
Tea Polysaccharides and Their Bioactivities

 ,
, 
Abstract
:1. Introduction
2. Polysaccharides in Tea
2.1. Basic Composition of Tea Polysaccharides (TPS)
2.2. TPS Variation Between Tea Cultivars and Plant Organs
2.3. Effect of Tea Processing on TPS
2.4. Effect of Preparation Methods on TPS
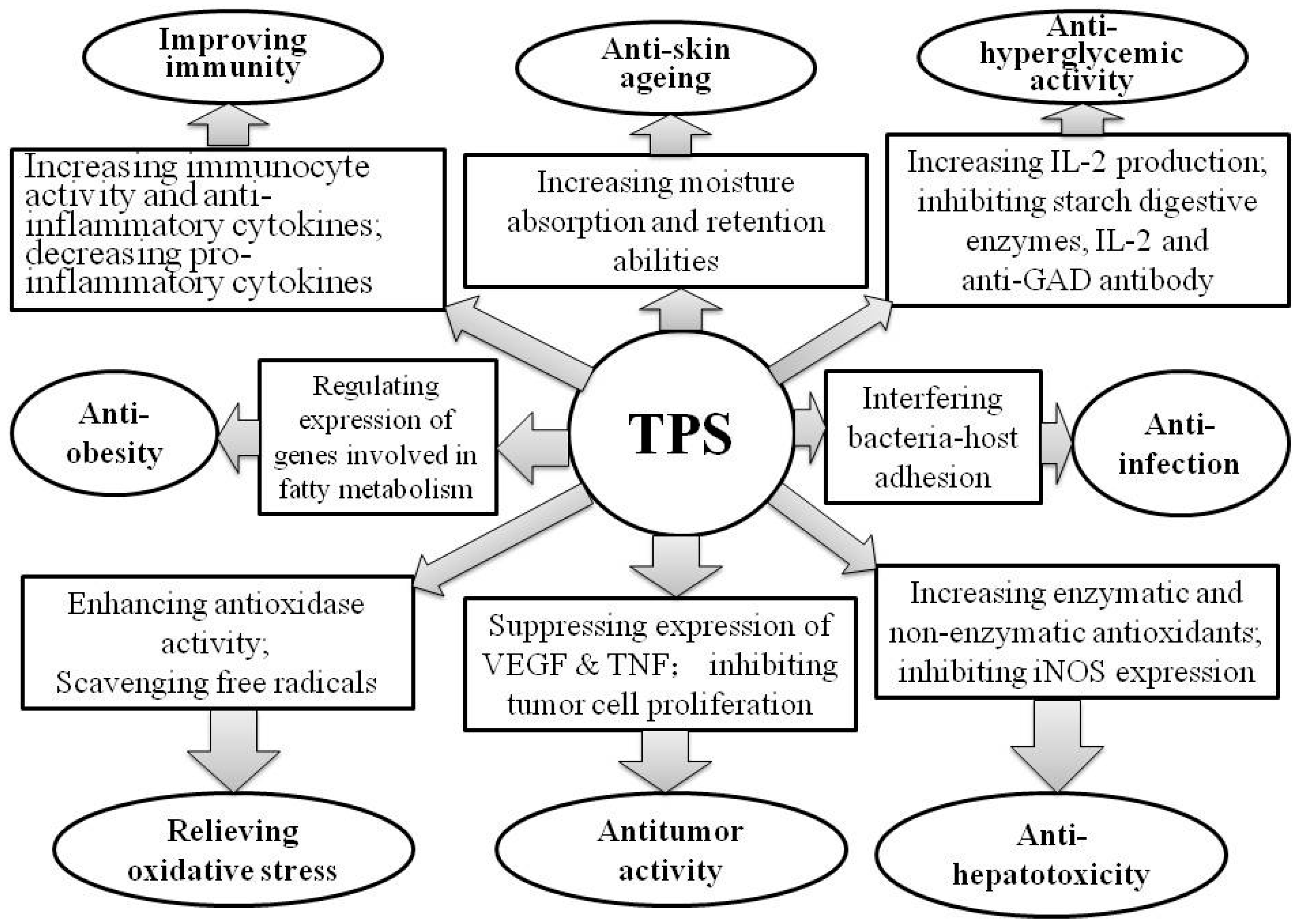
3. Bioactivities of TPS
3.1. Bioavailability and Toxicity of TPS
3.2. Alleviating Oxidative Stress
3.3. Antitumor
3.4. Anti-Hyperglycemia
3.5. Improving Immunity
3.6. Anti-Hepatotoxicity
3.7. Anti-Skin Aging
3.8. Anti-Infection of Pathogenic Bacteria
4. Inconsistent Results
4.1. Inconsistent Chemical Composition of TPS
4.2. Controversial Antioxidant Activities
4.3. Inconsistent Relationship between Single Bioactive Compound Content and Bioactivity
4.4. Inconsistent Relationship between TPS Molecular Weight (MW) and Bioactivity
4.5. Further Study Suggestions
5. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Full Name | Abbreviation | Full Name |
| ABTS | 2,2′-Azinobis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt | IGT | Impaired glucose tolerance |
| ALP | Alkaline phosphatase | IL | Interleukin |
| ALT | Alanine transaminase | iNOS | Inducible nitric oxide synthase |
| AOT | Sodium di-2-ethylhexyl sulfosuccinate | LPS | Lipopolysaccharide |
| Ara | Arabinose | Man | Mannose |
| AST | Aspartate transaminase | MDA | Malondialdehyde |
| ATPS | Acid tea polysaccharides | MIC | Minimum inhibitory concentration |
| ATSPS | Acid tea seed polysaccharides | MW | Molecular weight |
| BG | Blood glucose | nNOS | Neural nitric oxide synthase |
| BTPS | Black tea polysaccharides | NOD | Non-obese diabetic |
| BW | Body weight | NOS | Nitric oxide synthase |
| CAT | Catalase | NTPS | Neutral tea polysaccharides |
| CCl4 | Carbon tetrachloride | OECD | Organization for Economic Co-operation and Development |
| ConA | Concanavalin A | OTPS | Oolong tea polysacharides |
| DC | Dendritic cell | PCNA | Proliferating cell nuclear antigen |
| DM | Diabetes mellitus | PFPS | Pyracantha fortuneana Polysaccharides |
| DPPH | 1,1-Diphenyl-2-picrylhydrazyl | PTPS | Pu-erh tea polysaccharides |
| EGCG | Epigallocatechin gallate | REE | Rare earth elements |
| eNOS | Endothelial nitric oxide synthase | Rha | Rhamnose |
| ERBS | elements related to reducing blood sugar | Rib | Ribose |
| FRAP | Ferric-reducing antioxidant power | SOD | Superoxide dismutase |
| Fuc | Fucose | T1D | Type 1 diabetes |
| GAD | Glutamic acid decarboxylase | TC | Cholesterol |
| Gal | Galactose | TCR | T cell antigen receptor |
| GalA | Galacturonic acid | TFPPS | Tea fruit peel polysaccharides |
| GHS | Globally Harmonized System | TFPS | Tea flower polysaccharides |
| Glc | Glucose | TG | Triglycerides |
| GSH-Px | Glutathione peroxidase | TLPS | Tea leaf polysaccharides |
| GTPS | Green tea polysacharides | TLR4 | Toll-like receptor 4 |
| GulA | Glucuronic acid | TNF-α | Tumor necrosis factor-alpha |
| HCC | Hepatocarcinoma | TPS | Tea polysaccharides |
| HDF | Human diploid fibroblast | TPSIC | TPS-iron complex |
| HG | Homogalacturonan | Treg | Regulatory T cell |
| HMPS | Hawk mature tea polysaccharides | TSPS | Tea seed polysaccharides |
| HUVE | Human umbilical vein endothelial | TPP | Tea polyphenols |
| IC50 | 50% Inhibitory concentration | UV | Ultraviolet |
| 8-iso-PGF2α | 8-Iso-prostaglandin F2 alpha | VEGF | Vascular endothelial growth factor |
| IFN-γ | Interferon-gamma | WBC | White blood cells |
| Ig | Immunoglobulin | Xyl | Xylose |
References
- Zheng, X.Q.; Li, Q.S.; Xiang, L.P.; Liang, Y.R. Recent advances in volatiles of teas. Molecules 2016, 21, 338. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.P.; Wang, A.; Ye, J.H.; Zheng, X.Q.; Polito, C.A.; Lu, J.L.; Li, Q.S.; Liang, Y.R. Suppressive effects of tea catechins on breast cancer. Nutrients 2016, 8, 458. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.R.; Liu, C.; Xiang, L.P.; Zheng, X.Q. Health benefits of theanine in green tea: A review. Trop. J. Pharm. Res. 2015, 14, 1943–1949. [Google Scholar] [CrossRef]
- Guo, L.; Liang, Q.; Du, X. Effects of molecular characteristics of tea polysaccharide in green tea on glass transitions of potato amylose, amylopectin and their mixtures. Food Hydrocolloid. 2011, 25, 486–494. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on mechanisms of in vitro antioxidant activity of polysaccharides. Oxid. Med. Cell. Longev. 2016, 64, 1–13. [Google Scholar]
- Ren, D.Y.; Hu, Y.Y.; Luo, Y.Y.; Yang, X.B. Selenium-containing polysaccharides from Ziyang green tea ameliorate high-fructose diet induced insulin resistance and hepatic oxidative stress in mice. Food Funct. 2015, 6, 3342–3350. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wu, J.; Zhang, Y.; Chen, H.; Wang, Y.F. Physicochemical characterization of puerh tea polysaccharides and their antioxidant and alpha-glycosidase inhibition. J. Funct. Foods 2014, 6, 545–554. [Google Scholar] [CrossRef]
- Wang, Y.F.; Shao, S.H.; Xu, P.; Chen, H.; Lin-Shiau, S.Y.; Deng, Y.T.; Lin, J.K. Fermentation process enhanced production and bioactivities of oolong tea polysaccharides. Food Res. Int. 2012, 46, 158–166. [Google Scholar] [CrossRef]
- Chen, H.X.; Zhang, M.; Qu, Z.S.; Xie, B.J. Compositional analysis and preliminary toxicological evaluation of a tea polysaccharide conjugate. J. Agric. Food Chem. 2007, 55, 2256–2260. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Yang, X.; Zhao, Y.; Tuan, Y.; Yang, Y.; Wang, Z. Separation and quantification of component monosaccharides of the tea polysaccharides from Gynostemma. pentaphyllum by HPLC with indirect UV detection. Food Chem. 2009, 112, 742–746. [Google Scholar] [CrossRef]
- Zhou, X.L.; Wang, D.F.; Sun, P.N.; Bucheli, P.; Li, L.; Hou, Y.F.; Wang, J.F. Effects of soluble tea polysaccharides on hyperglycemia in alloxan-diabetic mice. J. Agric. Food Chem. 2007, 55, 5523–5528. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.F.; Liu, Y.Y.; Huo, J.L.; Zhao, T.T.; Ren, J.; Wei, X.L. Effect of different drying methods on chemical composition and bioactivity of tea polysaccharides. Int. J. Biol. Macromol. 2013, 62, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Wei, G.D.; Liu, F.; Banerjee, G.; Joshi, M.; Bligh, S.W.A.; Shi, S.S.; Lian, H.; Fan, H.W.; Gu, X.L.; Wang, S.C. Characterization of two homogalacturonan pectins with immunomodulatory activity from green tea. Int. J. Mol. Sci. 2014, 15, 9963–9978. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M.; Xia, W.S. Isolation and analysis of a novel acidic polysaccharide with glucokinase-stimulating activity from coarse green tea. J. Food Biochem. 2006, 30, 187–202. [Google Scholar] [CrossRef]
- Wang, Y.F.; Peng, Y.H.; Wei, X.L.; Yang, Z.W.; Xiao, J.B.; Jin, Z.Y. Sulfation of tea polysaccharides: Synthesis, characterization and hypoglycemic activity. Int. J. Biol. Macromol. 2010, 46, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Ye, Y.; Cheng, H.; Jiang, Y.W.; Wu, Y.L. Thermal effects on the stability and antioxidant activity of an acid polysaccharide conjugate derived from green tea. J. Agric. Food Chem. 2009, 57, 5795–5798. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, C.; Zhao, G.; Wei, Z.; Tao, Y.; Liang, X. Composition, characteristic and activity of rare earth element-bound polysaccharide from tea. Biosci. Biotechnol. Biochem. 2001, 65, 1987–1992. [Google Scholar] [CrossRef] [PubMed]
- TsuBaki, S.; Sakumoto, S.; Uemura, N.; Azuma, J. Compositional analysis of leaf cuticular membranes isolated from tea plants (Camellia sinensis L.). Food Chem. 2012, 138, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.F.; Wang, C.H.; Li, J.; Zhao, G.W. Components and activity of polysaccharides from coarse tea. J. Agric. Food Chem. 2001, 49, 507–510. [Google Scholar] [CrossRef]
- Wang, Y.F.; Yang, Z.W.; Wei, X.L. Sugar compositions, alpha-glucosidase inhibitory and amylase inhibitory activities of polysaccharides from leaves and flowers of Camellia sinensis obtained by different extraction methods. Int. J. Biol. Macromol. 2010, 47, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.L.; Mao, F.F.; Cai, X.; Wang, Y.F. Composition and bioactivity of polysaccharides from tea seeds obtained by water extraction. Int. J. Biol. Macromol. 2011, 49, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.F.; Wang, J.; Wu, J.; Xu, P.; Wang, Y.Q.; Gao, J.J.; Hochstetter, D. In vitro antioxidant activity and potential inhibitory action against α-glucosidase of polysaccharides from fruit peel of tea (Camellia sinensis L.). J. Zhejiang Univ. Sci. B (Biomed. Biotech.) 2014, 15, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.J.; Ding, C.B.; Yuan, S.; Zhang, Z.W.; Chen, Y.E.; Du, L.; Yuan, M. Extraction, purification and characterization of polysaccharides from Hawk tea. Carbohyd. Polym. 2014, 99, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Takeo, T.; Unno, T.; Kinogasa, H.; Yayabe, F.; Motoyama, M. The chemical properties and functional effects of polysaccharides dissolved in green tea infusion. Nippon Shokuhin. Kagaku Kaishi 1998, 45, 270–272. [Google Scholar] [CrossRef]
- Chen, H.X.; Zhang, M.; Xie, B.J. Components and antioxidant activity of polysaccharide conjugate from green tea. Food Chem. 2005, 90, 17–21. [Google Scholar] [CrossRef]
- Xiao, J.B.; Huo, J.L.; Jiang, H.X.; Yang, F. Chemical compositions and bioactivities of crude polysaccharides from tea leaves beyond their useful date. Int. J. Biol. Macromol. 2011, 49, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.F.; Yang, X.B.; Lu, X.S.; Wang, D.Y.; Zhao, Y. Protective effects of Keemun black tea polysaccharides on acute carbon tetrachloride-caused oxidative hepatotoxicity in mice. Food Chem. Toxicol. 2013, 58, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.X.; Qu, Z.S.; Fu, L.L.; Dong, P.; Zhang, X. Physicochemical properties and antioxidant capacity of 3 polysaccharides from green tea, oolong tea, and black tea. J. Food Sci. 2009, 74, C469–C474. [Google Scholar] [CrossRef] [PubMed]
- Du, W.H.; Peng, S.M.; Liu, Z.H.; Shi, L.; Tan, L.F.; Zou, X.Q. Hypoglycemic effect of the water extract of pu-erh tea. J. Agric. Food Chem. 2012, 60, 10126–10132. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.S.; Hu, X.J.; Peng, C.X.; Zhou, H.J. The molecular composition and spectral properties of polysaccharide isolated from Pu-erh teaand its material. Spectrosc. Spect. Anal. 2010, 30, 1960–1964. [Google Scholar]
- Zhang, Z.S.; Wang, X.M.; Li, J.F.; Wang, G.Z.; Mao, G.X. Extraction and free radical scavenging activity of polysaccharide from′Anji Baicha′ (Camellia sinensis (L.) O. Kuntze). Int. J. Biol. Macromol. 2016, 84, 161–165. [Google Scholar] [CrossRef] [PubMed]
- TsuBaki, S.; Iida, H.; Sakamoto, M.; Azuma, J.I. Microwave heating of tea residue yields polysaccharides, polyphenols, and plant biopolyester. J. Agric. Food. Chem. 2008, 56, 11293–11299. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.H.; Shin, K.S.; Park, Y.; Yu, K.W.; Suh, H.J.; Choi, H.S. Biotransformation of catechin and extraction of active polysaccharide from green tea leaves via simultaneous treatment with tannase and pectinase. J. Sci. Food Agric. 2015, 95, 2337–2344. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, H.X.; Zhang, N.; Chen, S.H.; Tian, J.G.; Zhang, Y.; Wang, Z.S. Extrusion treatment for improved physicochemical and antioxidant properties of high-molecular weight polysaccharides isolated from coarse tea. Food Res. Int. 2013, 53, 726–731. [Google Scholar] [CrossRef]
- Chen, M.; Xiong, L.Y. Supercritical extraction technology in tea polysaccharide extracting application. Adv. Mater. Res. 2012, 1683–1688. [Google Scholar] [CrossRef]
- Li, S.; Cao, X. Extraction of tea polysaccharides (TPS) using anionic reverse micellar system. Sep. Purif. Technol. 2014, 122, 306–314. [Google Scholar] [CrossRef]
- Chen, H.X.; Zhang, M.; Qu, Z.S.; Xie, B.J. Antioxidant activities of different fractions of polysaccharide conjugates from green tea (Camellia sinensis). Food Chem. 2008, 106, 559–563. [Google Scholar] [CrossRef]
- Yang, P.J.; Zhou, M.D.; Zhou, C.Y.; Wang, Q.; Zhang, F.F.; Chen, J. Separation and purification of both tea seed polysaccharide and saponin from camellia cake extract using macroporous resin. J. Sep. Sci. 2015, 38, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.F.; Nie, S.P.; Han, C.; Chen, Y.Q.; Xie, M.Y.; Gong, D.M. Regulation of maturation and function of dendritic cells by tea glycoprotein. Eur. Food Res. Technol. 2012, 235, 1023–1032. [Google Scholar]
- United Nations. Globally Harmonized System of Classification and Labelling of Chemicals (GHS), Third revised edition; United Nations: New York, NY, USA, 2009. [Google Scholar]
- Zhou, Y.B.; Wang, D.F.; Wan, X.C.; Zhang, L.; Du, X.F.; Hu, W.S. Effect of tea polysaccharide addition on the properties of bread made from two flours. J. Food Process. Pres. 2009, 33, 798–813. [Google Scholar] [CrossRef]
- Zhang, H.H.; Sun, B.H.; Zhang, S.K.; Zhu, Y.J.; Tian, Y.Q. Inhibition of wheat starch retrogradation by tea derivatives. Carbohyd. Polym. 2015, 134, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Mellinger, C.G.; Cipriani, T.R.; Noleto, G.R.; Carbonero, E.R.; Oliveira, M.B.M.; Gorin, P.A.J.; Iacomini, M. Chemical and immunological modifications of an arabinogalactan present in tea preparations of Phyllanthus. niruri after treatment with gastric fluid. Int. J. Biol. Macromol. 2008, 43, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.M.; Wang, D.F.; Hou, Y.F.; Buchili, P.; Sun, L.P. Preparation, characterization, bioavailability in vitro and in vivo of tea polysaccharides–iron complex. Eur. Food Res. Technol. 2013, 236, 341–350. [Google Scholar] [CrossRef]
- Yang, J.J.; Chen, B.; Qu, Y. Pharmacological evaluation of tea polysaccharides with antioxidant activity in gastric cancer mice. Carbohyd. Polym. 2012, 90, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.M. Evaluation of antioxidant activity of polysaccharides isolated from Camellia sinensis (tea) in exhausting training mice. J. Med. Plants Res. 2011, 5, 791–795. [Google Scholar]
- Wang, Y.F.; Mao, F.F.; Wei, X.L. Characterization and antioxidant activities of polysaccharides from leaves, flowers and seeds of green tea. Carbohyd. Polym. 2012, 88, 146–153. [Google Scholar] [CrossRef]
- Xu, R.; Ye, H.; Sun, Y.; Tu, Y.; Zeng, X. Preparation, preliminary characterization, antioxidant, hepatoprotective and antitumor activities of polysaccharides from the flower of tea plant (Camellia sinensis). Food Chem. Toxicol. 2012, 50, 2473–2480. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Y.; Huangfu, L.T.; Dong, L.L.; Liu, S.L. Functional groups and antioxidant activities of polysaccharides from five categories of tea. Ind. Crop. Prod. 2014, 58, 31–35. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, Y.; Yang, F. EGCG enhanced the antioxidant potential of purified fractions from crude tea polysaccharides. Free Radical Bio. Med. 2012, 53, S90. [Google Scholar] [CrossRef]
- Yuan, C.F.; Li, Z.H.; Peng, F.; Xiao, F.X.; Ren, D.M.; Xue, H.; Chen, T.; Mushtaq, G.; Kamal, M.A. Combination of selenium-enriched green tea polysaccharides and Huo-ji polysaccharides synergistically enhances antioxidant and immune activity in mice. J. Sci. Food Agric. 2015, 95, 3211–3217. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Mao, F.; Liu, Y.; Wei, X. Purification, characterization and biological activities in vitro of polysaccharides extracted from tea seeds. Int. J. Biol. Macromol. 2013, 62, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Zhang, D.; Zhang, Y.; Wen, Y.; Li, L.; Zheng, L. Tumoricidal effects of a selenium (Se)-polysaccharide from Ziyang green tea on human osteosarcoma U-2 OS cells. Carbohyd. Polym. 2013, 98, 1186–1190. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Ling, Z.J.; He, P.M.; Xiong, C.Y. Immunomodulatory and antitumor activity of polysaccharide isolated from tea plant flower. Prog. Biochem. Biophys. 2010, 37, 646–653. [Google Scholar] [CrossRef]
- Fan, Y.L.; Wang, W.H.; Hu, Y.S.; Rong, Q.; Teng, A.G.; Liu, A.J. Comparison of the roles of tea polysaccharides and polyphenols in growth inhibition of hepatoma h22 cells in mice. Adv. Mate. Re. 2013, 690, 1193–1197. [Google Scholar] [CrossRef]
- Chen, B.; Zhou, W.; Ning, M.; Wang, Z.; Zou, L.; Zhang, H.; Wang, Q. Evaluation of antitumour activity of tea carbohydrate polymers in hepatocellular carcinoma animals. Int. J. Biol. Macromol. 2012, 50, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Chen, H.; Wang, Y.; Hochstetter, D.; Zhou, T.; Wang, Y. Oral administration of puerh tea polysaccharides lowers blood glucose levels and enhances antioxidant status in alloxan-induced diabetic mice. J. Food Sci. 2012, 77, H246–H252. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.Y.; Nie, S.P.; Fu, B.Q.; Wang, X.R. Determination of elements related to reducing g blood sugar (ERBS) in tea and tea polysaccharide by ICP-MS. Spectrosc. Spect. Anal. 2006, 26, 1710–1715. [Google Scholar]
- Stehouwer, C.D.A.; Lambert, J.; Donker, A.J.M.; vanHinsbergh, V.W.M. Endothelial dysfunction and pathogenesis of diabetic angiopathy. Cardiovasc. Res. 1997, 34, 55–68. [Google Scholar] [CrossRef]
- Hink, U.; Li, H.G.; Mollnau, H.; Oelze, M.; Matheis, E.; Hartmann, M.; Skatchkov, M.; Thaiss, F.; Stahl, R.A.K.; Warnholtz, A.; et al. Mechanisms underlying endothelial dysfunction in diabetes mellitus. Circ. Res. 2001, 88, H14–H22. [Google Scholar] [CrossRef]
- Dimmeler, S.; Zeiher, A.M. Endothelial cell apoptosis in angiogenesis and vessel regression. Circ. Res. 2000, 87, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Wang, Y.F.; Wu, Y.L.; Han, B.Y.; Zhu, Y.J.; Tang, X.L.; Sun, Q.L. Green tea polysaccharide-conjugates protect human umbilical vein endothelial cells against impairments triggered by high glucose. Int. J. Biol. Macromol. 2011, 49, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.J.; Sim, L.; Kuntz, D.A.; Hahn, D.; Johnston, B.D.; Ghavami, A.; Szczepina, M.G.; Kumar, N.S.; Sterchi, E.E.; Nichols, B.L.; Pinto, B.M.; Rose, D.R. Inhibition of recombinant human Maltase glucoamylase by salacinol and derivatives. FEBS J. 2006, 273, 2673–2683. [Google Scholar] [CrossRef] [PubMed]
- Hanhineva, K.; Törrönen, R.; Bondia-Pons, I.; Pekkinen, J.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Impact of dietary polyphenols on carbohydrate metabolism. Int. J. Mol. Sci. 2010, 11, 1365–1402. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Fang, Y.P.; Nishinari, K.; We, H.; Sun, C.C.; Li, J.R.; Jiang, Y.W. Physicochemical characteristics of polysaccharide conjugates prepared from fresh tea leaves and their improving impaired glucose tolerance. Carbohyd. Poly. 2014, 112, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.L.; Cai, X.; Xiong, S.L.; Wang, Y.F. Hypoglycemic effect of oral crude tea flower polysaccharides on alloxan modeling SPrague–Dawley rats and the possible mechanism. CyTA-J. Food 2012, 10, 325–332. [Google Scholar] [CrossRef]
- Deng, Y.T.; Lin-Shiau, S.Y.; Shyur, L.F.; Lin, J.K. Pu-erh tea polysaccharides decrease blood sugar by inhibition of α-glucosidase activity in vitro and in mice. Food Funt. 2015, 6, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, C.J.; Ward, N.C.; Pugliese, A.; Malek, T.R. Promoting immune regulation in type 1 diabetes using low-dose interleUKin-2. Curr. Diab. Rep. 2016, 16, 46. [Google Scholar] [CrossRef] [PubMed]
- Brizeno, L.A.C.; Assreuy, A.M.S.; Alves, A.P.N.N.; Sousa, F.B.; Silva, P.G.D.; de Sousa, S.C.O.M.; Lascane, N.A.S.; Evangelista, J.S.A.M.; Mota, M.R.L. Delayed healing of oral mucosa in a diabetic rat model: Implication of TNF-α, IL-1β and FGF-2. Life Sci. 2016, 155, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Q.; Chen, H.X.; Wang, J.; Wang, X.M.; Hu, B.; Lv, F.N. Involvement of the PI3K/Akt signal pathway in the hypoglycemic effects of tea polysaccharides on diabetic mice. Int. J. Biol. Macromol. 2015, 81, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.C.; Lara, G.M.; Linden, R.; Feksa, L.R.; Tavares, R.G.; Almeida, S.E.D.; Berlese, D.B. Frequency of the anti-glutamic acid decarboxylase immunological marker in patients with diabetes duration longer than three years in southern Brazil. Sao Paulo Med. J. 2011, 129, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Lin, Z.; Ye, Y.; Zhang, R.; Yin, J.F.; Jiang, Y.W.; Wan, H.T. Suppression of diabetes in non-obese diabetic (NOD) mice by oral administration of water-soluble and alkali-soluble polysaccharide conjugates prepared from green tea. Carbohyd. Polym. 2010, 82, 28–33. [Google Scholar] [CrossRef]
- Monobe, M.; Ema, K.; Tokuda, Y.; Maeda-Yamamoto, M. Enhancement of the phagocytic activity of macrophage-like cells with a crude polysaccharide derived from green tea (Camellia sinensis) extract. Biosci. Biotechnol. Biochem. 2010, 74, 1306–1308. [Google Scholar] [CrossRef] [PubMed]
- Monobe, M.; Ema, K.; Kato, F.; Maeda-Yamamoto, M. Immunostimulating activity of a crude polysaccharide derived from green tea (Camellia sinensis) extract. J. Agric. Food Chem. 2008, 56, 1423–1427. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Xiong, C.Y.; Shi, J.; Gao, Y.; Chen, Y.S.; Ling, Z.J.; He, P.M. Isolation, chemical characterization and antioxidant activities of a water-soluble polysaccharide fraction of tea (Camellia sinensis) flower. J. Food Biochem. 2012, 36, 46–55. [Google Scholar] [CrossRef]
- Lu, X.S.; Zhao, Y.; Sun, Y.F.; Yang, S.; Yang, X.B. Characterisation of polysaccharides from green tea of Huangshan Maofeng with antioxidant and hepatoprotective effects. Food Chem. 2013, 141, 3415–3423. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, A.E.; Piao, W.; Shoenfelt, J.; Rhee, S.H.; Chen, H.; Basu, S.; Wahl, L.M.; Fenton, M.J.; Vogel, S.N. Role of TLR4 tyrosine phosphorylation in signal transduction and endotoxin tolerance. J. Biol. Chem. 2007, 282, 16042–16053. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, G.; Li, C.J.; Zhang, M.; Zhao, H.; Sheng, J.; Shi, W. Pu-erh tea reduces nitric oxide levels in rats by inhibiting inducible nitric oxide synthase expression through Toll-like receptor 4. Int. J. Mol. Sci. 2012, 13, 7174–7185. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Rong, H.; Chu, X.H.; Dou, T.F.; Gao, L.; Gu, D.H.; Lin, Q.Y.; Cao, Z.H.; Ge, C.R.; Jia, J.J.; Xu, Z.Q. Effects of natural antioxidant extracts on the molecular mechanism of anti-aging in HDF. J. Yunnan Agric. Univ. 2015, 30, 219–227. [Google Scholar]
- Wei, X.L.; Liu, Y.; Xiao, J.B.; Wang, Y.F. Protective effects of tea polysaccharides and polyphenols on skin. J. Agric. Food Chem. 2009, 57, 7757–7762. [Google Scholar] [CrossRef] [PubMed]
- Skovgaard, G.R.L.; Jensen, A.S.; Sigler, M.L. Effect of a novel dietary supplement on skin aging in post-menopaUSAl women. Eur. J. Clin. Nutr. 2006, 60, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Shim, J.S.; Lee, J.S.; Kim, J.K.; Yang, I.S.; Chung, M.S.; Kim, K.H. Inhibition of pathogenic bacterial adhesion by acidic polysaccharide from green tea (Camellia sinensis). J. Agric. Food Chem. 2006, 54, 8717–8723. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Shim, J.S.; Chung, M.S.; Lim, S.T.; Kim, K.H. In vitro anti-adhesive activity of green tea extract against pathogen adhesion. Phytother. Res. 2009, 23, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L.A. Host-pathogen interactions: Basic concepts of microbial commensalism, colonization, infection, and disease. Infect. Immun. 2000, 68, 6511–6518. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Yu, Q.Y.; Shi, J.; Xiong, C.Y.; Ling, Z.J.; He, P.M. Structural characterization and antioxidant activities of 2 water-soluble polysaccharide fractions purified fromtea (Camellia sinensis) flower. J. Food Sci. 2011, 76, C462–C471. [Google Scholar]
- Wang, H.L.; Zhao, Y.; Andrae-Marobela, K.; Okatch, H.; Xiao, J.B. Tea polysaccharides as food antioxidants: An old wOman′s tale? Food Chem. 2013, 138, 1923–1927. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.X.; Zhang, M.; Xie, B.J. Quantification of uronic acids in tea polysaccharide conjugates and their antioxidant properties. J. Agric. Food Chem. 2004, 52, 3333–3336. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Guo, Y.; Liu, R.; Wang, K.; Zhang, M. Black tea polyphenols and polysaccharides improve body composition, increase fecal fatty acids, and regulate fat metabolism in high-fat diet-induced obese rats. Food Funct. 2016, 7, 2469–2478. [Google Scholar] [CrossRef] [PubMed]
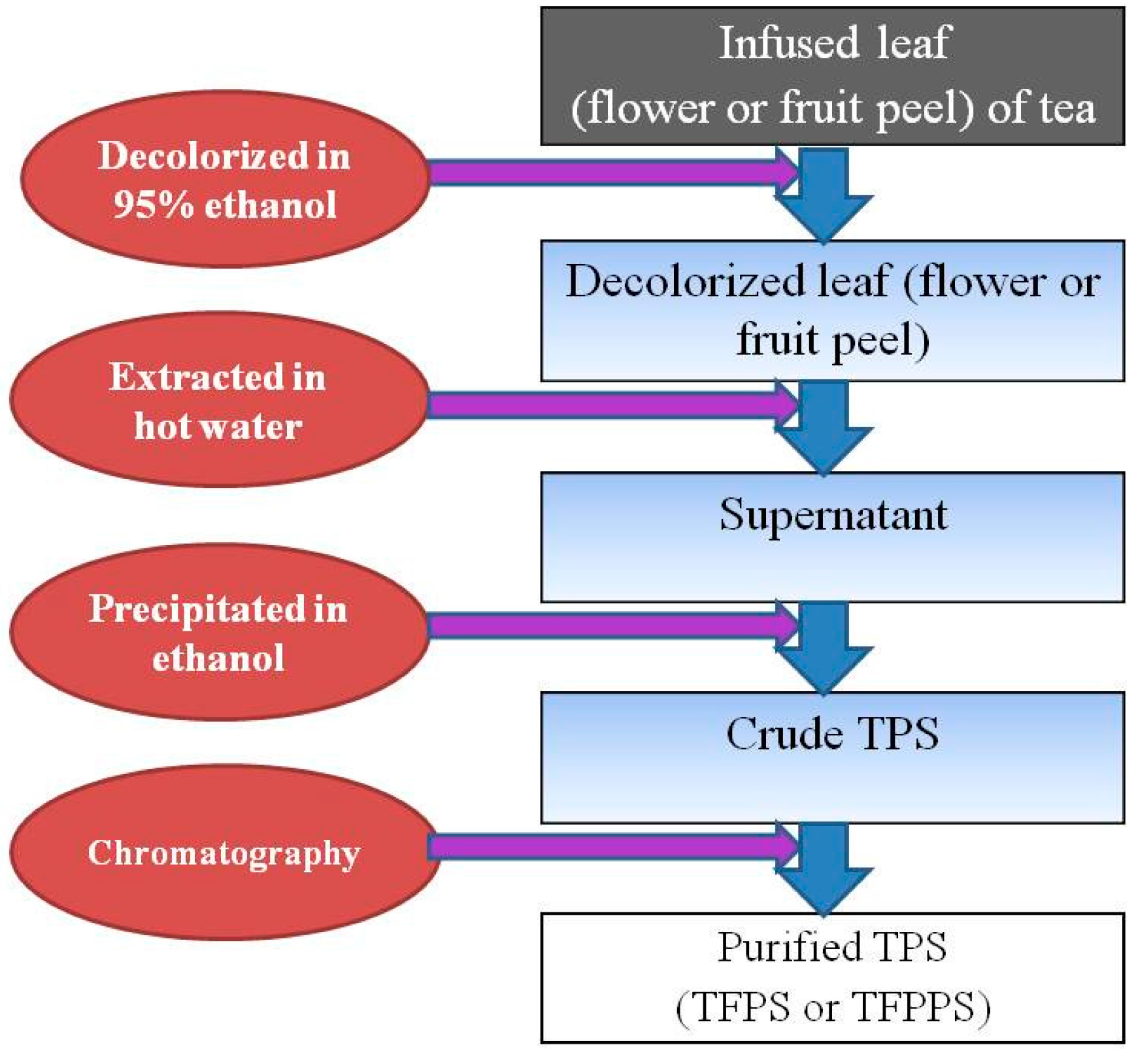
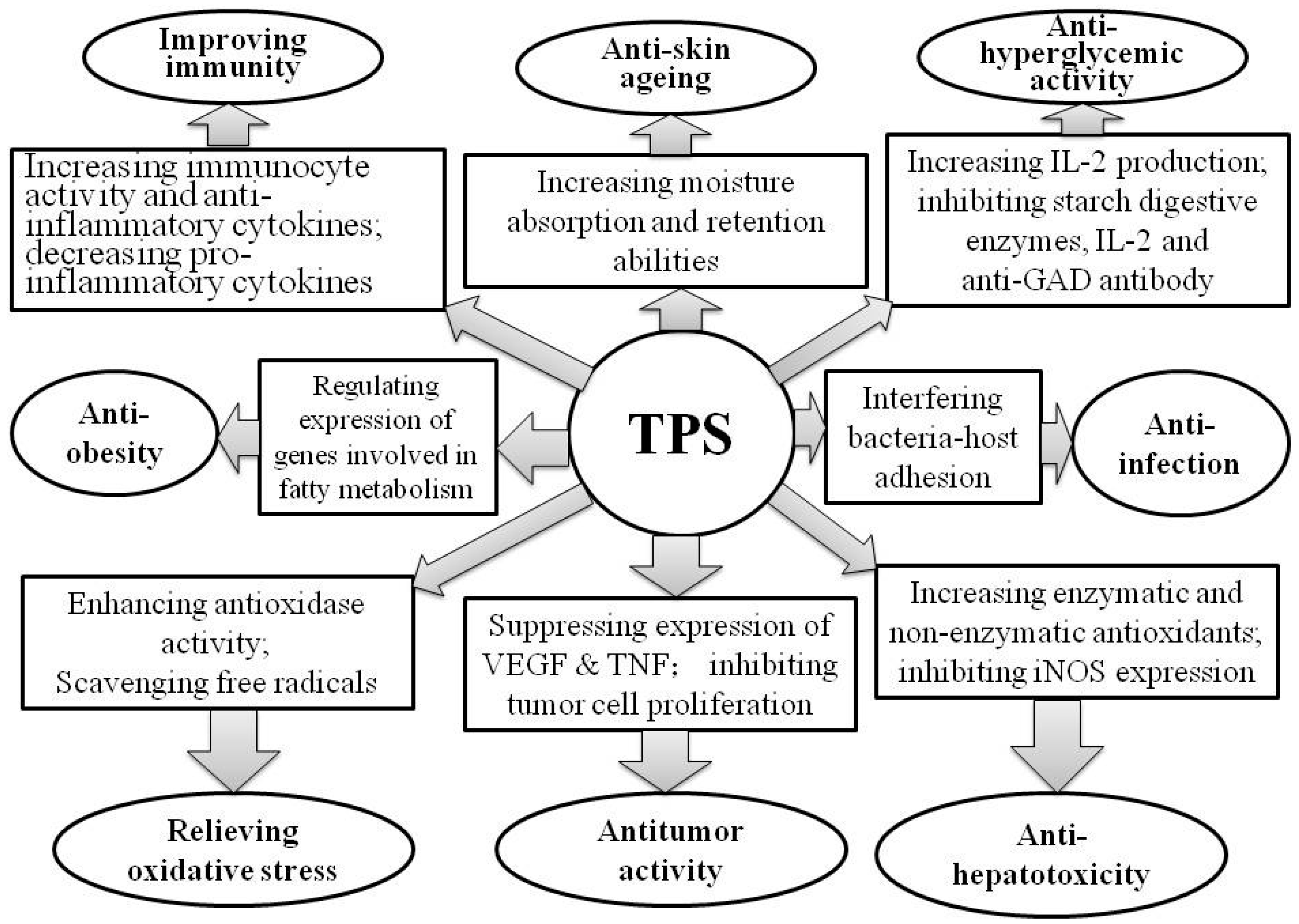

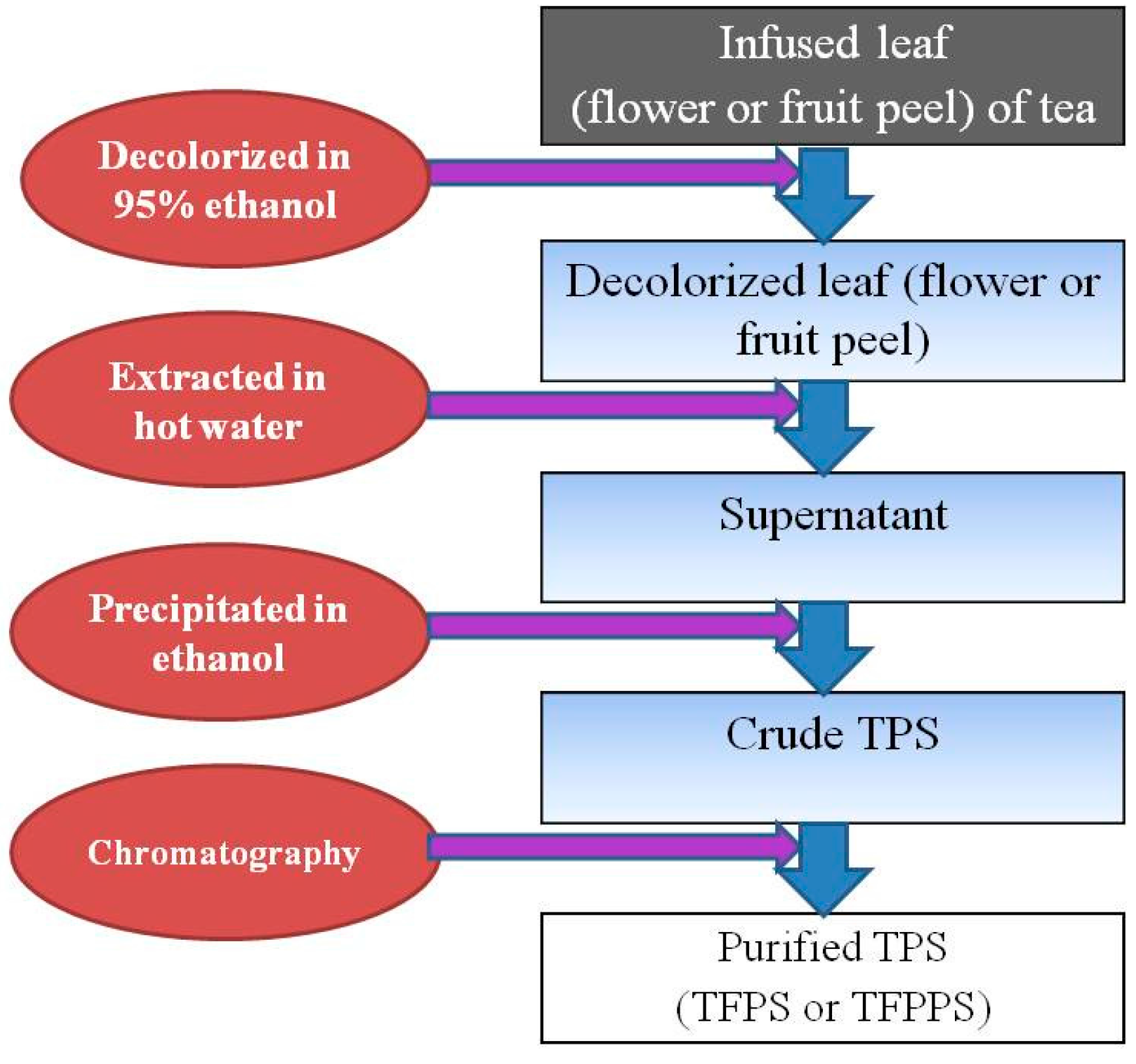{kind=link}
{kind=link}
| Preparation Method | Raw Materials | Major Components | MW Distribution | Reference |
|---|---|---|---|---|
| Separated by DEAE-cellulose column and dried by freeze drying | Green tea | TPS was composed of Rha, Rib, Ara, Xyl, Man, Glu, Gal in molar contents of 53.92, 13.07, 94.56, 38.61, 24.75, 85.38, 124.50 mM. | TPS had a largely undispersed MW distribution. | Guo et al., 2011 [4]. |
| Extracted in hot water and precipitated in ethanol, then lyophilized | Oolong teas Tieguanyin (TTPS), Fenghuangdancong (FTPS) and Dahongpao (DTPS) | TTPS contained 5.57% protein, 27.56% neutral sugars and 45.89% uronic acid; FTPS contained 7.68% protein, 16.71% neutral sugars and 56.46% uronic acid; DTPS contained 9.30% protein, 20.74% neutral sugars and 63.11% uronic acid | 7–817 kDa for TTPS, 14–930 kDa for FTPS and 42–264 kDa for DTPS | Wang et al., 2012 [8] |
| RP-C18 column chromatography | Green tea | Man, Rib, Rha, GulA, GalA, Glu, Xyl, Gal And Ara in molar contents of 16.3, 10.3, 47.1, 5.6, 24.0, 128.4, 25.0, 101.4 and 71.1 μM | Lv et al., 2009 [10] | |
| Extracted in hot water and precipitated in ethanol, then dried by freeze drying (TPS-F), spray drying (TPS-S), vacuum drying (TPS-V) and microwave-vacuum drying (TPS-M) | Green tea | Total sugar content was 42.71% in TPS-F, 42.01% in TPS-V, 41.16% in TPS-S and 41.08% in TPS-M. Protein content was 5.5% in TPS-F, 8.34% in TPS-V, 4.10% in TPS-S and 5.75% in TPS-M. Polyphenols content was 10.41% in TPS-F, 13.22% in TPS-V, 9.56% in TPS-S and 10.78% in TPS-M | 3.3–952.5 kDa for TPS-F, 3.4–910.9 kDa for TPS-V, 3.3–969.1 kDa for TPS-S and 3.5–915.7 kDa for TPS-M | Wang et al., 2013 [12] |
| Hot water extraction and followed by 40% (TPS1) and 70% (TPS2) ethanol precipitation | Green tea | TPS1 showed stronger phagocytosis-enhancing activity than TPS2. | TPS1-2a had MW 20 kDa and TPS1-2b had a MW 22 kDa. | Wang et al., 2014 [13]. |
| Anion-exchange chromatography | Green tea, | Water-soluble TPS was separated into 5 fractions among which fraction C was further separated into FC-1 and FC-2. FC-1 is a novel polysaccharide which is composed of Rha, Ara, Man, Glc and Gal in the ratio of 12.57:22.95:4.4:39.34:20.77, with 8% GalA. | FC-1 has a MW about 60 kDa. | Wang et al., 2006 [14] |
| Extraction in hot water and precipitated using 95% ethanol | Green tea | TPS was composed of Ara, Xyl, Fuc, Glc, and Gal, with extraction yield 2.3–5.8 g per kg dry tea. | 107–110 kDa | Wang et al., 2001 [19] |
| Hot water extraction and followed by 95% ethanol precipitation | Green tea (GTPS), Oolong tea (OTPS), Black tea (BTPS) | Protein content was 32.6% in GTPS, 32.7% in OTPS and 38.0% in BTPS. Uronic acid content was 20.8% in GTPS, 25.5% in OTPS and 16.1% in BTPS. Neutral sugar content was 27.3% in GTPS, 26.5% in OTPS and 18.8% in BTPS. Extraction yield was 4.0% for GTPS, 4.6% for OTPS and 4.2% for BTPS. | 9.2–251.5 KDa for GTPS ; 5.3–100.9 kDa for OTPS and 3.8–32.7 KDa for BTPS | Chen et al., 2009 [28] |
| Absorbent chromatography and ion exchange chromatography | Green tea | TPS-1 was composed of Ara, Rib, Xyl, Glc, Gal and Man, with 30.0% uronic acid and 2.8% protein. TPS-2 was composed of Ara, Rib, Xyl, Glc and Man with 47.6% uronic acid and 3.8% protein. TPS-3 was composed of Ara, Rib, Xyl, Glc and Gal with 51.8% uronic acid and 4.0% protein. | 268 kDa for TPS-1, 118 kDa for TPS-2 and 42 kDa for TPS-3. | Chen et al. 2008 [37] |
| Extraction in hot water and precipitated in 30%–60% final ethanol | Green tea (GTPS), Dark tea (DTPS), Oolong tea (OTPS), White tea (WTPS) and Black tea (BTPS) | Sulfuric radical content was 0.37%–0.91% in GTPS, 0.55%–1.78% in DTPS, 2.77%–3.44% in OTPS, 0.99%–1.21% in WTPS and 1.65%–2.13% in BTPS. Uronic acid content was 3.40%–2.18% in GTPS, 9.90%–6.05% in DTPS, 6.42%–3.71% in OTPS, 56.51%–2.03% in WTPS and 3.21%–1.95% in BTPS | Zhao et al., 2014 [49] | |
| Extraction in hot water and precipitated using 95% ethanol | Tea flower (TFPS) | TFPS-1 was composed of Glc:Xyl:Rha:Gal in ratio of 1.0:1.2:0.81:0.98; TFPS-2 comprised Glc:Xyl:Rha:Ara in ratio of 1.0:0.76:2.3:2.3. | 167.5 kDa for TFPS-1 and 10.1 kDa for TFPS-2 | Han et al., 2011 [85] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, L.-L.; Fu, Q.-Y.; Xiang, L.-P.; Zheng, X.-Q.; Lu, J.-L.; Ye, J.-H.; Li, Q.-S.; Polito, C.A.; Liang, Y.-R. Tea Polysaccharides and Their Bioactivities. Molecules 2016, 21, 1449. https://doi.org/10.3390/molecules21111449
Du L-L, Fu Q-Y, Xiang L-P, Zheng X-Q, Lu J-L, Ye J-H, Li Q-S, Polito CA, Liang Y-R. Tea Polysaccharides and Their Bioactivities. Molecules. 2016; 21(11):1449. https://doi.org/10.3390/molecules21111449
Chicago/Turabian StyleDu, Ling-Ling, Qiu-Yue Fu, Li-Ping Xiang, Xin-Qiang Zheng, Jian-Liang Lu, Jian-Hui Ye, Qing-Sheng Li, Curt Anthony Polito, and Yue-Rong Liang. 2016. "Tea Polysaccharides and Their Bioactivities" Molecules 21, no. 11: 1449. https://doi.org/10.3390/molecules21111449
APA StyleDu, L.-L., Fu, Q.-Y., Xiang, L.-P., Zheng, X.-Q., Lu, J.-L., Ye, J.-H., Li, Q.-S., Polito, C. A., & Liang, Y.-R. (2016). Tea Polysaccharides and Their Bioactivities. Molecules, 21(11), 1449. https://doi.org/10.3390/molecules21111449