
Oleanolic Acid, a Compound Present in Grapes and Olives, Protects against Genotoxicity in Human Mammary Epithelial Cells



Abstract
:
1. Introduction
2. Results
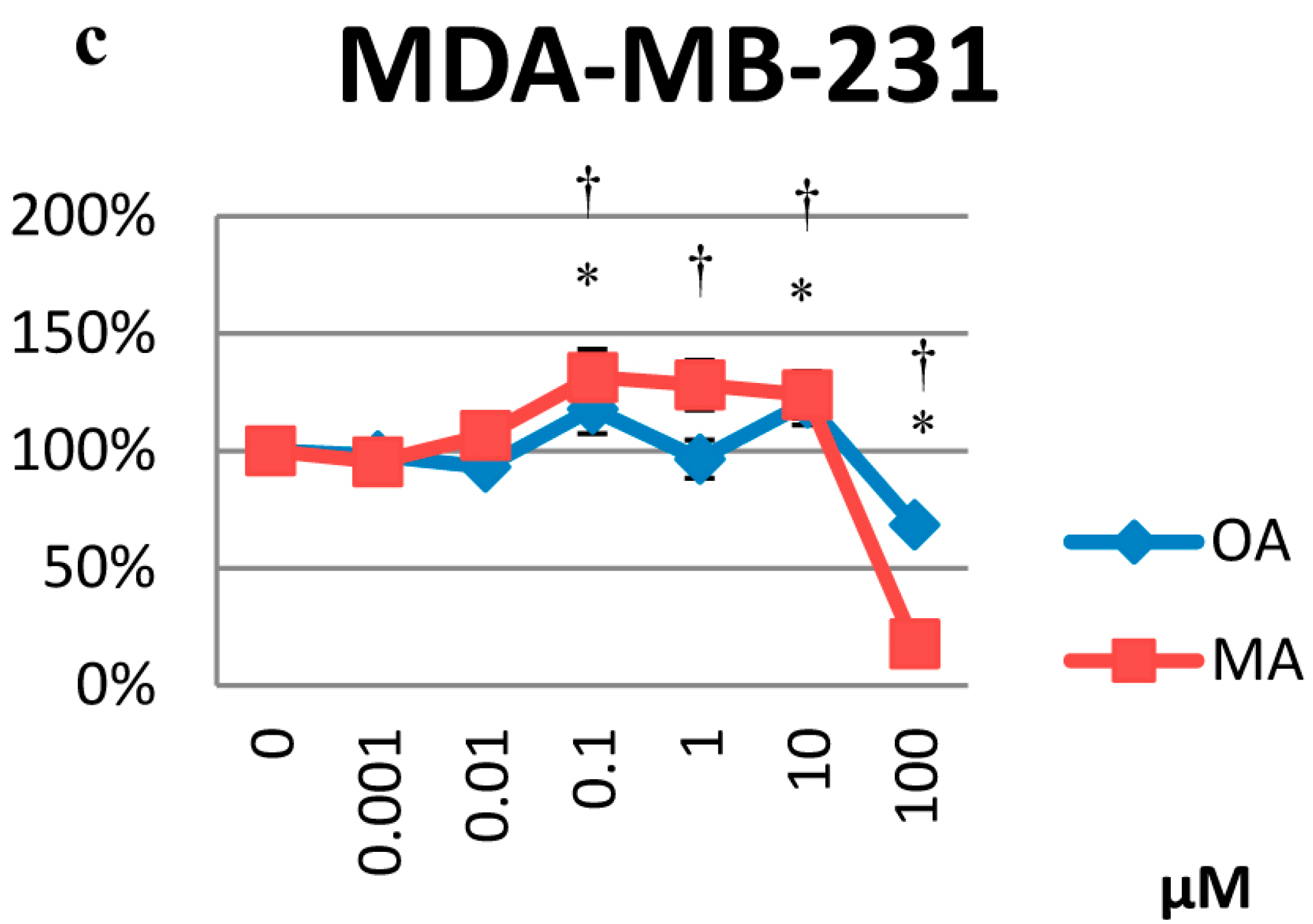
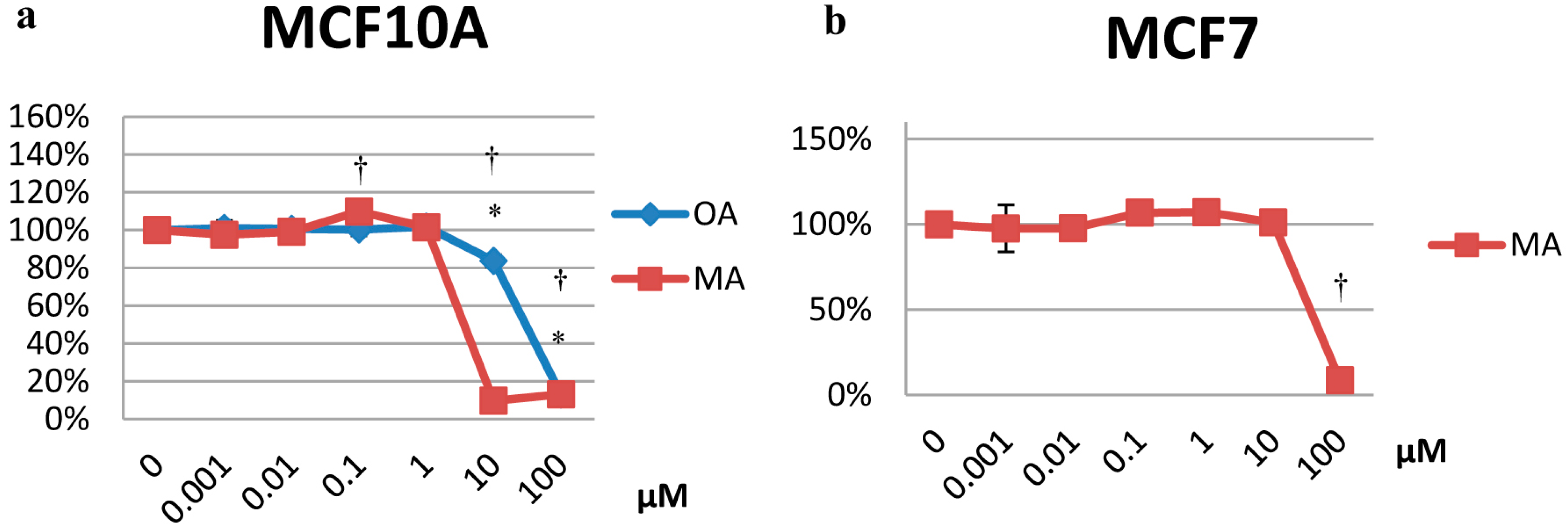
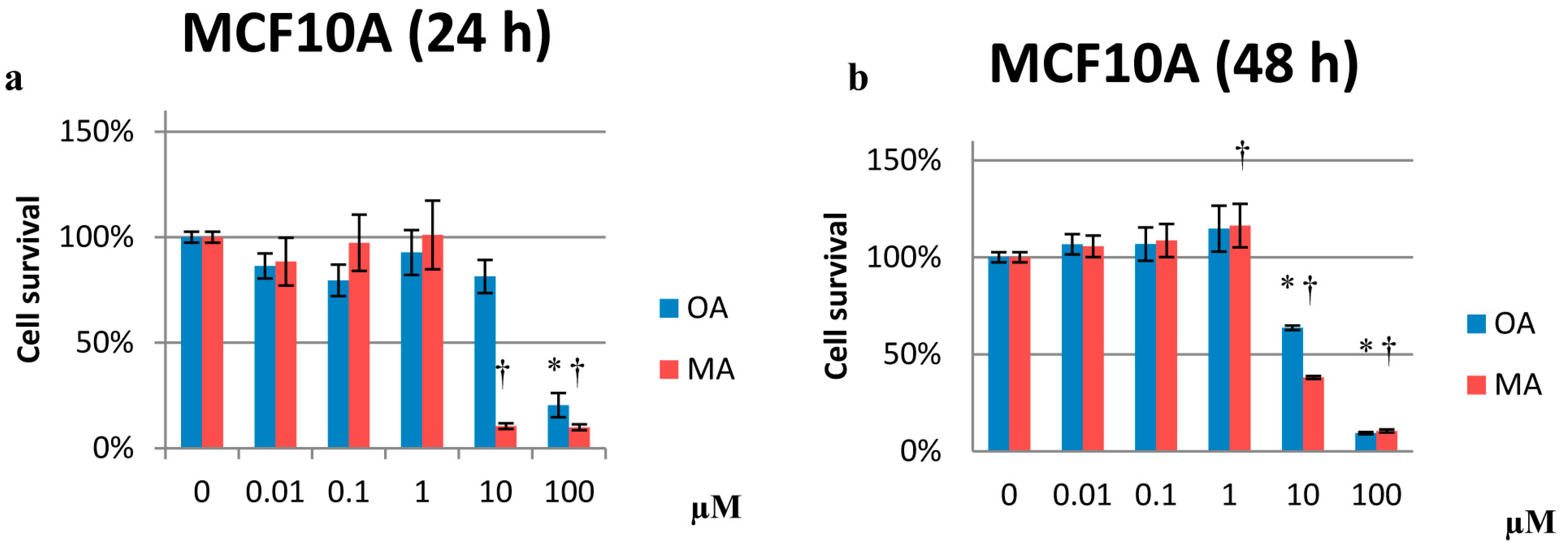
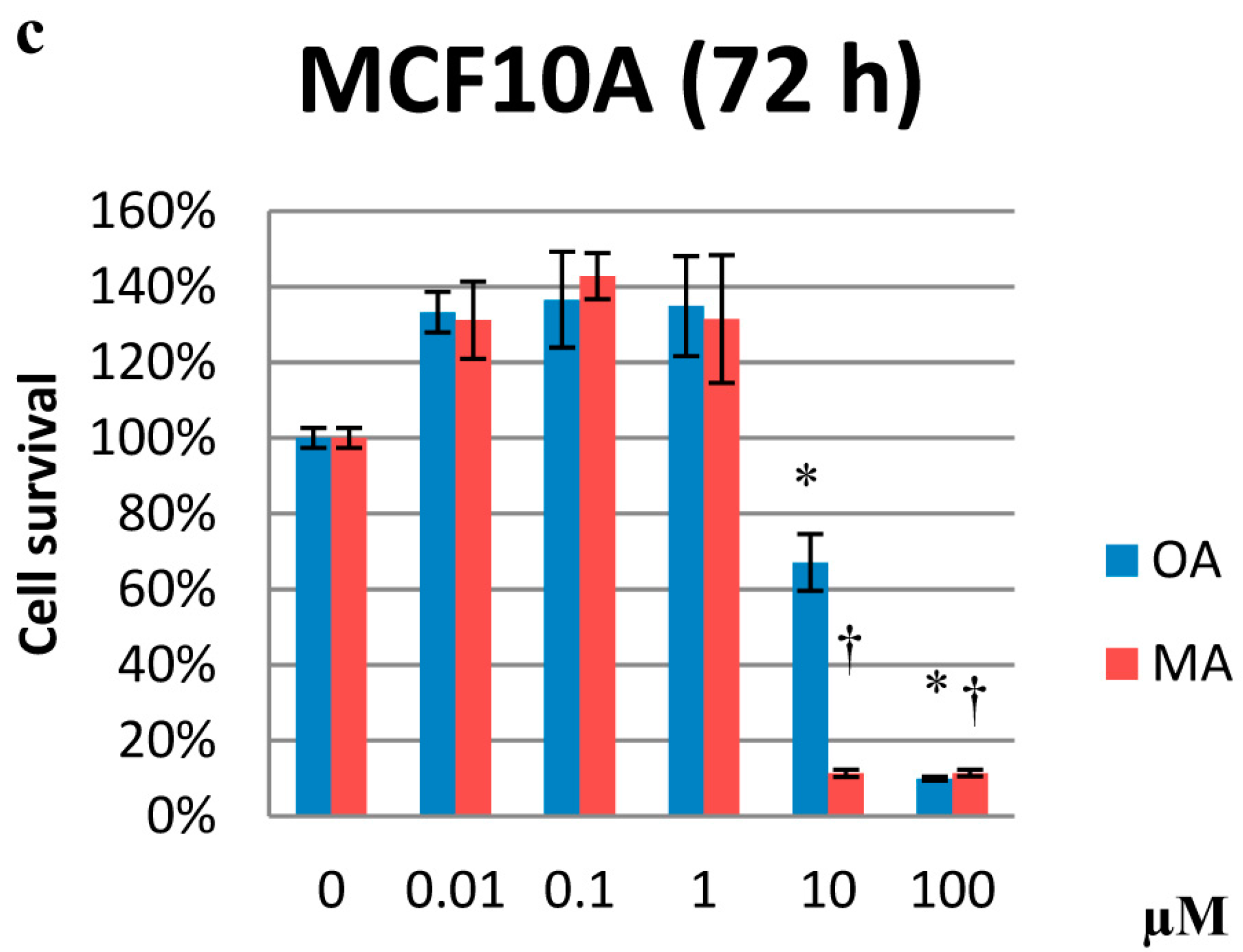
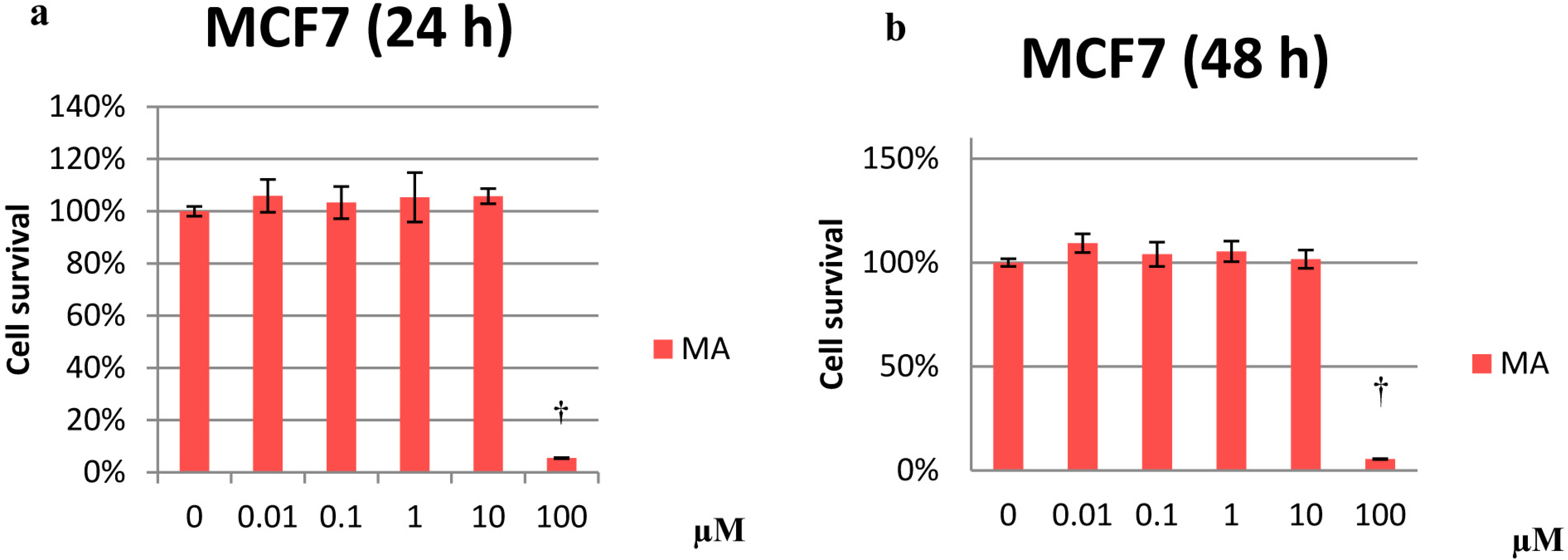
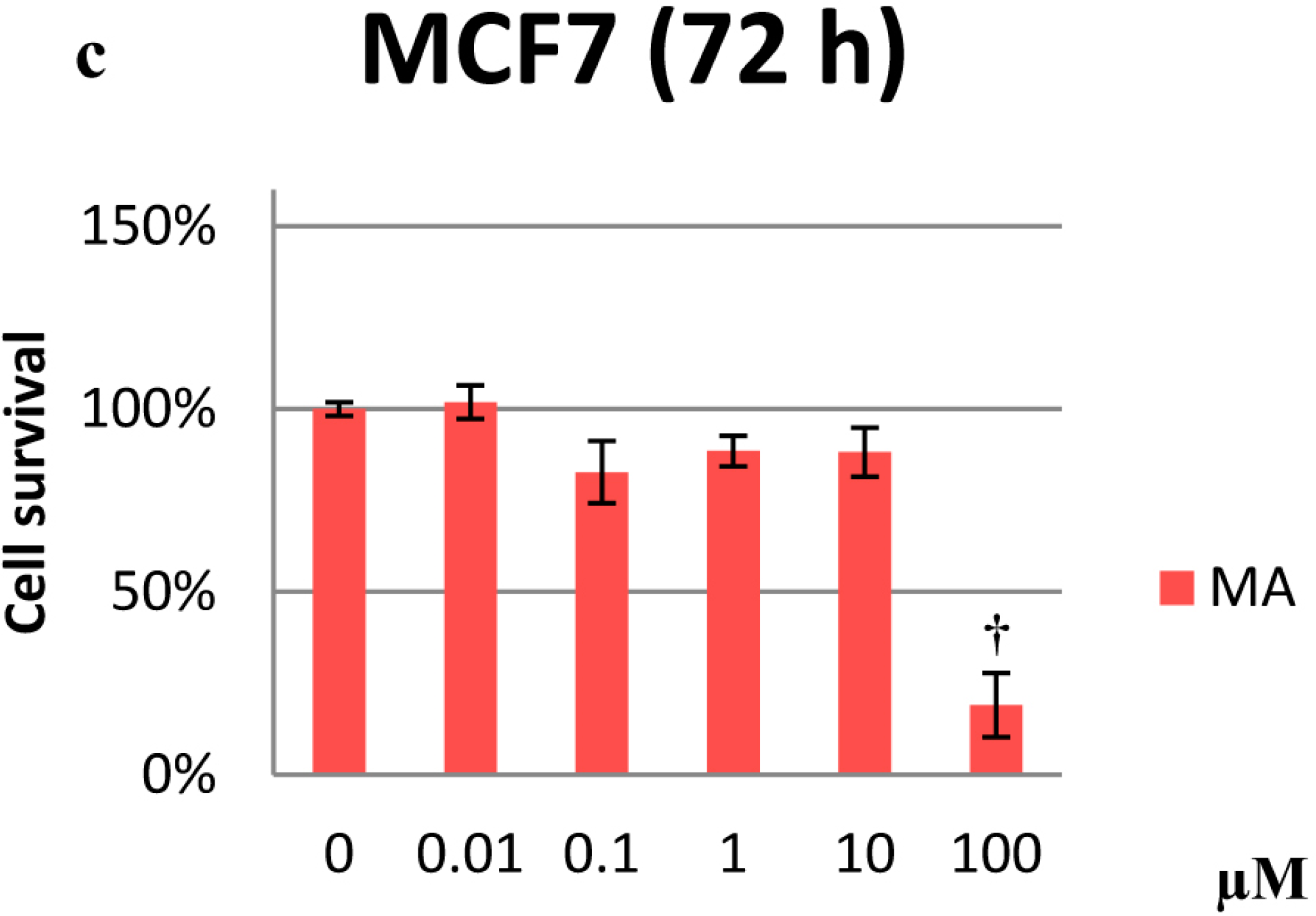
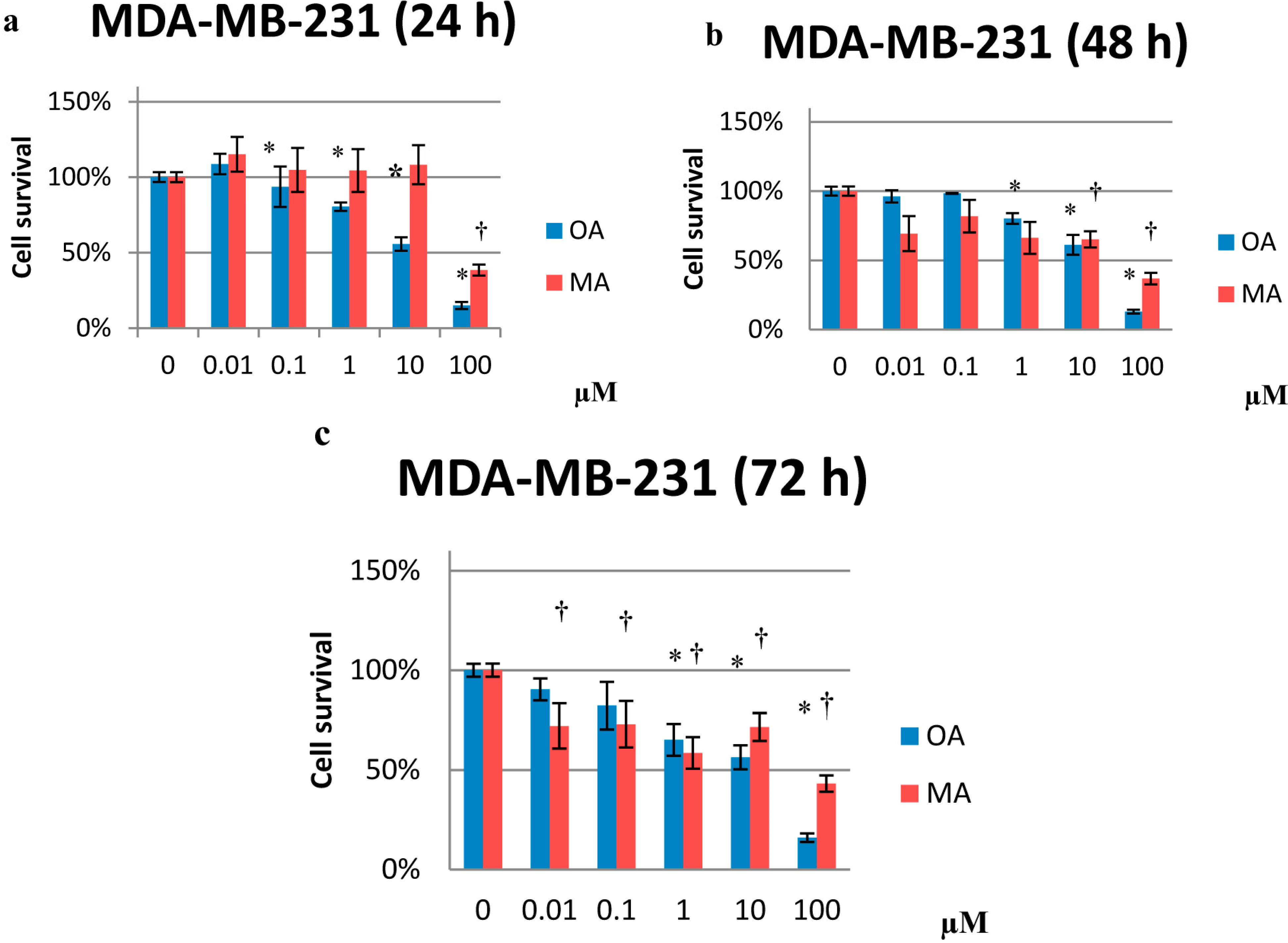
2.1. Cytotoxicity Effects

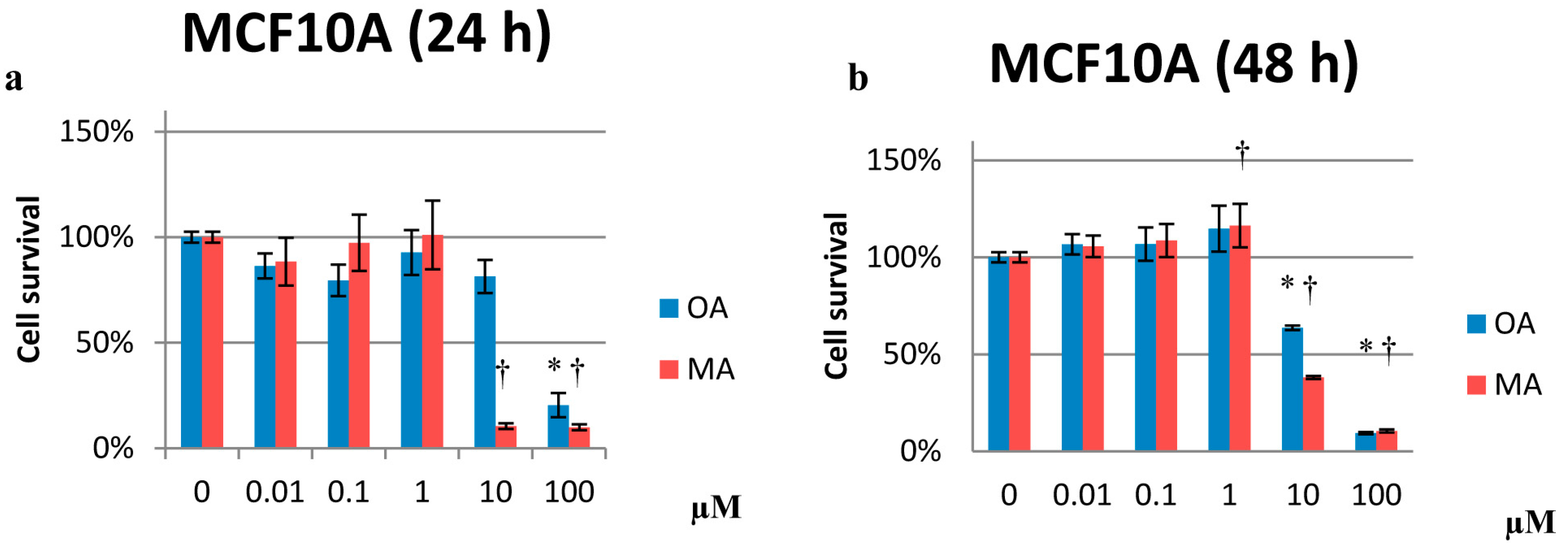
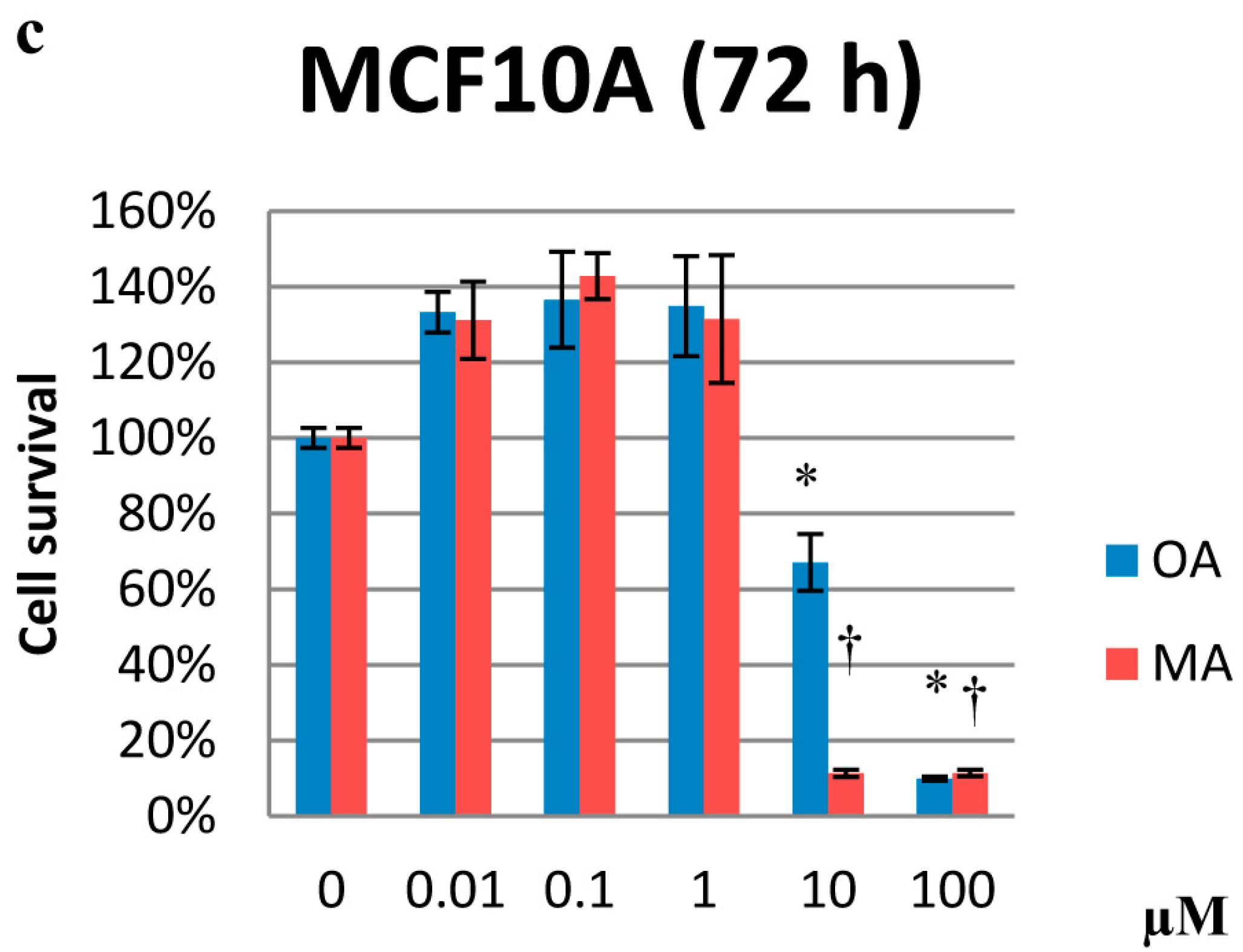
2.2. Effects on Proliferation
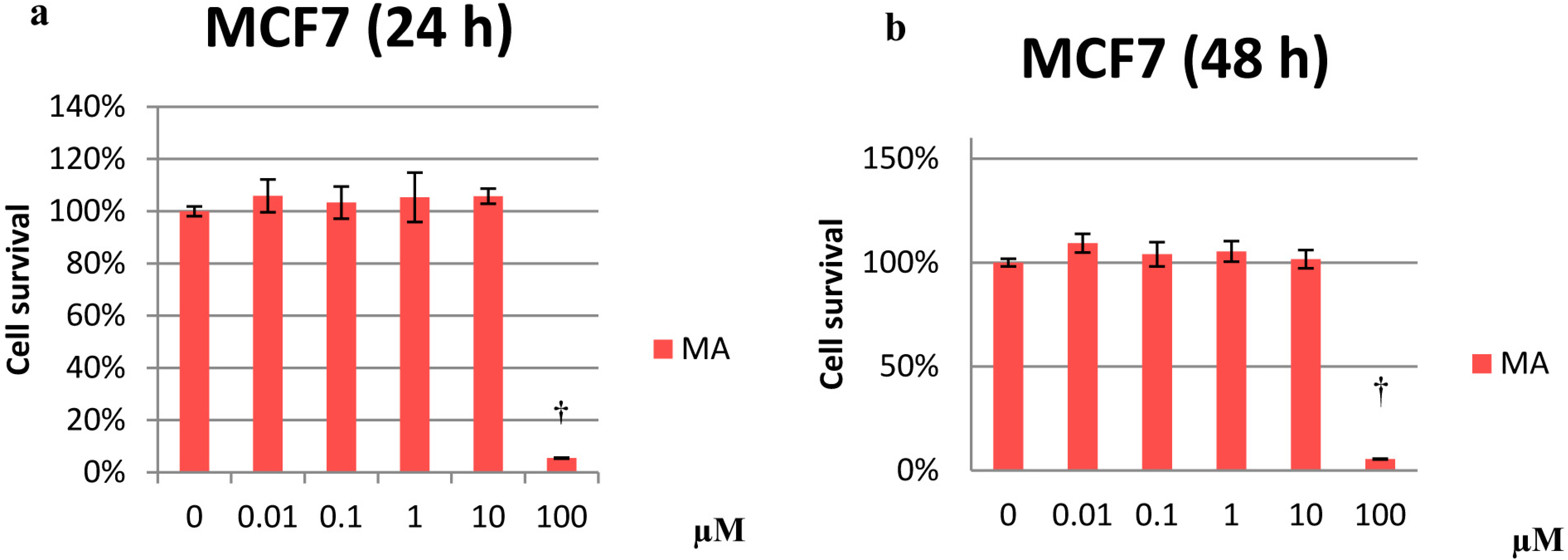
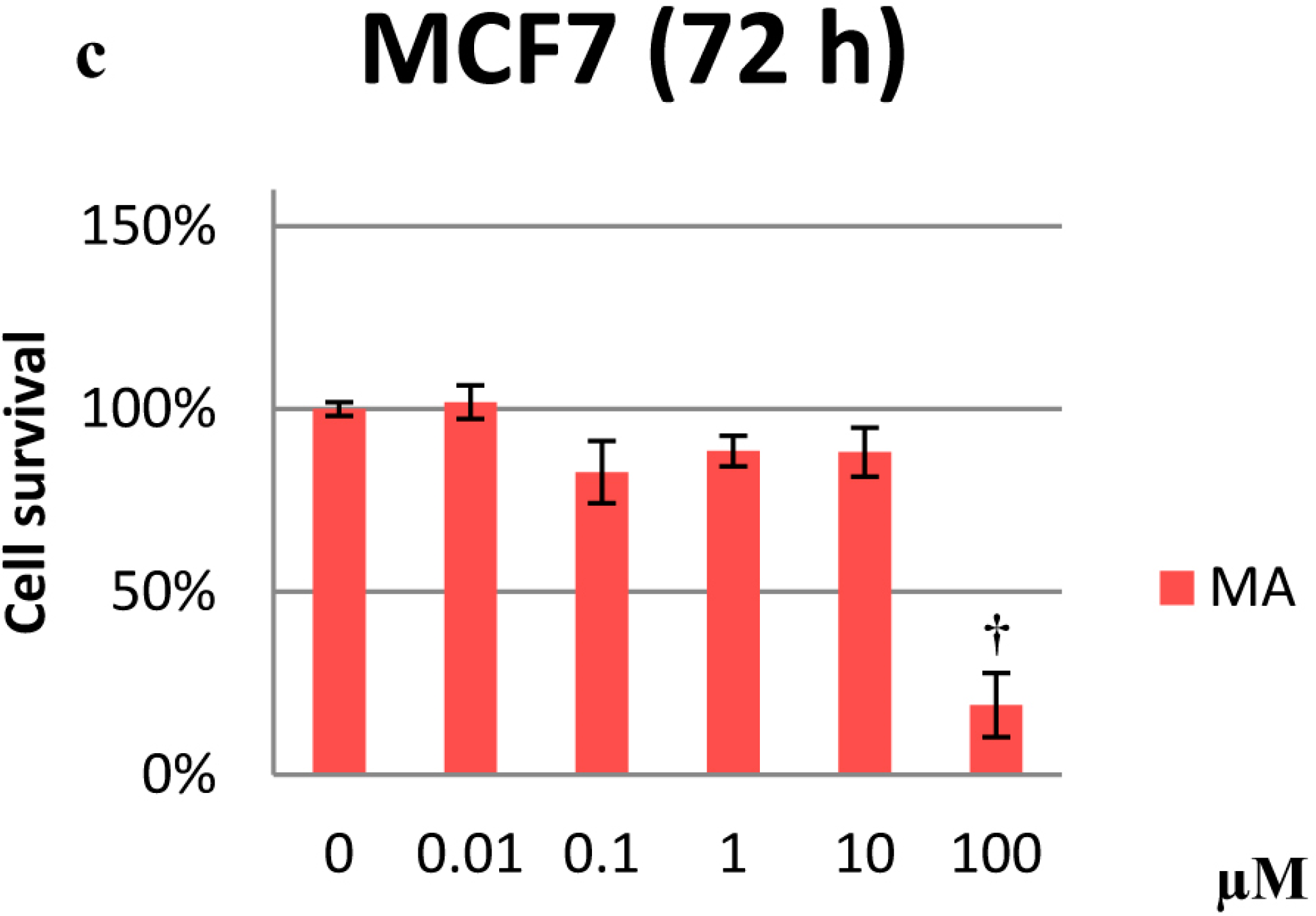

2.3. Effects on the Cell Cycle

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Percentage of Cells | ||||||||
|---|---|---|---|---|---|---|---|---|
| MDA-MB-231 | MCF10A | |||||||
| Sub-G0/G1 | G0/G1 | S | G2/M | Sub-G0/G1 | G0/G1 | S | G2/M | |
| Control | 0.84 ± 0.16 | 65.38 ± 1.25 | 16.18 ± 1.08 | 16.44 ± 2.05 | 0.39 ± 0.15 | 58.22 ± 2.93 | 15.90 ± 1.43 | 24.54 ± 1.51 |
| OA 0.1 µM | 0.75 ± 0.25 | 59.04 ± 2.99 * | 17.31 ± 1.58 | 21.89 ± 2.09 * | 0.54 ± 0.27 | 56.86 ± 4.60 | 16.54 ± 2.83 | 24. 27 ± 1.18 |
| OA 1 µM | 0.78 ± 0.25 | 59.73 ± 1.98 | 17.60 ± 1.66 | 20.76 ± 1.54 | 0.37 ± 0.15 | 58.23 ± 3.63 | 15.68 ± 1.86 | 25.08 ± 1.23 |
| OA 10 µM | 0.76 ± 0.37 | 61.06 ± 1.85 | 15.04 ± 1.46 | 21.36 ± 0.91 * | 0.84 ± 0. 25 | 71.45 ± 6.63 * | 11.58 ± 2.54 | 15.14 ± 4.22 * |
| MA 0.1 µM | 1.42 ± 0.49 | 61.88 ± 0.73 | 16.86 ± 2.25 | 19.28 ± 1.61 | 0.37 ± 0.13 | 58.77 ± 1.75 | 15.17 ± 0.20 | 24.43 ± 2.26 |
| MA 1 µM | 0.72 ± 0.49 | 61.90 ± 0.52 | 16.66 ± 1.13 | 19.59 ± 0.84 | 0.59 ± 0.16 | 58.81 ± 3.82 | 16.58 ± 2.65 | 22.92 ± 1.88 |
| MA 10 µM | 0.72 ± 0.01 | 62.36 ± 0.65 | 16.56 ± 1.12 | 19.75 ± 1.64 | 64.68 ± 1.92 * | 21.96 ± 1.82 * | 8.01 ± 1.15* | 4.56 ± 1.24 * |
2.4. Analysis of Apoptosis
| Percentage of Cells | ||||||
|---|---|---|---|---|---|---|
| MDA-MB-231 | MCF10A | |||||
| Live | Apoptotic | Death | Live | Apoptotic | Death | |
| Control | 87.64 ± 3.16 | 8.92 ± 2.15 | 1.33 ± 0.48 | 92.43 ± 1.43 | 5.92 ± 1.40 | 1.63 ± 0.53 |
| OA 0.1 µM | 90.66 ± 4.28 | 8.43 ± 4.04 | 0.90 ± 0.36 | 94.43 ± 0.71 | 3.57 ± 1.31 | 1.97 ± 0.61 |
| OA 1 µM | 86.72 ± 3.27 | 11.83 ± 3.28 | 1.43 ± 0.24 | 94.91 ± 0.74 | 2.16 ± 0.79 | 2.90 ± 1.00 |
| OA 10 µM | 88.22 ± 2.78 | 10.20 ± 3.36 | 1.56 ± 0.61 | 70.40 ± 16.09 | 17.18 ± 8.22 * | 12.41 ± 7.89 |
| MA 0.1 µM | 90.81 ± 3.29 | 8.12 ± 2.65 | 1.05 ± 0.67 | 92.38 ± 2.01 | 6.35 ± 2.30 | 1.26 ± 0.33 |
| MA 1 µM | 89.43 ± 5.38 | 7.70 ± 3.06 | 2.85 ± 2.33 | 92.35 ± 1.30 | 5.80 ± 1.84 | 1.83 ± 0.63 |
| MA 10 µM | 88.86 ± 2.41 | 10.13 ± 2.16 | 0.98 ± 0.30 | 5.64 ± 2.31 | 78.17 ± 8.92 * | 16.17 ± 7.01 |
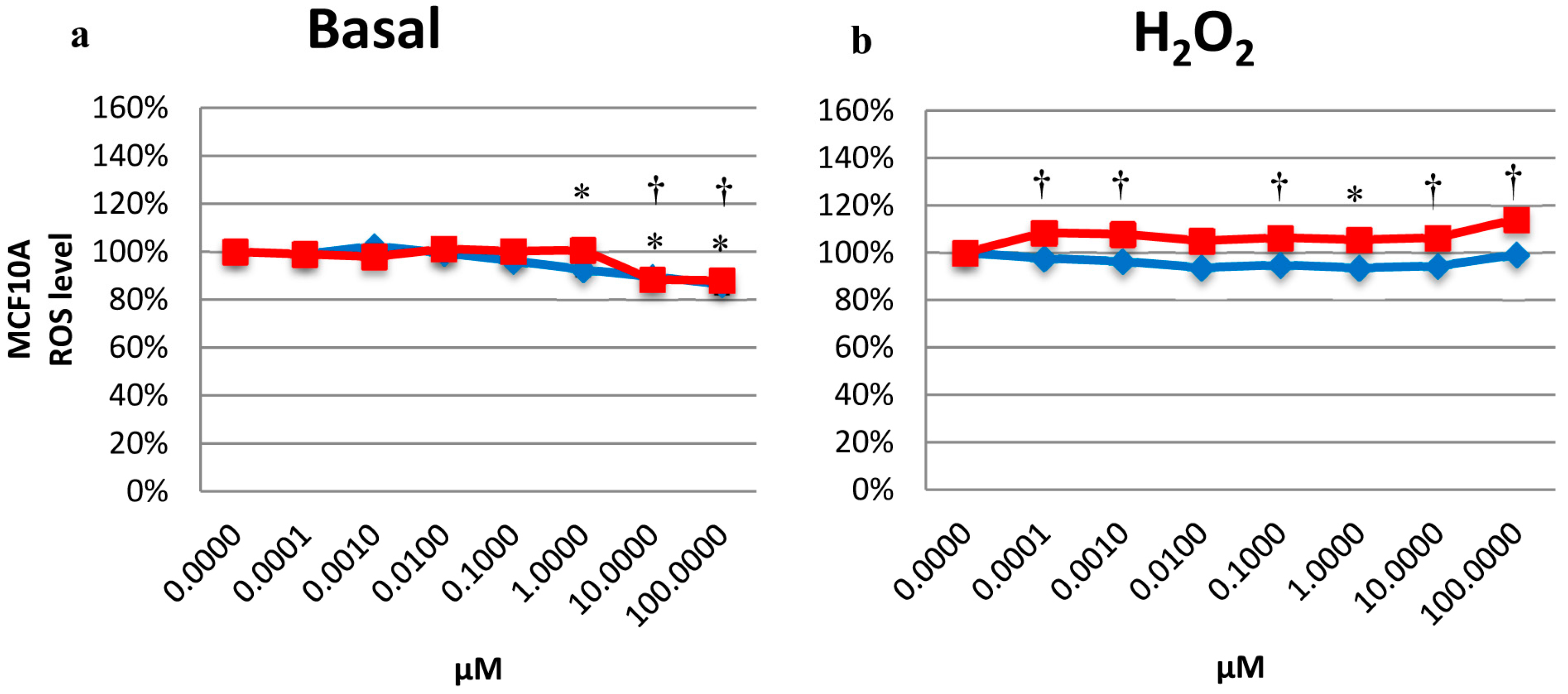
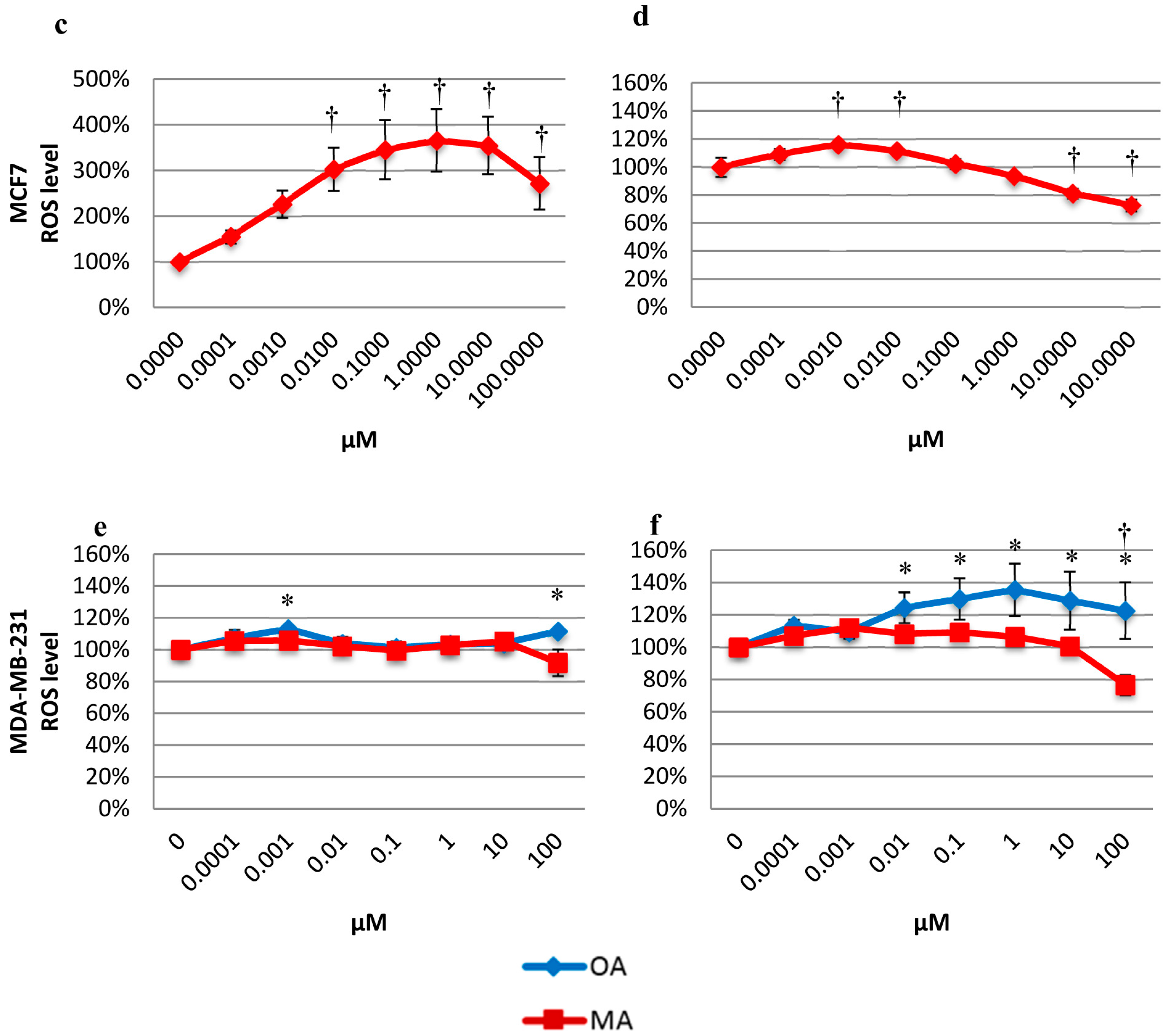
2.5. Effects on the Intracellular ROS Level
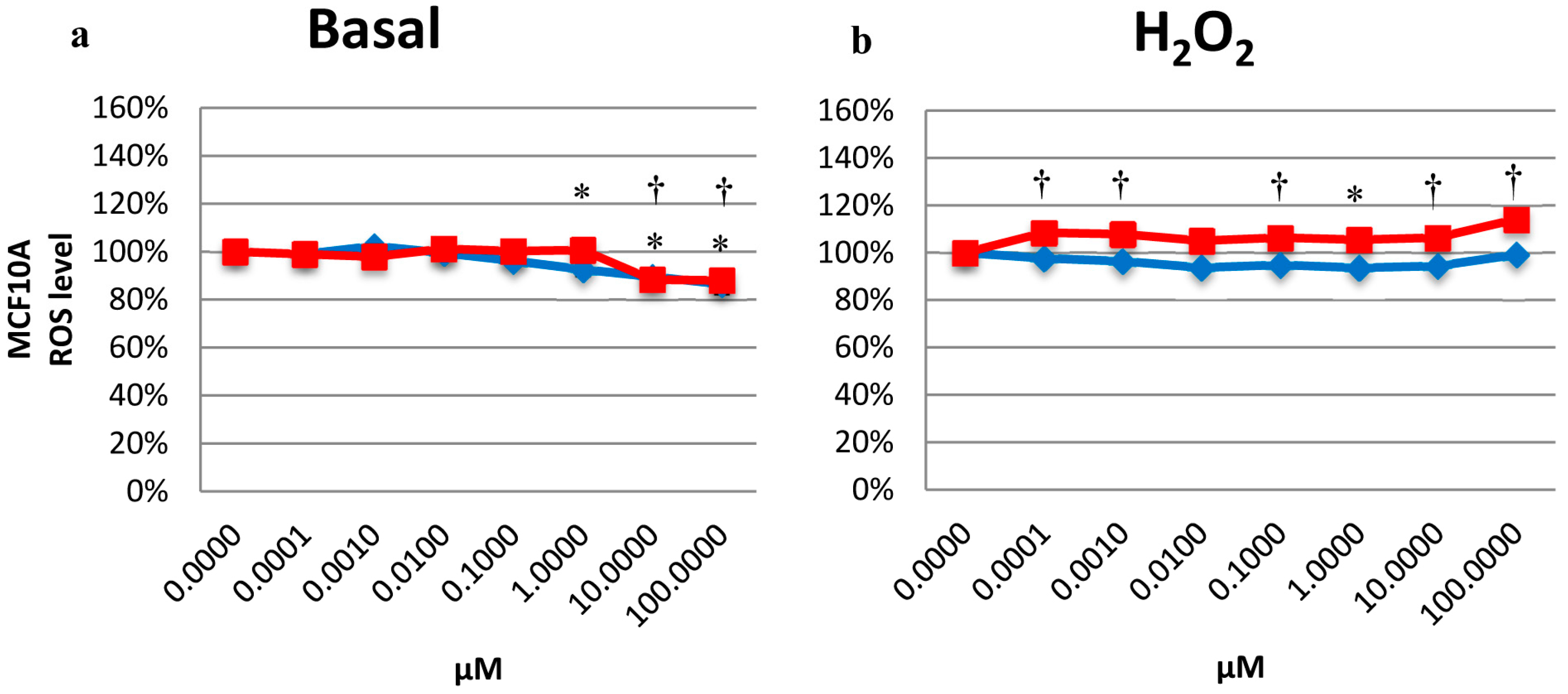
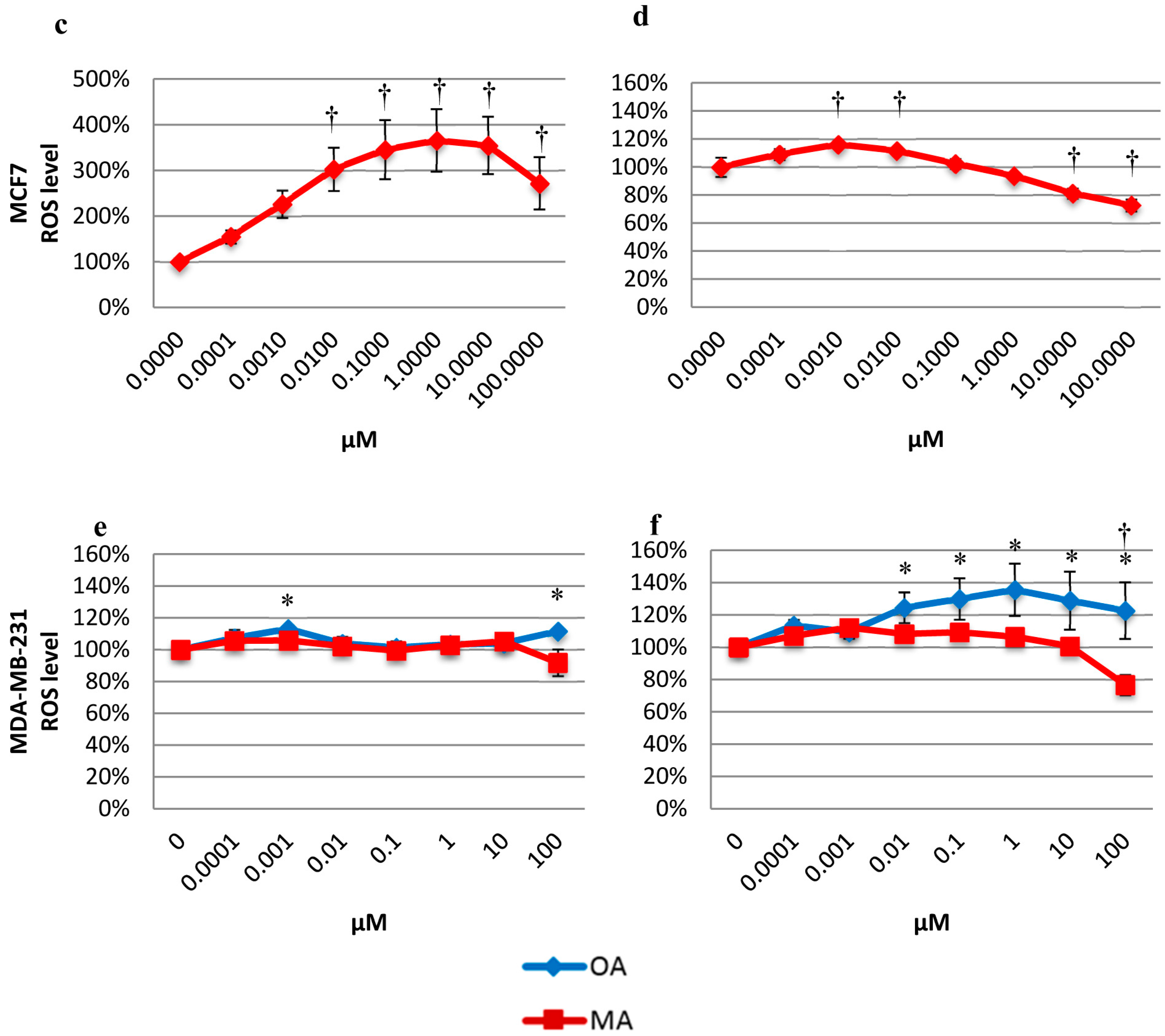
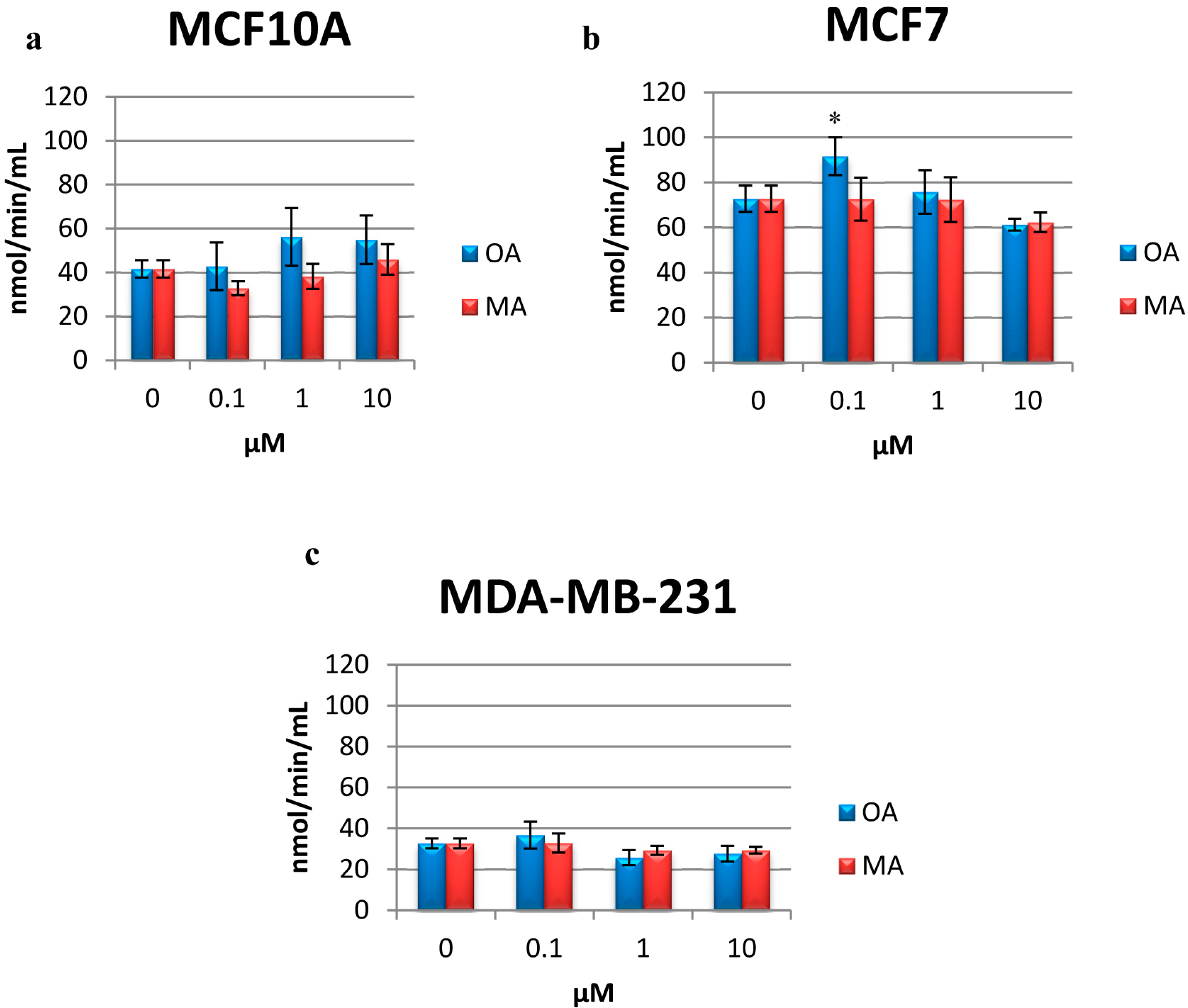
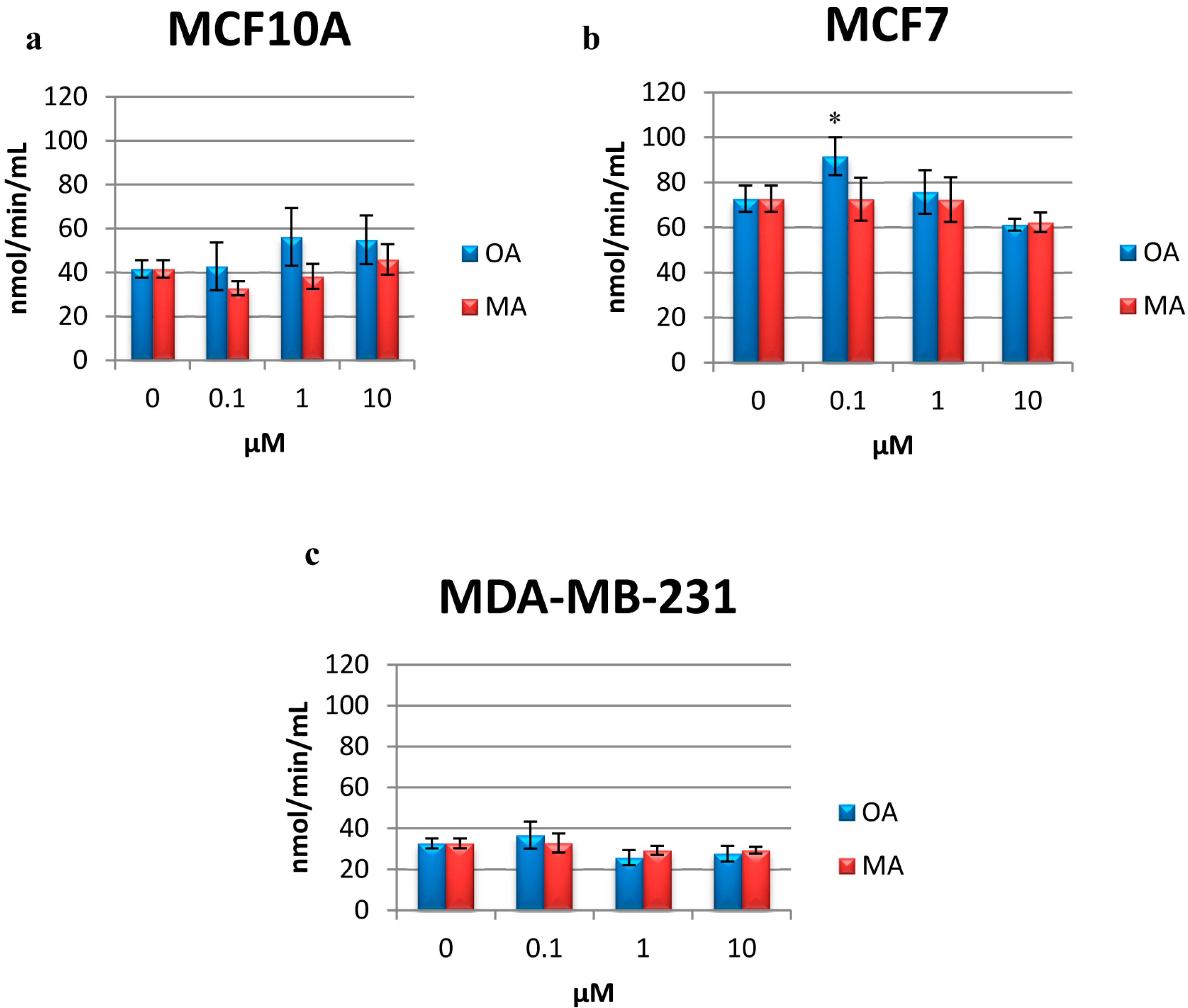
2.6. Determination of CAT Activity
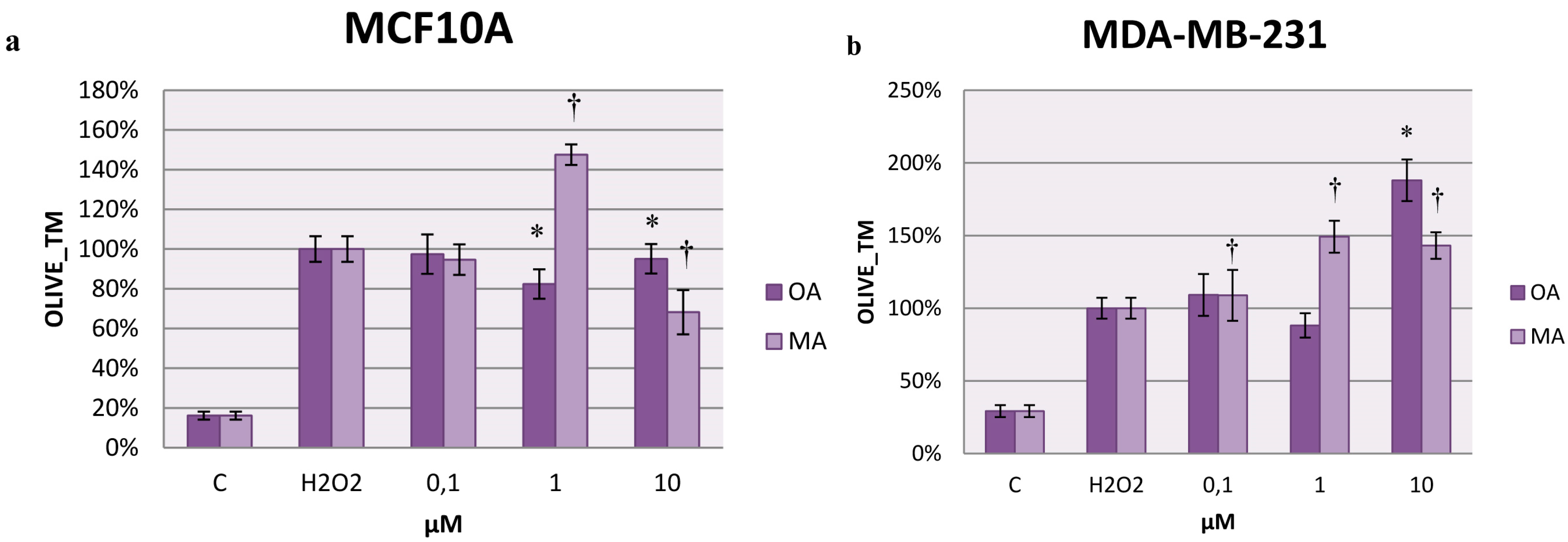
2.7. Effects on H2O2-Induced DNA Damage
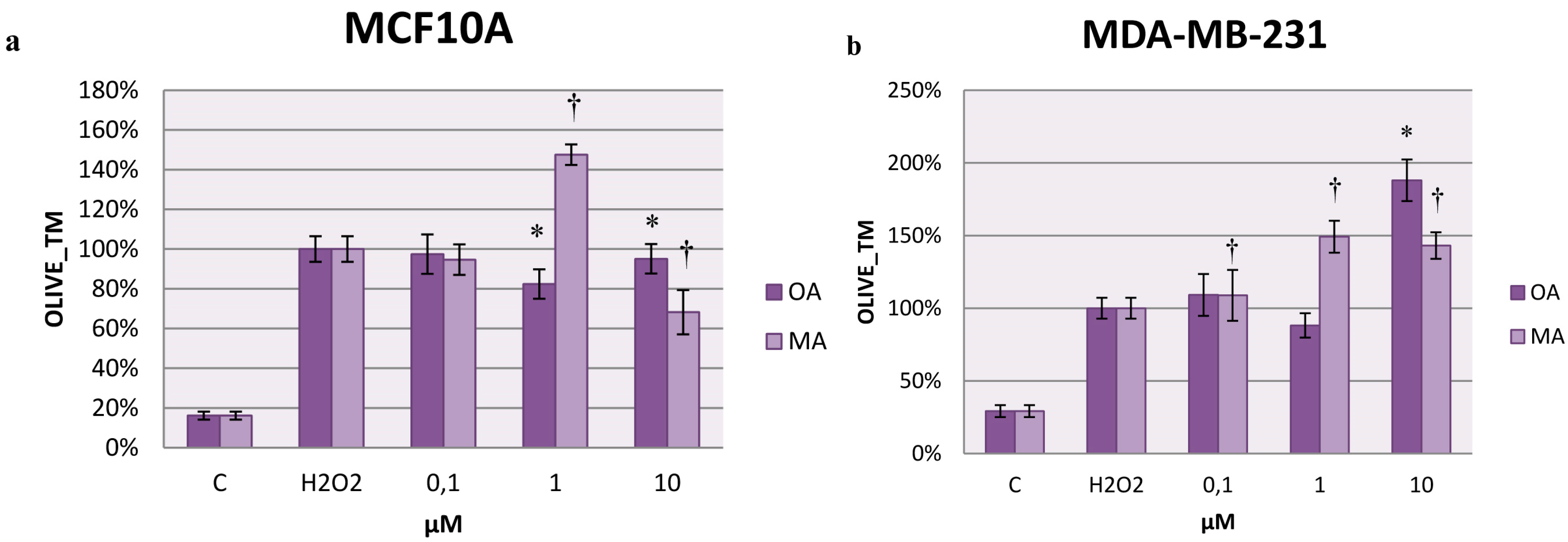
3. Discussion
4. Experimental Section
4.1. Chemicals
4.2. Cell Culture and Treatments
4.3. Cytotoxicity Assay
4.4. Cell Proliferation Assay
4.5. Cell Cycle Assay
4.6. Apoptosis Assay
4.7. Detection of Intracellular Reactive Oxygen Species
4.8. Determination of Catalase (CAT) Activity
4.9. Alkaline Single-Cell Gel Electrophoresis (Comet Assay)
4.10. Slide Scoring and Analysis
4.11. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, Y.; Jayaprakasam, B.; Seeram, N.P.; Olson, L.K.; DeWitt, D.; Nair, M.G. Insulin Secretion and Cyclooxygenase Enzyme Inhibition by Cabernet Sauvignon Grape Skin Compounds. J. Agric. Food Chem. 2004, 52, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Glensk, M.; Glinski, J.A.; Wlodarczyk, M.; Stefanowicz, P. Determination of Ursolic and Oleanolic Acid in Sambuci Fructus. Chem. Biodivers. 2014, 11, 1939–1944. [Google Scholar] [CrossRef] [PubMed]
- Szakiel, A.; Mroczek, A. Distribution of Triterpene Acids and their Derivatives in Organs of Cowberry (Vaccinium vitis-idaea L.) Plant. Acta Biochim. Pol. 2007, 54, 733–740. [Google Scholar] [PubMed]
- Szakiel, A.; Paczkowski, C.; Koivuniemi, H.; Huttunen, S. Comparison of the Triterpenoid Content of Berries and Leaves of Lingonberry Vaccinium vitis-idaea from Finland and Poland. J. Agric. Food Chem. 2012, 60, 4994–5002. [Google Scholar] [CrossRef] [PubMed]
- Allouche, Y.; Jimenez, A.; Uceda, M.; Aguilera, M.P.; Gaforio, J.J.; Beltran, G. Triterpenic Content and Chemometric Analysis of Virgin Olive Oils from Forty Olive Cultivars. J. Agric. Food Chem. 2009, 57, 3604–3610. [Google Scholar] [CrossRef] [PubMed]
- Pensec, F.; Paczkowski, C.; Grabarczyk, M.; Wozniak, A.; Benard-Gellon, M.; Bertsch, C.; Chong, J.; Szakiel, A. Changes in the Triterpenoid Content of Cuticular Waxes during Fruit Ripening of Eight Grape (Vitis vinifera) Cultivars Grown in the Upper Rhine Valley. J. Agric. Food Chem. 2014, 62, 7998–8007. [Google Scholar] [CrossRef] [PubMed]
- Yunoki, K.; Sasaki, G.; Tokuji, Y.; Kinoshita, M.; Naito, A.; Aida, K.; Ohnishi, M. Effect of Dietary Wine Pomace Extract and Oleanolic Acid on Plasma Lipids in Rats Fed High-Fat Diet and its DNA Microarray Analysis. J. Agric. Food Chem. 2008, 56, 12052–12058. [Google Scholar] [CrossRef] [PubMed]
- Buckland, G.; Travier, N.; Cottet, V.; Gonzalez, C.A.; Lujan-Barroso, L.; Agudo, A.; Trichopoulou, A.; Lagiou, P.; Trichopoulos, D.; Peeters, P.H.; et al. Adherence to the Mediterranean Diet and Risk of Breast Cancer in the European Prospective Investigation into Cancer and Nutrition Cohort Study. Int. J. Cancer 2013, 132, 2918–2927. [Google Scholar] [CrossRef] [PubMed]
- Patlolla, J.M.; Rao, C.V. Triterpenoids for Cancer Prevention and Treatment: Current Status and Future Prospects. Curr. Pharm. Biotechnol. 2012, 13, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Allouche, Y.; Warleta, F.; Campos, M.; Sanchez-Quesada, C.; Uceda, M.; Beltran, G.; Gaforio, J.J. Antioxidant, Antiproliferative, and Pro-Apoptotic Capacities of Pentacyclic Triterpenes found in the Skin of Olives on MCF-7 Human Breast Cancer Cells and their Effects on DNA Damage. J. Agric. Food Chem. 2011, 59, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Warleta, F.; Campos, M.; Allouche, Y.; Sanchez-Quesada, C.; Ruiz-Mora, J.; Beltran, G.; Gaforio, J.J. Squalene Protects against Oxidative DNA Damage in MCF10A Human Mammary Epithelial Cells but Not in MCF7 and MDA-MB-231 Human Breast Cancer Cells. Food Chem. Toxicol. 2010, 48, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Warleta, F.; Quesada, C.S.; Campos, M.; Allouche, Y.; Beltran, G.; Gaforio, J.J. Hydroxytyrosol Protects Against Oxidative DNA Damage in Human Breast Cells. Nutrients 2011, 3, 839–857. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Quesada, C.; Lopez-Biedma, A.; Gaforio, J.J. The Differential Localization of a Methyl Group Confers a Different Anti-Breast Cancer Activity to Two Triterpenes Present in Olives. Food Funct. 2015, 6, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Allouche, Y.; Beltran, G.; Gaforio, J.J.; Uceda, M.; Mesa, M.D. Antioxidant and Antiatherogenic Activities of Pentacyclic Triterpenic Diols and Acids. Food Chem. Toxicol. 2010, 48, 2885–2890. [Google Scholar] [CrossRef] [PubMed]
- Senthil, S.; Sridevi, M.; Pugalendi, K.V. Cardioprotective Effect of Oleanolic Acid on Isoproterenol-Induced Myocardial Ischemia in Rats. Toxicol. Pathol. 2007, 35, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Graham, V.S.; Lawson, C.; Wheeler-Jones, C.P.; Perona, J.S.; Ruiz-Gutierrez, V.; Botham, K.M. Triacylglycerol-Rich Lipoproteins Derived from Healthy Donors Fed Different Olive Oils Modulate Cytokine Secretion and Cyclooxygenase-2 Expression in Macrophages: The Potential Role of Oleanolic Acid. Eur. J. Nutr. 2012, 51, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yang, Z.; Zhai, C.; Qiu, W.; Li, D.; Yi, Z.; Wang, L.; Tang, J.; Qian, M.; Luo, J.; et al. Maslinic Acid Potentiates the Anti-Tumor Activity of Tumor Necrosis Factor Alpha by Inhibiting NF-kappaB Signaling Pathway. Mol. Cancer. 2010, 9. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Nho, C.W.; Kwon, D.Y.; Kang, Y.H.; Lee, K.W.; Park, J.H. Maslinic Acid Inhibits the Metastatic Capacity of DU145 Human Prostate Cancer Cells: Possible Mediation via Hypoxia-Inducible Factor-1alpha Signalling. Br. J. Nutr. 2013, 109, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bai, H.; Zhang, X.; Liu, J.; Cao, P.; Liao, N.; Zhang, W.; Wang, Z.; Hai, C. Inhibitory Effect of Oleanolic Acid on Hepatocellular Carcinoma via ERK-p53-Mediated Cell Cycle Arrest and Mitochondrial-Dependent Apoptosis. Carcinogenesis 2013, 34, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Liu, M.; Liu, H.; Wang, H.; Wang, F.; Zhang, Y.; Han, L.; Lin, X. Oleanolic Acid Arrests Cell Cycle and Induces Apoptosis via ROS-Mediated Mitochondrial Depolarization and Lysosomal Membrane Permeabilization in Human Pancreatic Cancer Cells. J. Appl. Toxicol. 2013, 33, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Rufino-Palomares, E.E.; Reyes-Zurita, F.J.; Garcia-Salguero, L.; Mokhtari, K.; Medina, P.P.; Lupianez, J.A.; Peragon, J. Maslinic Acid, a Triterpenic Anti-Tumoural Agent, Interferes with Cytoskeleton Protein Expression in HT29 Human Colon-Cancer Cells. J. Proteomics 2013, 83, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Tena, S.; Reyes-Zurita, F.J.; Diaz-Moralli, S.; Vinardell, M.P.; Reed, M.; Garcia-Garcia, F.; Dopazo, J.; Lupianez, J.A.; Gunther, U.; Cascante, M. Maslinic Acid-Enriched Diet Decreases Intestinal Tumorigenesis in Apc(Min/+) Mice through Transcriptomic and Metabolomic Reprogramming. PLoS ONE 2013, 8, e59392. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.Z.; Xuan, Y.Y.; Ruan, S.Q.; Sun, M. Proliferation-Inhibiting and Apoptosis-Inducing Effects of Ursolic Acid and Oleanolic Acid on Multi-Drug Resistance Cancer Cells in Vitro. Chin. J. Integr. Med. 2011, 17, 607–611. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, Y.; Hu, H.; Zhang, Z. In Vitro and in Vivo Antimammary Tumor Activities and Mechanisms of the Apple Total Triterpenoids. J. Agric. Food Chem. 2012, 60, 9430–9436. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, B.; Maurya, R.; Siddiqui, J.A.; Bid, H.K.; Rajendran, S.M.; Yadav, P.P.; Konwar, R. In Vitro Anti-Breast Cancer Activity of Ethanolic Extract of Wrightia Tomentosa: Role of Pro-Apoptotic Effects of Oleanolic Acid and Urosolic Acid. J. Ethnopharmacol. 2012, 142, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Ponou, B.K.; Teponno, R.B.; Ricciutelli, M.; Nguelefack, T.B.; Quassinti, L.; Bramucci, M.; Lupidi, G.; Barboni, L.; Tapondjou, L.A. Novel 3-Oxo- and 3,24-Dinor-2,4-Secooleanane-Type Triterpenes from Terminalia Ivorensis A. Chem. Biodivers. 2011, 8, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Quesada, C.; Lopez-Biedma, A.; Warleta, F.; Campos, M.; Beltran, G.; Gaforio, J.J. Bioactive Properties of the Main Triterpenes found in Olives, Virgin Olive Oil, and Leaves of Olea europaea. J. Agric. Food Chem. 2013, 61, 12173–12182. [Google Scholar] [CrossRef] [PubMed]
- Saeidnia, S.; Abdollahi, M. Antioxidants: Friends or Foe in Prevention or Treatment of Cancer: The Debate of the Century. Toxicol. Appl. Pharmacol. 2013, 271, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A Simple Technique for Quantitation of Low Levels of DNA Damage in Individual Cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Janicke, R.U.; Sprengart, M.L.; Wati, M.R.; Porter, A.G. Caspase-3 is required for DNA Fragmentation and Morphological Changes Associated with Apoptosis. J. Biol. Chem. 1998, 273, 9357–9360. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, S.; Archer, S.L.; Allalunis-Turner, J.; Haromy, A.; Beaulieu, C.; Thompson, R.; Lee, C.T.; Lopaschuk, G.D.; Puttagunta, L.; Bonnet, S.; et al. A Mitochondria-K+ Channel Axis is Suppressed in Cancer and its Normalization Promotes Apoptosis and Inhibits Cancer Growth. Cancer Cell. 2007, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Zurita, F.J.; Rufino-Palomares, E.E.; Lupianez, J.A.; Cascante, M. Maslinic Acid, a Natural Triterpene from Olea europaea L., Induces Apoptosis in HT29 Human Colon-Cancer Cells via the Mitochondrial Apoptotic Pathway. Cancer Lett. 2009, 273, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Zurita, F.J.; Pachon-Pena, G.; Lizarraga, D.; Rufino-Palomares, E.E.; Cascante, M.; Lupianez, J.A. The Natural Triterpene Maslinic Acid Induces Apoptosis in HT29 Colon Cancer Cells by a JNK-p53-Dependent Mechanism. BMC Cancer 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Chu, R.; Zhao, X.; Griffin, C.; Staub, R.E.; Shoemaker, M.; Climent, J.; Leitman, D.; Cohen, I.; Shtivelman, E.; Fong, S. Selective Concomitant Inhibition of mTORC1 and mTORC2 Activity in Estrogen Receptor Negative Breast Cancer Cells by BN107 and Oleanolic Acid. Int. J. Cancer 2010, 127, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, M.S.; Garcia, M.; Pujol, P.; Fontana, J.A.; Rochefort, H. Why are Estrogen-Receptor-Negative Breast Cancers More Aggressive than the Estrogen-Receptor-Positive Breast Cancers? Invasion Metastasis 1994, 14, 329–336. [Google Scholar] [PubMed]
- Pratheeshkumar, P.; Kuttan, G. Oleanolic Acid Induces Apoptosis by Modulating p53, Bax, Bcl-2 and Caspase-3 Gene Expression and Regulates the Activation of Transcription Factors and Cytokine Profile in B16F. J. Environ. Pathol. Toxicol. Oncol. 2011, 30, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Vera-Ramirez, L.; Sanchez-Rovira, P.; Ramirez-Tortosa, M.C.; Ramirez-Tortosa, C.L.; Granados-Principal, S.; Lorente, J.A.; Quiles, J.L. Free Radicals in Breast Carcinogenesis, Breast Cancer Progression and Cancer Stem Cells. Biological Bases to Develop Oxidative-Based Therapies. Crit. Rev. Oncol. Hematol. 2011, 80, 347–368. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Hang, T.J.; Wang, Y.; Jiang, L.; Wu, X.L.; Zhang, Z.; Shen, J.; Zhang, Y. Determination of Oleanolic Acid in Human Plasma and Study of its Pharmacokinetics in Chinese Healthy Male Volunteers by HPLC Tandem Mass Spectrometry. J. Pharm. Biomed. Anal. 2006, 40, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Rada, M.; Ruiz-Gutierrez, V.; Guinda, A. Determination of Triterpenic Acids in Human Serum by High-Performance Liquid Chromatography: Triterpenoid Interaction with Serum Protein. J. Agric. Food Chem. 2011, 59, 2308–2313. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Mena, G.; Juan, M.E.; Garcia-Granados, A.; Planas, J.M. Determination of Maslinic Acid, a Pentacyclic Triterpene from Olives, in Rat Plasma by High-Performance Liquid Chromatography. J. Agric. Food Chem. 2012, 60, 10220–10225. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, D.A.; Shoemaker, R.H.; Paull, K.D.; Monks, A.; Tierney, S.; Nofziger, T.H.; Currens, M.J.; Seniff, D.; Boyd, M.R. Evaluation of a Soluble tetrazolium/formazan Assay for Cell Growth and Drug Sensitivity in Culture using Human and Other Tumor Cell Lines. Cancer Res. 1988, 48, 4827–4833. [Google Scholar] [PubMed]
- Lee, D.H.; Lim, B.S.; Lee, Y.K.; Yang, H.C. Effects of Hydrogen Peroxide (H2O2) on Alkaline Phosphatase Activity and Matrix Mineralization of Odontoblast and Osteoblast Cell Lines. Cell Biol. Toxicol. 2006, 22, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds AO (oleanolic acid) and MA (maslinic acid) are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Quesada, C.; López-Biedma, A.; Gaforio, J.J. Oleanolic Acid, a Compound Present in Grapes and Olives, Protects against Genotoxicity in Human Mammary Epithelial Cells. Molecules 2015, 20, 13670-13688. https://doi.org/10.3390/molecules200813670
Sánchez-Quesada C, López-Biedma A, Gaforio JJ. Oleanolic Acid, a Compound Present in Grapes and Olives, Protects against Genotoxicity in Human Mammary Epithelial Cells. Molecules. 2015; 20(8):13670-13688. https://doi.org/10.3390/molecules200813670
Chicago/Turabian StyleSánchez-Quesada, Cristina, Alicia López-Biedma, and José J. Gaforio. 2015. "Oleanolic Acid, a Compound Present in Grapes and Olives, Protects against Genotoxicity in Human Mammary Epithelial Cells" Molecules 20, no. 8: 13670-13688. https://doi.org/10.3390/molecules200813670
APA StyleSánchez-Quesada, C., López-Biedma, A., & Gaforio, J. J. (2015). Oleanolic Acid, a Compound Present in Grapes and Olives, Protects against Genotoxicity in Human Mammary Epithelial Cells. Molecules, 20(8), 13670-13688. https://doi.org/10.3390/molecules200813670