anti-inflammatory phytochemical, induces expression of proinflammatory cytokine genes in human colon adenocarcinoma cell lines. Biofactors 2004, 21, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Tsim, S.; O’Dowd, C.; Milroy, R.; Davidson, S. Staging of non-small cell lung cancer (Nsclc): A review. Respir. Med. 2010, 104, 1767–1774. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, K.; Matsuo, Y.; Shamoto, T.; Koide, S.; Ochi, N.; Takahashi, H.; Wakasugi, T.; Funahashi, H.; Kimura, M.; Sato, M. Zerumbone inhibits tumor angiogenesis in gastric cancer. J. Surg. Res. 2012, 172, 193. [Google Scholar] [CrossRef]
- Tsuboi, K.; Matsuo, Y.; Shamoto, T.; Shibata, T.; Koide, S.; Morimoto, M.; Guha, S.; Sung, B.; Aggarwal, B.B.; Takahashi, H. Zerumbone inhibits tumor angiogenesis via Nf-κB in gastric cancer. Oncol. Rep. 2014, 31, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Somchit, M.; Mak, J.; Ahmad Bustamam, A.; Zuraini, A.; Arifah, A.; Adam, Y.; Zakaria, Z. Zerumbone isolated from Zingiber zerumbet inhibits inflammation and pain in rats. J. Med. Plants Res. 2012, 6, 177–180. [Google Scholar] [CrossRef]
- Ganabadi, S.; Kadir, S.F.A. Zitumboihi’s effect on major histocompatibility complex type II cells in synovial membrane of osteoarthritic joint. Res. J. Vet. Sci. 2009, 2, 14–20. [Google Scholar] [CrossRef]
- Chien, T.-Y.; Huang, S.K.-H.; Lee, C.-J.; Tsai, P.-W.; Wang, C.-C. Antinociceptive and anti-inflammatory effects of zerumbone against mono-iodoacetate-induced arthritis. Int. J. Mol. Sci. 2016, 17, 249. [Google Scholar] [CrossRef] [PubMed]
- Wenhong, D.; Jia, Y.; Weixing, W.; Xiaoyan, C.; Chen, C.; Sheng, X.; Hao, J. Zerumbone attenuates the severity of acute necrotizing pancreatitis and pancreatitis-induced hepatic injury. Mediat. Inflamm. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Al-Saffar, F.; Ganabadi, S.; Fakurazi, S.; Yaakub, H. Zerumbone significantly improved immunoreactivity in the synovium compared to channa striatus extract in monosodium iodoacetate (Mia)-induced knee osteoarthritis in rat. J. Med. Plants Res. 2011, 5, 1701–1710. [Google Scholar]
- Chen, B.-Y.; Lin, D.P.-C.; Wu, C.-Y.; Teng, M.-C.; Sun, C.-Y.; Tsai, Y.-T.; Su, K.-C.; Wang, S.-R.; Chang, H.-H. Dietary zerumbone prevents mouse cornea from Uvb-induced photokeratitis through inhibition of Nf-κB, Inos, and Tnf-α expression and reduction of mda accumulation. Mol. Vis. 2011, 17, 854. [Google Scholar] [PubMed]
- Hamdi, O.A.A.; Ye, L.J.; Alfarizal, M.N. Neuroprotective and antioxidant constituents from Curcuma zedoaria rhizomes. Rec. Natl. Prod. 2015, 9, 349. [Google Scholar]
- Sidahmed, H.M.A.; Hashim, N.M.; Abdulla, M.A.; Ali, H.M.; Mohan, S.; Abdelwahab, S.I.; Taha, M.M.E.; Fai, L.M.; Vadivelu, J. Antisecretory, gastroprotective, antioxidant and anti-helicobcter pylori activity of zerumbone from Zingiber zerumbet (L.) Smith. PLoS ONE 2015, 10, e0121060. [Google Scholar] [CrossRef] [PubMed]
- Mesomo, M.C.; de Paula Scheer, A.; Perez, E.; Ndiaye, P.M.; Corazza, M.L. Ginger (Zingiber officinale r.) extracts obtained using supercritical co 2 and compressed propane: Kinetics and antioxidant activity evaluation. J. Supercrit. Fluids 2012, 71, 102–109. [Google Scholar] [CrossRef]
- Shin, J.-W.; Murakami, A.; Ohigashi, H.; Johnson, D.; Johnson, J.; Na, H.-K.; Surh, Y.-J. Zerumbone, a Sesquiterpene Derived from Tropical Ginger, Induces Heme Oxygenase-1 Expression via Activation of Nrf2 Signaling in Mouse Epidermal Cells and Hairless Mouse Skin in Vivo. In Proceedings of the AACR Annual Meeting, San Diego, CA, USA, 12–16 April 2008. [Google Scholar]
- Baiubon, P.; Kunanusorn, P.; Khonsung, P.; Chiranthanut, N.; Panthong, A.; Rujjanawate, C. Gastroprotective activity of the rhizome ethanol extract of Zingiber simaoense Y. Qian in rats. J. Ethnopharmacol. 2016, 194, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Chantharangsikul, G.; Kitpati, W.; Soonthornchareonnon, N.; Sailasuta, A.; Itharat, A.; Suvitayavat, W. Mucus secretion stimulation: A mechanism in gastroprotective effect of Zingiber officinale. Thai J. Pharm. Sci. (TJPS) 2016, 40, 1–8. [Google Scholar]
- Kusters, J.G.; van Vliet, A.H.; Kuipers, E.J. Pathogenesis of helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.S.; Srinivas, P.; Negi, P.; Bettadaiah, B. Antibacterial and antimutagenic activities of novel zerumbone analogues. Food Chem. 2013, 141, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Rana, V.S.; Ahluwalia, V.; Shakil, N.A.; Prasad, L. Essential oil composition, antifungal, and seedling growth inhibitory effects of zerumbone from Zingiber zerumbet smith. J. Essent. Oil Res. 2017, 29, 320–329. [Google Scholar] [CrossRef]
- Tzeng, T.-F.; Lu, H.-J.; Liou, S.-S.; Chang, C.J.; Liu, I.-M. Lipid-lowering effects of zerumbone, a natural cyclic sesquiterpene of Zingiber zerumbet smith, in high-fat diet-induced hyperlipidemic hamsters. Food Chem. Toxicol. 2014, 69, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, V.; Boffa, I.; De Masi, G.; Zollo, M. Natural compounds for pediatric cancer treatment. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 389, 1–19. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Cell Line | Biological Effects of ZER | References |
|---|---|---|---|
| Pancreatic | INS-1 rat pancreatic b cells | Enhances the viability of INS-1 b cells (high glucose). ZER can attenuate significantly the apoptosis of high glucose-induced INS-1 cells. | [77] |
| Human pancreatic carcinoma (PaCa) | Excellent inhibitor of Jak2/Stat3, which inhibits the growing of promigratory gene as well as the expression and migration of cancer cells. | [78] | |
| Human pancreatic carcinoma (PANC-1 and SW1990) | Effect on cell viability inhibition and induces apoptosis in time-dependent manner. | [79] | |
| Human pancreatic carcinoma (PaCa) | By inhibition of NF-1D705B and NF-κB-dependent proangiogenic gene products, ZER can inhibit PaCa-associated angiogenesis. | [80] | |
| Lung | ATCC-HTB-57 cells | Some novel compounds fabricated through azazerumbone conjugation with 2,4-dihydroxychalcones. These compounds showed anti-proliferative activity against the LU-1, Hep-G2, MCF-7 and SW480 human cancer cell lines. | [81] |
| TGF-β1-stimulated human (A549) cells | Demonstrates the anti-EMT and anti-metastatic properties of zerumbone in A549 lung cancer cells under TGF-β1-stimulation. | [82] | |
| Human small cell lung carcinoma (NCI-H187) | Inhibits the HSP 27 protein as a monomeric form of. Derivative of ZER induces strong cytotoxicity. | [83] | |
| Human non-small cell lung cancer (NSCLC) cells | The viability of NSCLC cells, significantly impaired by treatments of in a dose-dependent manner and NSCLC. | [84] | |
| Liver | ATCC-HTB-22 cells | Some novel compounds fabricated through azazerumbone conjugation with 2,4-dihydroxychalcones. These compounds showed anti-proliferative activity against the LU-1, Hep-G2, MCF-7 and SW480 human cancer cell lines. | [81] |
| HepG2 cells | Some novel compounds fabricated through azazerumbone conjugation with 2,4-dihydroxychalcones. These compounds showed anti-proliferative activity against the LU-1, Hep-G2, MCF-7 and SW480 human cancer cell lines. | [64] | |
| Murine hepatoma cells (Hepa1c1c7) | Increases proteasome activity, p62 and microtubule-associated protein 1 light-chain 3 (LC3)-II. | [85] | |
| Breast | ATCC-HTB-22 cells | The conjugation of azazerumbone and 2,4-dihydroxychalcones use for the preparation of novel target compounds. The anti-proliferative activity of these compounds against the LU-1, Hep-G2, MCF-7 and SW480 human cancer cell lines improves compared to azazerumbone or ZER. | [81] |
| MDA-MB-231, MCF-7, SUM159 cells | Exposure of cells to ZER resultes in increased cleavage of Notch2 in each cell line.Notch2 activation by ZER inhibits its proapoptotic and anti-migratory response. | [86] | |
| kinase κB (IKKβ) and the Nuclear factor κB (NF-κB) component proteins | Inhibits the IKKβ kinase that activates the NF-κB and also binds to the NF-κB complex in the TNF pathway. Blocking both proteins can lead to inhibition of cell proliferating proteins to be downregulated and possibly ultimate induction of apoptosis. | ||
| Human mammary gland adenocarcinoma (MDA-MB-231) cell line | Suppresses the proliferation of MDA-MB-231 cells. | [87] | |
| Hs578T and MDA-MB231 cells | IL-1β-induced IL-8 and MMP-3 expression, migration and invasion decrease. | ||
| MCF-7 and MDAMB-231 human cells | Induces significant expression of DR4. Activation of Bax and Bak and is not cytotoxic. | [88] | |
| Human mammary adenocarcinoma MDA-MB-231 Cell Line | ZER and ZER-NLC markedly suppressed the proliferation ofMDA-MB-231 cells. They arrested MDA-MB-231 cell cycle at the G2/M phase. | [89] | |
| SKBR3 breast cancer cells | ZER downregulated the level of CD44 expression in CD44+. The induction of CD44 expression by EGFR ligands, EGF or TGF-α, was significantly decreased by ZER treatment. | [90] | |
| Leukemia | MDA-MB-231Cell Line | Proliferation of MDA-MB significantly suppressed by ZER. | [88] |
| CML-K562 cells | Inhibits K562 cell proliferation and colony formation capability. | [91] | |
| WEHI-3B cells | The growth of leukemia cells inhibits. | [81] | |
| Human myeloid leukemia (HL60) | Drops off the percentage of HL60 cell viability. | [92] | |
| T-acute lymphoblastic leukemia, CEM-ss cells | Cytotoxic influence on CEM-ss cells and able to apoptosis the T-acute lymphoblastic leukemia. | [93] | |
| Human T-cell acute lymphoblastic leukemia (Jurkat) cells | Used as a system with sustained-release drug carrier mechanism. ZER activated the caspase-3 and caspase-9 and induced intrinsic apoptotic pathway, cytochrome c release from mitochondria, and PARP cleavage. | [94] | |
| Human peripheral blood lymphocytes (PBL) | The overall clastogenic effect not significant and is a cytotoxic but not a clastogenic substance in human PBL. | [36] | |
| Mice thymocytes and splenocytes human PBMC | Proliferation in stimulates time- and dose-dependent manner of human PBMC and mice cells upregulates human cytokine immunomodulatory. | [73] | |
| Human peripheral blood lymphocytes | At high concentrations induces an apparent substantial increase in the micronuclei frequency. | [95] | |
| Colon | Cells of mice thymocytes, mice splenocytes and human human peripheral blood mononuclear | ZEr activated the mice thymocytes, splenocytes and PBMC with dosage dependent manne. | [96] |
| Colorectal cancer cells, (CRC) cells | ZER enhanced radiation-induced cell cycle arrest (G2/M), increased radiation-induced apoptosis and enhanced radiation-induced DNA damage. | [97] | |
| Prostate | Hormone refractory prostate cancer (HRPC) cell lines | Induces antiproliferative and apoptotic influence on PC-3 and DU-145, 2 human hormonerefractory prostate cancer (HRPC) cell lines. | [81] |
| Brain | Human meningioma cell lines (IOMM-Lee, CH157MN) | Induces apoptosis with enhanced phosphorylation of glycogen synthase kinase 3 β (GSK3β) via inhibition of the Wnt5/β-catenin pathway. | |
| Human brain malignant glioma (GBM8401) | GBM8401 cells death induction with a dose-dependent pattern. | [98] | |
| Human brain malignant glioma (U87MG) | Transfection of GBM 8401 cells with WT IKKα inhibite ZER-induced apoptosis, and ZER markedly decreases IKKα phosphorylation levels with a time-dependent pattern. | [99] | |
| Kidney | Human RCC cell line 786-O | Suppresses STAT3 activation with a dose- and time-dependent pattern in RCC cells. | [100] |
| Normal African green monkey kidney cells | Nonsignificant cytotoxicity with 30 µM IC50. | [74] | |
| Ovarian | Normal Chinese hamster ovary cells (CHO) | Has genotoxic produces and cytotoxic influences in high concentrations. | [74] |
| Miscellaneous | human umbilical vein endothelial cells (HUVECs) | Inhibits HUVECs proliferation, migration and tubule formation. | [101] |
| Human oral cancer (KB) | ZER derivatives induces strong cytotoxicity. | [83] | |
| Gastric | Umbilical vein endothelial cells (HUVECs) | Proliferation of cell, VEGF expression and NF-κB activity in AGS cells inhibited by ZER. Reduction in both VEGF expression and NF-κB activity in AGS cells. | [21] |
| Human gastric adenocarcinoma (AGS) | Inhibits tumor angiogenesis via reduction of VEGF production and NF-κB activity. | [71] | |
| Skin | Murine epidermal cells (JB6 Cl41) | Murine epidermal cells (JB6 Cl41) Induces heme oxygenase-1 expression by activation of Nrf2. | [83] |
| Organ | Animal Model | ZER Route | Biological Effects of ZER | References |
|---|---|---|---|---|
| Lung | BALB/c female mice | Intraperitoneal injection | Effectively controls the growth of tumor and metastasis via delayed progression cancer cell cycle and apoptosis. | [39] |
| Kidney | Adult male Sprague Dawley rats | Injected intra articularly | Significantly induction in cytosolic glutathione-S-transferase enzyme activity. | [87] |
| Six-week-old athymic nu/nu female mice | Injection | STAT3 activation is inhibited in tissues of tumor and the human RCC xenograft tumors growth. | [101] | |
| Leukemia | Male BALB/c mice | Intraperitoneal injection | The growth of leukemia cells inhibits. | [97] |
| Chinese Hamster Ovary (CHO) cells and rat bone marrow polychromatic erythrocytes (PCEs) | Intraperitoneal injection | The leukemia cells number in the spleen of BALB/c leukemia mice markedly decreases after 28 days of orally treatment with different doses of ZER-NLC. Inhibits cell proliferation and causes cytotoxicity in the rat bone marrow. | [83] | |
| Miscellaneous | 28-Days-old C57BL/6 male mice | Intraperitoneal injection | Significant decreases in content of vascularization and hemoglobin in the plugs from ZER-treated mice, than control mice. | [71] |
| Syrian golden hamsters | Oral dose | Decreases hepatic mRNA levels of sterol regulatory element-binding protein-1c and its lipogenic target genes, included fatty acid synthase, acetyl-CoA carboxylase 1, and stearoyl-CoA desaturase 1. | [102] | |
| Male Sprague Dawley rats | Intraperitoneal injection | Increase with dose-dependent manner in MN production. No significant effect on human PBL by the overall clastogenic. | [97] | |
| Male Wistar rats | Oral dose | Decreases infiltration of macrophages, IL-1, IL-6, and TNF-α produced by p38 mitogen-activated protein kinase activation. | [103] | |
| Breast | Female severe combined immune deficient (SCID) mouse | Intraperitoneal injection | Retards growth of orthotopic MDA-MB-231 xenografts in association with induction in apoptosis and suppression of cell proliferation (Ki-67 expression). | [90] |
| 4T1 challenged mice | Oral feeding | ZER controlled the growth of tumor and metastasis by delaying the cancer cell cycle progression and apoptosis. | [39] | |
| Skin | Female HR-1 hairless mice | Topical application | increases of Nrf2 nuclear translocation followed by the promoter activity of HO-1, and also enhances Nrf2 direct binding to the antioxidant response element. | [104] |
| Liver | Male Sprague Dawley rats | Oral dose | Upregulates heat shock protein expressions in the liver Confers thermoresistant phenotype. | [85] |
| Male golden Syrian Hamsters | Oral dose | Improves dyslipidemia by modulating the genes expression involved in the lipolytic and lipogenic pathways of lipids metabolism Decreases hepatic mRNA levels of fatty acid synthase, malic enzyme, sterol-regulatory element binding protein, and 3-hydroxy-3-methyl-glutaryl-CoA reductase. | [21] | |
| Colon | Pathogen-free male Sprague–Dawley rats | Intraperitoneal injection | Lowers expression of PCNA is observed in the rat liver Increases Bax and decreases Bcl-2 protein expression in the liver. | [105] |
| Paw | Mice | Intraperitoneal injection | Significantly inhibited the production of paw edema induced by carrageenan in dose-dependent. | [102] |
| Eye | Female imprinting control region (ICR) mice | Oral dose | Inhibits the expressions of NF-κB, iNOS, and TNF-α. Abrogates nuclear translocation of NF-κB. | [106] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalantari, K.; Moniri, M.; Boroumand Moghaddam, A.; Abdul Rahim, R.; Bin Ariff, A.; Izadiyan, Z.; Mohamad, R. A Review of the Biomedical Applications of Zerumbone and the Techniques for Its Extraction from Ginger Rhizomes. Molecules 2017, 22, 1645. https://doi.org/10.3390/molecules22101645
Kalantari K, Moniri M, Boroumand Moghaddam A, Abdul Rahim R, Bin Ariff A, Izadiyan Z, Mohamad R. A Review of the Biomedical Applications of Zerumbone and the Techniques for Its Extraction from Ginger Rhizomes. Molecules. 2017; 22(10):1645. https://doi.org/10.3390/molecules22101645
Chicago/Turabian StyleKalantari, Katayoon, Mona Moniri, Amin Boroumand Moghaddam, Raha Abdul Rahim, Arbakariya Bin Ariff, Zahra Izadiyan, and Rosfarizan Mohamad. 2017. "A Review of the Biomedical Applications of Zerumbone and the Techniques for Its Extraction from Ginger Rhizomes" Molecules 22, no. 10: 1645. https://doi.org/10.3390/molecules22101645
APA StyleKalantari, K., Moniri, M., Boroumand Moghaddam, A., Abdul Rahim, R., Bin Ariff, A., Izadiyan, Z., & Mohamad, R. (2017). A Review of the Biomedical Applications of Zerumbone and the Techniques for Its Extraction from Ginger Rhizomes. Molecules, 22(10), 1645. https://doi.org/10.3390/molecules22101645