Evaluation of the Efficiency of Different Disruption Methods on Yeast Cell Wall Preparation for β-Glucan Isolation

Abstract
:1. Introduction
2. Results and Discussion
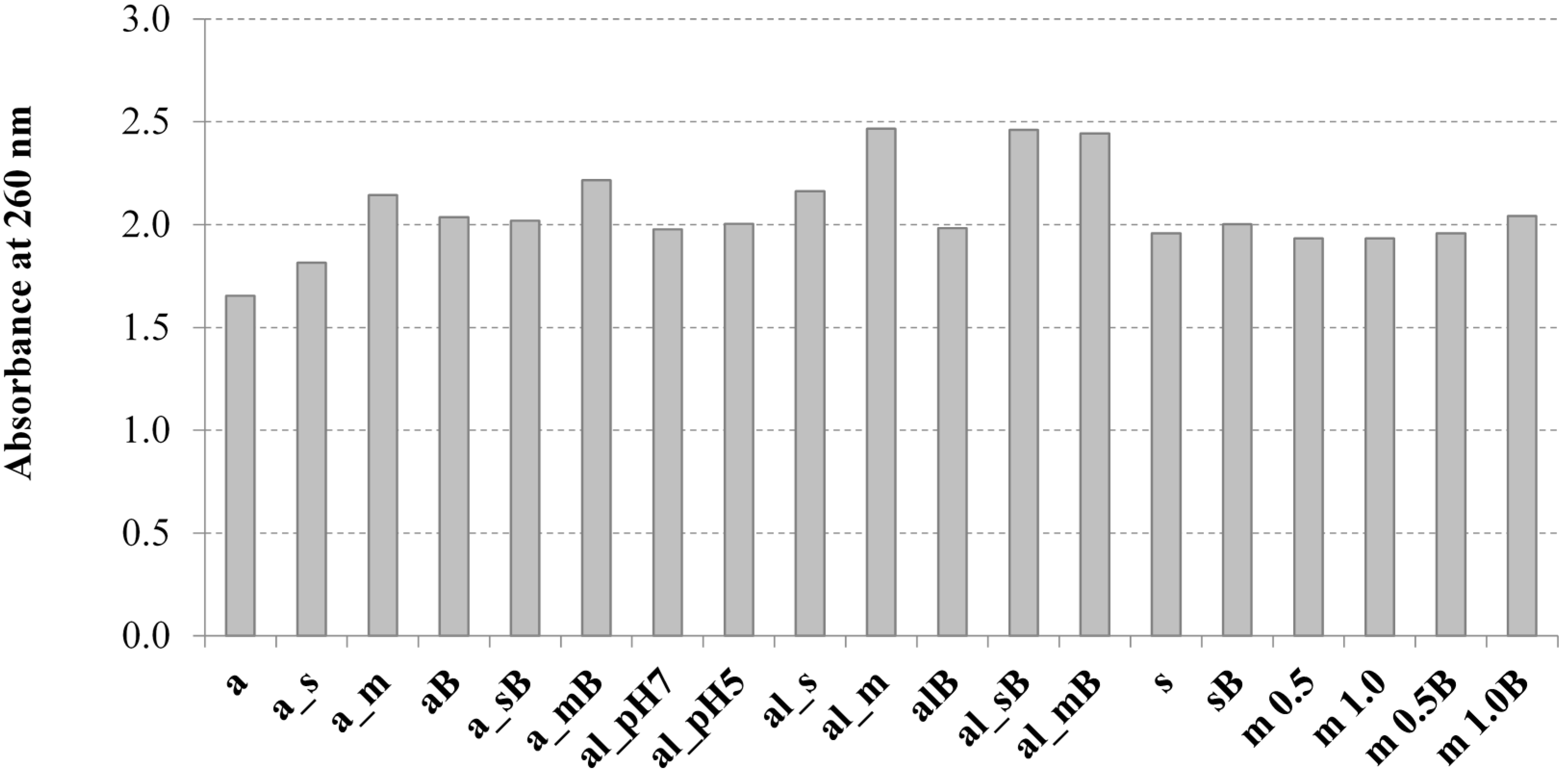
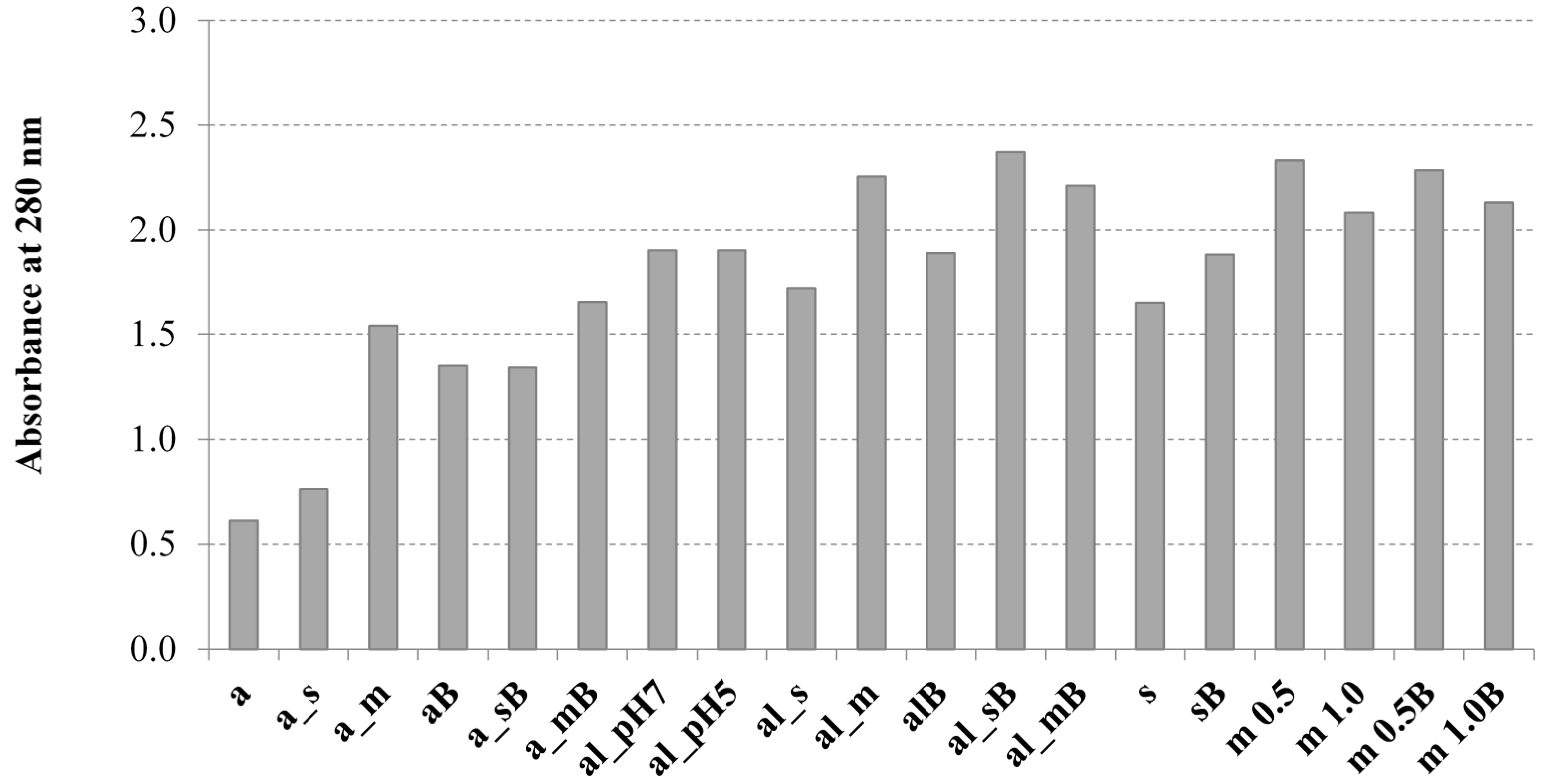
2.1. The Influence of the Tested Methods on the Solubilization of Yeast Cell Biomass

{kind=link}
{kind=link}
{kind=link}
{kind=link}
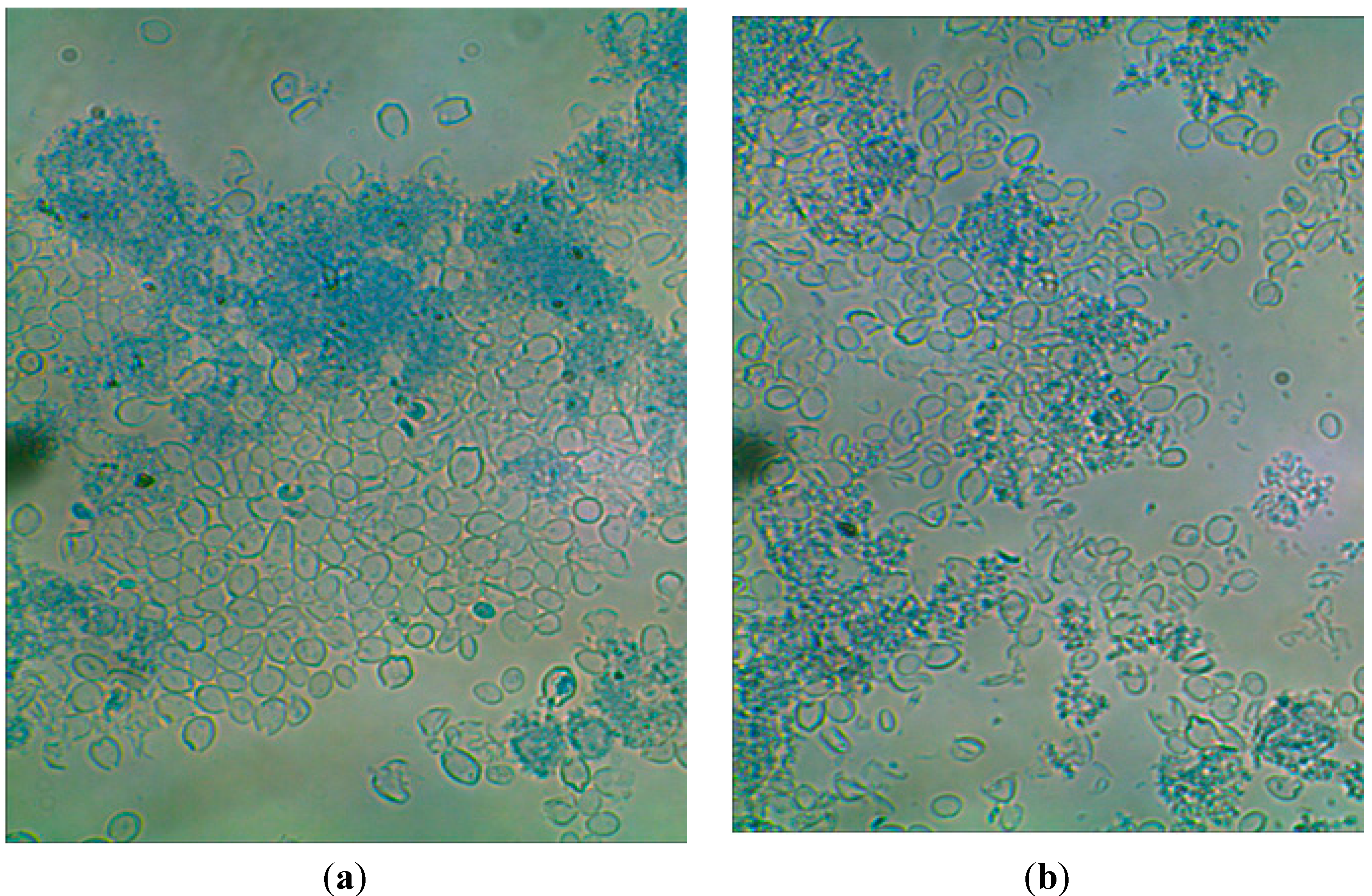{kind=link}
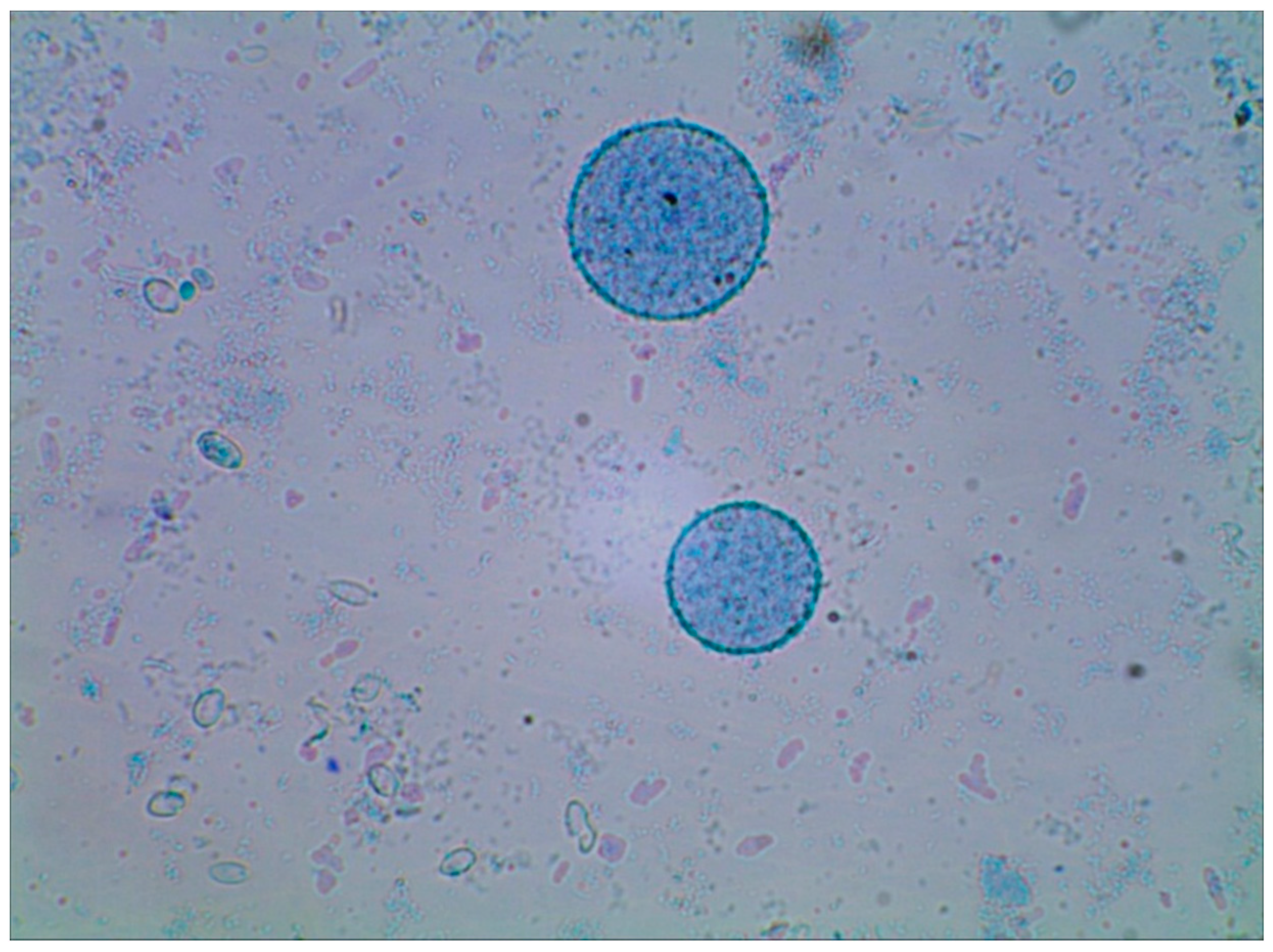{kind=link}
{kind=link}
| Experimental System | Solubilised Material (%) | Total Saccharides | β(1,3)/(1,6)-glucan | Crude Proteins |
|---|---|---|---|---|
| ± SD (g/100 g d.m. Preparation) | ||||
| Biomass | - | 42.5 ± 2.0 A,B | 7.7 ± 0.3 B | 45.3 ± 2.6 F |
| a * | 18.4 ± 0.6 A,** | 47.0 ± 3.7 A,B,C,D | 5.7 ± 0.1 A | 53.6 ± 1.5 F,G |
| a_s | 25.5 ± 0.5 B | 39.4 ± 5.6 A | 6.1 ± 0.4 A | 54.3 ± 0.6 F,G |
| a_m | 30.5 ± 0.5 D | 41.5 ± 4.8 A | 6.4 ± 0.4 A | 55.8 ± 1.1 G |
| aB | 27.8 ± 0.3 C | 44.8 ± 3.4 A,B,C | 5.7 ± 0.3 A | 51.5 ± 0.5 F |
| a_sB | 26.3 ± 0.5 B,C | 45.0 ± 0.9 A,B,C | 5.8 ± 0.4 A | 54.2 ± 0.3 F,G |
| a_mB | 41.4 ± 0.4 E | 43.7 ± 1.6 A,B | 6.6 ± 0.1 A | 56.0 ± 0.6 G |
| al_pH7 | 55.6 ± 1.2 H,I | 57.2 ± 1.4 D,F,G,H | 12.9 ± 0.4 D,E | 35.0 ± 0.1 A |
| al_pH5 | 53.5 ± 0.8 G,H | 63.5 ± 1.7 G,H | 12.5 ± 0.4 D | 35.2 ± 0.7 A |
| al_s | 52.6 ± 0.8 F,G | 60.5 ± 1.2 F,G,H | 15.5 ± 0.4 G | 36.5 ± 0.5 A,B |
| al_m | 62.9 ± 0.5 K,L | 61.2 ± 1.7 F,G,H | 14.5 ± 0.6 F,G | 36.7 ± 0.6 A,B |
| alB | 56.1 ± 0.3 I | 64.4 ± 1.5 H | 12.0 ± 0.2 C,D | 36.2 ± 0.7 A,B |
| al_sB | 60.5 ± 0.5 J | 62.5 ± 2.9 F,G,H | 12.2 ± 0.2 C,D | 36.6 ± 0.3 A,B |
| al_mB | 60.7 ± 1.5 J | 57.3 ± 1.6 E,F,G,H | 11.2 ± 0.2 C | 42.9 ± 0.4 D,E,F |
| s | 33.2 ± 0.4 D | 47.9 ± 2.1A,B,C,D,E | 6.4 ± 0.2 A | 44.3 ± 0.1 E,F |
| sB | 52.7 ± 0.6 G | 58.8 ± 3.0 F,G,H | 12.9 ± 0.3 D,E | 40.8 ± 0.4 C,D,E |
| m 0.5 | 64.4 ± 0.8 L,M | 59.7 ± 2.8 F,G,H | 13.8 ± 0.6 E.F | 34.8 ± 0.8 A |
| m 1.0 | 61.2 ± 0.3 J,K | 56.3 ± 2.3C,D,E,F,G,H | 12.1 ± 0.5 C,D | 37.8 ± 0.9 A,B,C |
| m 0.5B | 66.5 ± 0.8 M | 53.1 ± 3.4 B,C,D,E,F | 14.1 ± 0.6 F | 37.4 ± 0.8 A,B,C |
| m 1.0B | 61.5 ± 0.5 J,K | 54.0 ± 5.1 C,D,E,F,G | 12.4 ± 0.3 D | 39.6 ± 0.6 B,C,D |
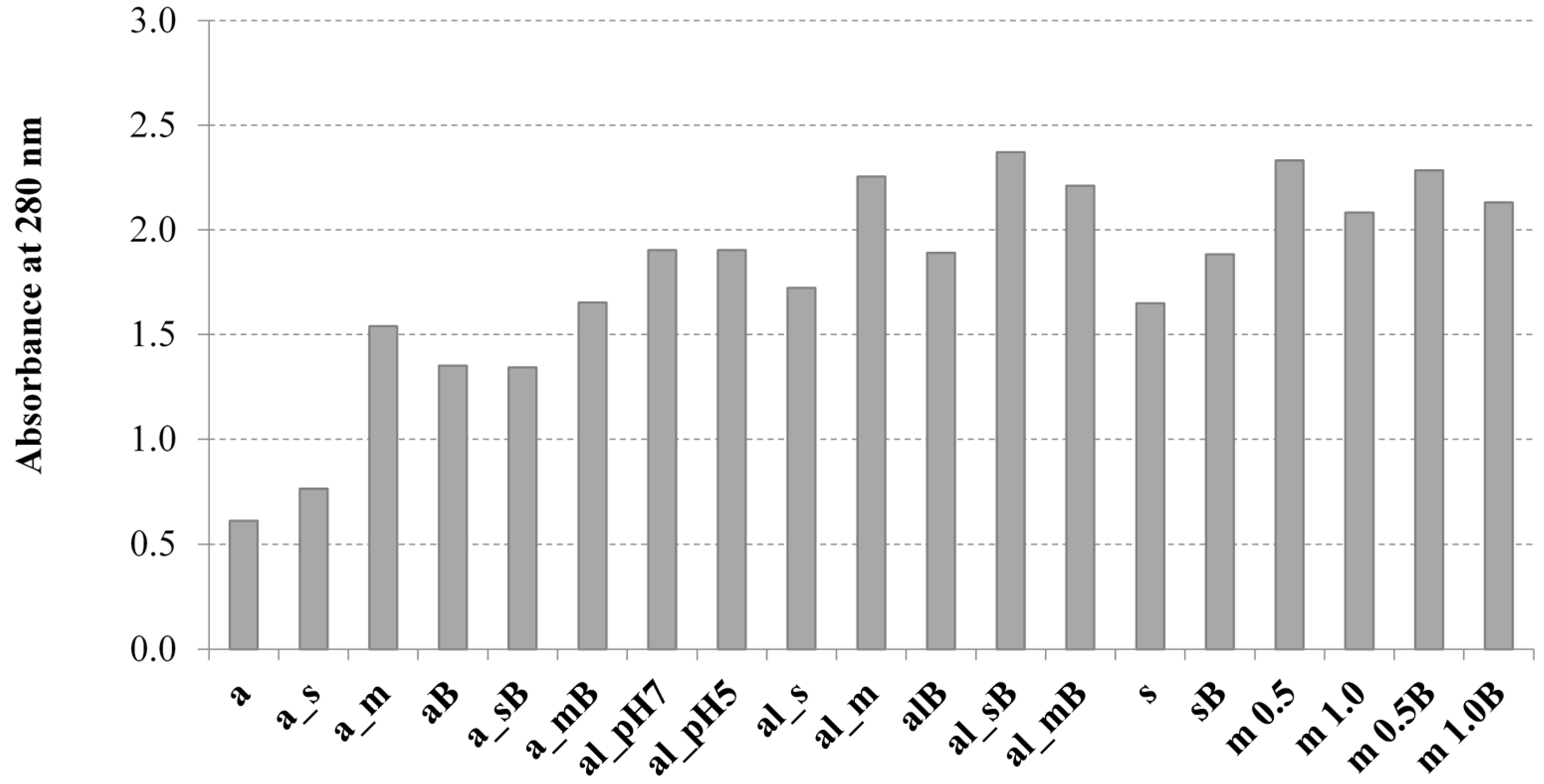
2.2. Effect of the Analyzed Disruption Methods on Contents of Total Saccharides, β(1,3)/(1,6)-Glucans, and Proteins in S. Cerevisiae Cell Wall Preparations
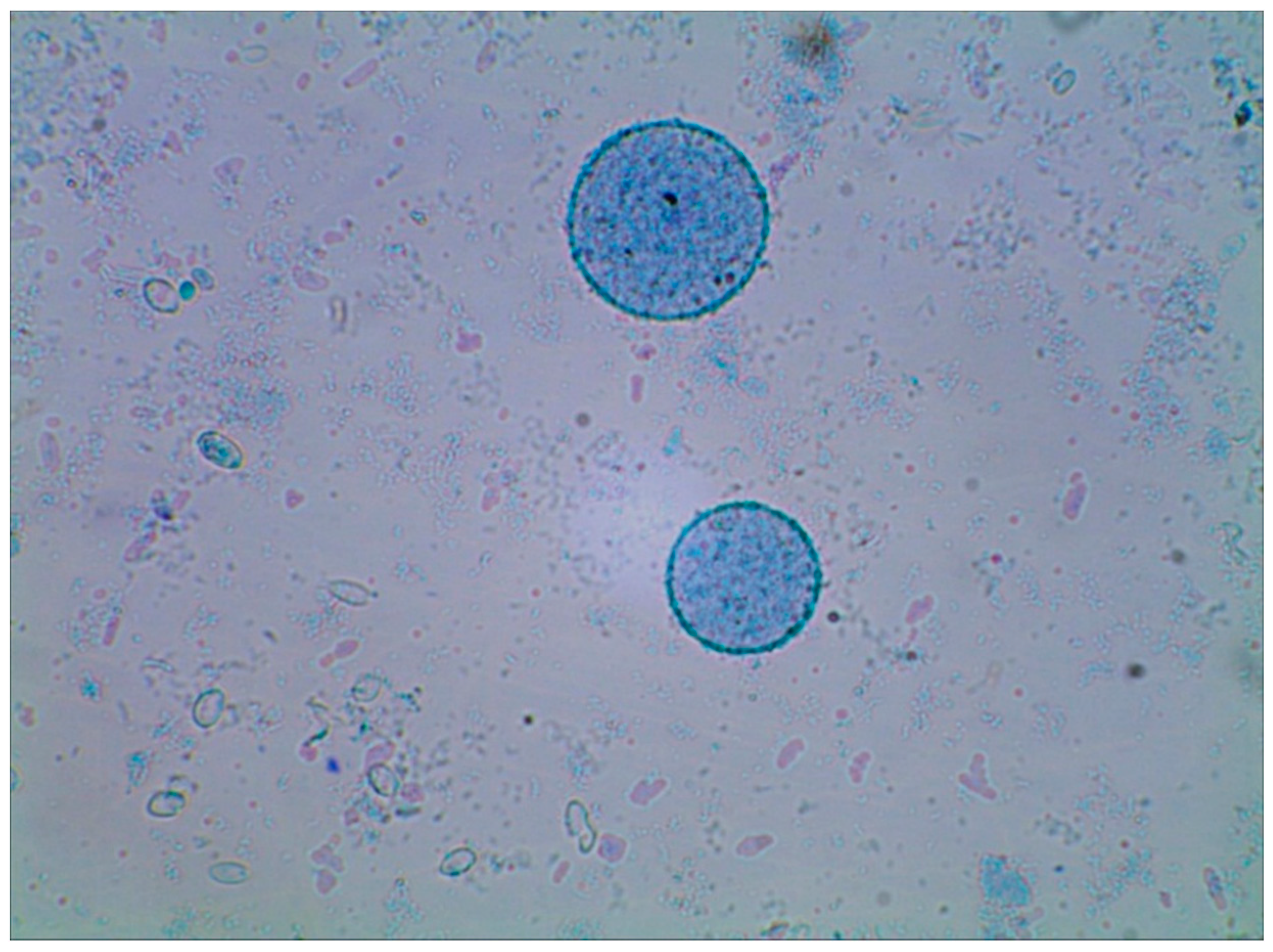
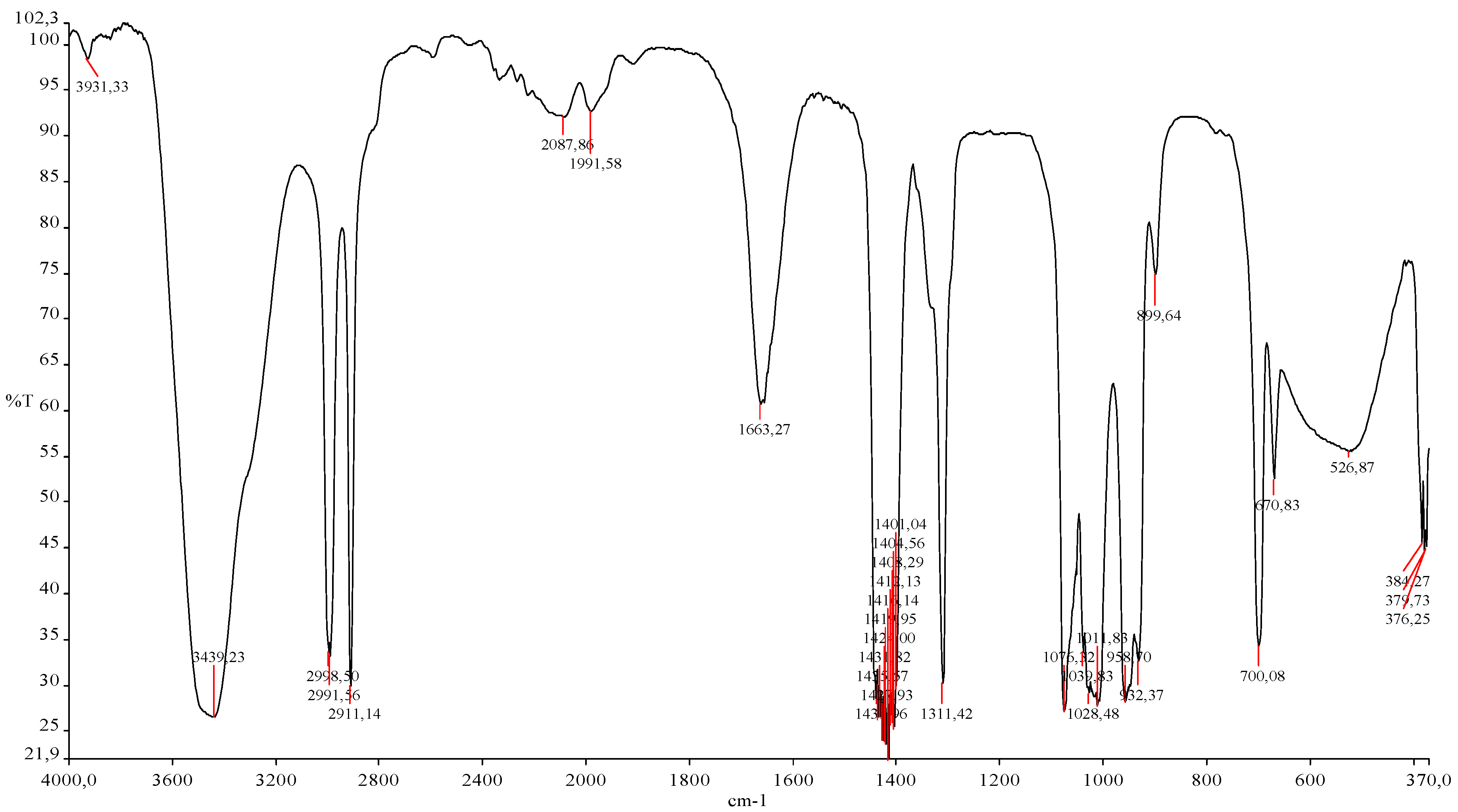
2.3. Spectral Analysis of Cell Wall Preparations Using Fourier Transform Infrared Spectroscopy (FTIR)
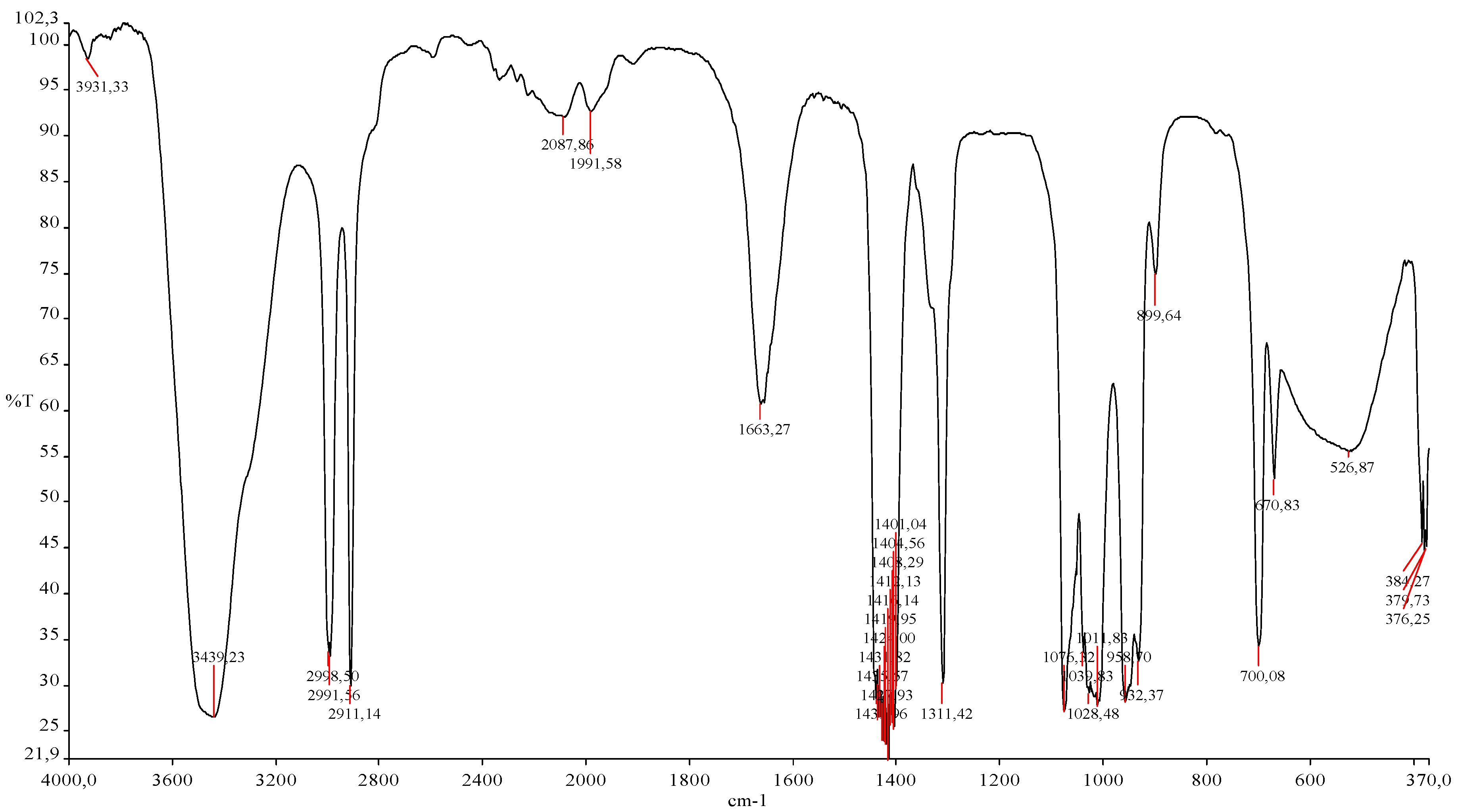
2.4. Discussion
3. Experimental Section
3.1. Biological Material
3.2. Preparation of Yeast Suspension for Disruption
3.3. Procedures for Yeast Cells Disruption with the Selected Methods
3.3.1. Autoclaving Process
3.3.2. Autolysis
3.3.3. Homogenization in a Bead Mill
| Type of Method | Experimental System | Preparation of Suspensions | |||
|---|---|---|---|---|---|
| Cell Suspensions in Deionized Water | Abbr. * | Cell Suspensions in 10 mM Tris-HCl Buffer (pH 8.0) | Abbr. * | ||
| Individual methods | Autoclaving | 115 °C, 10 min | a | 115 °C, 10 min | aB |
| Autolysis | pH 5.0, 50 °C, 24 h, 200 rpm | al_pH5 | 50 °C, 24 h, 200 rpm | alB | |
| Autolysis | pH 7.0, 50 °C, 24 h, 200 rpm | al_pH7 | |||
| Sonication | 4 × 5 min/2 min, pulser 80%, power 80% | s | 4 × 5 min/2 min, pulser 80%, power 80% | sB | |
| Bead mill | Ø 0.5 mm, 5 × 3 min/3 min | m 0.5 | Ø 0.5 mm, 5 × 3 min/3 min | m 0.5B | |
| Bead mill | Ø 1 mm, 5 × 3 min/3 min | m 1.0 | Ø 1 mm, 5 × 3 min/3 min | m 1.0B | |
| Coupled methods | Autoclaving | 115 °C, 10 min | a_s | 115 °C, 10 min | a_sB |
| Sonication | 4 × 5 min/2 min, pulser 80%, power 80% | 4 × 5 min/2 min, pulser 80%, power 80% | |||
| Autoclaving | 115 °C, 10 min | a_m | 115 °C, 10 min | a_mB | |
| Bead mill | Ø 0.5 mm, 5 × 3 min/3 min | Ø 0.5 mm, 5 × 3 min/3 min | |||
| Autolysis | pH 5.0, 50 °C, 24 h, 200 rpm | al_m | pH 5.0, 50 °C, 24 h, 200 rpm | al_mB | |
| Bead mill | Ø 0.5 mm, 5 × 3 min/3 min | Ø 0.5 mm, 5 × 3 min/3 min | |||
| Autolysis | pH 5.0, 50 °C, 24 h, 200 rpm | al_s | pH 5.0, 50 °C, 24 h, 200 rpm | al_sB | |
| Sonication | 4 × 5 min/2 min, pulser 80%, power 80% | 4 × 5 min/2 min, pulser 80%, power 80% | |||
3.3.4. Sonication
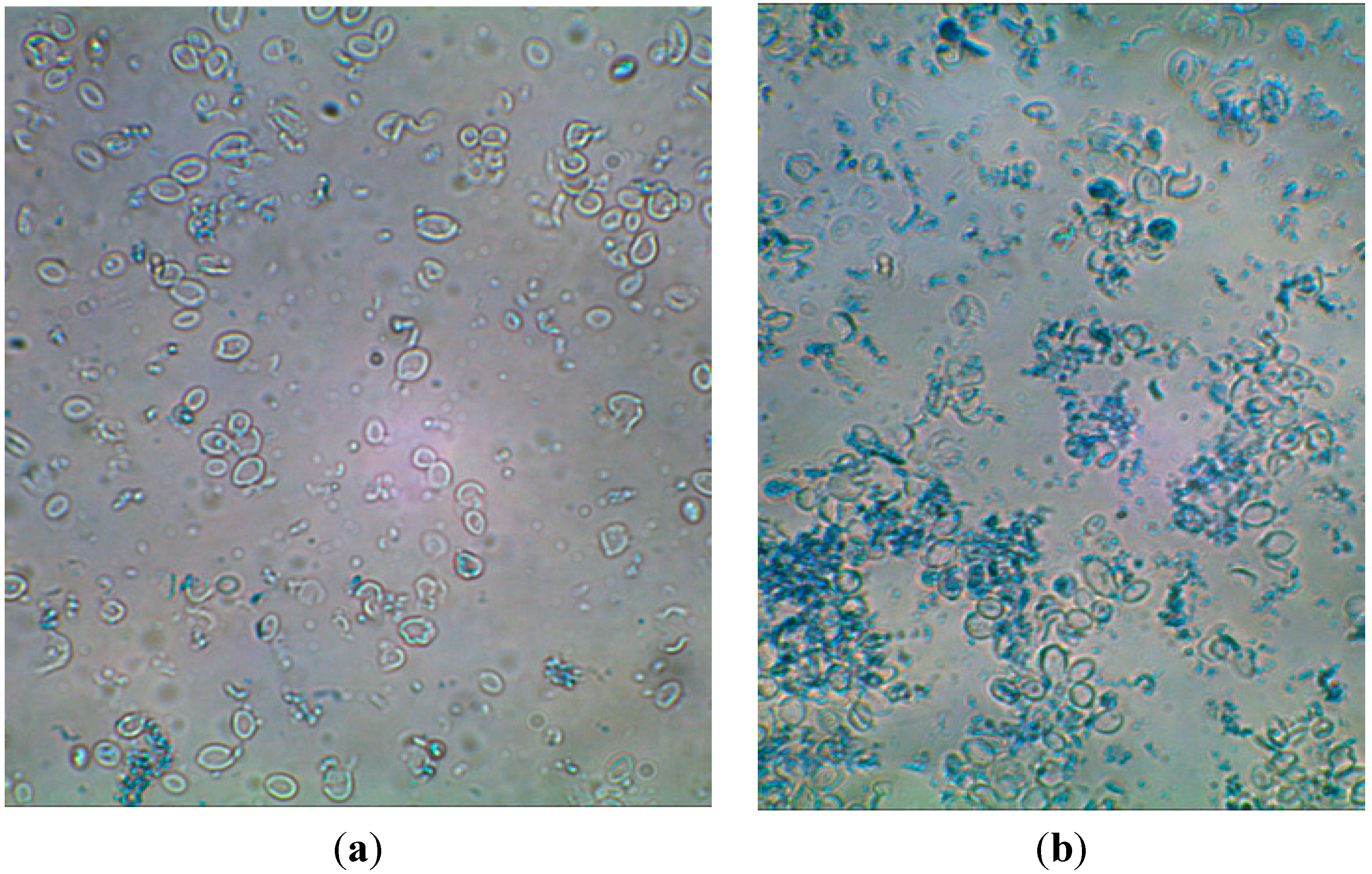
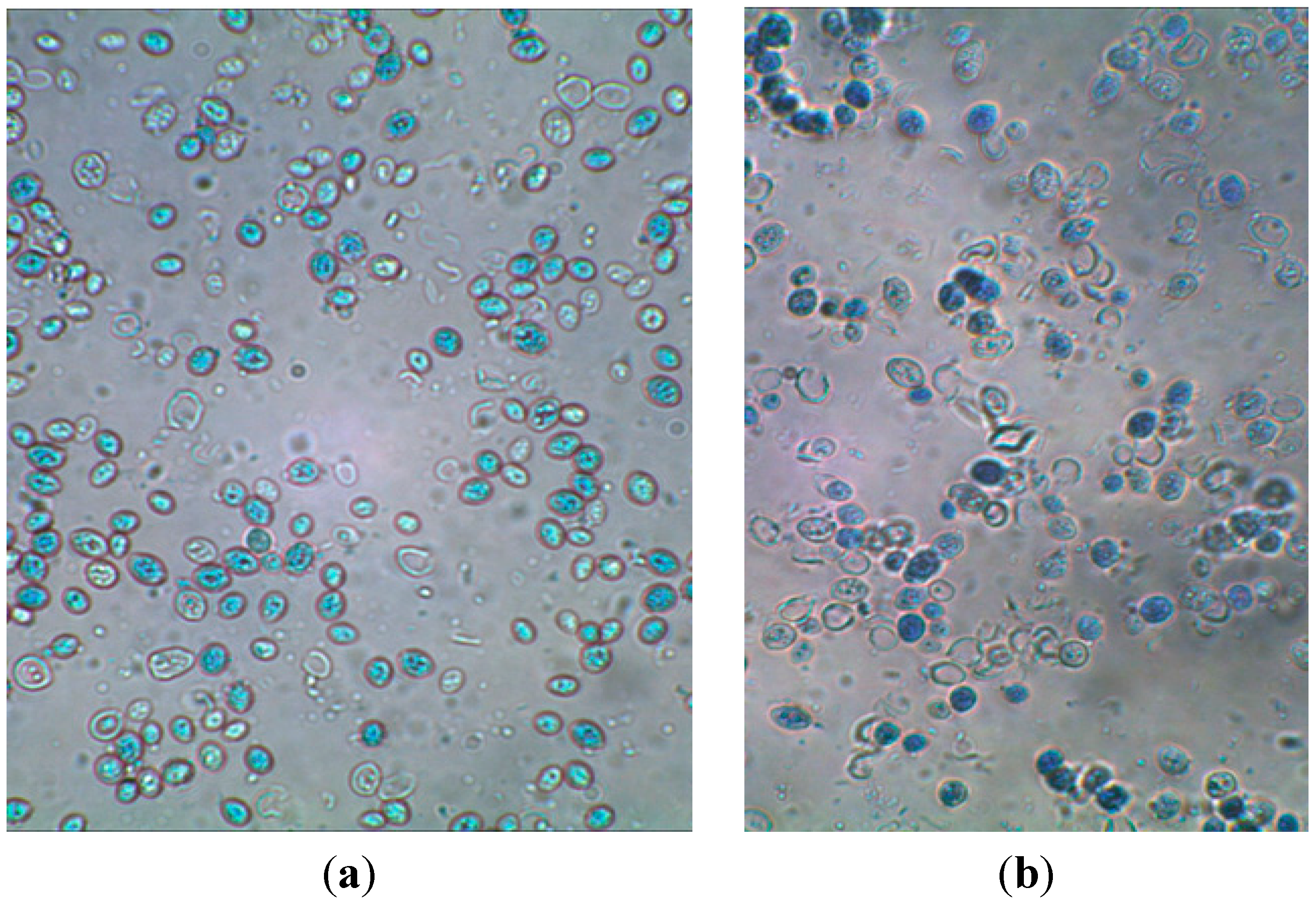
3.4. Microscope Photos
3.5. Cell Wall Preparations
3.6. Determination of the Percentage of Solubilised Material
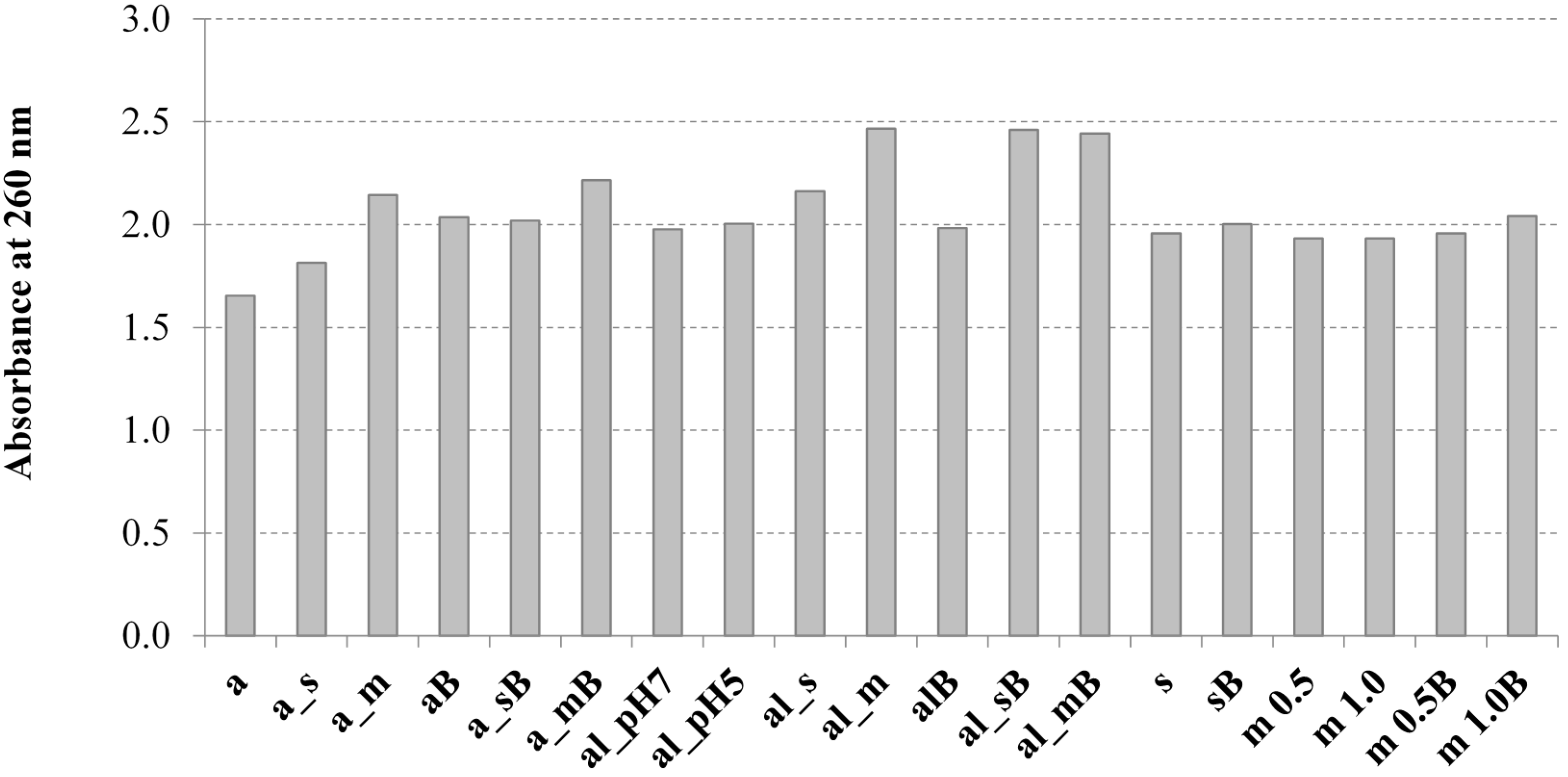
3.7. Determination of the UV Absorption of Yeast Cell Solubilised Material
3.8. Characteristics of Produced Cell Walls Preparations
3.8.1. Total Saccharides Content
3.8.2. β(1,3)(1,6)-Glucans Content
3.8.3. Total Nitrogen Content
3.8.4. FTIR Spectral Analysis
3.9. Statistical Analysis of Results
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Brown, G.D.; Gordon, S. Fungal β-glucans and mammalian immunity. Immunity 2003, 19, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Seviour, R. Medicinal importance of fungal β-(1→3),(1→6)-glucans. Mycol. Res. 2007, 111, 635–652. [Google Scholar] [CrossRef] [PubMed]
- Jouany, J.P.; Yiannikouris, A.; Bertin, G. The chemical bonds between mycotoxins and cell wall components of Saccharomyces cerevisiae have been identified. Arch. Zootech. 2005, 8, 26–50. [Google Scholar]
- Kogan, G.; Kocher, A. Role of yeast cell wall polysaccharides in pig nutrition and health protection. Livest. Sci. 2007, 109, 161–165. [Google Scholar] [CrossRef]
- Latgé, J.P. Tasting the fungal cell wall. Cell. Microbiol. 2010, 12, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Soltanian, S.; Thai, T.Q.; Dhont, J.; Sorgeloos, P.; Bossier, P. The protective effect against Vibrio campbellii in Artemia nauplii by pure β-glucan and isogenic yeast cells differing in β-glucan and chitin content operated with a source-dependent time lag. Fish Shellfish Immunol. 2007, 23, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Thanardkit, P.; Khunrae, P.; Suphantharika, M.; Verduyn, C. Glucan from spent brewer’s yeast: Preparation, analysis and use as a potential immunostimulant in shrimp feed. World J. Microbiol. Biotechnol. 2002, 18, 527–539. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vetvickova, J. β(1–3)-d-glucan affects adipogenesis, wound healing and inflammation. Orient. Pharm. Exp. Med. 2011, 11, 169–175. [Google Scholar] [CrossRef]
- Yoshida, Y.; Yokoi, W.; Ohishi, K.; Ito, M.; Naito, E.; Sawada, H. Effects of the cell wall of Kluyveromyces marxianus YIT 8292 on the plasma cholesterol and fecal sterol excretion in rats fed on a high-cholesterol diet. Biosci. Biotechnol. Biochem. 2005, 69, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Valasgues, G.L.J.; de Lima, F.O.; Boffo, E.F.; Santos, J.D.G.; da Silva, B.C.; de Assis, S. Extraction optimization and antinociceptive activity of (1–3)-β-glucan from Rhodororula mucilaginosa. Carbohydr. Polym. 2014, 105, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Yun, H.S. Production of soluble β-glucan from the cell wall of Saccharomyces cerevisiae. Enzym. Microb. Technol. 2006, 39, 496–500. [Google Scholar] [CrossRef]
- Liu, D.; Zeng, X.A.; Sun, D.W.; Han, Z. Disruption and proteins release by ultrasonication of yeast cells. Innov. Food Sci. Emerg. 2013, 18, 132–137. [Google Scholar] [CrossRef]
- Middelberg, A.J. Process-scale disruption of microorganisms. Biotechnol. Adv. 1995, 13, 491–551. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Fleet, G. Degradation of DNA during the autolysis of Saccharomyces cerevisiae. J. Ind. Microbiol. Biotechnol. 2003, 30, 175–182. [Google Scholar]
- Geciova, J.; Bury, D.; Jelen, P. Methods for disruption of microbial cells for potential use in the dairy industry—A review. Int. Dairy J. 2002, 12, 541–553. [Google Scholar] [CrossRef]
- Hernawan, T.; Fleet, G. Chemical and cytological changes during the autolysis of yeast. J. Ind. Microbiol. 1995, 14, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Wang, Q.; Cui, S.W.; Liu, H.Z. A new isolation method of β-d-glucans from spent yeast Saccharomyces cerevisiae. Food Hydrocoll. 2008, 22, 239–247. [Google Scholar] [CrossRef]
- Ŕeháćek, J.; Beran, K.; Bićik, V. Disintegration of microorganisms and preparation of yeast cell walls in a new type of disintegrator. Appl. Microbiol. 1969, 17, 462–466. [Google Scholar] [PubMed]
- Dallies, N.; Francoi, J.; Paquet, V. A new method for quantitative determination of polysaccharides in the yeast cell wall. Application to the cell wall defective mutants of Saccharomyces cerevisiae. Yeast 1998, 14, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Freimund, S.; Sauter, M.; Käppeli, O.; Dutler, H. A new non-degrading isolation process for 1, 3-β-d-glucan of high purity from baker's yeast Saccharomyces cerevisiae. Carbohydr. Polym. 2003, 54, 159–171. [Google Scholar] [CrossRef]
- Hunter, K.W.; Gult, R.A.; Berner, M.D. Preparation of microparticulate β-glucan from Saccharomyces cerevisiae for use in immune potentiation. Lett. Appl. Microbiol. 2002, 35, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Javmen, A.; Grigiškis, S.; Gliebutè, R. β-Glucan extraction from Saccharomyces cerevisiae yeast using Actinomycetes rutgersensis 88 yeast lysis enzymatic complex. Biologija 2012, 58, 51–59. [Google Scholar] [CrossRef]
- Kath, F.; Kulicke, W.M. Mild enzymatic isolation of mannan and glucan from yeast Saccharomyces cervisiae. Angew. Macromol. Chem. 1999, 268, 59–68. [Google Scholar] [CrossRef]
- Magnani, M.; Calliari, C.M.; de Macedo, J.F.C.; Mori, M.P.; de Syllos Cólus, I.M.; Casto-Gomez, R.J.H. Optimized methodology for extraction of (1→3)(1→6)-β-d-glucan from Saccharomyces cerevisiae and in vitro evaluation of the cytotoxicity and genotoxicity of the corresponding carboxymethyl derivative. Carbohydr. Polym. 2009, 78, 658–665. [Google Scholar] [CrossRef]
- Magnelli, P.; Cipollo, J.F.; Abeijon, C. A refined method for the determination of Saccharomyces cerevisiae cell wall composition and β-1,6-glucan fine structure. Anal. Biochem. 2002, 301, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Marinescu, G.; Stoicescu, A. Researches concerning the preparation of spent brewer’s yeast β-glucans. J. Agroaliment. Proc. Technol. 2009, 15, 547–553. [Google Scholar]
- Nguyen, T.H.; Fleet, G.H.; Rogers, P.L. Composition of the cell walls of several yeast species. Appl. Microbiol. Biotechnol. 1998, 50, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Suphantharika, M.; Khunrae, P.; Thanardkit, P.; Verduyn, C. Preparation of spent brewer’s yeast β-glucans with a potential application as an immunostimulant for black tiger shrimp Penaeus monodon. Bioresour. Technol. 2003, 88, 55–56. [Google Scholar] [CrossRef] [PubMed]
- Zehner-Krpan, V.; Petravić-Tominac, V.; Galović, P.; Galović, V.; Filipović-Grčić, J.; Srečec, S. Application of different drying methods on β-glucan isolated from spent brewer’s yeast using alkaline procedure. Agric. Conspec. Sci. 2010, 75, 45–50. [Google Scholar]
- Klimek-Ochab, M.; Brzezinska-Rodak, M.; Żymanczyk-Duda, E.; Lejczak, B.; Kafarski, P. Comparative study of fungal cell disruption-scope and limitation of the methods. Folia Microbiol. 2011, 56, 469–475. [Google Scholar] [CrossRef]
- Zhao, J.; Fleet, G. Degradation of RNA during the autolysis of Saccharomyces cerevisiae. Produces predominantly ribonucleotides. J. Ind. Microbiol. Biotechnol. 2005, 32, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Young, S.H.; Dong, W.J.; Jacobs, R.R. Observation of partially opened triple-helix conformation in (1→3)-β-glucan by fluorescence resonance energy transfer spectroscopy. J. Biol. Chem. 2000, 275, 111874–111879. [Google Scholar]
- Novák, M.; Synytsya, A.; Gedeon, O.; Slepička, P.; Procházka, V.; Synytsya, A.; Blahovec, J.; Hejlová, A.; Čopíková, J. Yeast β(1,3),(1,6)-d-glucan films: Preparation and characterization of some structural and physical properties. Carbohydr. Polym. 2012, 87, 2496–2504. [Google Scholar] [CrossRef]
- Hromádková, Z.; Ebringerová, A.; Sasinková, V.; Šandula, J.; Hříbalová, V.; Omelková, J. Influence of the drying method on the physical properties and immunomodulatory activity of the particulate (1→3)-β-d-glucan from Saccharomyces cerevisiae. Carbohydr. Polym. 2003, 51, 9–15. [Google Scholar] [CrossRef]
- Taboury, J.A.; Liquier, J.; Taillandier, E. Characterization of DNA structures by infrared spectroscopy: double heli cal forms of poly(dG-dC) poly(dG-dC) poly(dD8G-dC), and poly(dG-dm5C)·poly(dG-dm5C). Can. J. Chem. 1985, 63, 1904–1909. [Google Scholar] [CrossRef]
- Banyay, M.; Sarkar, M.; Gräslund, A. A libr ary of IR band of nucleic acids in solution. Biophys. Chem. 2003, 104, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Galichet, A.; Sockalingum, G.D.; Belarbi, A.; Manfait, M. FTIR spectroscopic analysis of Saccharomyces cerevisiae cell walls: Study of an anomalous exhibiting a pink-colored cell phenotype. FEMS Microbiol. Lett. 2001, 197, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Križková, L.; Ďuračková, Z.; Šandula, J.; Sasinková, V.; Krajčovič, J. Antioxidative and antimutagenic activity of yeast cell wall mannans in vitro. Mut. Res. 2001, 497, 213–222. [Google Scholar] [CrossRef]
- Adt, I.; Toubas, D.; Pinon, J.-M.; Manfait, M.; Sockalingum, G.D. FTIR spectroscopy as a potential tool to analyse structural modifications during morphogenesis of Candida albicans. Arch. Microbiol. 2006, 185, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Tajmir-Rhiai, H.A.; N’soukpoé-Kossi, C.N.; Joly, D. Structural analysis of protein-DNA and protein-RNA interactions by FTIR, UV-visible and CD spectroscopic methods. Spectroscopy 2009, 23, 81–101. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, S.; Wu, T. Combination of induced autolysis and sodium hypochlorite oxidation for the production of Saccharomyces cerevisiae (1→3)-β-d-glucan. World J. Microbiol. Biotechnol. 2003, 19, 947–952. [Google Scholar] [CrossRef]
- Thammakiti, S.; Suphantharika, M.; Phaesuwan, T.; Verduyn, C. Preparation of spent brewer’s yeast β-glucans for potential application in the food industry. Int. J. Food Sci. Technol. 2004, 39, 21–29. [Google Scholar] [CrossRef]
- Danielson, M.E.; Dauth, R.; Elmasry, N.A.; Langesly, R.R.; Magee, A.S.; Will, P.M. Enzymatic method to measure β-1,3-β-1,6-glucan content in extracts and formulated products (GEM Assay). J. Agric. Food Chem. 2010, 58, 10305–10308. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bzducha-Wróbel, A.; Błażejak, S.; Kawarska, A.; Stasiak-Różańska, L.; Gientka, I.; Majewska, E. Evaluation of the Efficiency of Different Disruption Methods on Yeast Cell Wall Preparation for β-Glucan Isolation. Molecules 2014, 19, 20941-20961. https://doi.org/10.3390/molecules191220941
Bzducha-Wróbel A, Błażejak S, Kawarska A, Stasiak-Różańska L, Gientka I, Majewska E. Evaluation of the Efficiency of Different Disruption Methods on Yeast Cell Wall Preparation for β-Glucan Isolation. Molecules. 2014; 19(12):20941-20961. https://doi.org/10.3390/molecules191220941
Chicago/Turabian StyleBzducha-Wróbel, Anna, Stanisław Błażejak, Anna Kawarska, Lidia Stasiak-Różańska, Iwona Gientka, and Ewa Majewska. 2014. "Evaluation of the Efficiency of Different Disruption Methods on Yeast Cell Wall Preparation for β-Glucan Isolation" Molecules 19, no. 12: 20941-20961. https://doi.org/10.3390/molecules191220941
APA StyleBzducha-Wróbel, A., Błażejak, S., Kawarska, A., Stasiak-Różańska, L., Gientka, I., & Majewska, E. (2014). Evaluation of the Efficiency of Different Disruption Methods on Yeast Cell Wall Preparation for β-Glucan Isolation. Molecules, 19(12), 20941-20961. https://doi.org/10.3390/molecules191220941