Maslinic Acid, a Natural Phytoalexin-Type Triterpene from Olives — A Promising Nutraceutical?

Abstract
:1. Introduction
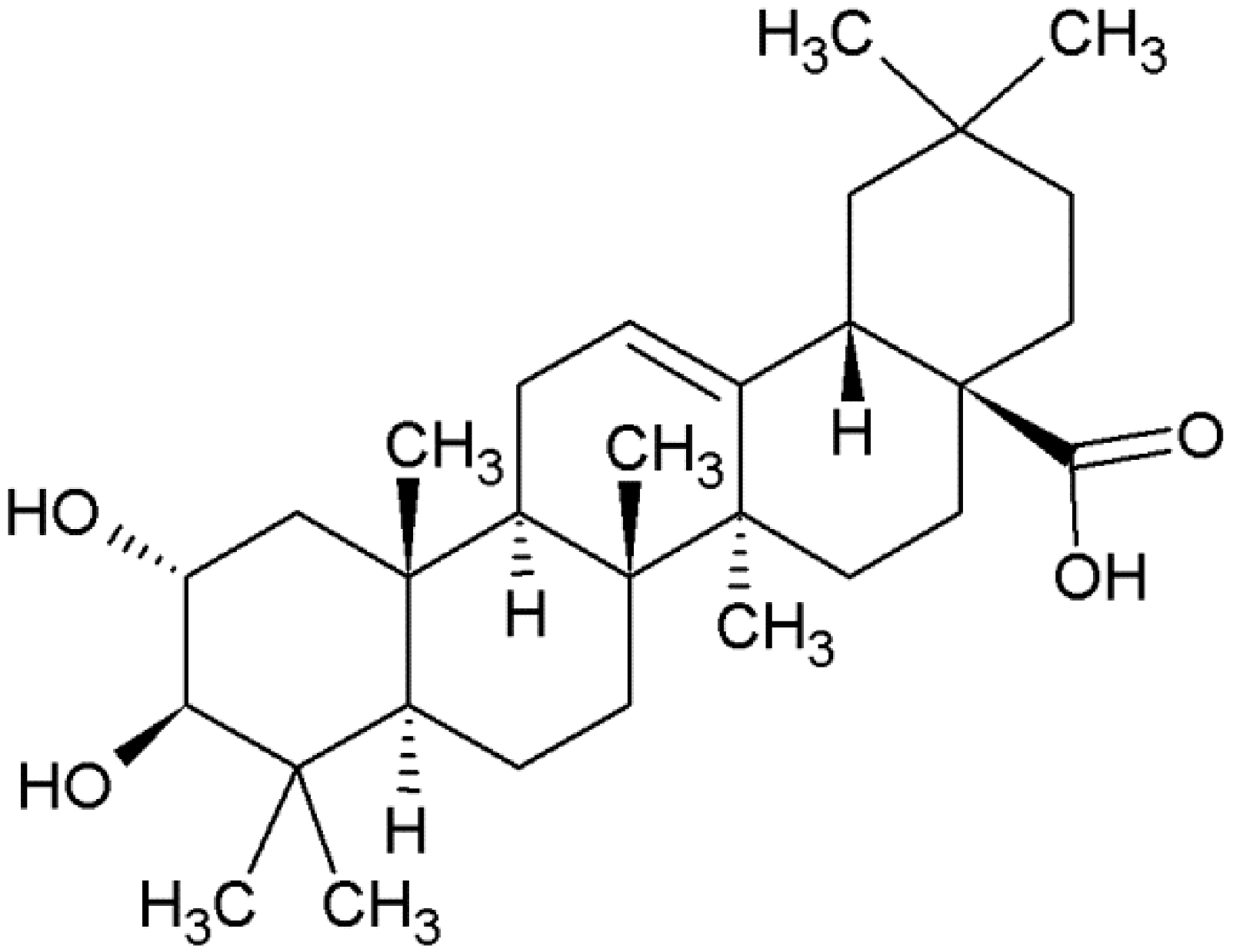
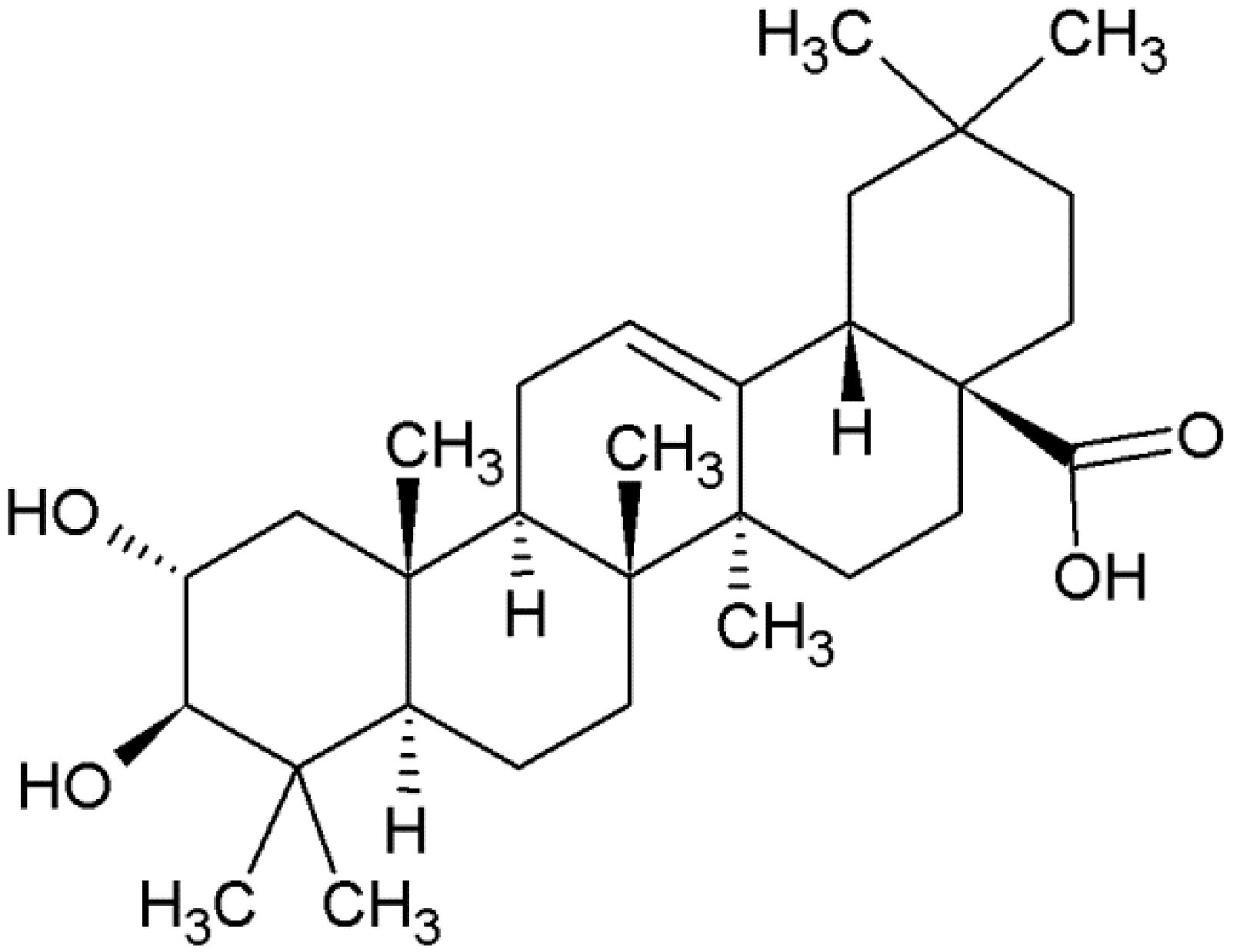
1.1. Biosynthesis and Role as a Phytoalexin
1.2. Natural Sources

{kind=link}
| Maslinic Acid (mg/kg Dry Weight) | References | |
|---|---|---|
| Table olives | ||
| Kalamata (plain black) | 1318 | [21] |
| Hojiblanca (plain green) | 905 | [21] |
| Gordal (plain green) | 414 | [21] |
| Manzanilla (plain green) | 384 | [21] |
| Cacereña (plain black) | 295 | [21] |
| Fresh vegetables | ||
| Spinach | 1260 | [22] |
| Eggplant | 840 | [22] |
| Aromatic herbs | ||
| Brown mustard | 330 | [22] |
| Leaf mustard | 1740 | [23] |
| Basil | 350, 320 | [22,23] |
| Cooked legumes | ||
| Small lentils | 26.3 | [24] |
| Large lentils | 39.5 | [24] |
| Chickpeas | 61.9 | [24] |
| Fresh fruits | ||
| Mandarin | 1.18 | [25] |
| Pomegranate | 10.8 | [25] |
2. Biological Effects
2.1. Maslinic Acid and Cancer
2.1.1. Maslinic Acid Exerts an Anti-Proliferative Activity through Arresting Cell Cycle and Activates Both the Intrinsic and the Extrinsic Apoptotic Pathways in Vitro
| Origin | Cell Line | IC50 (μM) | References |
|---|---|---|---|
| Human colorectal adenocarcinoma | HT-29 | 101.2 | [27] |
| HT-29 | 61 | [28] | |
| Caco-2 | 85 | [28] | |
| Caco-2 | 15.4 | [49] | |
| Human hepatocellular carcinoma | HepG2 | 69.1 | [49] |
| Human breast adenocarcinoma | MCF-7 | 136.0 | [49] |
| Human salivary gland adenoid cystic carcinoma | ACC-2 (low metastasis) | 43.7 | [46] |
| ACC-M (high metastasis) | 45.8 | [46] | |
| Human transitional cell urinary bladder carcinoma | T24 | 33.0 | [47] |
| 253J | 71.8 | [47] | |
| TCCSUP | 28.0 | [47] | |
| Human transitional cell urinary bladder papilloma | RT4 | 42.7 | [47] |
| Human synovial sarcoma | SW 982 | 45.3 | [48] |
| Human uterus leiomyosarcoma | SK-UT-1 | 59.1 | [48] |
2.1.2. Maslinic Acid Targets Other Cancer-Related Signaling Pathways
2.1.3. The Antitumor Activity of Maslinic Acid also Occurs in Vivo
2.2. Maslinic Acid and Diabetes
2.3. Maslinic Acid as Antioxidant and Anti-Inflammatory
2.4. Maslinic Acid and Cardioprotection
2.5. Maslinic Acid and Neuroprotection
2.6. Maslinic Acid as Antiparasitic
2.7. Maslinic Acid and Growth
2.8. Other Biological Activities
3. Conclusion and Future Prospects
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sánchez-González, M.; Lozano-Mena, G.; Juan, M.E.; García-Granados, A.; Planas, J.M. Assessment of the safety of maslinic acid, a bioactive compound from Olea europaea L. Mol. Nutr. Food Res. 2013, 57, 339–346. [Google Scholar] [CrossRef]
- Bächler, L. Chemische Untersuchungen über die Früchte von Crataegus oxyacantha L. (Monographie der Mehlbeeren); Universität Basel: Basel, Switzerland, 1927. [Google Scholar]
- Tschesche, R.; Fugmann, R. Crataegolsäure, ein neues triterpenoid aus Crataegus oxyacantha. Ein beitrag zur konstitution der α-amyrine. Chem. Ber. 1951, 84, 810–826. [Google Scholar]
- Tschesche, R.; Heesch, A.; Fugmann, R. Über triterpenoide, III. Mitteil.: Zur kenntnis der crataegolsäure. Chem. Ber. 1953, 86, 626–629. [Google Scholar]
- Caglioti, L.; Cainelli, G.; Minutilli, F. Constitution of maslinic acid. Chim. Ind. 1961, 43, 278. [Google Scholar]
- Caputo, R.; Mangoni, L.; Monaco, P.; Previtera, L. Triterpenes in husks of Olea europaea. Phytochemistry 1974, 13, 1551–1552. [Google Scholar] [CrossRef]
- Vioque, A.; Morris, L. Minor components of olive oils. I. Triterpenoid acids in an acetone-extracted orujo oil. J. Am. Oil Chem. Soc. 1961, 38, 458–488. [Google Scholar]
- Bianchi, G.; Pozzi, N.; Vlahov, G. Pentacyclic triterpene acids in olives. Phytochemistry 1994, 37, 205–207. [Google Scholar] [CrossRef]
- Xu, R.; Fazio, G.C.; Matsuda, S.P.T. On the origins of triterpenoid skeletal diversity. Phytochemistry 2004, 65, 261–291. [Google Scholar] [CrossRef]
- Tschesche, R.; Poppel, G.; Über Triterpene, V. Zur Kenntnis der crataegolsäure und über zwei neue triterpencarbonsäuren aus Crataegus oxyacantha L. Chem. Ber. 1959, 92, 320–328. [Google Scholar] [CrossRef]
- Stiti, N.; Triki, S.; Hartmann, M.A. Formation of triterpenoids throughout Olea europaea fruit ontogeny. Lipids 2007, 42, 55–67. [Google Scholar] [CrossRef]
- Kombargi, W.S.; Michelakis, S.E.; Petrakis, C.A. Effect of olive surface waxes on oviposition by Bactrocera oleae (Diptera: Tephritidae). J. Econ. Entomol. 1998, 91, 993–998. [Google Scholar]
- Pungitore, C.R.; García, M.; Gianello, J.C.; Sosa, M.E.; Tonn, C.E. Insecticidal and antifeedant effects of Junellia aspera (Verbenaceae) triterpenes and derivatives on Sitophilus oryzae (Coleoptera: Curculionidae). J. Stored Prod. Res. 2005, 41, 433–443. [Google Scholar] [CrossRef]
- Lu, H.; Xi, C.; Chen, J.; Li, W. Determination of triterpenoid acids in leaves of Eriobotrya japonica collected at in different seasons. Zhongguo Zhong Yao Za Zhi 2009, 34, 2353–2355. [Google Scholar]
- Banno, N.; Akihisa, T.; Tokuda, H.; Yasukawa, K.; Taguchi, Y.; Akazawa, H.; Ukiya, M.; Kimura, Y.; Suzuki, T.; Nishino, H. Anti-inflammatory and antitumor-promoting effects of the triterpene acids from the leaves of Eriobotrya japonica. Biol. Pharm. Bull. 2005, 28, 1995–1999. [Google Scholar] [CrossRef]
- Kim, D.H.; Han, K.M.; Chung, I.S.; Kim, D.K.; Kim, S.H.; Kwon, B.M.; Jeong, T.S.; Park, M.H.; Ahn, E.M.; Baek, N.I. Triterpenoids from the flower of Campsis grandiflora K. Schum. as human acyl-CoA: Cholesterol acyltransferase inhibitors. Arch. Pharm. Res. 2005, 28, 550–556. [Google Scholar] [CrossRef]
- Xu, H.X.; Zeng, F.Q.; Wan, M.; Sim, K.Y. Anti-HIV triterpene acids from Geum japonicum. J. Nat. Prod. 1996, 59, 643–645. [Google Scholar]
- Yoshida, T.; Okuda, T.; Memon, M.U.; Shingu, T. Tannins of rosaceous medicinal plants. Part 2. Gemins A, B, and C, new dimeric ellagitannins from Geum japonicum. J. Chem. Soc. Perkin Trans. 1985, 1, 315–321. [Google Scholar]
- Zou, Z.M.; Cong, P.Z. Studies on the chemical constituents from roots of Agastache rugosa. Yao Xue Xue Bao 1991, 26, 906–910. [Google Scholar]
- Shin, S.; Kang, C.A. Antifungal activity of the essential oil of Agastache rugosa Kuntze and its synergism with ketoconazole. Lett. Appl. Microbiol. 2003, 36, 111–115. [Google Scholar] [CrossRef]
- Romero, C.; García, A.; Medina, E.; Ruiz-Méndez, M.V.; de Castro, A.; Brenes, M. Triterpenic acids in table olives. Food Chem. 2010, 118, 670–674. [Google Scholar] [CrossRef]
- Lin, C.C.; Huang, C.Y.; Mong, M.C.; Chan, C.Y.; Yin, M.C. Antiangiogenic potential of three triterpenic acids in human liver cancer cells. J. Agric. Food Chem. 2011, 59, 755–762. [Google Scholar]
- Yin, M.C.; Lin, M.C.; Mong, M.C.; Lin, C.Y. Bioavailability, distribution, and antioxidative effects of selected triterpenes in mice. J. Agric. Food Chem. 2012, 60, 7697–7701. [Google Scholar]
- Kalogeropoulos, N.; Chiou, A.; Ioannou, M.; Karathanos, V.T.; Hassapidou, M.; Andrikopoulos, N.K. Nutritional evaluation and bioactive microconstituents (phytosterols, tocopherols, polyphenols, triterpenic acids) in cooked dry legumes usually consumed in the Mediterranean countries. Food Chem. 2010, 121, 682–690. [Google Scholar] [CrossRef]
- Li, G.L.; You, J.M.; Song, C.H.; Xia, L.; Zheng, J.; Suo, Y.R. Development of a new HPLC method with precolumn fluorescent derivatization for rapid, selective and sensitive detection of triterpenic acids in fruits. J. Agric. Food Chem. 2011, 59, 2972–2979. [Google Scholar] [CrossRef]
- Juan, M.E.; Wenzel, U.; Ruiz-Gutiérrez, V.; Planas, J.M.; Daniel, H. Maslinic acid, a natural compound from olives, induces apoptosis in HT-29 human colon cancer cell line. In Proceedings of the Experimental Biology 2005 Meeting, 35th International Congress of Physiological Sciences, San Diego, CA, USA, 31 March–6 April 2005.
- Juan, M.E.; Planas, J.M.; Ruiz-Gutiérrez, V.; Daniel, H.; Wenzel, U. Anti-proliferative and apoptosis-inducing effects of maslinic and oleanolic acids, two pentacyclic triterpenes from olives, on HT-29 colon cancer cells. Br. J. Nutr. 2008, 100, 36–43. [Google Scholar]
- Reyes, F.J.; Centelles, J.J.; Lupiáñez, J.A.; Cascante, M. (2α,3β)-2,3-dihydroxyolean-12-en-28-oic acid, a new natural triterpene from Olea europea, induces caspase dependent apoptosis selectively in colon adenocarcinoma cells. FEBS Lett. 2006, 580, 6302–6310. [Google Scholar] [CrossRef]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef]
- Reyes-Zurita, F.J.; Rufino-Palomares, E.E.; Lupiáñez, J.A.; Cascante, M. Maslinic acid, a natural triterpene from Olea europaea L., induces apoptosis in HT29 human colon-cancer cells via the mitochondrial apoptotic pathway. Cancer Lett. 2009, 273, 44–54. [Google Scholar] [CrossRef]
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef]
- Reyes-Zurita, F.J.; Pachón-Peña, G.; Lizárraga, D.; Rufino-Palomares, E.E.; Cascante, M.; Lupiáñez, J.A. The natural triterpene maslinic acid induces apoptosis in HT29 colon cancer cells by a JNK-p53-dependent mechanism. BMC Cancer 2011, 154:1–154:13. [Google Scholar]
- Tsuruta, F.; Sunayama, J.; Mori, Y.; Hattori, S.; Shimizu, S.; Tsujimoto, Y.; Yoshioka, K.; Masuyama, N.; Gotoh, Y. JNK promotes Bax translocation to mitochondria through phosphorylation of 14-3-3 proteins. EMBO J. 2004, 23, 1889–1899. [Google Scholar] [CrossRef]
- Deng, Y.; Ren, X.; Yang, L.; Lin, Y.; Wu, X. A JNK-dependent pathway is required for TNFα-induced apoptosis. Cell 2003, 115, 61–70. [Google Scholar] [CrossRef]
- Wilkinson, J.C.; Wilkinson, A.S.; Scott, F.L.; Csomos, R.A.; Salvesen, G.S.; Duckett, C.S. Neutralization of Smac/Diablo by inhibitors of apoptosis (IAPs). A caspase-independent mechanism for apoptotic inhibition. J. Biol. Chem. 2004, 279, 51082–51090. [Google Scholar]
- Miyashita, T.; Krajewski, S.; Krajewska, M.; Wang, H.G.; Lin, H.K.; Liebermann, D.A.; Hoffman, B.; Reed, J.C. Tumor suppressor p53 is a regulator of bcl-2 and bax gene expression in vitro and in vivo. Oncogene 1994, 9, 1799–1805. [Google Scholar]
- Micheau, O.; Tschopp, J. Induction of TNF receptor I-mediated apoptosis via two sequential signaling complexes. Cell 2003, 114, 181–190. [Google Scholar] [CrossRef]
- Napetschnig, J.; Wu, H. Molecular basis of NF-κB signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef]
- Karin, M. Nuclear factor-κB in cancer development and progression. Nature 2006, 441, 431–436. [Google Scholar] [CrossRef]
- Stennicke, H.R.; Jürgensmeier, J.M.; Shin, H.; Deveraux, Q.; Wolf, B.B.; Yang, X.; Zhou, Q.; Ellerby, H.M.; Ellerby, L.M.; Bredesen, D.; et al. Pro-caspase-3 is a major physiologic target of caspase-8. J. Biol. Chem. 1998, 273, 27084–27090. [Google Scholar] [CrossRef]
- Li, C.; Yang, Z.; Zhai, C.; Qiu, W.; Li, D.; Yi, Z.; Wang, L.; Tang, J.; Qian, M.; Luo, J.; et al. Maslinic acid potentiates the anti-tumor activity of tumor necrosis factor α by inhibiting NF-κB signaling pathway. Mol. Cancer 2010, 73:1–73:13. [Google Scholar]
- Hsum, Y.W.; Yew, W.T.; Hong, P.L.; Soo, K.K.; Hoon, L.S.; Chieng, Y.C.; Mooi, L.Y. Cancer chemopreventive activity of maslinic acid: Suppression of COX-2 expression and inhibition of NF-κB and AP-1 activation in Raji cells. Planta Med. 2011, 77, 152–157. [Google Scholar] [CrossRef]
- Dannenberg, A.J.; Subbaramaiah, K. Targeting cyclooxygenase-2 in human neoplasia: Rationale and promise. Cancer Cell 2003, 4, 431–436. [Google Scholar] [CrossRef]
- Schulze-Osthoff, K.; Ferrari, D.; Riehemann, K.; Wesselborg, S. Regulation of NF-kappa B activation by MAP kinase cascades. Immunobiology 1997, 198, 35–49. [Google Scholar] [CrossRef]
- Wu, D.M.; Zhao, D.; Li, D.Z.; Xu, D.Y.; Chu, W.F.; Wang, X.F. Maslinic acid induces apoptosis in salivary gland adenoid cystic carcinoma cells by Ca2+-evoked p38 signaling pathway. Naunyn Schmiedebergs Arch. Pharmacol. 2011, 383, 321–330. [Google Scholar] [CrossRef]
- Zhang, S.; Ding, D.; Zhang, X.; Shan, L.; Liu, Z. Maslinic acid induced apoptosis in bladder cancer cells through activating p38 MAPK signaling pathway. Mol. Cell Biochem. 2014, 392, 281–287. [Google Scholar] [CrossRef]
- Villar, V.H.; Vögler, O.; Barceló, F.; Gómez-Florit, M.; Martínez-Serra, J.; Obrador-Hevia, A.; Martín-Broto, J.; Ruiz-Gutiérrez, V.; Alemany, R. Oleanolic and maslinic acid sensitize soft tissue sarcoma cells to doxorubicin by inhibiting the multidrug resistance protein MRP-1, but not P-glycoprotein. J. Nutr. Biochem. 2014, 25, 429–438. [Google Scholar] [CrossRef]
- He, X.; Liu, R.H. Triterpenoids isolated from apple peels have potent anti-proliferative activity and may be partially responsible for apple’s anticancer activity. J. Agric. Food Chem. 2007, 55, 4366–4370. [Google Scholar] [CrossRef]
- Zhong, H.; de Marzo, A.M.; Laughner, E.; Lim, M.; Hilton, D.A.; Zagzag, D.; Buechler, P.; Isaacs, W.B.; Semenza, G.L.; Simons, J.W. Overexpression of hypoxia-inducible factor 1α in common human cancers and their metastases. Cancer Res. 1999, 59, 5830–5835. [Google Scholar]
- Rankin, E.B.; Giaccia, A.J. The role of hypoxia-inducible factors in tumorigenesis. Cell Death Differ. 2008, 15, 678–685. [Google Scholar] [CrossRef]
- Bourboulia, D.; Stetler-Stevenson, W.G. Matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs): Positive and negative regulators in tumor cell adhesion. Semin. Cancer Biol. 2010, 20, 161–168. [Google Scholar] [CrossRef]
- Mekkawy, A.H.; Morris, D.L.; Pourgholami, M.H. Urokinase plasminogen activator system as a potential target for cancer therapy. Future Oncol. 2009, 5, 1487–1499. [Google Scholar] [CrossRef]
- Park, S.Y.; Nho, C.W.; Kwon, D.Y.; Kang, Y.H.; Lee, K.W.; Park, J.H. Maslinic acid inhibits the metastatic capacity of DU145 human prostate cancer cells: Possible mediation via hypoxia-inducible factor-1α signalling. Br. J. Nutr. 2013, 109, 210–222. [Google Scholar] [CrossRef]
- Pialoux, V.; Mounier, R.; Brown, A.D.; Steinback, C.D.; Rawling, J.M.; Poulin, M.J. Relationship between oxidative stress and HIF-1α mRNA during sustained hypoxia in humans. Free Radic. Biol. Med. 2009, 46, 321–326. [Google Scholar] [CrossRef]
- Sánchez-Tena, S.; Reyes-Zurita, F.J.; Díaz-Moralli, S.; Vinardell, M.P.; Reed, M.; García-García, F.; Dopazo, J.; Lupiáñez, J.A.; Günther, U.; Cascante, M. Maslinic acid-enriched diet decreases intestinal tumorigenesis in ApcMin/+ mice through transcriptomic and metabolomic reprogramming. PLoS One. 2013, 8, e59392. [Google Scholar]
- Wen, X.; Sun, H.; Liu, J.; Wu, G.; Zhang, L.; Wu, X.; Ni, P. Pentacyclic triterpenes. Part 1: The first examples of naturally occurring pentacyclic triterpenes as a new class of inhibitors of glycogen phosphorylases. Bioorg. Med. Chem. Lett. 2005, 15, 4944–4948. [Google Scholar] [CrossRef]
- Wen, X.; Sun, H.; Liu, J.; Cheng, K.; Zhang, P.; Zhang, L.; Hao, J.; Zhang, L.; Ni, P.; Zographos, S.E.; et al. Naturally occurring pentacyclic triterpenes as inhibitors of glycogen phosphorylase: Synthesis, structure-activity relationships, and X-ray crystallographic studies. J. Med. Chem. 2008, 51, 3540–3554. [Google Scholar] [CrossRef]
- Liu, J.; Sun, H.; Duan, W.; Mu, D.; Zhang, L. Maslinic acid reduces blood glucose in KK-Ay mice. Biol. Pharm. Bull. 2007, 30, 2075–2078. [Google Scholar] [CrossRef]
- Khathi, A.; Serumula, M.R.; Myburg, R.B.; van Heerden, F.R.; Musabayane, C.T. Effects of Syzygium aromaticum-derived triterpenes on postprandial blood glucose in streptozotocin-induced diabetic rats following carbohydrate challenge. PLoS One 2013, 8, e81632. [Google Scholar]
- Guan, T.; Qian, Y.; Tang, X.; Huang, M.; Huang, L.; Li, Y.; Sun, H. Maslinic acid, a natural inhibitor of glycogen phosphorylase, reduces cerebral ischemic injury in hyperglycemic rats by GLT-1 up-regulation. J. Neurosci. Res. 2011, 89, 1829–1839. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Chen, Y.P.; Mao, L.F.; Shang, J.; Sun, H.B.; Zhang, L.Y. Maslinic acid modulates glycogen metabolism by enhancing the insulin signaling pathway and inhibiting glycogen phosphorylase. Chin. J. Nat. Med. 2014, 12, 259–265. [Google Scholar]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef]
- Montilla, M.P.; Agil, A.; Navarro, M.C.; Jiménez, M.I.; García-Granados, A.; Parra, A.; Cabo, M.M. Antioxidant activity of maslinic acid, a triterpene derivative obtained from Olea europaea. Planta Med. 2003, 69, 472–474. [Google Scholar] [CrossRef]
- Wang, R.; Wang, W.; Wang, L.; Liu, R.; Ding, Y.; Du, L. Constituents of the flowers of Punica granatum. Fitoterapia 2006, 77, 534–537. [Google Scholar] [CrossRef]
- Allouche, Y.; Beltrán, G.; Gaforio, J.J.; Uceda, M.; Mesa, M.D. Antioxidant and antiatherogenic activities of pentacyclic triterpenic diols and acids. Food Chem. Toxicol. 2010, 48, 2885–2890. [Google Scholar]
- Márquez-Martín, A.; de la Puerta, R.; Fernández-Arche, A.; Ruiz-Gutiérrez, V.; Yaqoob, P. Modulation of cytokine secretion by pentacyclic triterpenes from olive pomace oil in human mononuclear cells. Cytokine 2006, 36, 211–217. [Google Scholar] [CrossRef]
- Huang, L.; Guan, T.; Qian, Y.; Huang, M.; Tang, X.; Li, Y.; Sun, H. Anti-inflammatory effects of maslinic acid, a natural triterpene, in cultured cortical astrocytes via suppression of nuclear factor-kappa B. Eur. J. Pharmacol. 2011, 672, 169–174. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, R.; Perona, J.S.; Herrera, M.D.; Ruiz-Gutiérrez, V. Triterpenic compounds from “orujo” olive oil elicit vasorelaxation in aorta from spontaneously hypertensive rats. J. Agric. Food Chem. 2006, 54, 2096–2102. [Google Scholar] [CrossRef]
- Liu, J.; Sun, H.; Wang, X.; Mu, D.; Liao, H.; Zhang, L. Effects of oleanolic acid and maslinic acid on hyperlipidemia. Drug Dev. Res. 2007, 68, 261–266. [Google Scholar] [CrossRef]
- Hussain Shaik, A.; Rasool, S.N.; Abdul Kareem, M.; Krushna, G.S.; Akhtar, P.M.; Devi, K.L. Maslinic acid protects against isoproterenol-induced cardiotoxicity in albino Wistar rats. J. Med. Food 2012, 15, 741–746. [Google Scholar] [CrossRef]
- Mackness, M.; Mackness, B. Targeting paraoxonase-1 in atherosclerosis. Expert Opin. Ther. Targets 2013, 17, 829–837. [Google Scholar] [CrossRef]
- Ros, E.; Martínez-González, M.A.; Estruch, R.; Salas-Salvadó, J.; Fitó, M.; Martínez, J.A.; Corella, D. Mediterranean diet and cardiovascular health: Teachings of the PREDIMED Study. In Proceedings of the IUNS 20th Congress of Nutrition, Granada, Spain, 15–20 September 2013; pp. 330S–336S.
- Qian, Y.; Guan, T.; Tang, X.; Huang, L.; Huang, M.; Li, Y.; Sun, H. Maslinic acid, a natural triterpenoid compound from Olea europaea, protects cortical neurons against oxygen-glucose deprivation-induced injury. Eur. J. Pharmacol. 2011, 670, 148–153. [Google Scholar] [CrossRef]
- Moncada, S.; Erusalimsky, J.D. Does nitric oxide modulate mitochondrial energy generation and apoptosis? Nat. Rev. Mol. Cell Biol. 2002, 3, 214–220. [Google Scholar] [CrossRef]
- Moro, M.A.; de Alba, J.; Leza, J.C.; Lorenzo, P.; Fernández, A.P.; Bentura, M.L.; Boscá, L.; Rodrigo, J.; Lizasoain, I. Neuronal expression of inducible nitric oxide synthase after oxygen and glucose deprivation in rat forebrain slices. Eur. J. Neurosci. 1998, 10, 445–456. [Google Scholar] [CrossRef]
- Lau, A.; Tymianski, M. Glutamate receptors, neurotoxicity and neurodegeneration. Pflugers Arch. 2010, 460, 525–542. [Google Scholar] [CrossRef]
- Kanai, Y.; Hediger, M.A. The glutamate/neutral amino acid transporter family SLC1: Molecular, physiological and pharmacological aspects. Pflügers Archiv 2004, 447, 469–479. [Google Scholar] [CrossRef]
- Qian, Y.; Guan, T.; Tang, X.; Huang, L.; Huang, M.; Li, Y.; Sun, H.; Yu, R.; Zhang, F. Astrocytic glutamate transporter-dependent neuroprotection against glutamate toxicity: An in vitro study of maslinic acid. Eur. J. Pharmacol. 2011, 651, 59–65. [Google Scholar] [CrossRef]
- Kagansky, N.; Levy, S.; Knobler, H. The role of hyperglycemia in acute stroke. Arch. Neurol. 2001, 58, 1209–1212. [Google Scholar] [CrossRef]
- Acebey-Castellón, I.L.; Voutquenne-Nazabadioko, L.; Mai, D.T.H.; Roseau, N.; Bouthagane, N.; Muhammad, D.; le Debar, M.E.; Gangloff, S.C.; Litaudon, M.; Sevenet, T.; et al. Triterpenoid saponins from Symplocos lancifolia. J. Nat. Prod. 2011, 74, 163–168. [Google Scholar] [CrossRef]
- De Pablos, L.M.; González, G.; Rodrigues, R.; García-Granados, A.; Parra, A.; Osuna, A. Action of a pentacyclic triterpenoid, maslinic acid, against Toxoplasma gondii. J. Nat. Prod. 2010, 73, 831–834. [Google Scholar] [CrossRef]
- De Pablos, L.M.; dos Santos, M.F.; Montero, E.; García-Granados, A.; Parra, A.; Osuna, A. Anticoccidial activity of maslinic acid against infection with Eimeria tenella in chickens. Parasitol. Res. 2010, 107, 601–604. [Google Scholar] [CrossRef]
- Moneriz, C.; Marín-García, P.; García-Granados, A.; Bautista, J.M.; Diez, A.; Puyet, A. Parasitostatic effect of maslinic acid. I. Growth arrest of Plasmodium falciparum intraerythrocytic stages. Malar. J. 2011, 82:1–82:10. [Google Scholar]
- Moneriz, C.; Marín-García, P.; Bautista, J.M.; Diez, A.; Puyet, A. Parasitostatic effect of maslinic acid. II. Survival increase and immune protection in lethal Plasmodium yoelii-infected mice. Malar. J. 2011, 103:1–103:9. [Google Scholar]
- Moneriz, C.; Mestres, J.; Bautista, J.M.; Diez, A.; Puyet, A. Multi-targeted activity of maslinic acid as an antimalarial natural compound. FEBS J. 2011, 278, 2951–2961. [Google Scholar] [CrossRef]
- Fernández-Navarro, M.; Peragón, J.; Esteban, F.J.; de la Higuera, M.; Lupiáñez, J.A. Maslinic acid as a feed additive to stimulate growth and hepatic protein-turnover rates in rainbow trout (Onchorhynchus mykiss). Comp. Biochem. Physiol. C 2006, 144, 130–140. [Google Scholar]
- Fernández-Navarro, M.; Peragón, J.; Amores, V.; de la Higuera, M.; Lupiáñez, J.A. Maslinic acid added to the diet increases growth and protein-turnover rates in the white muscle of rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. C 2008, 147, 158–167. [Google Scholar]
- Rufino-Palomares, E.; Reyes-Zurita, F.J.; Fuentes-Almagro, C.A.; de la Higuera, M.; Lupiáñez, J.A.; Peragón, J. Proteomics in the liver of gilthead sea bream (Sparus aurata) to elucidate the cellular response induced by the intake of maslinic acid. Proteomics 2011, 11, 3312–3325. [Google Scholar] [CrossRef]
- Sultana, N.; Lee, N.H. Antielastase and free radical scavenging activities of compounds from the stems of Cornus kousa. Phytother Res. 2007, 21, 1171–1176. [Google Scholar] [CrossRef]
- Ullah, F.; Hussain, H.; Hussain, J.; Bukhari, I.A.; Khan, M.T.; Choudhary, M.I.; Gilani, A.H.; Ahmad, V.U. Tyrosinase inhibitory pentacyclic triterpenes and analgesic and spasmolytic activities of methanol extracts of Rhododendron collettianum. Phytother Res. 2007, 21, 1076–1081. [Google Scholar] [CrossRef]
- Li, C.; Yang, Z.; Li, Z.; Ma, Y.; Zhang, L.; Zheng, C.; Qiu, W.; Wu, X.; Wang, X.; Li, H.; et al. Maslinic acid suppresses osteoclastogenesis and prevents ovariectomy-induced bone loss by regulating RANKL-mediated NF-κB and MAPK signaling pathways. J. Bone Miner. Res. 2011, 26, 644–656. [Google Scholar] [CrossRef]
- Nieto, F.R.; Cobos, E.J.; Entrena, J.M.; Parra, A.; García-Granados, A.; Baeyens, J.M. Antiallodynic and analgesic effects of maslinic acid, a pentacyclic triterpenoid from Olea europaea. J. Nat. Prod. 2013, 76, 737–740. [Google Scholar] [CrossRef]
- Prades, J.; Vögler, O.; Alemany, R.; Gómez-Florit, M.; Funari, S.S.; Ruiz-Gutiérrez, V.; Barceló, F. Plant pentacyclic triterpenic acids as modulators of lipid membrane physical properties. Biochim. Biophys. Acta 2011, 1808, 752–760. [Google Scholar] [CrossRef]
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lozano-Mena, G.; Sánchez-González, M.; Juan, M.E.; Planas, J.M. Maslinic Acid, a Natural Phytoalexin-Type Triterpene from Olives — A Promising Nutraceutical? Molecules 2014, 19, 11538-11559. https://doi.org/10.3390/molecules190811538
Lozano-Mena G, Sánchez-González M, Juan ME, Planas JM. Maslinic Acid, a Natural Phytoalexin-Type Triterpene from Olives — A Promising Nutraceutical? Molecules. 2014; 19(8):11538-11559. https://doi.org/10.3390/molecules190811538
Chicago/Turabian StyleLozano-Mena, Glòria, Marta Sánchez-González, M. Emília Juan, and Joana M. Planas. 2014. "Maslinic Acid, a Natural Phytoalexin-Type Triterpene from Olives — A Promising Nutraceutical?" Molecules 19, no. 8: 11538-11559. https://doi.org/10.3390/molecules190811538
APA StyleLozano-Mena, G., Sánchez-González, M., Juan, M. E., & Planas, J. M. (2014). Maslinic Acid, a Natural Phytoalexin-Type Triterpene from Olives — A Promising Nutraceutical? Molecules, 19(8), 11538-11559. https://doi.org/10.3390/molecules190811538