
What Characteristics Confer Proteins the Ability to Induce Allergic Responses? IgE Epitope Mapping and Comparison of the Structure of Soybean 2S Albumins and Ara h 2

Abstract
:1. Introduction
2. Results
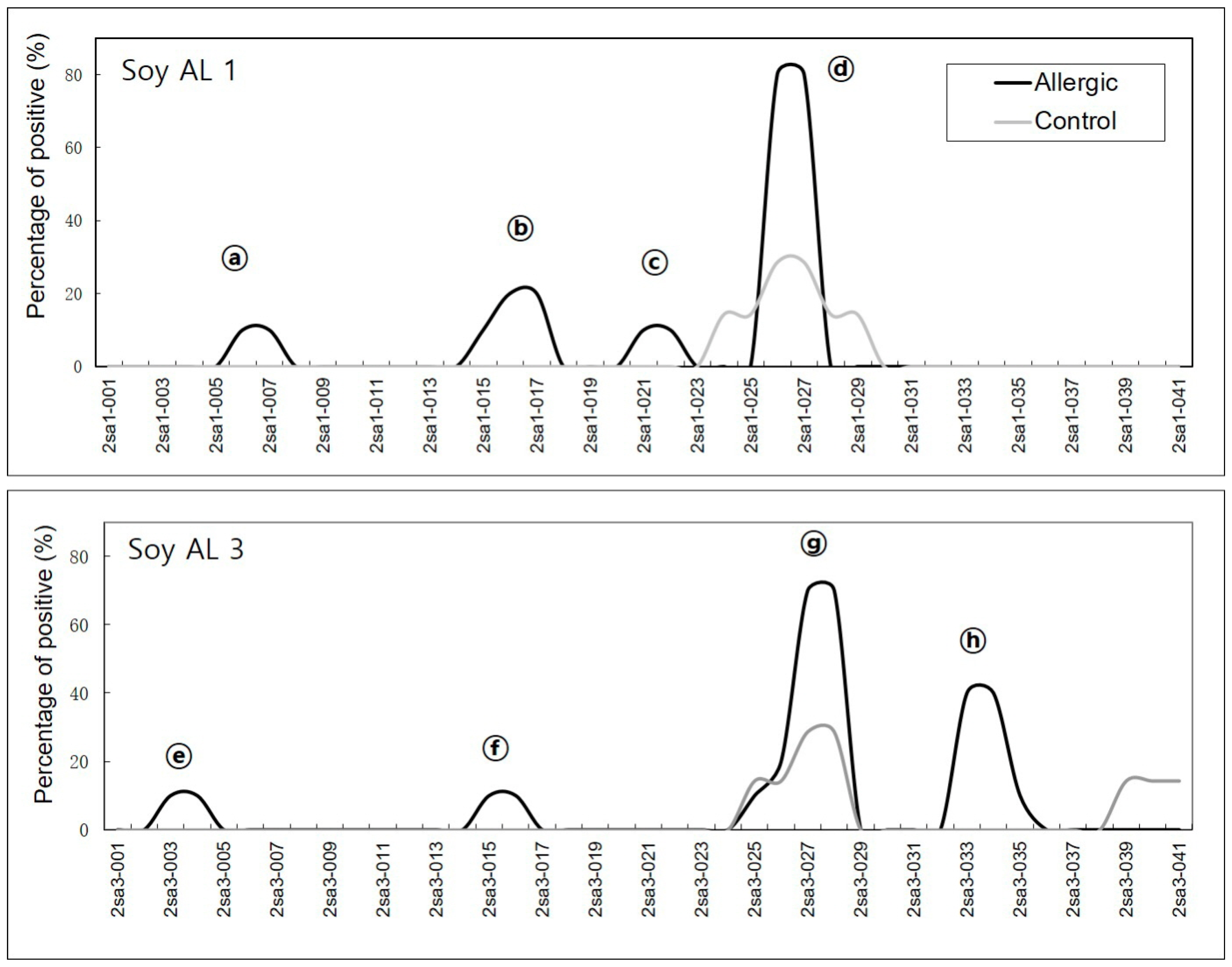
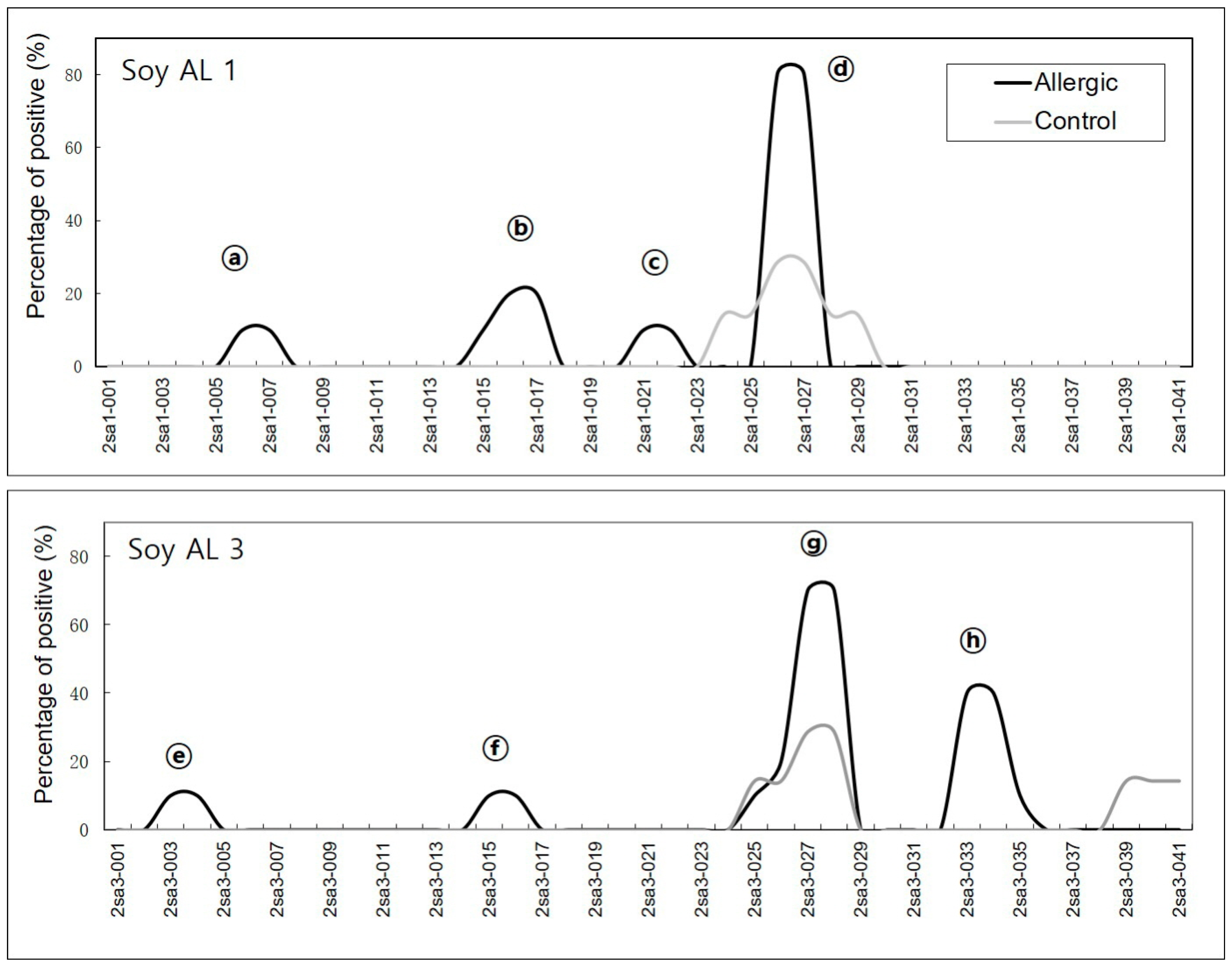
2.1. Detection of Sequential Epitopes
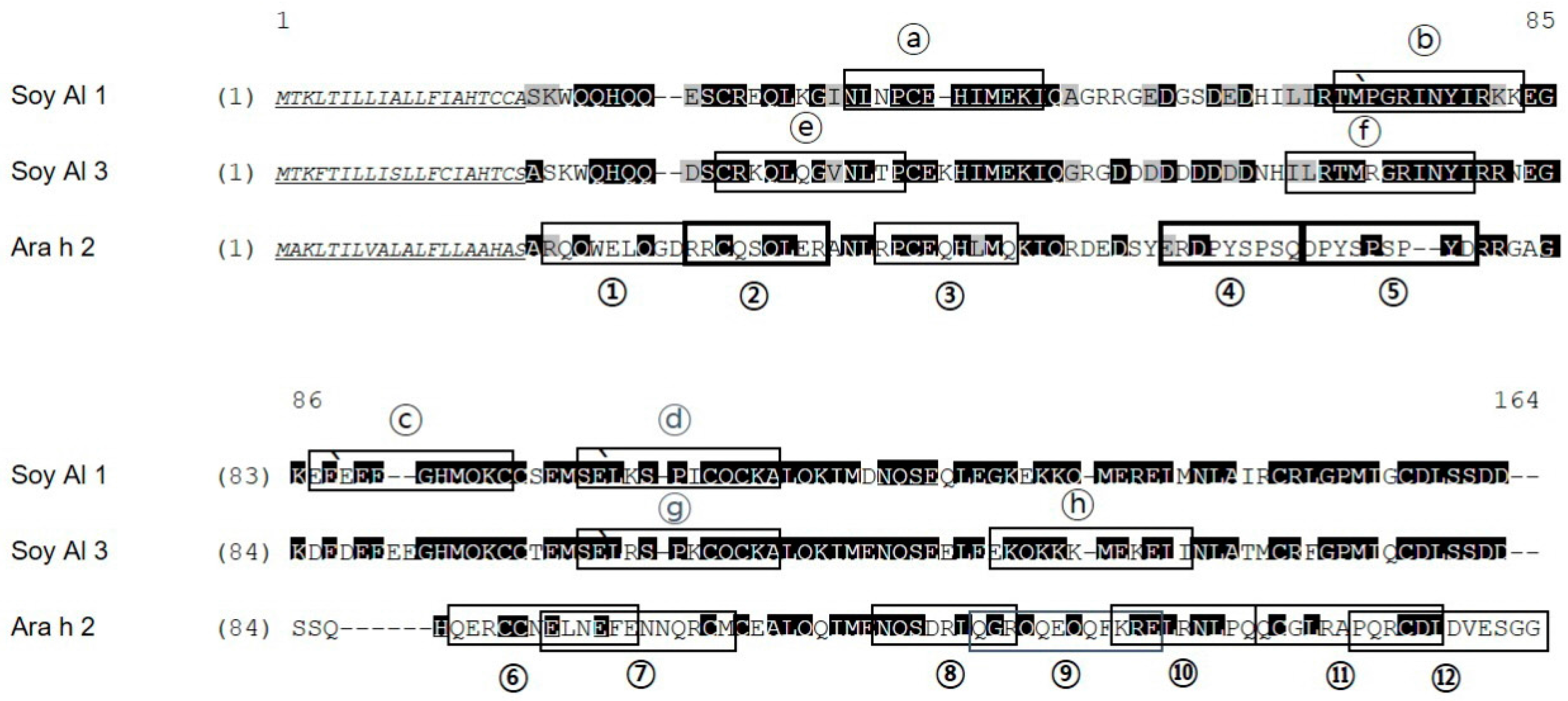
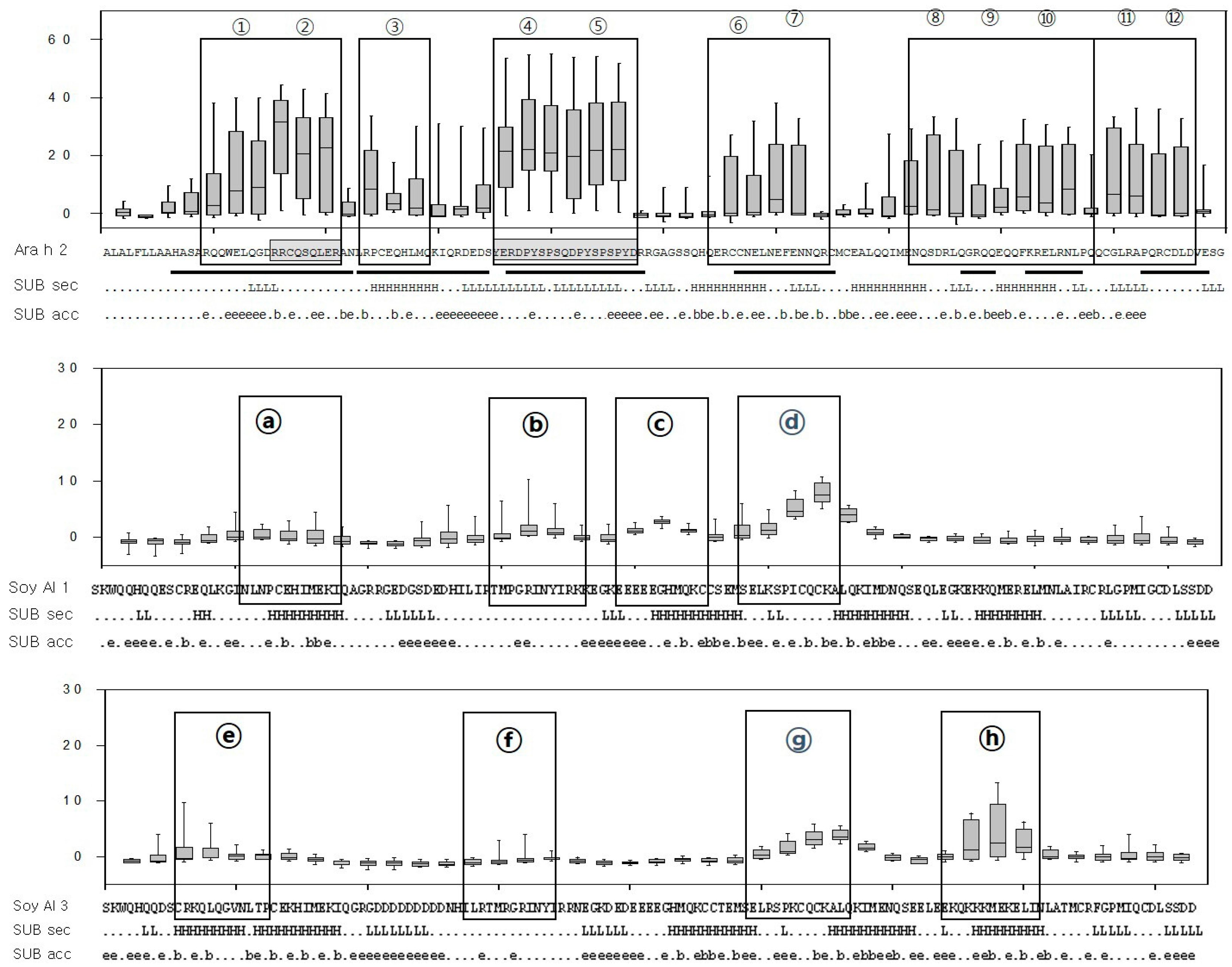
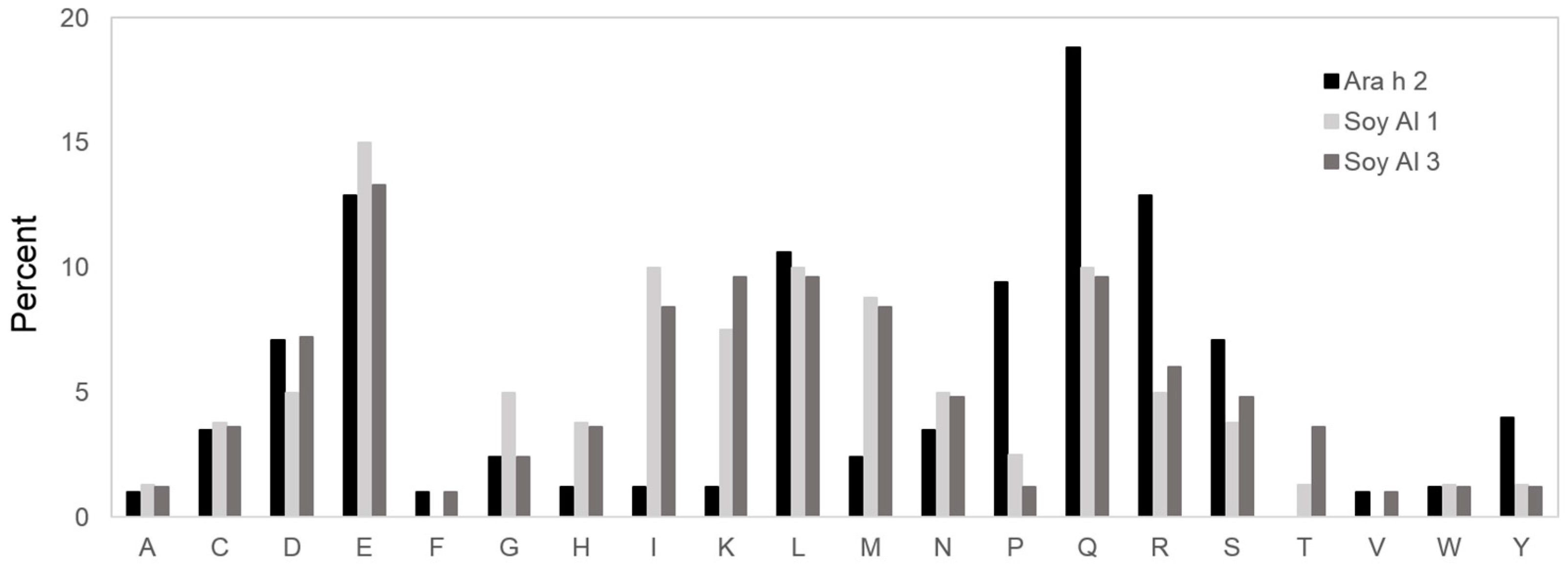
2.2. Structure Comparison
3. Discussion
4. Materials and Methods
4.1. Patient Sera
4.2. Microarray
4.3. Structure Comparison
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Soy Al 1 | soybean 2S albumin 1 |
| Soy Al 3 | soybean 2S albumin 3 |
References
- Bannon, G.A. What makes a food protein an allergen? Curr. Allergy Asthma Rep. 2004, 4, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Breiteneder, H.; Mills, E.N. Molecular properties of food allergens. J. Allergy Clin. Immunol. 2005, 115, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C. Structural biology of allergens. J. Allergy Clin. Immunol. 2000, 106, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Huby, R.D.; Dearman, R.J.; Kimber, I. Why are some proteins allergens? Toxicol. Sci. 2000, 55, 235–246. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Evaluation of Allergenicity of Genetically Modified Foods; Report of a Joint FAO/WHO Expert Consultation on Allergenicity of Foods Derived from Biotechnology; FAO/WHO: Rome, Italy, 2001; pp. 10–13. [Google Scholar]
- Breiteneder, H.; Radauer, C. A classification of plant food allergens. J. Allergy Clin. Immunol. 2004, 113, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, J.A.; Griffiths-Jones, S.; Shewry, P.R.; Breiteneder, H.; Mills, E.N. Structural relatedness of plant food allergens with specific reference to cross-reactive allergens: An in silico analysis. J. Allergy Clin. Immunol. 2005, 115, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Moreno, F.J.; Clemente, A. 2S albumin storage proteins: What makes them food allergens? Open Biochem. J. 2008, 2, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Bashir, M.E.; Hubatsch, I.; Leinenbach, H.P.; Zeppezauer, M.; Panzani, R.C.; Hussein, I.H. Ric c 1 and Ric c 3, the allergenic 2S albumin storage proteins of Ricinus communis, complete primary structures and phylogenetic relationships. Int. Arch. Allergy Immunol. 1998, 115, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Beyer, K.; Bardina, L.; Grishina, G.; Sampson, H.A. Identification of sesame seed allergens by 2-dimensional proteomics and Edman sequencing: Seed storage proteins as common food allergens. J. Allergy Clin. Immunol. 2002, 110, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Nordlee, J.A.; Taylor, S.L.; Townsend, J.A.; Thomas, L.A.; Bush, R.K. Identification of a Brazil-nut allergen in transgenic soybeans. N. Engl. J. Med. 1996, 334, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Puumalainen, T.J.; Poikonen, S.; Kotovuori, A.; Vaali, K.; Kalkkinen, N.; Reunala, T.; Turjanmaa, K.; Palosuo, T. Napins, 2S albumins, are major allergens in oilseed rape and turnip rape. J. Allergy Clin. Immunol. 2006, 117, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Robotham, J.M.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Linear IgE epitope mapping of the English walnut (Juglans regia) major food allergen, Jug r 1. J. Allergy Clin. Immunol. 2002, 109, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Robotham, J.M.; Wang, F.; Seamon, V.; Teuber, S.S.; Sathe, S.K.; Sampson, H.A.; Beyer, K.; Seavy, M.; Roux, K.H. Ana o 3, an important cashew nut (Anacardium occidentale L.) allergen of the 2S albumin family. J. Allergy Clin. Immunol. 2005, 115, 1284–1290. [Google Scholar] [CrossRef] [PubMed]
- Sathe, S.K.; Kshirsagar, H.H.; Roux, K.H. Advances in seed protein research: A prospective on seed allergens. J. Food Sci. 2005, 70, R93–R120. [Google Scholar] [CrossRef]
- Kattan, J.D.; Sampson, H.A. Clinical reactivity to soy is best identified by component testing to Gly m 8. J. Allergy Clin. Immunol. Pract. 2015, 3, 970–972. [Google Scholar] [CrossRef] [PubMed]
- Ebisawa, M.; Brostedt, P.; Sjölander, S.; Sato, S.; Borres, M.P.; Ito, K. Gly m 2S albumin is a major allergen with a high diagnostic value in soybean-allergic children. J. Allergy Clin. Immunol. 2013, 132, 976–978. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, K.; Schweimer, K.; Reese, G.; Randow, S.; Suhr, M.; Becker, W.M.; Vieths, S.; Rösch, P. Structure and stability of 2S albumin-type peanut allergens: Implications for the severity of peanut allergic reactions. Biochem. J. 2006, 395, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Shewry, P.R.; Archer, D.B.; Beyer, K.; Niggemann, B.; Haas, H.; Wilson, P.; Alcocer, M.J.C. The potential allergenicity of two 2S albumins from soybean (Glycine max): A protein microarray approach. Int. Arch. Allergy Immunol. 2006, 141, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Shreffler, W.G.; Beyer, K.; Chu, T.H.; Burks, A.W.; Sampson, H.A. Microarray immunoassay: Association of clinical history, in vitro IgE function, and heterogeneity of allergenic peanut epitopes. J. Allergy Clin. Immunol. 2004, 113, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Bardina, L.; Shreffler, W.G.; Andreae, D.A.; Ge, Y.; Wang, J.; Bruni, F.M.; Fu, Z.; Han, Y.; Sampson, H.A. Development of a novel peptide microarray for large-scale epitope mapping of food allergens. J. Allergy Clin. Immunol. 2009, 124, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Stanley, J.S.; King, N.; Burks, A.W.; Huang, S.K.; Sampson, H.; Cockrell, G.; Helm, R.M.; West, C.M.; Bannon, G.A. Identification and mutational analysis of the immunodominant IgE binding epitopes of the major peanut allergen Ara h 2. Arch. Biochem. Biophys. 1997, 342, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Pascal, M.; Konstantinou, G.N.; Masilamani, M.; Lieberman, J.; Sampson, H.A. In silico prediction of Ara h 2 T cell epitopes in peanut-allergic children. Clin. Exp. Allergy. 2013, 43, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Otsu, K.; Guo, R.; Dreskin, S.C. Epitope analysis of Ara h 2 and Ara h 6: Characteristic patterns of IgE-binding fingerprints among individuals with similar clinical histories. Clin. Exp. Allergy 2015, 45, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, D.C.; Berzofsky, J.A.; East, I.J.; Gurd, F.R.N.; Hannum, C.; Leach, S.J.; Margoliash, E.; Michael, J.G.; Miller, A.; Prager, E.M.; et al. The antigenic structure of proteins: A reappraisal. Annu. Rev. Immunol. 1984, 2, 67–101. [Google Scholar] [CrossRef] [PubMed]
- Hopp, T.P. Protein surface analysis. Methods for identifying antigenic determinants and other interaction sites. J. Immunol. Methods 1986, 88, 1–18. [Google Scholar] [CrossRef]
- Berzofsky, J.A. Intrinsic and extrinsic factors in protein antigenic structure. Science 1985, 229, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, N.D.; Mayrose, I.; Halperin, D.; Yekutieli, D.; Gershoni, J.M.; Pupko, T. Computational characterization of B-cell epitopes. Mol. Immunol. 2007, 45, 3477–3489. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gordo, M.; Lin, J.; Bardina, L.; Pastor-Vargas, C.; Cases, B.; Vivanco, F.; Cuesta-Herranz, J.; Sampson, H.A. Epitope mapping of Atlantic salmon major allergen by peptide microarray immunoassay. Int. Arch. Allergy Immunol. 2012, 157, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Novotny, J.; Handschumacher, M.; Haber, E.; Bruccoleri, R.E.; Carlson, W.B.; Fanning, D.W.; Smith, J.A.; Rose, G.D. Antigenic determinants in proteins coincide with surface regions accessible to large probes (antibody domains). Proc. Natl. Acad. Sci. USA 1986, 83, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Alcocer, M.J.; Murtagh, G.J.; Wilson, P.B.; Progias, P.; Lin, J.; Archer, D.B. The major human structural IgE epitope of the Brazil nut allergen Ber e 1: A chimaeric and protein microarray approach. J. Mol. Biol. 2004, 343, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Chruszcz, M.; Maleki, S.J.; Majorek, K.A.; Demas, M.; Bublin, M.; Solberg, R.; Hurlburt, B.K.; Ruan, S.; Mattisohn, C.P.; Breiteneder, H.; et al. Structural and immunologic characterization of Ara h 1, a major peanut allergen. J. Biol. Chem. 2011, 286, 39318–39327. [Google Scholar] [CrossRef] [PubMed]
- Barre, A.; Borges, J.P.; Culerrier, R.; Rouge, P. Homology modelling of the major peanut allergen Ara h 2 and surface mapping of IgE-binding epitopes. Immunol. Lett. 2005, 100, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Berchtold, M.W.; Brinkmeier, H.; Müntener, M. Calcium ion in skeletal muscle: Its crucial role for muscle function, plasticity, and disease. Physiol. Rev. 2000, 80, 1215–1265. [Google Scholar] [PubMed]
- Tsai, C.J.; Lin, S.L.; Wolfson, H.J.; Nussinov, R. Studies of protein-protein interfaces: A statistical analysis of the hydrophobic effect. Protein Sci. 1997, 6, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Bogan, A.A.; Thorn, K.S. Anatomy of hot spots in protein interfaces. J. Mol. Biol. 1998, 280, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kühne, Y.; Reese, G.; Ballmer-Weber, B.K.; Niggemann, B.; Hanschmann, K.M.; Vueths, S.; Holzhauser, T. A novel multipeptide microarray for the specific and sensitive mapping of linear IgE-binding epitopes of food allergens. Int. Arch. Allergy Immunol. 2015, 166, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Herian, A.M.; Taylor, S.L.; Bush, R.K. Identification of soybean allergens by immunoblotting with sera from soy-allergic adults. Int. Arch. Allergy Appl. Immunol. 1990, 92, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Zapatero, L.; Caravaca, F.; Carreira, J. Identification of soybean proteins responsible for respiratory allergies. Int. Arch. Allergy Appl. Immunol. 1991, 95, 53–57. [Google Scholar] [PubMed]
- Ogawa, T.; Bando, N.; Tsuji, H.; Okajima, H.; Nishikawa, K.; Sasaoka, K. Investigation of the IgE-binding proteins in soybeans by immunoblotting with the sera of the soybean-sensitive patients with atopic-dermatitis. J. Nutr. Sci. Vitaminol. 1991, 37, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Calamelli, E.; Caffarelli, C.; Ricci, G. Peanut sensitization profiles in Italian children and adolescents with specific IgE to peanuts. BioMed Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- Dearman, R.; Alcocer, M.; Kimber, I. Influence of plant lipids on immune responses in mice to the major Brazil nut allergen Ber e 1. Clin. Exp. Allergy 2007, 37, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A. Utility of food-specific IgE concentrations in predicting symptomatic food allergy. J. Allergy Clin. Immunol. 2001, 107, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Bardina, L.; Shreffler, W.G. Microarrayed allergen molecules for diagnostics of allergy. Methods Mol. Biol. 2009, 524, 259–272. [Google Scholar] [PubMed]
- Lin, J.; Bruni, F.M.; Fu, Z.; Maloney, J.; Bardina, L.; Boner, A.L.; Gimenez, G.; Sampson, H.A. A bioinformatics approach to identify patients with symptomatic peanut allergy using peptide microarray immunoassay. J. Allergy Clin. Immunol. 2012, 129, 1321.e5–1328.e5. [Google Scholar] [CrossRef] [PubMed]
- Flinterman, A.E.; Knol, E.F.; Lencer, D.A.; Bardina, L.; den Hartog Jager, C.F.; Lin, J.; Pasmans, S.G.M.A.; Bruijnzeel-Koomen, C.A.F.M.; Sampson, H.A.; van Hoffen, E.; et al. Peanut epitopes for IgE and IgG4 in peanut-sensitized children in relation to severity of peanut allergy. J. Allergy Clin. Immunol. 2008, 121, 737.e10–743.e10. [Google Scholar] [CrossRef] [PubMed]
- Rost, B.; Yachdav, G.; Liu, J. PredictProtein. Nucleic Acids Res. 2004, 32, W321–W326. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modeling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L. SWISS-MODEL: Modeling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Serum samples were kindly supplied by the Food Allergy Resource Initiative, which is supported by funding from Food Allergy Research and Education and maintained at the Elliot and Roslyn Jaffe Food Allergy Institute, Icahn School of Medicine at Mount Sinai, NY, USA.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Age (year) | Challenge Type | Soybean Specific IgE (kUA/L) | Peanut Specific IgE (kUA/L) | Other Atopy |
|---|---|---|---|---|---|
| 1 | 4 | Open | 12.7 | 100 | None |
| 2 | 4 | Single Blind | 5.31 | 33.3 | None |
| 3 | 4 | Open | 77.7 | 100 | AD, asthma |
| 4 | 4 | Double Blind | 16.1 | 57.3 | AD, asthma |
| 5 | 4 | Double Blind | 77.4 | 100 | AD, asthma |
| 6 | 5 | Open | 0.53 | 17.1 | AD, asthma |
| 7 | 6 | Double Blind | 16.9 | 100 | AD, asthma |
| 8 | 7 | Double Blind | 35.3 | 100 | AD, AR, asthma |
| 9 | 9 | Double Blind | 6.64 | 15.4 | AR, asthma |
| 10 | 9 | Double Blind | 19.6 | 100 | AD, AR, asthma |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.; Lin, J.; Bardina, L.; Grishina, G.A.; Lee, C.; Seo, W.H.; Sampson, H.A. What Characteristics Confer Proteins the Ability to Induce Allergic Responses? IgE Epitope Mapping and Comparison of the Structure of Soybean 2S Albumins and Ara h 2. Molecules 2016, 21, 622. https://doi.org/10.3390/molecules21050622
Han Y, Lin J, Bardina L, Grishina GA, Lee C, Seo WH, Sampson HA. What Characteristics Confer Proteins the Ability to Induce Allergic Responses? IgE Epitope Mapping and Comparison of the Structure of Soybean 2S Albumins and Ara h 2. Molecules. 2016; 21(5):622. https://doi.org/10.3390/molecules21050622
Chicago/Turabian StyleHan, Youngshin, Jing Lin, Ludmilla Bardina, Galina A. Grishina, Chaeyoon Lee, Won Hee Seo, and Hugh A. Sampson. 2016. "What Characteristics Confer Proteins the Ability to Induce Allergic Responses? IgE Epitope Mapping and Comparison of the Structure of Soybean 2S Albumins and Ara h 2" Molecules 21, no. 5: 622. https://doi.org/10.3390/molecules21050622
APA StyleHan, Y., Lin, J., Bardina, L., Grishina, G. A., Lee, C., Seo, W. H., & Sampson, H. A. (2016). What Characteristics Confer Proteins the Ability to Induce Allergic Responses? IgE Epitope Mapping and Comparison of the Structure of Soybean 2S Albumins and Ara h 2. Molecules, 21(5), 622. https://doi.org/10.3390/molecules21050622