Resveratrol and Calcium Signaling: Molecular Mechanisms and Clinical Relevance


Abstract
:1. Introduction
1.1. Effects and Mechanisms of Action of Resveratrol as the Basis for Its Therapeutic Potential in Various Diseases
1.2. Key Signaling Proteins Controlling Cellular Calcium Homeostasis
2. Medical Relevance of Modulation of Cellular Calcium Signaling Mechanisms by Resveratrol
2.1. Modulation of Cellular Calcium Signaling Mechanisms by Resveratrol in Excitable Cells
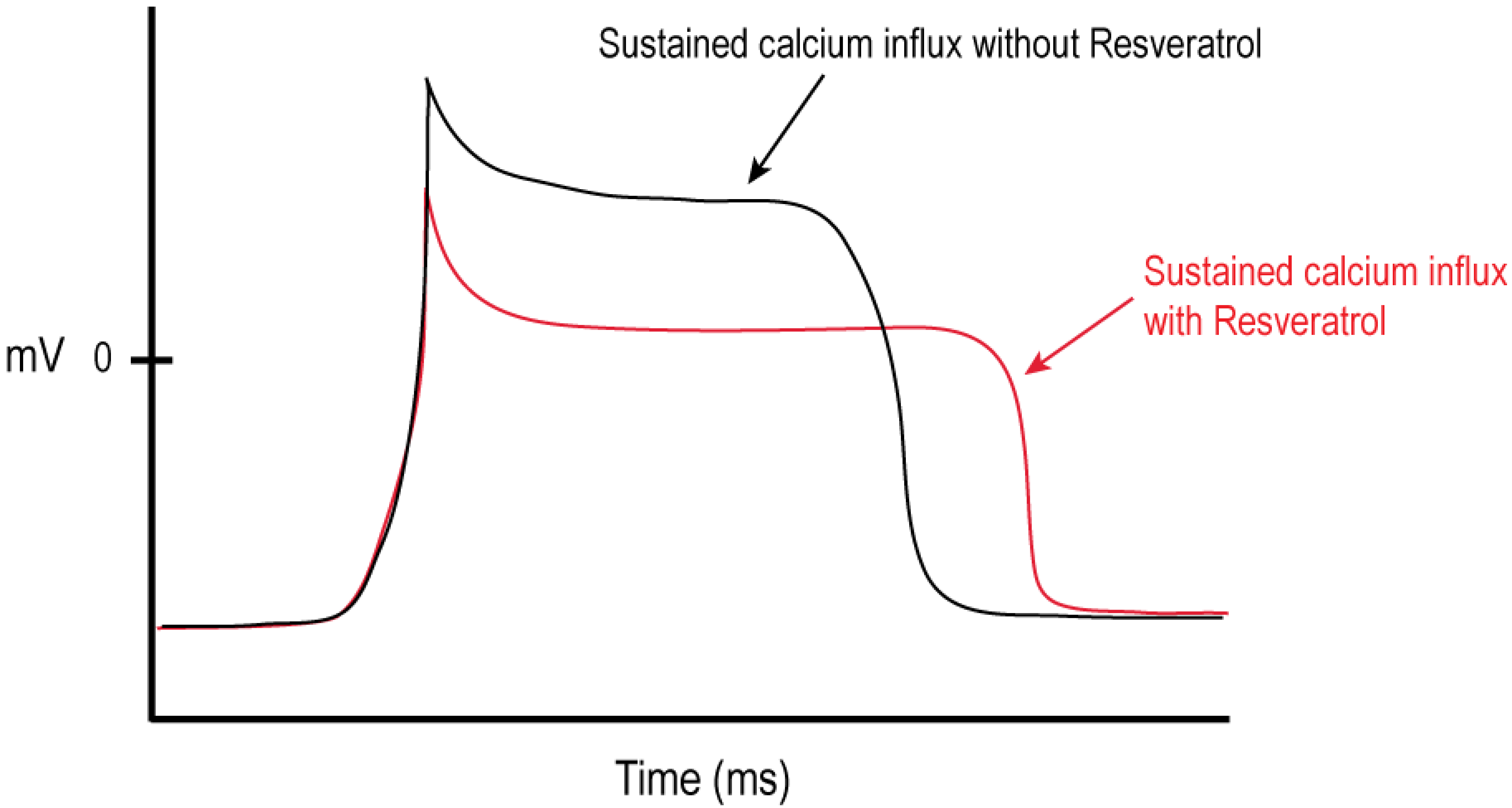
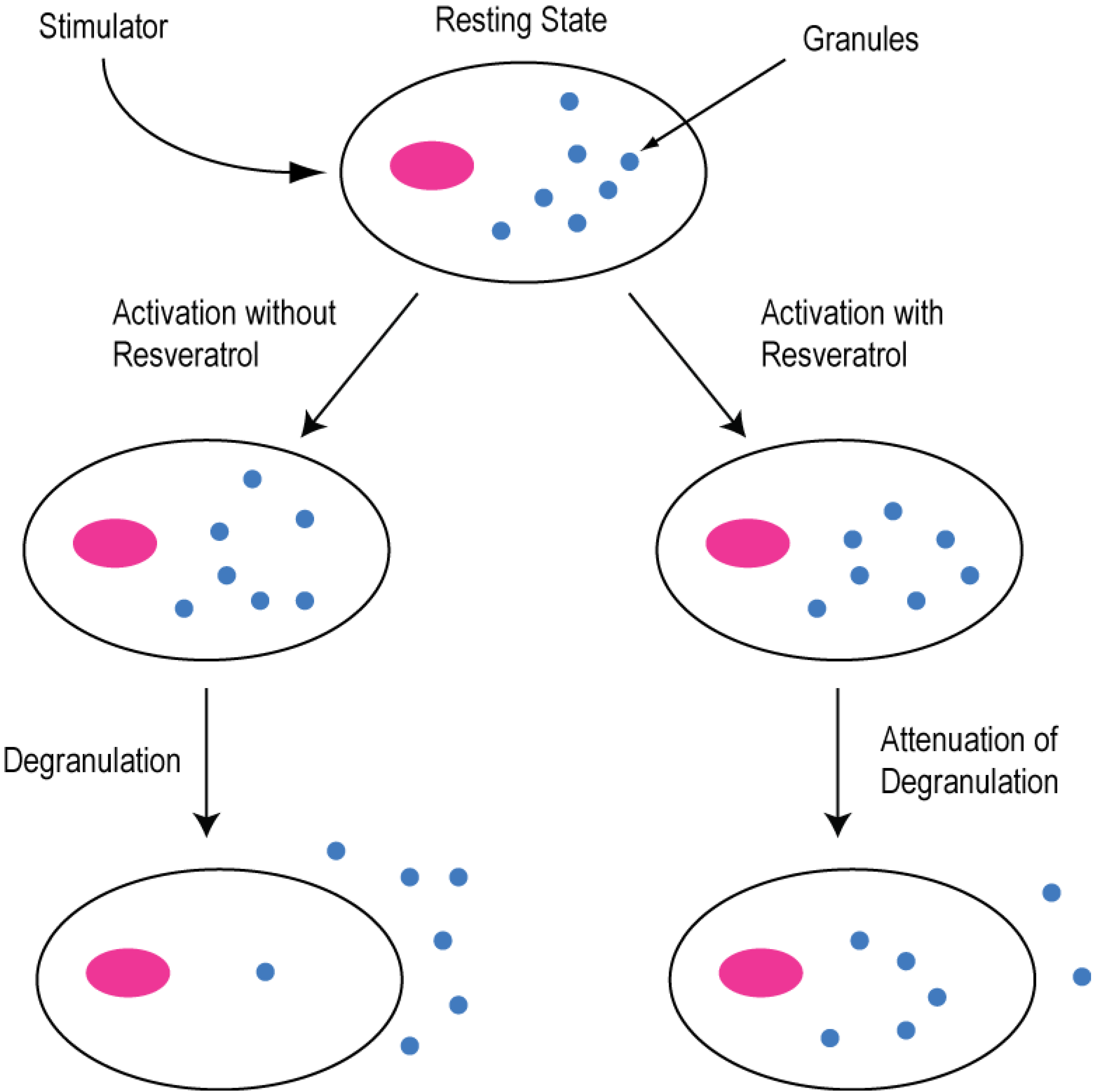
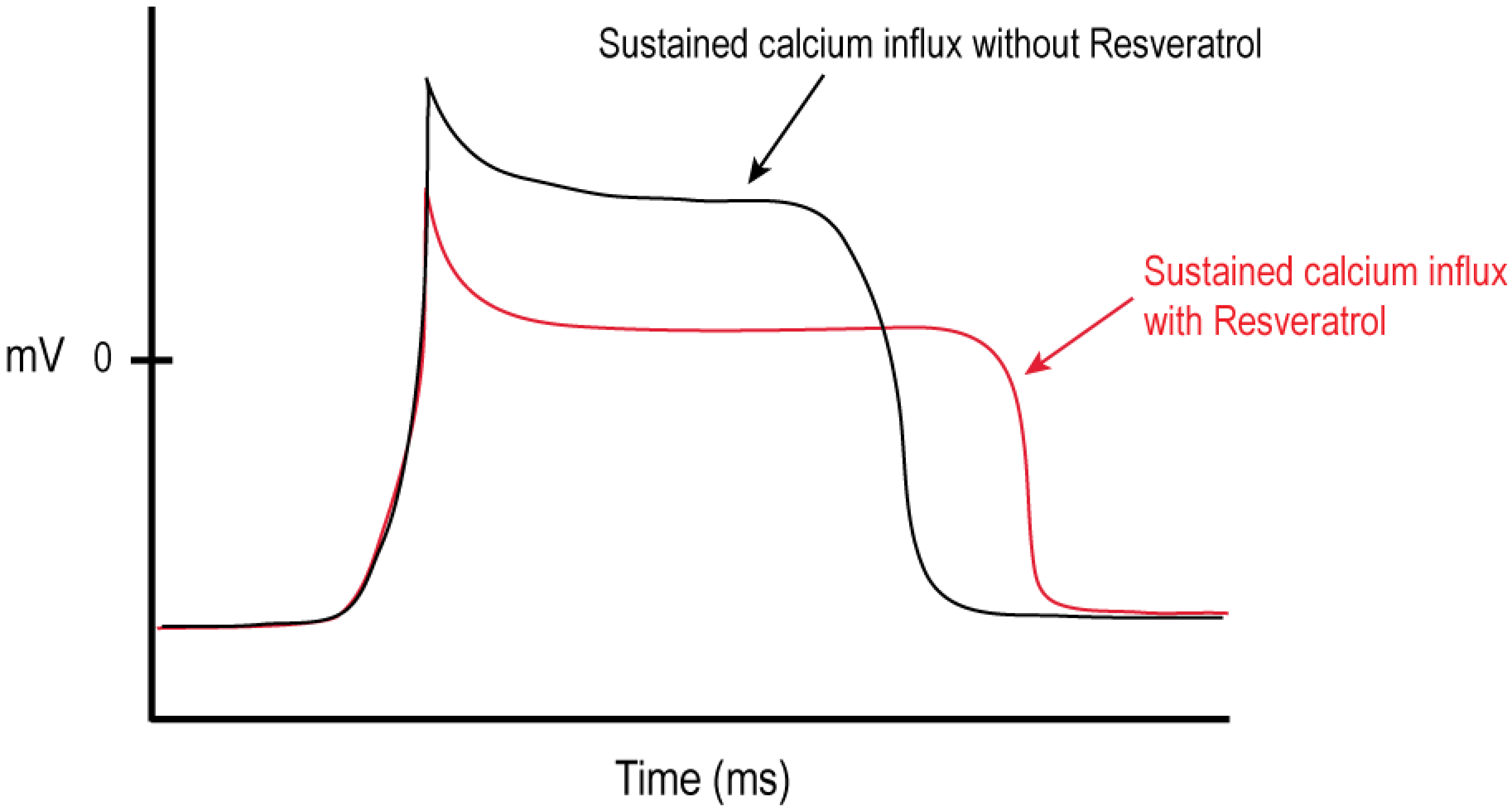
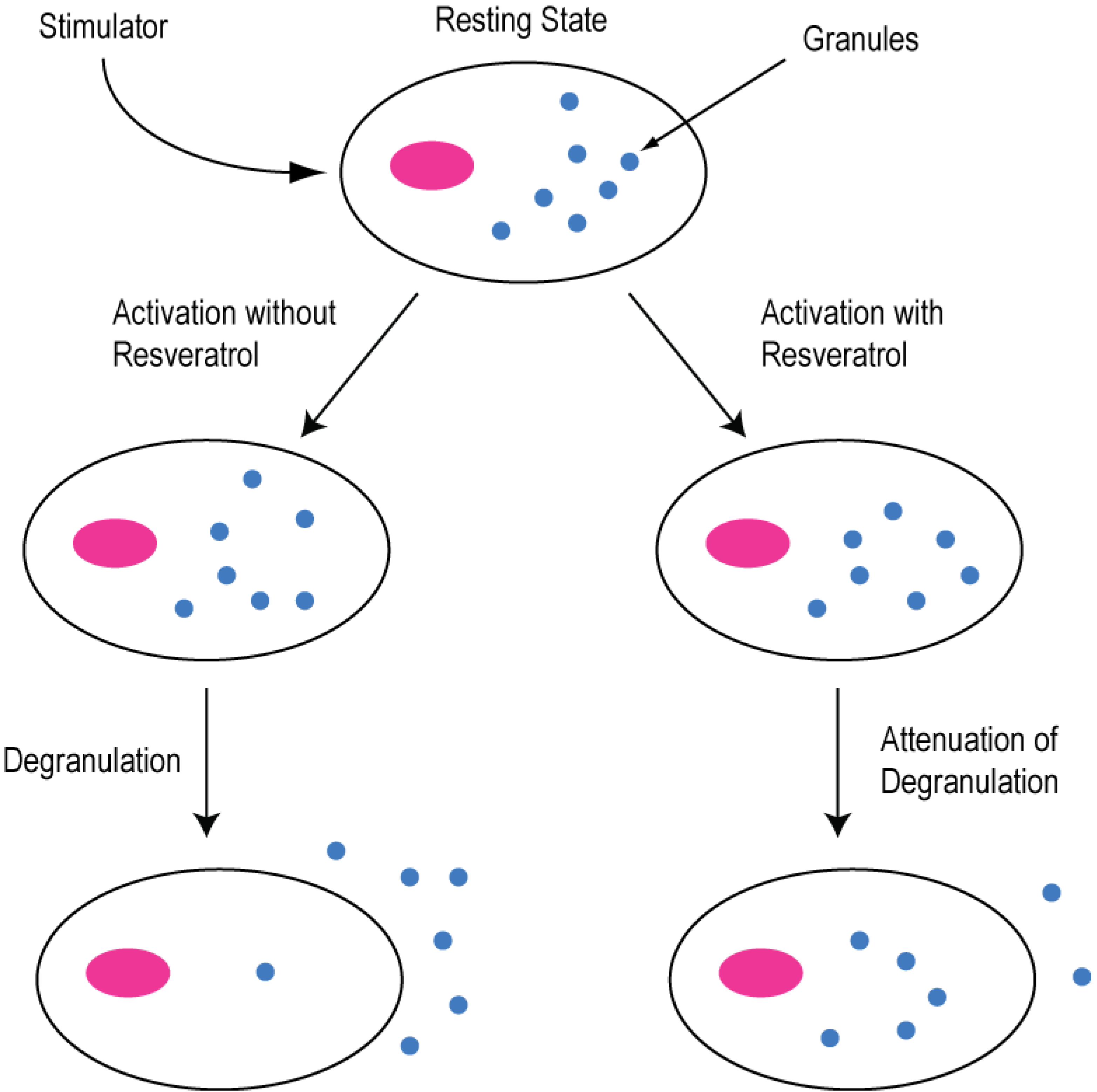
2.2. Modulation of Cellular Calcium Signaling Mechanisms by Resveratrol in Cancer and Immune Cells
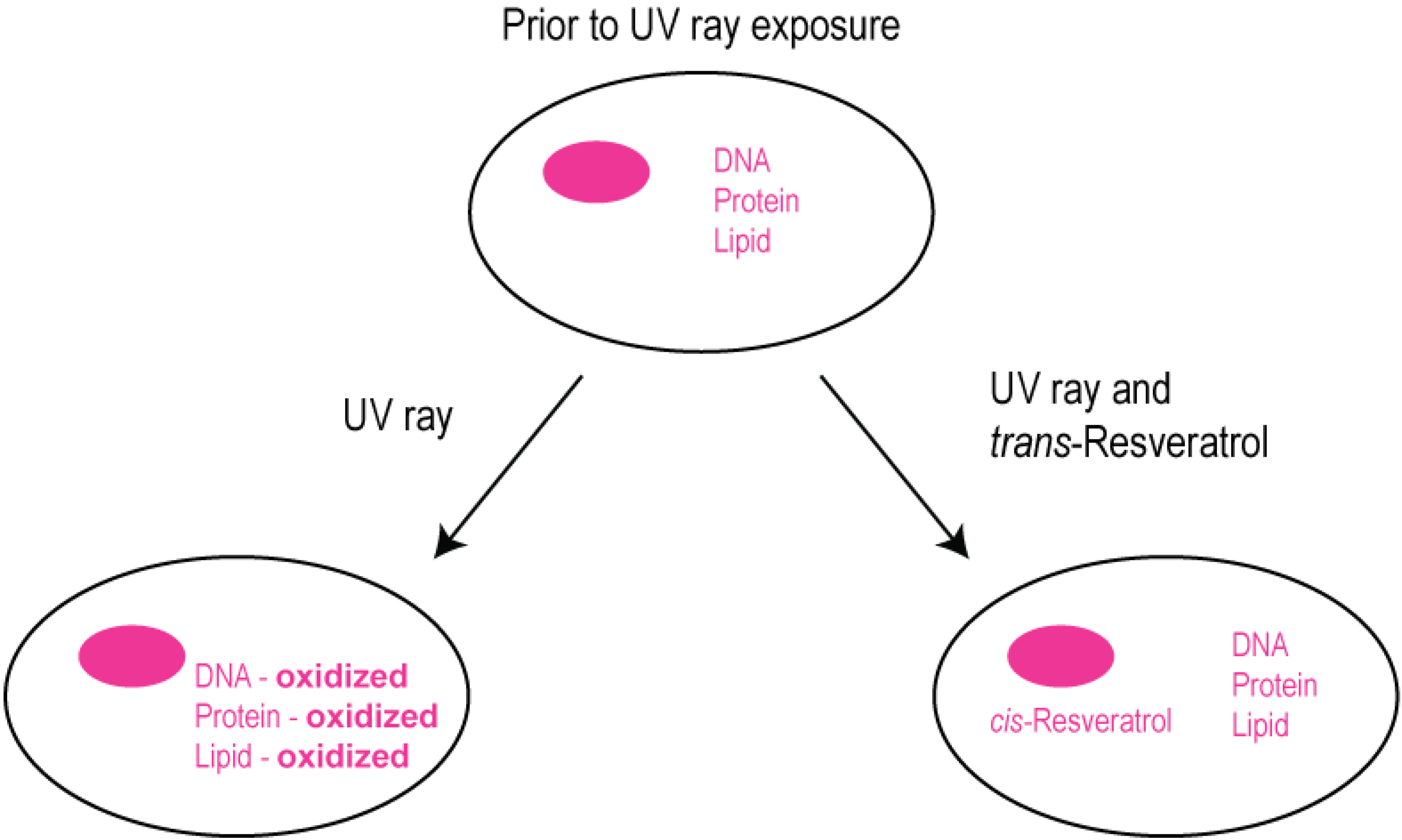
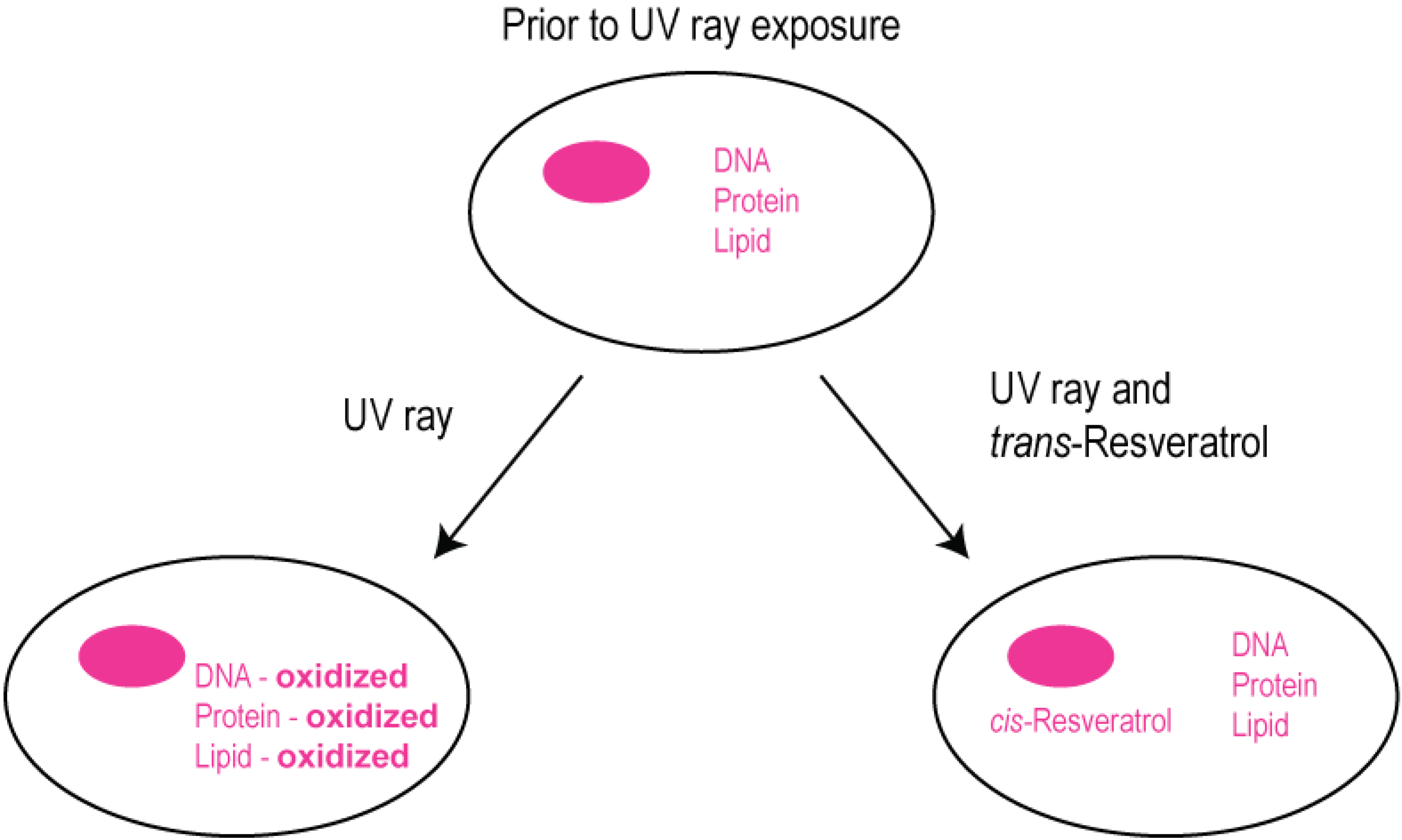
2.3. Modulation of Cellular Calcium Signaling Mechanisms by Resveratrol in Human Retinal Pigment Epithelial Cells
3. Therapeutic Potential of Resveratrol via the Modulation of Cellular Calcium Signaling
3.1. Intracellular Calcium Channels
3.2. Store-Operated Calcium Channel
3.3. Voltage-Gated Calcium Channels
3.4. Calcium-Activated Potassium Channels

{kind=link}
{kind=link}
{kind=link}
| Protein | Modulatory Action | Therapeutic Application | References |
|---|---|---|---|
| Intracellular calciumchannels | TBD | Potential for excitatory neuronal, cardiac, inflammatory and autoimmune diseases | --- |
| Calcium-release activated channels | No direct effects | No direct disease amelioration | [60] |
| Store-operated calcium entry | Activation of store-operated calcium entry | Diseases of prolonged calcium influx such as immune and inflammatory diseases | [33] |
| Voltage-gated calcium channels | Dose-dependent inhibition of L- and T-type channels | Prevention of uncontrolled excitability | [26,27] |
| Calcium-activated potassium channels | Indirect inhibition, likely via modulation of voltage-gated calcium channels | Modulation of action potentials (particularly in cardiac and neurological disorders) | [19,47,74] |
| SERCA | Indirect up-regulation via SIRT1 activation | SIRT1 down regulation disorders, cancer | [26,27] |
| PMCA | Indirect PMCA degradation via calpain activation | Cancer | [27] |
3.5. SERCA and PMCA
3.6. Mitochondrial Calcium Signaling
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jeandet, P.; Delaunois, B.; Conreux, A.; Donnez, D.; Nuzzo, V.; Cordelier, S.; Clément, C.; Courot, E. Biosynthesis, metabolism, molecular engineering and biological functions of stilbene phytoalexins in plants. BioFactors 2010, 36, 331–341. [Google Scholar]
- Jeandet, P.; Clément, C.; Courot, E.; Cordelier, S. Modulation of phytoalexin biosynthesis in engineered plants for disease resistance. Int. J. Mol. Sci. 2013, 14, 14136–14170. [Google Scholar]
- Neves, A.R.; Lucio, M.; Lima, J.L.; Reis, S. Resveratrol in medicinal chemistry: A critical review of its pharmacokinetics, drug-delivery, and membrane interactions. Curr. Med. Chem. 2012, 19, 1663–1681. [Google Scholar]
- Richard, J.L. Coronary risk factors. The French paradox. Arch. Mal. Coeur Vaiss. 1987, 80, 17–21. [Google Scholar]
- St Leger, A.S.; Cochrane, A.L.; Moore, F. Factors associated with cardiac mortality in developed countries with particular reference to the consumption of wine. Lancet 1979, 1, 1017–1020. [Google Scholar]
- Criqui, M.H.; Ringel, B.L. Does diet or alcohol explain the French paradox? Lancet 1994, 344, 1719–1723. [Google Scholar]
- Kroon, P.A.; Iyer, A.; Chunduri, P.; Chan, V.; Brown, L. The cardiovascular nutrapharmacology of resveratrol: Pharmacokinetics, molecular mechanisms and therapeutic potential. Curr. Med. Chem. 2010, 17, 2442–2455. [Google Scholar]
- Bagatini, P.B.; Saur, L.; Rodrigues, M.F.; Bernardino, G.C.; Paim, M.F.; Coelho, G.P.; Silva, D.V.; de Oliveira, R.M.; Schirmer, H.; Souto, A.A.; et al. The role of calcium channel blockers and resveratrol in the prevention of paraquat-induced parkinsonism in Drosophila melanogaster: A locomotor analysis. Invertebr. Neurosci. 2011, 11, 43–51. [Google Scholar]
- Granados-Soto, V.; Arguelles, C.F.; Ortiz, M.I. The peripheral antinociceptive effect of resveratrol is associated with activation of potassium channels. Neuropharmacology 2002, 43, 917–923. [Google Scholar]
- Park, S.J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar]
- Athar, M.; Back, J.H.; Tang, X.; Kim, K.H.; Kopelovich, L.; Bickers, D.R.; Kim, A.L. Resveratrol: A review of preclinical studies for human cancer prevention. Toxicol. Appl. Pharm. 2007, 224, 274–283. [Google Scholar]
- Yuan, M.; Li, J.; Lv, J.; Mo, X.; Yang, C.; Chen, X.; Liu, Z.; Liu, J. Polydatin (PD) inhibits IgE-mediated passive cutaneous anaphylaxis in mice by stabilizing mast cells through modulating Ca(2)(+) mobilization. Toxicol. Appl. Pharm. 2012, 264, 462–469. [Google Scholar]
- Venkatesan, B.; Valente, A.J.; Reddy, V.S.; Siwik, D.A.; Chandrasekar, B. Resveratrol blocks interleukin-18-EMMPRIN cross-regulation and smooth muscle cell migration. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H874–H886. [Google Scholar]
- Naylor, J.; Al-Shawaf, E.; McKeown, L.; Manna, P.T.; Porter, K.E.; O’Regan, D.; Muraki, K.; Beech, D.J. TRPC5 channel sensitivities to antioxidants and hydroxylated stilbenes. J. Biol. Chem. 2011, 286, 5078–5086. [Google Scholar]
- Wang, Q.; Xu, J.; Rottinghaus, G.E.; Simonyi, A.; Lubahn, D.; Sun, G.Y.; Sun, A.Y. Resveratrol protects against global cerebral ischemic injury in gerbils. Brain Res. 2002, 958, 439–447. [Google Scholar]
- Zhang, H.; Schools, G.P.; Lei, T.; Wang, W.; Kimelberg, H.K.; Zhou, M. Resveratrol attenuates early pyramidal neuron excitability impairment and death in acute rat hippocampal slices caused by oxygen-glucose deprivation. Exp. Neurol. 2008, 212, 44–52. [Google Scholar]
- Chen, W.P.; Su, M.J.; Hung, L.M. In vitro electrophysiological mechanisms for antiarrhythmic efficacy of resveratrol, a red wine antioxidant. Eur. J. Pharmacol. 2007, 554, 196–204. [Google Scholar]
- Chen, Y.R.; Yi, F.F.; Li, X.Y.; Wang, C.Y.; Chen, L.; Yang, X.C.; Su, P.X.; Cai, J. Resveratrol attenuates ventricular arrhythmias and improves the long-term survival in rats with myocardial infarction. Cardiovasc. Drug. Ther. 2008, 22, 479–485. [Google Scholar]
- Calderone, V.; Martelli, A.; Testai, L.; Martinotti, E.; Breschi, M.C. Functional contribution of the endothelial component to the vasorelaxing effect of resveratrol and NS 1619, activators of the large-conductance calcium-activated potassium channels. Naunyn-Schmiedeberg’s Arch. Pharm. 2007, 375, 73–80. [Google Scholar]
- Gojkovic-Bukarica, L.; Novakovic, A.; Kanjuh, V.; Bumbasirevic, M.; Lesic, A.; Heinle, H. A role of ion channels in the endothelium-independent relaxation of rat mesenteric artery induced by resveratrol. J. Pharmacol. Sci. 2008, 108, 124–130. [Google Scholar]
- Gao, X.; Xu, Y.X.; Janakiraman, N.; Chapman, R.A.; Gautam, S.C. Immunomodulatory activity of resveratrol: Suppression of lymphocyte proliferation, development of cell-mediated cytotoxicity, and cytokine production. Biochem. Pharmacol. 2001, 62, 1299–1308. [Google Scholar]
- Sharma, S.; Anjaneyulu, M.; Kulkarni, S.K.; Chopra, K. Resveratrol, a polyphenolic phytoalexin, attenuates diabetic nephropathy in rats. Pharmacology 2006, 76, 69–75. [Google Scholar]
- Sheu, S.J.; Wu, T.T. Resveratrol protects against ultraviolet A-mediated inhibition of the phagocytic function of human retinal pigment epithelial cells via large-conductance calcium-activated potassium channels. Kaohsiung J. Med. Sci. 2009, 25, 381–388. [Google Scholar]
- Sheu, S.J.; Bee, Y.S.; Chen, C.H. Resveratrol and large-conductance calcium-activated potassium channels in the protection of human retinal pigment epithelial cells. J. Ocul. Pharmacol. 2008, 24, 551–555. [Google Scholar]
- Pirola, L.; Frojdo, S. Resveratrol: One molecule, many targets. IUBMB Life 2008, 60, 323–332. [Google Scholar]
- Sulaiman, M.; Matta, M.J.; Sunderesan, N.R.; Gupta, M.P.; Periasamy, M.; Gupta, M. Resveratrol, an activator of SIRT1, upregulates sarcoplasmic calcium ATPase and improves cardiac function in diabetic cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H833–H843. [Google Scholar]
- Sareen, D.; Darjatmoko, S.R.; Albert, D.M.; Polans, A.S. Mitochondria, calcium, and calpain are key mediators of resveratrol-induced apoptosis in breast cancer. Mol. Pharmacol. 2007, 72, 1466–1475. [Google Scholar]
- Duncan, R.S.; Goad, D.L.; Grillo, M.A.; Kaja, S.; Payne, A.J.; Koulen, P. Control of intracellular calcium signaling as a neuroprotective strategy. Molecules 2010, 15, 1168–1195. [Google Scholar]
- Jakab, M.; Lach, S.; Bacova, Z.; Langeluddecke, C.; Strbak, V.; Schmidt, S.; Iglseder, E.; Paulmichl, M.; Geibel, J.; Ritter, M. Resveratrol inhibits electrical activity and insulin release from insulinoma cells by block of voltage-gated Ca+ channels and swelling-dependent Cl- currents. Cell Physiol. Biochem. 2008, 22, 567–578. [Google Scholar]
- Zhang, L.P.; Yin, J.X.; Liu, Z.; Zhang, Y.; Wang, Q.S.; Zhao, J. Effect of resveratrol on L-type calcium current in rat ventricular myocytes. Acta Pharmacol. Sin. 2006, 27, 179–183. [Google Scholar]
- Dobrydneva, Y.; Williams, R.L.; Blackmore, P.F. trans-Resveratrol inhibits calcium influx in thrombin-stimulated human platelets. Br. J. Pharmacol. 1999, 128, 149–157. [Google Scholar]
- Tian, X.M.; Zhang, Z.X. [Resveratrol promote permeability transition pore opening mediated by Ca2+]. Yao Xue Xue Bao 2003, 38, 81–84. [Google Scholar]
- Dobrydneva, Y.; Williams, R.L.; Blackmore, P.F. Diethylstilbestrol and other nonsteroidal estrogens: Novel class of store-operated calcium channel modulators. J. Cardiovasc. Pharmacol. 2010, 55, 522–530. [Google Scholar]
- Dobrydneva, Y.; Williams, R.L.; Katzenellenbogen, J.A.; Ratz, P.H.; Blackmore, P.F. Diethylstilbestrol and tetrahydrochrysenes are calcium channel blockers in human platelets: Relationship to the stilbene pharmacophore. Thromb. Res. 2003, 110, 23–31. [Google Scholar]
- Wang, L.; Ma, Q.; Chen, X.; Sha, H.; Ma, Z. Effects of resveratrol on calcium regulation in rats with severe acute pancreatitis. Eur. J. Pharmacol. 2008, 580, 271–276. [Google Scholar]
- Buluc, M.; Demirel-Yilmaz, E. Resveratrol decreases calcium sensitivity of vascular smooth muscle and enhances cytosolic calcium increase in endothelium. Vascul. Pharmacol. 2006, 44, 231–237. [Google Scholar]
- Campos-Toimil, M.; Elies, J.; Alvarez, E.; Verde, I.; Orallo, F. Effects of trans- and cis-resveratrol on Ca2+ handling in A7r5 vascular myocytes. Eur. J. Pharmacol. 2007, 577, 91–99. [Google Scholar]
- Sangeetha, M.K.; Eazhisai Vallabi, D.; Sali, V.K.; Thanka, J.; Vasanthi, H.R. Sub-acute toxicity profile of a modified resveratrol supplement. Food Chem. Toxicol. 2013, 59, 492–500. [Google Scholar]
- Cottart, C.H.; Nivet-Antoine, V.; Beaudeux, J.L. Review of recent data on the metabolism, biological effects, and toxicity of resveratrol in humans. Mol. Nutr. Food Res. 2013, 58, 7–21. [Google Scholar]
- Zhang, L.P.; Ma, H.J.; Zhao, J.; Wang, Q.S. Effects of resveratrol on delayed afterdepolarization and triggered activity induced by ouabain in guinea pig papillary muscles. Sheng Li Xue Bao 2005, 57, 361–366. [Google Scholar]
- Liu, Z.; Zhang, L.P.; Ma, H.J.; Wang, C.; Li, M.; Wang, Q.S. Resveratrol reduces intracellular free calcium concentration in rat ventricular myocytes. Sheng Li Xue Bao 2005, 57, 599–604. [Google Scholar]
- Campos-Toimil, M.; Elies, J.; Orallo, F. Trans- and cis-resveratrol increase cytoplasmic calcium levels in A7r5 vascular smooth muscle cells. Mol. Nutr. Food Res. 2005, 49, 396–404. [Google Scholar]
- Li, H.; Xia, N.; Forstermann, U. Cardiovascular effects and molecular targets of resveratrol. Nitric Oxide 2012, 26, 102–110. [Google Scholar]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discovery 2006, 5, 493–506. [Google Scholar]
- Elíes, J.; Cuíñas, A.; García-Morales, V.; Orallo, F.; Campos-Toimil, M. Trans-resveratrol simultaneously increases cytoplasmic Ca2+ levels and nitric oxide release in human endothelial cells. Mol. Nutr. Food Res. 2011, 55, 1237–1248. [Google Scholar]
- Protic, D.; Beleslin-Cokic, B.; Novakovic, R.; Kanjuh, V.; Heinle, H.; Scepanovic, R.; Gojkovic-Bukarica, L. Effect of Wine Polyphenol Resveratrol on the Contractions Elicited Electrically or by Norepinephrine in the Rat Portal Vein. Phytother. Res. 2013, 27, 1685–1693. [Google Scholar]
- Novakovic, A.; Bukarica, L.G.; Kanjuh, V.; Heinle, H. Potassium channels-mediated vasorelaxation of rat aorta induced by resveratrol. Basic Clin. Pharmacol. Toxicol. 2006, 99, 360–364. [Google Scholar]
- Su, H.C.; Hung, L.M.; Chen, J.K. Resveratrol, a red wine antioxidant, possesses an insulin-like effect in streptozotocin-induced diabetic rats. Am. J. Physiol. Endocrinol. Metab. 2006, 290, 1339–1346. [Google Scholar]
- Raval, A.P.; Dave, K.R.; Perez-Pinzon, M.A. Resveratrol mimics ischemic preconditioning in the brain. J. Cereb. Blood Flow Metab. 2006, 26, 1141–1147. [Google Scholar]
- Raval, A.P.; Lin, H.W.; Dave, K.R.; Defazio, R.A.; Della Morte, D.; Kim, E.J.; Perez-Pinzon, M.A. Resveratrol and ischemic preconditioning in the brain. Curr. Med. Chem. 2008, 15, 1545–1551. [Google Scholar]
- Li, M.; Wang, Q.S.; Chen, Y.; Wang, Z.M.; Liu, Z.; Guo, S.M. Resveratrol inhibits neuronal discharges in rat hippocampal CA1 area. Sheng Li Xue Bao 2005, 57, 355–360. [Google Scholar]
- Kundu, J.K.; Surh, Y.J. Cancer chemopreventive and therapeutic potential of resveratrol: Mechanistic perspectives. Cancer Lett. 2008, 269, 243–261. [Google Scholar]
- Le Ferrec, E.; Lagadic-Gossmann, D.; Rauch, C.; Bardiau, C.; Maheo, K.; Massiere, F.; le Vee, M.; Guillouzo, A.; Morel, F. Transcriptional induction of CYP1A1 by oltipraz in human Caco-2 cells is aryl hydrocarbon receptor- and calcium-dependent. J. Biol. Chem. 2002, 277, 24780–24787. [Google Scholar]
- Chen, Y.F.; Chen, Y.T.; Chiu, W.T.; Shen, M.R. Remodeling of calcium signaling in tumor progression. J. Biomed. Sci. 2013, 20. [Google Scholar] [CrossRef]
- Ma, X.; Tian, X.; Huang, X.; Yan, F.; Qiao, D. Resveratrol-induced mitochondrial dysfunction and apoptosis are associated with Ca2+ and mCICR-mediated MPT activation in HepG2 cells. Mol. Cell. Biochem. 2007, 302, 99–109. [Google Scholar]
- Yang, Y.; Paik, J.H.; Cho, D.; Cho, J.A.; Kim, C.W. Resveratrol induces the suppression of tumor-derived CD4+CD25+ regulatory T cells. Int. Immunopharmacol. 2008, 8, 542–547. [Google Scholar]
- Bishayee, A. Cancer prevention and treatment with resveratrol: From rodent studies to clinical trials. Cancer Prev. Res. 2009, 2, 409–418. [Google Scholar]
- Feng, Y.H.; Zhou, W.L.; Wu, Q.L.; Li, X.Y.; Zhao, W.M.; Zou, J.P. Low dose of resveratrol enhanced immune response of mice. Acta Pharmacol. Sin. 2002, 23, 893–897. [Google Scholar]
- Falchetti, R.; Fuggetta, M.P.; Lanzilli, G.; Tricarico, M.; Ravagnan, G. Effects of resveratrol on human immune cell function. Life Sci. 2001, 70, 81–96. [Google Scholar]
- Shin, D.H.; Seo, E.Y.; Pang, B.; Nam, J.H.; Kim, H.S.; Kim, W.K.; Kim, S.J. Inhibition of Ca2+-release-activated Ca2+ channel (CRAC) and K+ channels by curcumin in Jurkat-T cells. J. Pharmacol. Sci. 2011, 115, 144–154. [Google Scholar]
- Singh, N.P.; Hegde, V.L.; Hofseth, L.J.; Nagarkatti, M.; Nagarkatti, P. Resveratrol (trans-3,5,4'-trihydroxystilbene) ameliorates experimental allergic encephalomyelitis, primarily via induction of apoptosis in T cells involving activation of aryl hydrocarbon receptor and estrogen receptor. Mol. Pharmacol. 2007, 72, 1508–1521. [Google Scholar]
- Nour, V.; Trandafir, I.; Muntean, C. Ultraviolet irradiation of trans-resveratrol and HPLC determination of trans-resveratrol and cis-resveratrol in Romanian red wines. J. Chromatogr. Sci. 2012, 50, 920–927. [Google Scholar]
- Bertram, K.M.; Baglole, C.J.; Phipps, R.P.; Libby, R.T. Molecular regulation of cigarette smoke induced-oxidative stress in human retinal pigment epithelial cells: Implications for age-related macular degeneration. Am. J. Physiol. Cell Physiol. 2009, 297, C1200–C1210. [Google Scholar]
- King, R.E.; Kent, K.D.; Bomser, J.A. Resveratrol reduces oxidation and proliferation of human retinal pigment epithelial cells via extracellular signal-regulated kinase inhibition. Chem. Biol. Interact. 2005, 151, 143–149. [Google Scholar]
- Nagaoka, T.; Hein, T.W.; Yoshida, A.; Kuo, L. Resveratrol, a component of red wine, elicits dilation of isolated porcine retinal arterioles: Role of nitric oxide and potassium channels. Invest. Ophthalmol. Vis. Sci. 2007, 48, 4232–4239. [Google Scholar]
- Berridge, M.J. Neuronal calcium signaling. Neuron 1998, 21, 13–26. [Google Scholar]
- Wehrens, X.H.; Lehnart, S.E.; Marks, A.R. Intracellular calcium release and cardiac disease. Annu. Rev. Physiol. 2005, 67, 69–98. [Google Scholar]
- Feske, S. Calcium signalling in lymphocyte activation and disease. Nat. Rev. Immunol. 2007, 7, 690–702. [Google Scholar]
- Dorrie, J.; Gerauer, H.; Wachter, Y.; Zunino, S.J. Resveratrol induces extensive apoptosis by depolarizing mitochondrial membranes and activating caspase-9 in acute lymphoblastic leukemia cells. Cancer Res. 2001, 61, 4731–4739. [Google Scholar]
- Wallace, C.H.; Baczko, I.; Jones, L.; Fercho, M.; Light, P.E. Inhibition of cardiac voltage-gated sodium channels by grape polyphenols. Br. J. Pharmacol. 2006, 149, 657–665. [Google Scholar]
- Anekonda, T.S.; Quinn, J.F.; Harris, C.; Frahler, K.; Wadsworth, T.L.; Woltjer, R.L. L-type voltage-gated calcium channel blockade with isradipine as a therapeutic strategy for Alzheimer’s disease. Neurobiol. Dis. 2011, 41, 62–70. [Google Scholar]
- Cain, S.M.; Snutch, T.P. Voltage-gated calcium channels and disease. Biofactors 2011, 37, 197–205. [Google Scholar]
- Vergara, C.; Latorre, R.; Marrion, N.V.; Adelman, J.P. Calcium-activated potassium channels. Curr. Opin. Neurobiol. 1998, 8, 321–329. [Google Scholar]
- Chang, Y.; Wang, S.J. Inhibitory effect of glutamate release from rat cerebrocortical nerve terminals by resveratrol. Neurochem. Int. 2009, 54, 135–141. [Google Scholar]
- Chen, W.-P.; Chi, T.-C.; Chuang, L.-M.; Su, M.-J. Resveratrol enhances insulin secretion by blocking KATP and KV channels of beta cells. Eur. J. Pharmacol. 2007, 568, 269–277. [Google Scholar]
- Kalra, N.; Roy, P.; Prasad, S.; Shukla, Y. Resveratrol induces apoptosis involving mitochondrial pathways in mouse skin tumorigenesis. Life Sci. 2008, 82, 348–358. [Google Scholar]
- Sun, W.; Wang, W.; Kim, J.; Keng, P.; Yang, S.; Zhang, H.; Liu, C.; Okunieff, P.; Zhang, L. Anti-cancer effect of resveratrol is associated with induction of apoptosis via a mitochondrial pathway alignment. Adv. Exp. Med. Biol. 2008, 614, 179–186. [Google Scholar]
- Schmidt, A.; Oberle, N.; Weiß, E.M.; Vobis, D.; Frischbutter, S.; Baumgrass, R.; Falk, C.S.; Haag, M.; Brügger, B.; Lin, H.; et al. Human regulatory T cells rapidly suppress T cell receptor-induced Ca2+, NF-kappaB, and NFAT signaling in conventional T cells. Sci. Signal. 2011, 4. [Google Scholar] [CrossRef]
- Qureshi, A.A.; Guan, X.Q.; Reis, J.C.; Papasian, C.J.; Jabre, S.; Morrison, D.C.; Qureshi, N. Inhibition of nitric oxide and inflammatory cytokines in LPS-stimulated murine macrophages by resveratrol, a potent proteasome inhibitor. Lipids Health Dis. 2012, 11, 76. [Google Scholar]
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
McCalley, A.E.; Kaja, S.; Payne, A.J.; Koulen, P. Resveratrol and Calcium Signaling: Molecular Mechanisms and Clinical Relevance. Molecules 2014, 19, 7327-7340. https://doi.org/10.3390/molecules19067327
McCalley AE, Kaja S, Payne AJ, Koulen P. Resveratrol and Calcium Signaling: Molecular Mechanisms and Clinical Relevance. Molecules. 2014; 19(6):7327-7340. https://doi.org/10.3390/molecules19067327
Chicago/Turabian StyleMcCalley, Audrey E., Simon Kaja, Andrew J. Payne, and Peter Koulen. 2014. "Resveratrol and Calcium Signaling: Molecular Mechanisms and Clinical Relevance" Molecules 19, no. 6: 7327-7340. https://doi.org/10.3390/molecules19067327
APA StyleMcCalley, A. E., Kaja, S., Payne, A. J., & Koulen, P. (2014). Resveratrol and Calcium Signaling: Molecular Mechanisms and Clinical Relevance. Molecules, 19(6), 7327-7340. https://doi.org/10.3390/molecules19067327