
Phenolic Constituents of Medicinal Plants with Activity against Trypanosoma brucei

 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
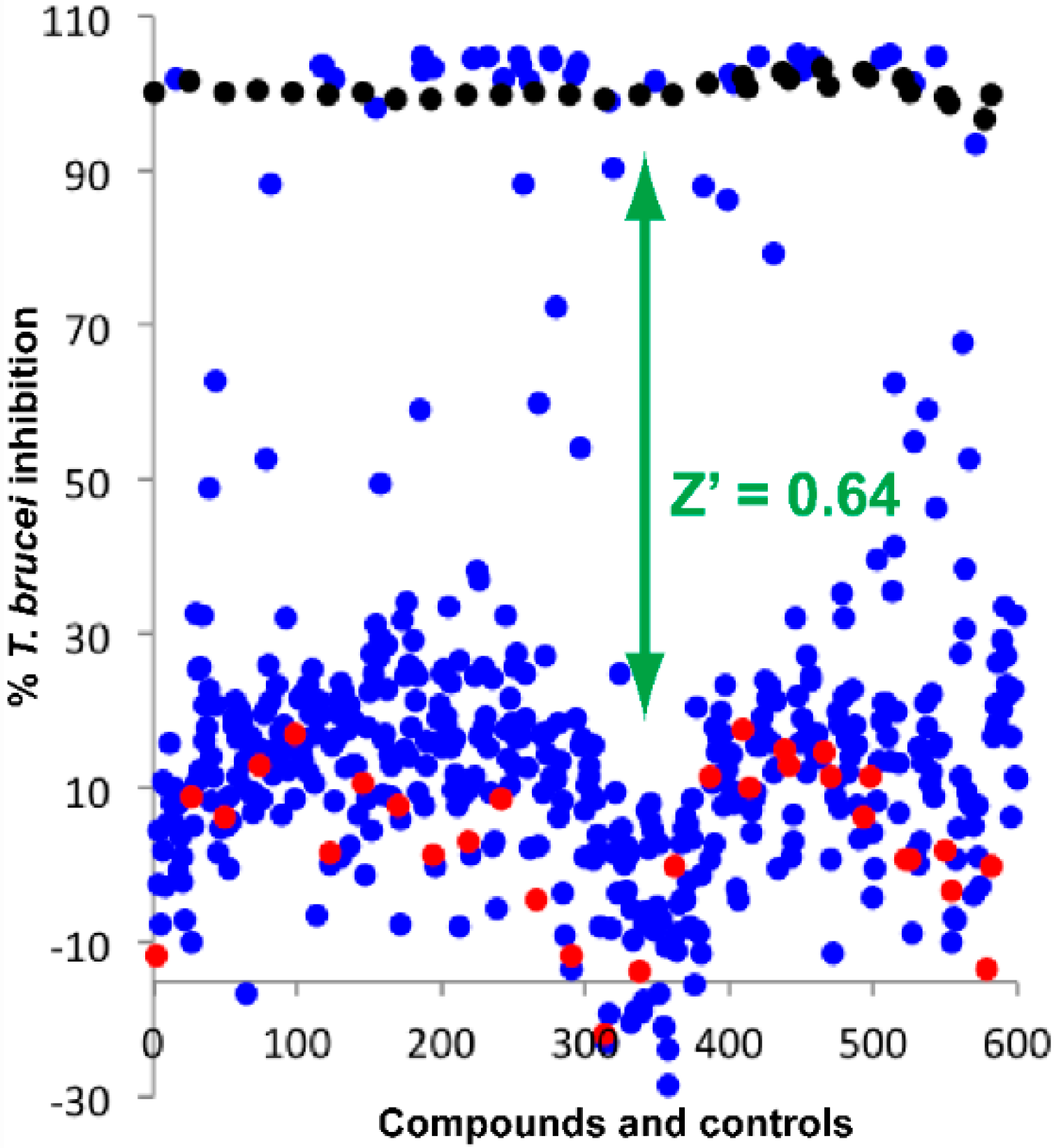
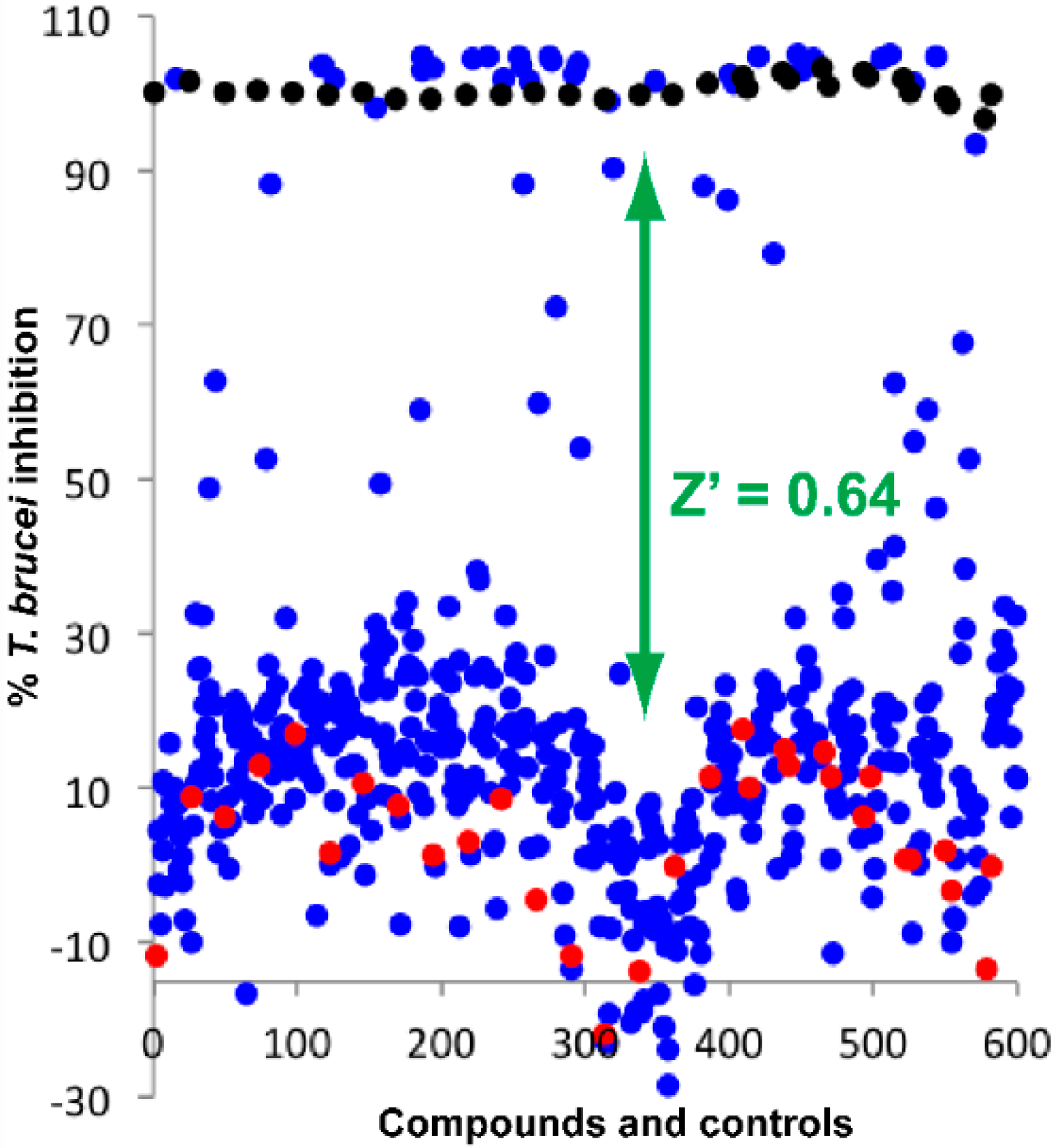
2.1. Screening of Natural Product Library
2.2. Biological Activity of the Screening Compounds
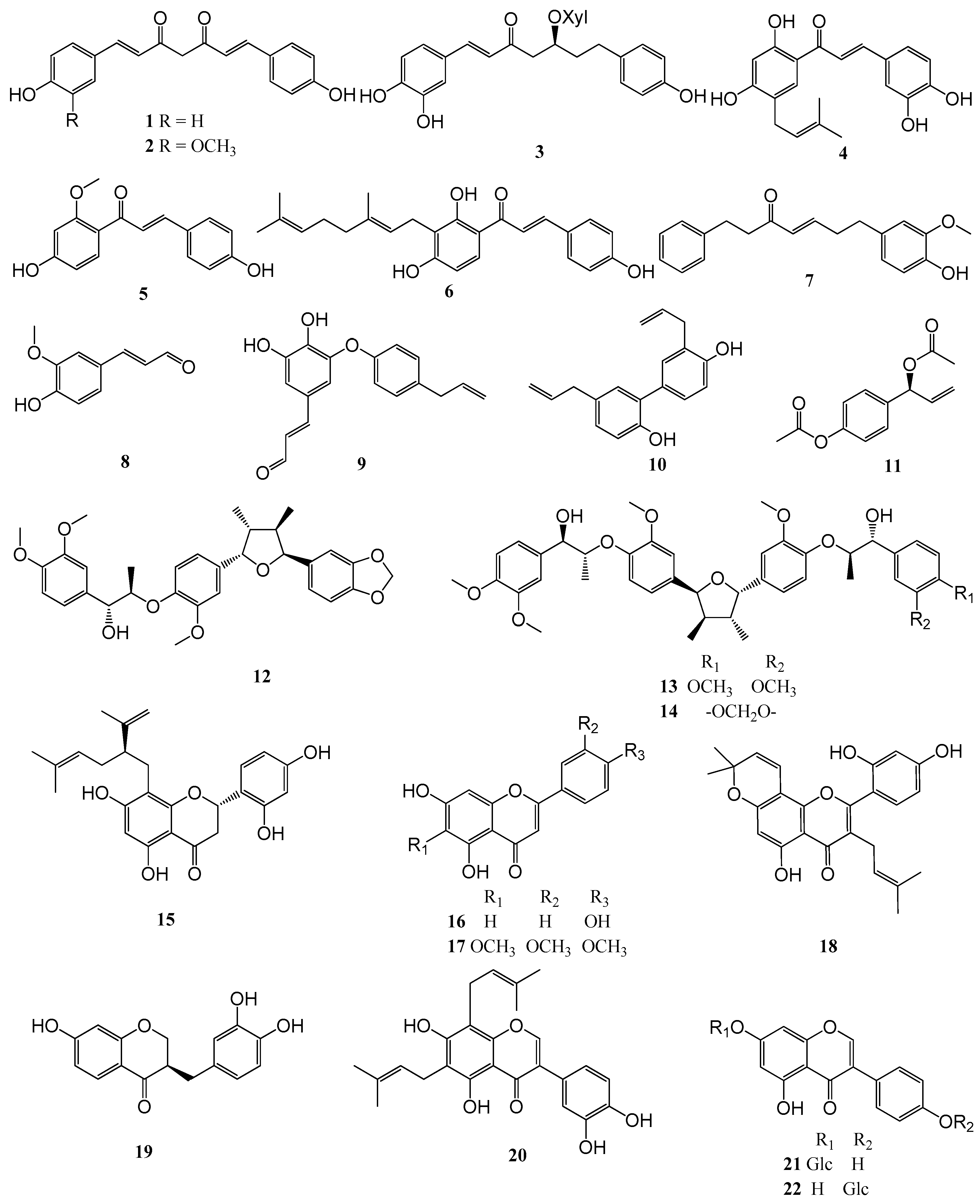
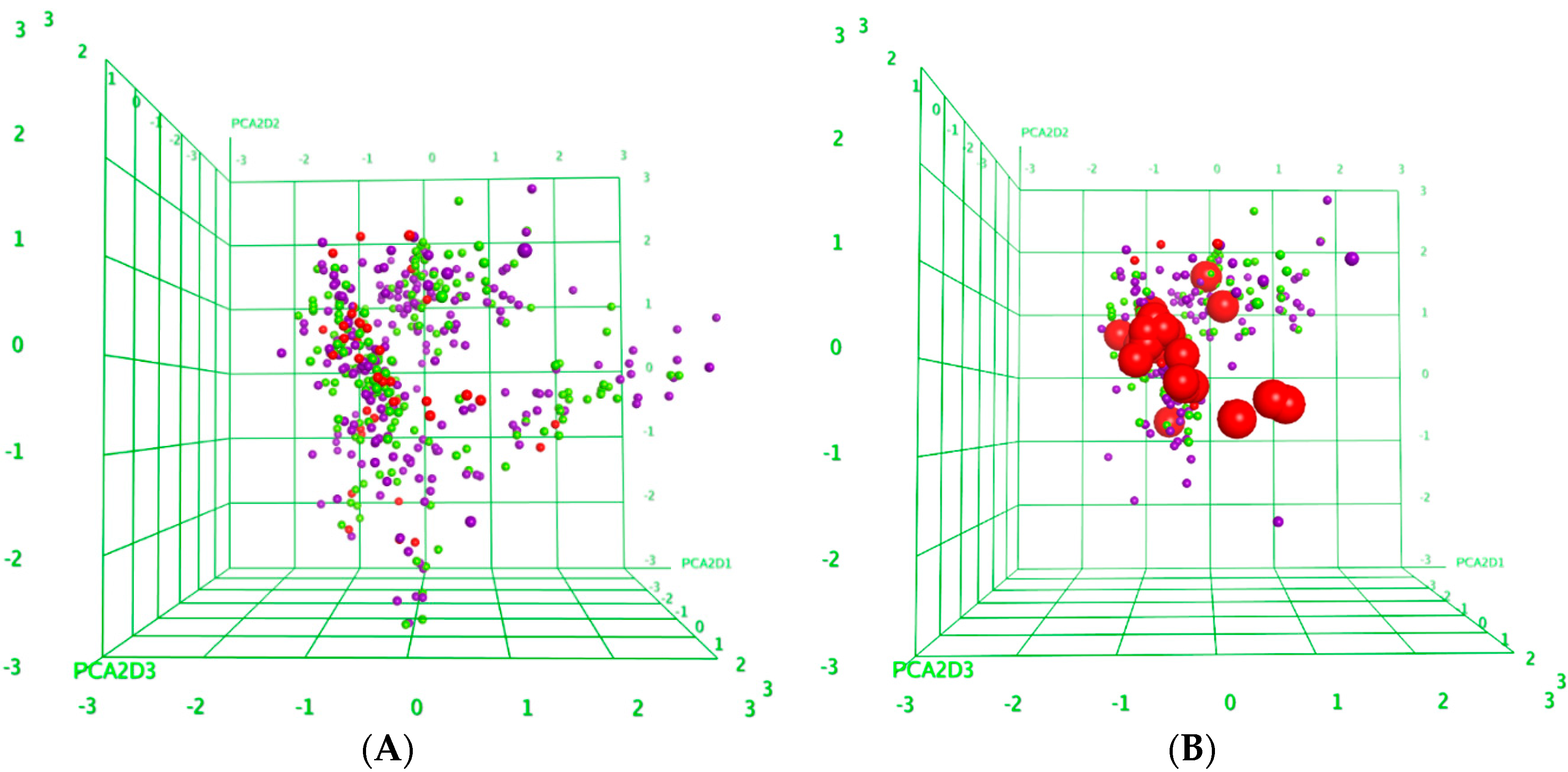
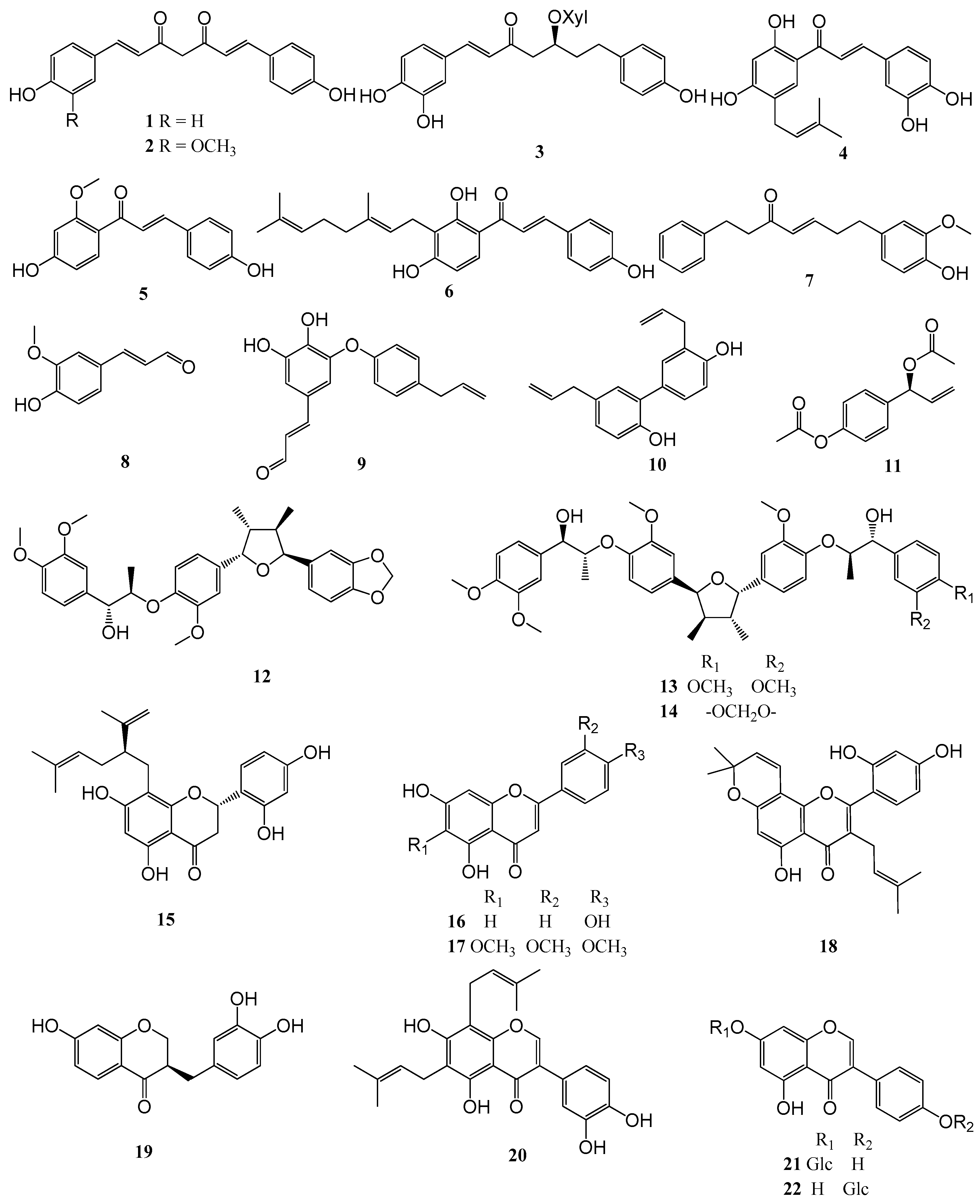
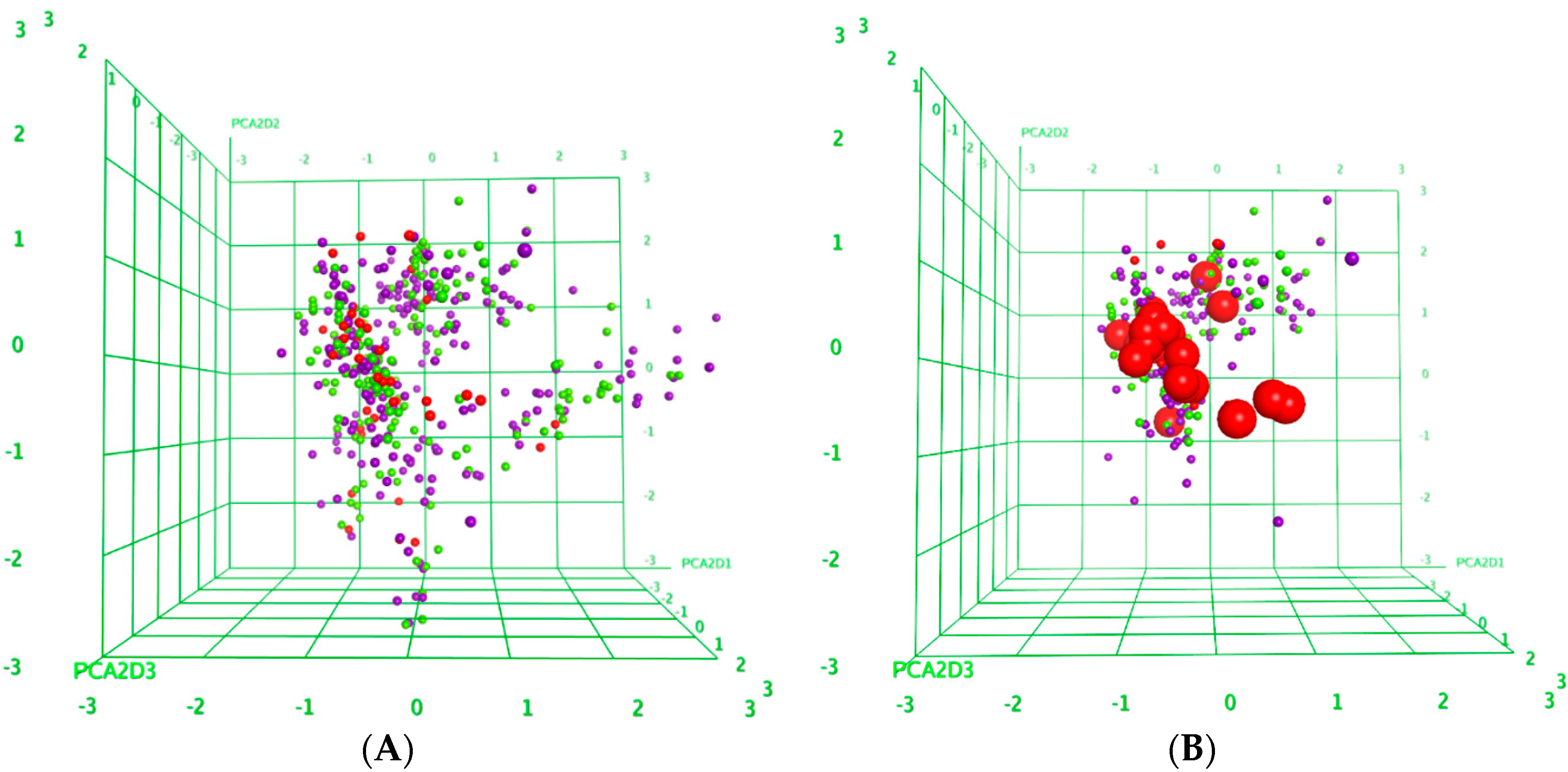
2.3. Structure-Activity Relationships
3. Materials and Methods
3.1. Test Compounds
3.2. Chemicals, Reagents, and Media
3.3. Anti-Trypanosomal Activity on T. brucei
3.4. Cytotoxicity Assays
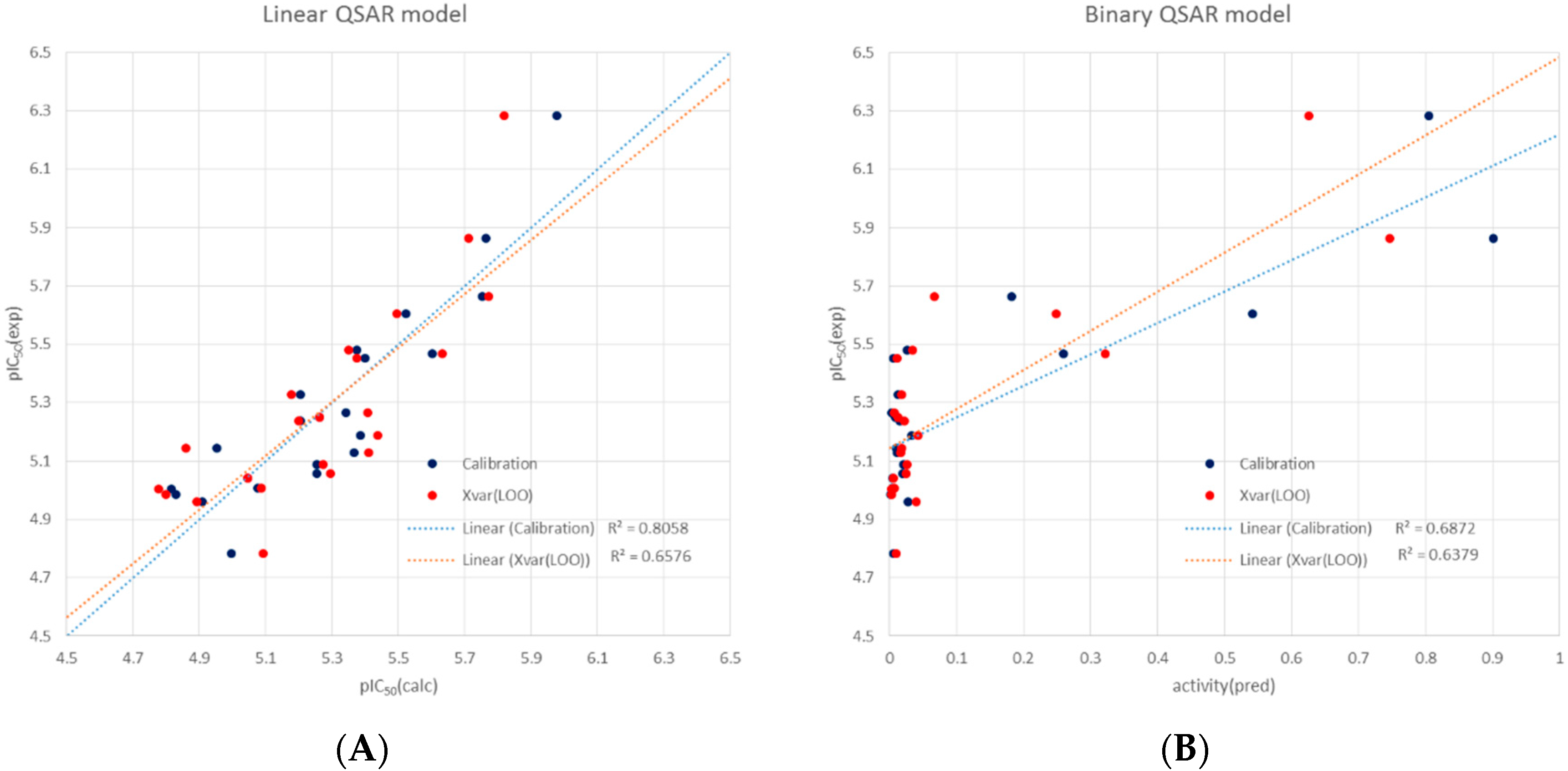
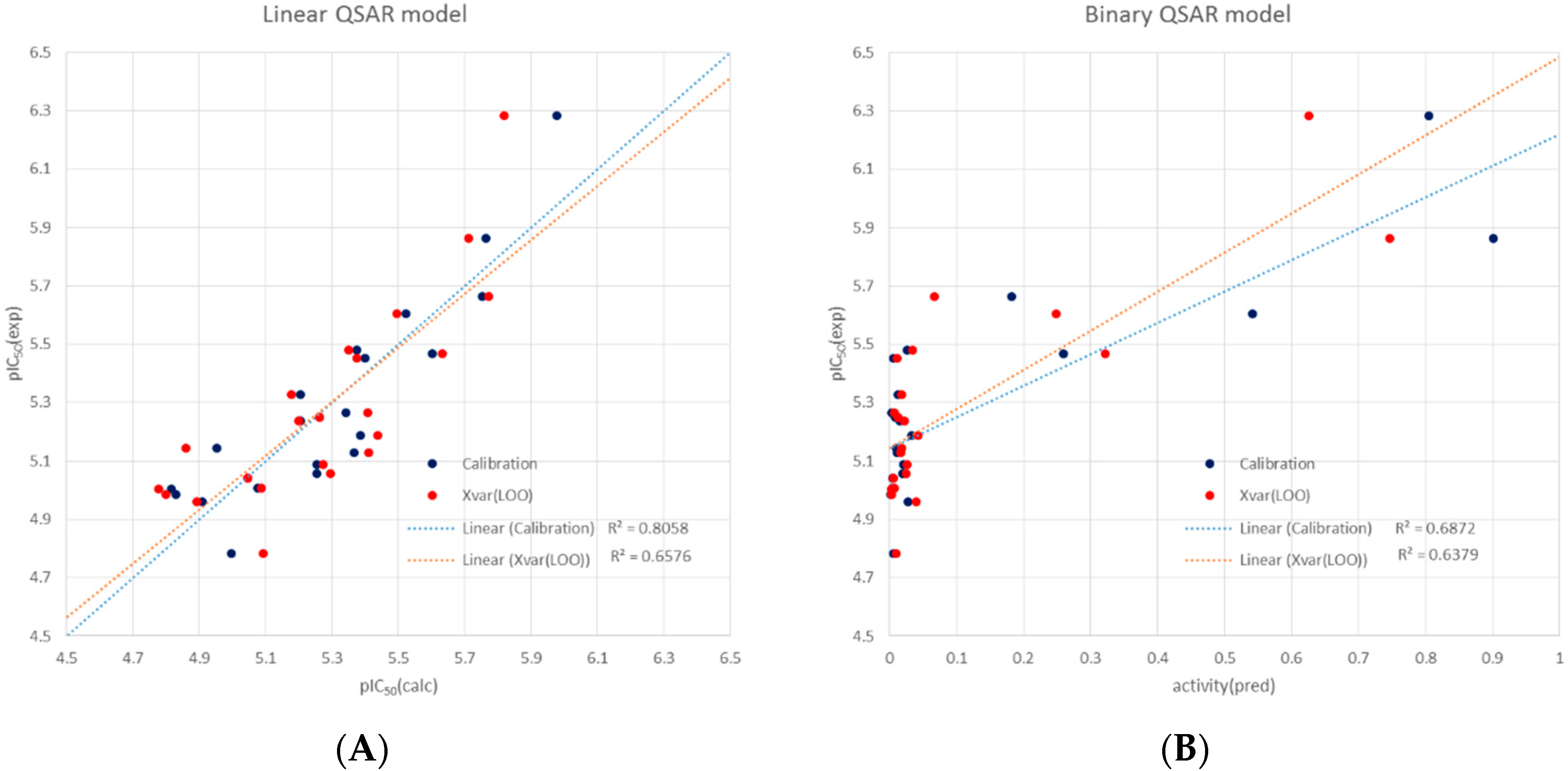
3.5. Molecular Modelling and QSAR analyses
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Barrett, M.P. Problems for the chemotherapy of human African trypanosomiasis. Curr. Opin. Infect. Dis. 2000, 13, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Olliario, P.; Lazdins, J.; Guhl, F. Developments in the treatment of leishmaniasis and trypanosomiasis. Expert Opin. Emerg. Drugs 2002, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hannaert, V. Sleeping sickness pathogen (Trypanosoma brucei) and natural products: Therapeutic targets and screening systems. Planta Med. 2011, 77, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Kristjanson, P.M.; Swallow, B.M.; Rowlands, G.J.; Kruska, R.L.; De Leeuw, P.N. Measuring the costs of African animal trypanosomosis, the potential benefits of control and returns to research. Agric. Syst. 1999, 59, 79–98. [Google Scholar] [CrossRef]
- Welburn, S.C.; Fèvre, E.M.; Coleman, P.G.; Odiit, M.; Maudlin, I. Sleeping sickness: A tale of two diseases. Trends Parasitol. 2001, 17, 19–24. [Google Scholar] [CrossRef]
- Simarro, P.P.; Cecchi, G.; Franco, J.R.; Paone, M.; Diarra, A.; Ruiz-Postigo, J.A.; Fèvre, E.M.; Mattioli, R.C.; Jannin, J.G. Estimating and mapping the population at risk of sleeping sickness. PLoS Negl. Trop. Dis. 2012, 6, e1859. [Google Scholar] [CrossRef] [PubMed]
- WHO. Control and Surveillance of African Trypanosomiasis Report of a WHO Expert Committee. WHO Tech. Rep. Ser. 2013, 984, 1–250. [Google Scholar]
- Kennedy, P.G.E. The continuing problem of human African trypanosomiasis (sleeping sickness). Ann. Neurol. 2008, 64, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Barrett, M.P. Potential new drugs for human African trypanosomiasis: Some progress at last. Curr. Opin. Infect. Dis. 2010, 23, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Ferrins, L.; Rahmani, R.S.; and Baell, J.B. Drug discovery and human African trypanosomiasis: A disease less neglected. Future Med. Chem. 2013, 5, 1801–1841. [Google Scholar] [CrossRef] [PubMed]
- Docampo, R.; Moreno, S.N. Current chemotherapy of human African trypanosomiasis. Parasitol. Res. 2003, 90, S10–S13. [Google Scholar] [PubMed]
- Pepin, J.; Milord, F. The treatment of human African trypanosomiasis. Adv. Parasitol. 1994, 33, 1–47. [Google Scholar] [PubMed]
- Schmidt, T.J.; Khalid, S.A.; Romanha, A.J.; Alves, T.M.A.; Biavatti, M.W.; Brun, R.; da Costa, F.B.; de Castro, S.L.; Ferreira, V.F.; de Lacerda, M.V.G.; et al. The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases—Part I. Curr. Med. Chem. 2012, 19, 2128–2175. [Google Scholar] [PubMed]
- Schmidt, T.J.; Khalid, S.A.; Romanha, A.J.; Alves, T.M.A.; Biavatti, M.W.; Brun, R.; Da Costa, F.B.; de Castro, S.L.; Ferreira, V.F.; de Lacerda, M.V.G.; et al. The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases—Part II. Curr. Med. Chem. 2012, 19, 2176–2228. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, D.; Kaiser, M.; Brun, R.; Yardley, V.; Schmidt, T.J.; Tosun, F.; Rüedi, P. Antitrypanosomal and antileishmanial activities of flavonoids and their analogues: In Vitro, in Vivo, structure-activity relationship, and quantitative structure-activity relationship studies. Antimicrob. Agents Chemother. 2006, 50, 1352–1364. [Google Scholar] [CrossRef] [PubMed]
- Changtam, C.; De Koning, H.P.; Ibrahim, H.; Sajid, M.S.; Gould, M.K.; Suksamrarn, A. Curcuminoid analogs with potent activity against Trypanosoma and Leishmania species. Eur. J. Med. Chem. 2010, 45, 941–956. [Google Scholar] [CrossRef] [PubMed]
- MOE—Molecular Operating Environment v. 2014.0901. For details on molecular descriptors see: http://www.chemcomp.com/journal/descr.htm. Available from Chemical Computing Group, Montreal, Canada: http://www.chemcomp.com/journal/descr.htm.
- Schmidt, T.J.; Heilmann, J. Quantitative Structure-Cytotoxicity Relationships of Sesquiterpene Lactones derived from Partial Charge (Q)-based Fractional Accessible Surface Area Descriptors (Q_frASAs). Quant. Struct. Act. Relat. 2002, 21, 276–287. [Google Scholar] [CrossRef]
- Cruciani, G.; Crivori, P.; Carrupt, P.-A.; Testa, B. Molecular fields in quantitative structure-permeation relationships: The VolSurf approach. J. Mol. Struct. 2000, 503, 17–30. [Google Scholar] [CrossRef]
- Thao, N.P.; No, J.H.; Luyen, B.T.T.; Yang, G.; Byun, S.Y.; Goo, J.; Kim, K.T.; Cuong, N.X.; Nam, N.H.; Minh, C.V.; Schmidt, T.J.; Kang, J.S.; Kim, Y.H. Secondary metabolites from vietnamese marine invertebrates with activity against Trypanosoma brucei and T. cruzi. Molecules 2014, 19, 7869–7880. [Google Scholar] [CrossRef] [PubMed]
- Thao, N.P.; Luyen, B.T.T.; Brun, R.; Kaiser, M.; Kiem, P.V.; Minh, C.V.; Schmidt, T.J.; Kang, J.S.; Kim, Y.H. Anti-protozoal activities of cembrane-type diterpenes from Vietnamese soft Corals. Molecules 2015, 20, 12459–12468. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds in this paper are available from the authors.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | T. brucei IC50 a (µM) | Cytotoxicity a CC50 Values (µM) | s.i b | Species | ||
|---|---|---|---|---|---|---|
| HEK239T | HepG2 | HEK239T | HepG2 | |||
| 1 | 9.84 ± 0.84 | >32.47 | >32.47 | >3.30 | >3.30 | Curcuma longa |
| 2 | 7.19 ± 1.02 | >21.35 | >29.55 | >2.97 | >4.11 | Curcuma longa |
| 3 | 9.12 ± 0.63 | >20.88 | >20.88 | >2.29 | >2.29 | Alnus japonica |
| 4 | 2.17 ± 0.50 | >28.81 | >29.19 | >13.28 | >13.54 | Broussonetia papyrifera |
| 5 | 10.37 ± 3.36 | >30.18 | >37.12 | >2.91 | >3.58 | Caesalpinia sappan |
| 6 | 7.42 ± 1.12 | >25.45 | >25.45 | >3.43 | >3.43 | Angelica keiskei |
| 7 | 2.48 ± 0.02 | >32.36 | >32.36 | >13.05 | >13.05 | Alpinia officinarum |
| 8 | 16.42 ± 2.78 | >48.11 | >56.97 | >2.93 | >3.47 | Cinnamomum cassia |
| 9 | 9.89 ± 0.51 | >33.82 | >33.82 | >3.42 | >3.42 | Machilus thunbergii |
| 10 | 8.76 ± 0.38 | >37.58 | >37.58 | >4.29 | >4.29 | Machilus thunbergii |
| 11 | 4.70 ± 1.53 | >23.22 | >29.99 | >4.94 | >6.38 | Alpinia galanga |
| 12 | 5.61 ± 0.09 | >16.21 | >18.63 | >2.89 | >3.32 | Saururus chinensis |
| 13 | 5.43 ± 0.26 | >11.84 | >13.68 | >2.18 | >2.52 | Saururus chinensis |
| 14 | 3.54 ± 0.49 | >11.04 | >13.91 | >3.12 | >3.93 | Saururus chinensis |
| 15 | 1.37 ± 0.01 | >23.76 | >23.76 | >17.34 | >17.34 | Sophora flavescens |
| 16 | 10.96 ± 2.64 | >37.04 | >37.04 | >3.38 | >3.38 | Agrimonia pilosa |
| 17 | 8.14 ± 1.37 | >29.14 | >29.14 | >3.58 | >3.58 | Artemisia vulgaris |
| 18 | 3.40 ± 0.15 | >23.73 | >23.73 | >6.98 | >6.98 | Morus alba |
| 19 | 5.80 ± 0.07 | >35.03 | >35.03 | >6.04 | >6.04 | Caesalpinia sappan |
| 20 | 0.52 ± 0.01 | >24.10 | >24.10 | >46.34 | >46.34 | Cudrania tricuspidata |
| 21 | 3.31 ± 0.75 | >23.13 | >23.13 | >6.99 | >6.99 | Glycine max |
| 22 | 6.50 ± 0.52 | >23.14 | >23.14 | >3.56 | >3.56 | Sophora japonica |
| Pentamidine c | 0.004 ± 0.003 | <0.40 | <0.40 | <26.7 | <26.7 | - |
| Chlorpromazine c | - | 19.89 | 16.50 | - | - | - |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.N.; No, J.H.; Lee, G.Y.; Li, W.; Yang, S.Y.; Yang, G.; Schmidt, T.J.; Kang, J.S.; Kim, Y.H. Phenolic Constituents of Medicinal Plants with Activity against Trypanosoma brucei. Molecules 2016, 21, 480. https://doi.org/10.3390/molecules21040480
Sun YN, No JH, Lee GY, Li W, Yang SY, Yang G, Schmidt TJ, Kang JS, Kim YH. Phenolic Constituents of Medicinal Plants with Activity against Trypanosoma brucei. Molecules. 2016; 21(4):480. https://doi.org/10.3390/molecules21040480
Chicago/Turabian StyleSun, Ya Nan, Joo Hwan No, Ga Young Lee, Wei Li, Seo Young Yang, Gyongseon Yang, Thomas J. Schmidt, Jong Seong Kang, and Young Ho Kim. 2016. "Phenolic Constituents of Medicinal Plants with Activity against Trypanosoma brucei" Molecules 21, no. 4: 480. https://doi.org/10.3390/molecules21040480
APA StyleSun, Y. N., No, J. H., Lee, G. Y., Li, W., Yang, S. Y., Yang, G., Schmidt, T. J., Kang, J. S., & Kim, Y. H. (2016). Phenolic Constituents of Medicinal Plants with Activity against Trypanosoma brucei. Molecules, 21(4), 480. https://doi.org/10.3390/molecules21040480