
Protective Effect of a (Poly)phenol-Rich Extract Derived from Sweet Cherries Culls against Oxidative Cell Damage

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterization of Ch-PRE
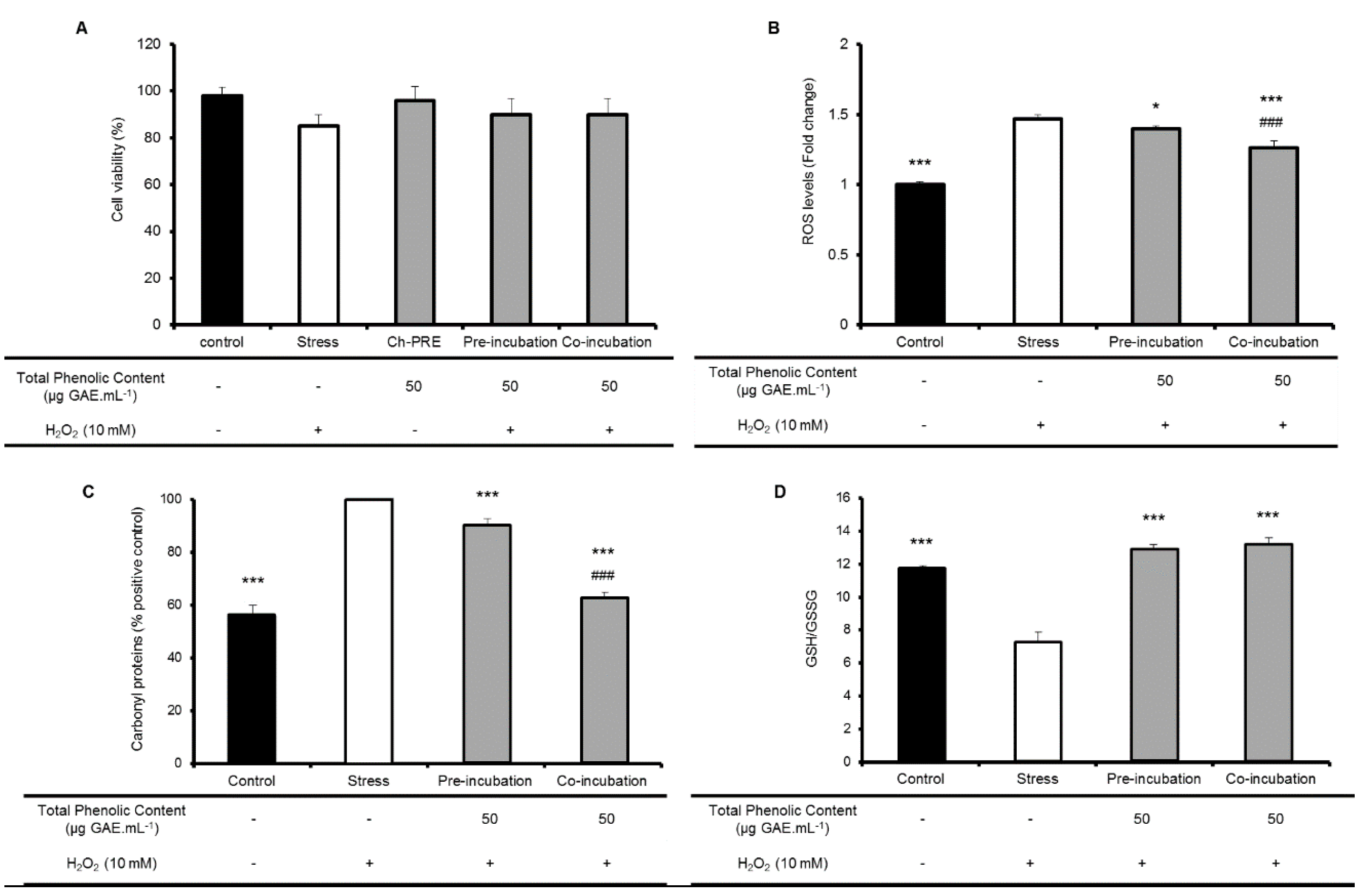
2.2. Protective Effect of Ch-PRE in Human Intestinal Epithelial Cells
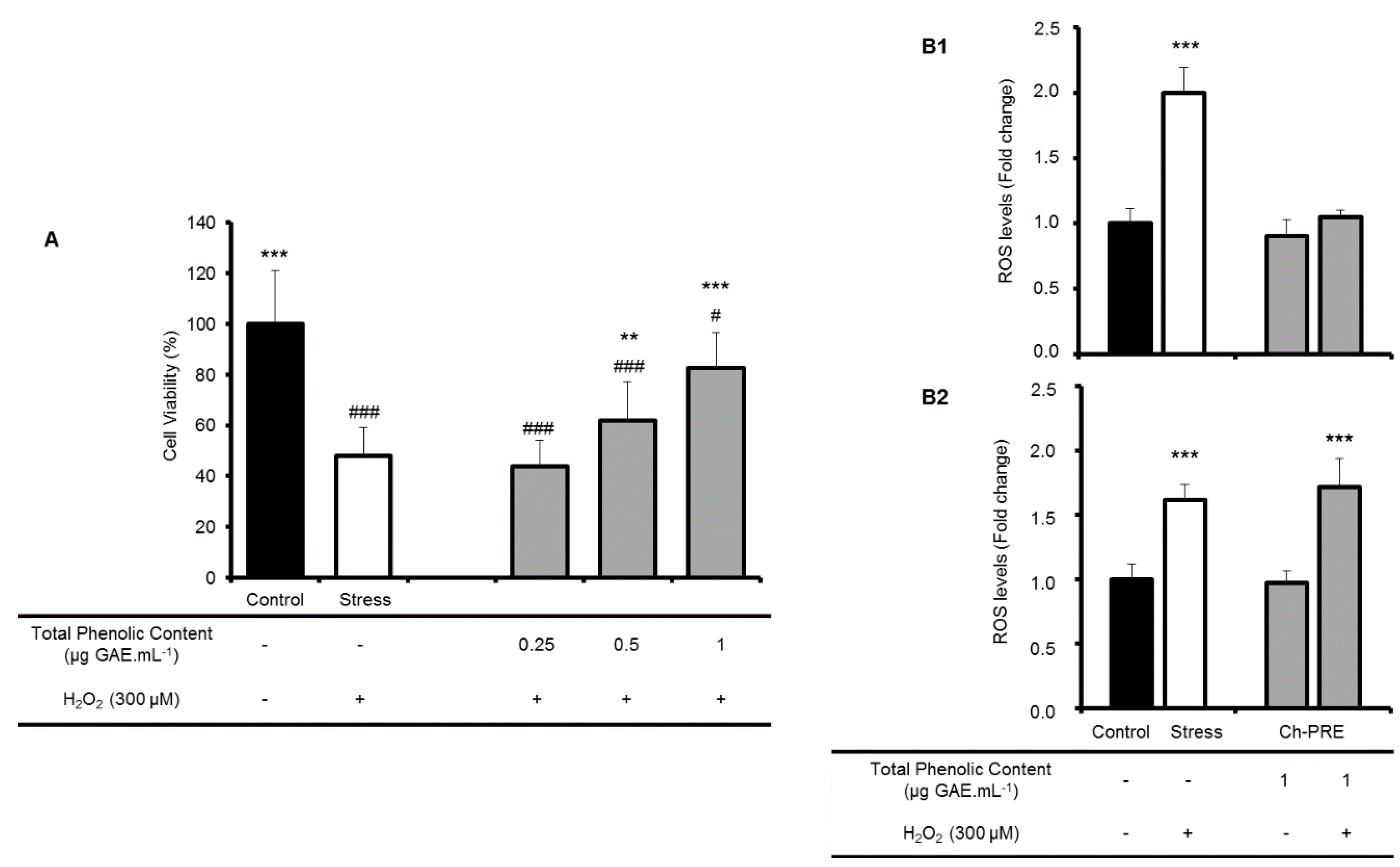
2.3. Protective Effect of Ch-PRE against Oxidative Stress in a Neurodegeneration Cell-Model
3. Experimental Section
3.1. Reagents
3.2. Raw Material
3.3. Preparation of Cherry (Poly)phenolic-Rich Extract (Ch-PRE)
3.4. Phytochemical and Antioxidant Characterization
3.4.1. Total Phenolic Content
3.4.2. HPLC-DAD-ED and LC-MS/MS Analysis
3.5. Oxygen Radical Absorbance Capacity (ORAC)
3.6. Hydroxyl Radical Adverting Capacity (HORAC)
3.7. Cell-Based Assays
3.7.1. Cell Culture
3.7.2. Intracellular Reactive Oxygen Species (ROS)
3.7.3. Determination of Protein Carbonyl Content on Intestinal Epithelial Cells
3.7.4. Glutathione (GSH) and Glutathione Disulfide (GSSG) Quantification on Intestinal Epithelial Cells
3.7.5. Evaluation of Neuroprotective Effect
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Halliwell, B. Free radicals and antioxidants—Quo vadis? Trends Pharmacol. Sci. 2011, 32, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Obrenovich, M.E.; Li, Y.; Parvathaneni, K.; Yendluri, B.B.; Palacios, H.H.; Leszek, J.; Aliev, G. Antioxidants in health, disease and aging. CNS Neurol. Disord. Drug Targets 2011, 10, 192–207. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, R.; Miranda, A.; Vergara, L. Modulation of endogenous antioxidant system by wine (poly)phenols in human disease. Clin. Chim. Acta 2011, 412, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Willcox, J.; Ash, S.; Catignani, G. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.; Mutlu, E.A.; Benson, M.; Fields, J.Z.; Banan, A.; Keshavarzian, A. Use of herbal preparations in the treatment of oxidant-mediated inflammatory disorders. Complement Ther. Med. 2007, 15, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Biswas, S.K.; Kirkham, P.A. Regulation of inflammation and redox signaling by dietary(poly)phenols. Biochem. Pharmacol. 2006, 72, 1439–1452. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, R.; Mozaffari, S.; Abdollahi, M. On the use of herbal medicines in management of inflammatory bowel diseases: A systematic review of animal and human studies. Dig. Dis. Sci. 2009, 54, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Ebrahime, A.; Scluesener, H. Natural (poly)phenols against neurodegenerative disorders: Potentials and pitfalls. Ageing Res. Rev. 2012, 11, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Macedo, D.; Tavares, L.R.; McDougall, G.J.; Miranda, H.V.; Stewart, D.; Ferreira, R.B.; Tenreiro, S.; Outeiro, T.F.; Santos, C.N. (Poly)phenols protect from α-synuclein toxicity by reducing oxidative stress and promoting autophagy. Hum. Mol. Genet. 2015, 24, 1717–1732. [Google Scholar] [CrossRef] [PubMed]
- McCune, L.; Kubota, C.; Stendell-Hollis, N.; Thomson, C. Cherries and Health: A Review. Crit. Rev. Food Sci. Nutr. 2011, 51. [Google Scholar] [CrossRef] [PubMed]
- Ship, J.; Abdel-Aal, E.M. Food Applications and Physiological Effects of Anthocyanins as Functional Food Ingredients. Open Food Sci. J. 2010, 4, 7–22. [Google Scholar] [CrossRef]
- Miguel, M.G. Anthocyanins: Antioxidant and/or anti-inflammatory activities. J. Appl. Pharm. Sci. 2011, 6, 7–15. [Google Scholar]
- Serra, A.; Duarte, R.; Bronze, M.; Duarte, C. Identification of bioactive response in traditional cherries from Portugal. Food Chem. 2011, 125, 318–325. [Google Scholar] [CrossRef]
- Ferretti, G.; Bacchetti, T.; Belleggia, A.; Neri, D. Cherry antioxidants: From farm to table. Molecules 2010, 15, 6993–7005. [Google Scholar] [CrossRef] [PubMed]
- Kappel, F.; FisherFleming, B.; Hogue, E. Fruit characteristics and sensory attributes of an ideal sweet cherry. HortScience 1996, 31, 443–446. [Google Scholar]
- Predieri, A.; Dris, R.; Rapparini, F. Influence of growing conditions on yield and quality of cherry: II. Fruit quality. J. Food Agric. Environ. 2004, 2, 307–309. [Google Scholar]
- Goncalves, B.; Landbo, A.K.; Knudsen, D.; Silva, A.P.; Moutinho-Pereira, J.; Rosa, E.; Meyer, A.S. Effect of ripeness and postharvest storage on the phenolic profiles of cherries (Prunus avium L.). J. Agric. Food Chem. 2004, 52, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Serra, A.T.; Seabra, I.J.; Braga, M.E.M.; Bronze, M.R.; de Sousa, H.C.; Duarte, C.M.M. Processing cherries (Prunus avium) using supercritical fluid technology. Part 1: Recovery of extract fractions rich in bioactive compounds. J. Supercrit. Fluids 2010, 55, 184–191. [Google Scholar] [CrossRef]
- Serra, A.T.; Matias, A.A.; Almeida, A.P.C.; Bronze, M.R.; Alves, P.M.; de Sousa, H.C.; Duarte, C.M.M. Processing cherries (Prunus avium) using supercritical fluid technology. Part 2. Evaluation of SCF extracts as promising natural chemotherapeutical agents. J. Supercrit. Fluids 2011, 55, 1007–1013. [Google Scholar] [CrossRef]
- Kammerer, D.R.; Saleh, Z.S.; Carle, R.; Stanley, R.A. Adsorptive recovery of phenolic compounds from apple juice. Eur. Food Res. Technol. 2007, 224, 605–613. [Google Scholar] [CrossRef]
- Kammerer, J.; Carle, R.; Kammerer, D.R. Adsorption and Ion Exchange: Basic Principles and Their Application in Food Processing. J. Agric. Food Chem. 2011, 59, 22–42. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.L.; Moure, A.; Domínguez, H.; Parajó, J.C. Recovery, concentration and purification of phenolic compounds by adsorption: A review. J. Food Eng. 2011, 105, 1–27. [Google Scholar] [CrossRef]
- Serra, A.T.; Poejo, J.; Matias, A.A.; Bronze, M.R.; Duarte, C.M.M. Evaluation of Opuntia spp. derived products as antiproliferative agents in human colon cancer cell line (HT29). Int. Food Res. 2013, 54, 892–901. [Google Scholar] [CrossRef]
- Matias, A.A.; Nunes, S.; Poejo, J.; Mecha, E.; Serra, A.T.; Bronze, M.R.; Duarte, C.M.M. Antioxidant and anti-inflammatory activity of a flavonoid-rich concentrate recovered from Opuntia ficus-indica juice. Food Funct. 2014, 5, 3269–3280. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, S.; Baum, M.; Eisenbrand, G.; Janzowski, C. Modulation of oxidative cell damage by reconstituted mixtures of phenolic apple juice extracts in human colon cell lines. Mol. Nutr. Food Res. 2006, 50, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Serra, A.T. Valorization of Traditional Portuguese Apples and Cherries—Biochemical Characterization and Development of Functional Ingredients. Ph.D. Thesis, Instituto de Tecnologia Química e Biológica, Universidade Nova de Lisboa, Lisbon, Portugal, 2010. [Google Scholar]
- Piccolella, S.; Fiorentino, A.; Pacifico, S.; D’Abrosca, B.; Uzzo, P.; Monaco, P. Antioxidant properties of sour cherries (Prunus cerasus L.): Role of colorless phytochemicals from the methanolic extract of ripe fruits. J. Agric. Food Chem. 2008, 56, 1928–1935. [Google Scholar] [CrossRef] [PubMed]
- Tavares, L.; Carrilho, D.; Tyagi, M.; Barata, D.; Serra, A.T.; Duarte, C.M.M.; Duarte, R.O.; Feliciano, R.P.; Bronze, M.R.; Chicau, P.; et al. Antioxidant Capacity of Macaronesian Traditional Medicinal Plants. Molecules 2010, 15, 2576–2592. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, R.P.; Bravo, M.N.; Pires, M.M.; Serra, A.T.; Duarte, C.M.; Boas, L.V.; Bronze, M.R. Phenolic Content and Antioxidant Activity of Moscatel Dessert Wines from the Setubal Region in Portugal. Food Anal. Method 2009, 2, 149–161. [Google Scholar] [CrossRef]
- Langerholc, T.; Maragkoudakisb, P.A.; Wollgastb, J.; Gradisnikc, L.; Cencica, A. Novel and established intestinal cell line models—An indispensable tool in food science and nutrition. Trends Food Sci. Technol. 2011, 22, S11–S20. [Google Scholar] [CrossRef]
- Williamson, G.; Manach, C. Bioavailability and bioefficacy of (poly)phenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005, 81, 243S–255S. [Google Scholar] [PubMed]
- Yi, W.; Akoh, C.C.; Fischer, J.; Krewer, G. Absorption of anthocyanins from blueberry extracts by Caco-2 human intestinal cell monolayers. J. Agric. Food Chem. 2006, 54, 5651–5658. [Google Scholar] [CrossRef] [PubMed]
- Deprez, S.; Mila, I.; Huneau, J.F.; Tome, D.; Scalbert, A. Transport of proanthocyanidin dimer, trimer, and polymer across monolayers of human intestinal epithelial Caco-2 cells. Antioxid. Redox Signal. 2001, 3, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Farrell, T.L.; Dew, T.P.; Poquet, L.; Hanson, P.; Williamson, G. Absorption and metabolism of chlorogenic acids in cultured gastric epithelial monolayers. Drug Metab. Dispos. 2011, 39, 2338–2346. [Google Scholar] [CrossRef] [PubMed]
- Konishi, Y.; Kobayashi, S. Transepithelial transport of chlorogenic acid, caffeic acid, and their colonic metabolites in intestinal Caco2 cell monolayers. J. Agric. Food Chem. 2004, 52, 2518–2526. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Galleano, M.; Verstraeten, S.V.; Oteiza, P.I. Basic biochemical mechanisms behind the health benefits of (poly)phenols. Mol. Aspects Med. 2010, 31, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Ballatori, N.; Krance, S.M.; Notenboom, S.; Shi, S.; Tieu, K.; Hammond, C.L. Glutathione dysregulation and the etiology and progression of human diseases. Biol. Chem. 2009, 390, 191–214. [Google Scholar] [CrossRef] [PubMed]
- Kasthuri, B.; Magalingam, A.; Kutty, R.; Nagaraja, H. Protective Mechanisms of Flavonoids in Parkinson’s Disease. Oxid. Med. Cell. Longev. 2015. [Google Scholar] [CrossRef]
- Viljanen, K.; Kylli, P.; Hubbermann, E.M.; Schwarz, K.; Heinonen, M. Anthocyanins antioxidant activity and partition behavior in Whey protein emulsion. J. Agric. Food Chem. 2005, 53, 2022–2027. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Schillig, S.; Schaffer, S.; Weber, C.C.; Eckert, G.P.; Müller, W.E. Flavonoids and the aging brain. J. Physiol. Pharmacol. 2005, 56, 23–36. [Google Scholar] [PubMed]
- Youdim, K.A.; Shukitt-Hale, B.; Joseph, J.A. Flavonoids and the brain: Interactions at the blood-brain barrier and their physiological effects on the central nervous system. Free Radic. Biol. Med. 2004, 37, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- Tavares, L.; Figueira, I.; McDougall, G.J.; Vieira, H.L.; Stewart, D.; Alves, P.M.; Ferreira, R.B.; Santos, C.N. Neuroprotective effects of digested (poly)phenols from wild blackberry species. Eur. J. Nutr. 2013, 52, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Cavazzoni, M.; Barogi, S.; Baracca, A.; Parenti Castelli, G.; Lenaz, G. The effect of aging and an oxidative stress on peroxide levels and the mitochondrial membrane potential in isolated rat hepatocytes. FEBS Lett. 1999, 449, 53–56. [Google Scholar] [CrossRef]
- Tabner, B.J.; El-Agnaf, O.M.; Turnbull, S.; German, M.J.; Paleologou, K.E.; Hayashi, Y.; Cooper, L.J.; Fullwood, N.J.; Allsop, D. Hydrogen peroxide is generated during the very early stages of aggregation of the amyloid peptides implicated in Alzheimer disease and familial British dementia. J. Biol. Chem. 2005, 280, 35789–35792. [Google Scholar] [CrossRef] [PubMed]
- Tavares, L.; Figueira, I.; Macedo, D.; McDougall, G.J.; Leitão, M.C.; Vieira, H.L.A.; Stewart, D.; Alves, P.M.; Ferreira, R.B.; Santos, C.N. Neuroprotective effect of blackberry (Rubus sp.) (poly)phenols is potentiated after simulated gastrointestinal digestion. Food Chem. 2012, 131, 1443–1452. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Remesy, C. Bioavailability and bioefficacy of (poly)phenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [PubMed]
- .Lu, Y.H.; Su, M.Y.; Huang, H.Y.; Lin, L.; Yuan, C.G. Protective effects of the citrus flavanones to PC12 cells against citotoxicity induced by hydrogen peroxide. Neurosci. Lett. 2010, 484, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Pavlica, S.; Gebhardt, R. Protective effects of flavonoids and two metabolites against oxidative stress in neuronal PC12 cells. Life Sci. 2010, 86, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Tarozzi, A.; Morroni, F.; Hrelia, S.; Angeloni, C.; Marchesi, A.; Cantelli-Forti, G.; Hrelia, P. Neuroprotective effects of anthocyanins and their in vivo metabolites in SH-SY5Y cells. Neurosci. Lett. 2007, 424, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Luo, J. Anthocyanins: Are they beneficial in treating ethanol neurotoxicity? Neurotox. Res. 2010, 17, 91–101. [Google Scholar]
- Min, J.; Yu, S.W.; Baek, S.H.; Nair, K.M.; Bae, O.N.; Bhatt, A.; Kassab, M.; Nair, M.G.; Maiid, A. Neuroprotective effect of cyanidin-3-O-glucoside anthocyanin in mice with focal cerebral ischemia. Neurosci. Lett. 2011, 500, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Son, T.G.; Camandola, S.; Mattson, M.P. Hormetic dietary phytochemicals. Neuromol. Med. 2008, 10, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.; Prior, R. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate flourescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Flanagan, J.; Deemer, E.; Prior, R.; Huang, D. Novel fluorometric assay for hydroxyl radical prevention capacity using fluorescein as the probe. J. Agric. Food Chem. 2002, 50, 2772–2777. [Google Scholar] [CrossRef] [PubMed]
- Ramful, D.; Tarnus, E.; Rondeau, P.; da Silva, C.R.; Bahorun, T.; Bourdon, E. Citrus Fruit Extracts Reduce Advanced Glycation End Products (AGEs)- and H2O2-Induced Oxidative Stress in Human Adipocytes. J. Agric. Food Chem. 2010, 58, 11119–11129. [Google Scholar] [CrossRef] [PubMed]
- Kand’ar, R.; Zakova, P.; Lotkova, H.; Kucera, O.; Cervinkova, Z. Determination of reduced and oxidized glutathione in biological samples using liquid chromatography with fluorimetric detection. J. Pharm. Biomed. Anal. 2007, 43, 1382–1387. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.


{kind=link}
{kind=link}
{kind=link}
| Peak | RT (min) | [M − H]− m/z 1 | MS/MS m/z 1 | Assigned Identity | Concentration 2 (mg·g−1 dw) |
|---|---|---|---|---|---|
| 1 | 18.28 | 353 | MS2 [353]: 135, 179, 195 | Neochlorogenic acid | 19.49 |
| 2 | 23.43 | 289 | MS2 [289]: 245, 205, 125, 109 | Catechin | 6.66 |
| 3 | 26.33 | 353 | MS2 [353]: 135, 179, 195 | Chlorogenic acid | 1.83 |
| 4 | 33.97 | 577 | MS2 [577]: 289, 125 | Procyanidin B2 | 0.98 |
| 8 | 51.91 | 609 | MS2 [609]: 301 | Quercetin-3-rutinoside | 17.00 |
| 9 | 52.85 | 463 | MS2 [463]: 301 | Quercetin-3-glucoside | 1.74 |
| 10 | 58.01 | 447 | MS2 [447]: 285 | Kaempferol-3-glucoside | 0.97 |
| 11 | 72.34 | 447 | MS2 [447]: 285 | Sakuranin | * |
| 12 | 92.12 | 285 | MS2 [285]: 243, 164 | Isosakuranetin | * |
| Peak | RT (min) | Assigned Identity | Concentration (mg·g−1 dw) |
|---|---|---|---|
| 5 | 44.73 | Cyanidin-3-glucoside | 14.04 |
| 6 | 46.56 | Cyanidin-3-rutinoside | 83.50 |
| 7 | 51.11 | Peonidin-3-glucoside | 1.60 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matias, A.A.; Rosado-Ramos, R.; Nunes, S.L.; Figueira, I.; Serra, A.T.; Bronze, M.R.; Santos, C.N.; Duarte, C.M.M. Protective Effect of a (Poly)phenol-Rich Extract Derived from Sweet Cherries Culls against Oxidative Cell Damage. Molecules 2016, 21, 406. https://doi.org/10.3390/molecules21040406
Matias AA, Rosado-Ramos R, Nunes SL, Figueira I, Serra AT, Bronze MR, Santos CN, Duarte CMM. Protective Effect of a (Poly)phenol-Rich Extract Derived from Sweet Cherries Culls against Oxidative Cell Damage. Molecules. 2016; 21(4):406. https://doi.org/10.3390/molecules21040406
Chicago/Turabian StyleMatias, Ana A., Rita Rosado-Ramos, Sara L. Nunes, Inês Figueira, Ana Teresa Serra, Maria R. Bronze, Claúdia N. Santos, and Catarina M. M. Duarte. 2016. "Protective Effect of a (Poly)phenol-Rich Extract Derived from Sweet Cherries Culls against Oxidative Cell Damage" Molecules 21, no. 4: 406. https://doi.org/10.3390/molecules21040406
APA StyleMatias, A. A., Rosado-Ramos, R., Nunes, S. L., Figueira, I., Serra, A. T., Bronze, M. R., Santos, C. N., & Duarte, C. M. M. (2016). Protective Effect of a (Poly)phenol-Rich Extract Derived from Sweet Cherries Culls against Oxidative Cell Damage. Molecules, 21(4), 406. https://doi.org/10.3390/molecules21040406