The “Funny” Current (If) Inhibition by Ivabradine at Membrane Potentials Encompassing Spontaneous Depolarization in Pacemaker Cells

Abstract
:1. Introduction
2. Results
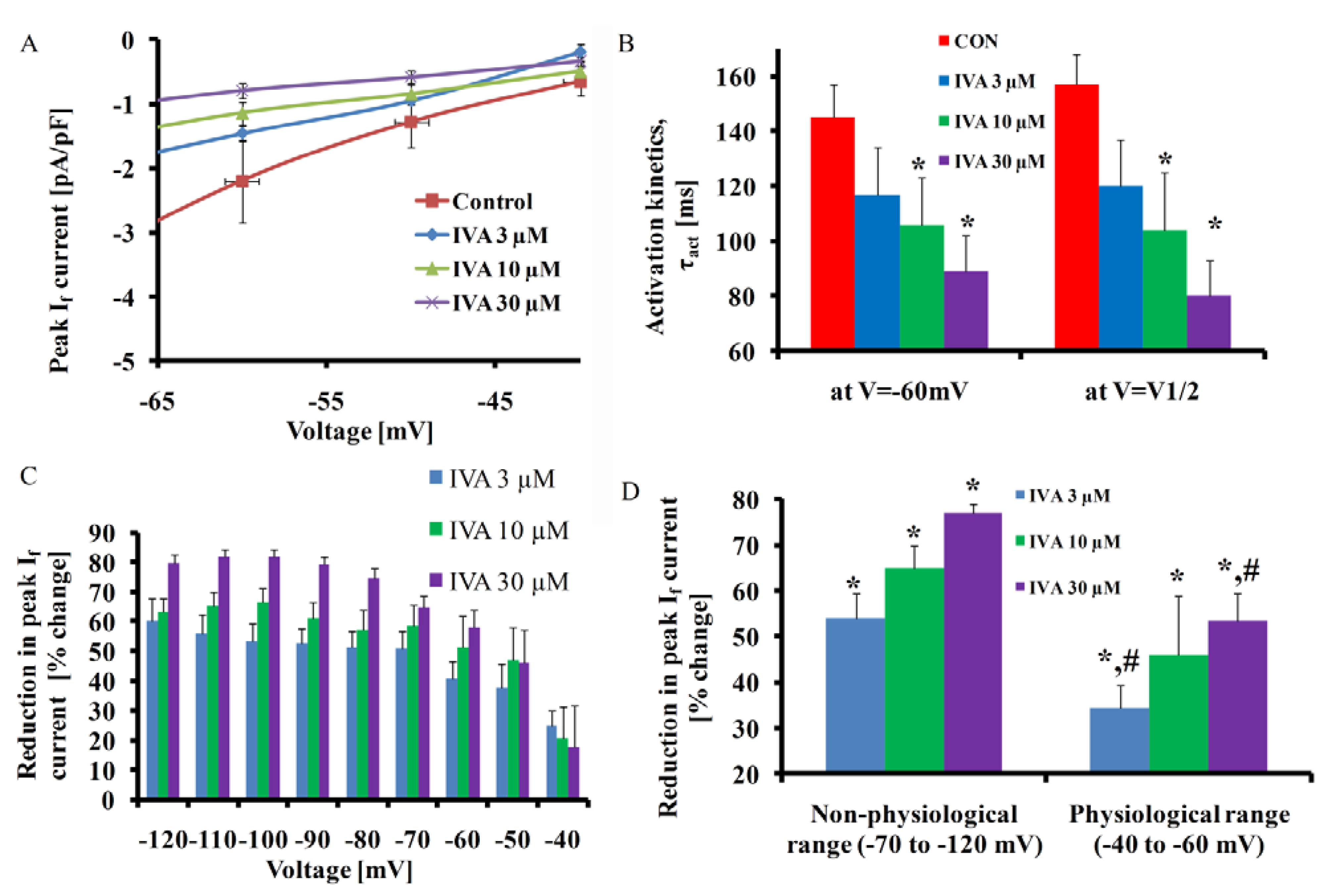
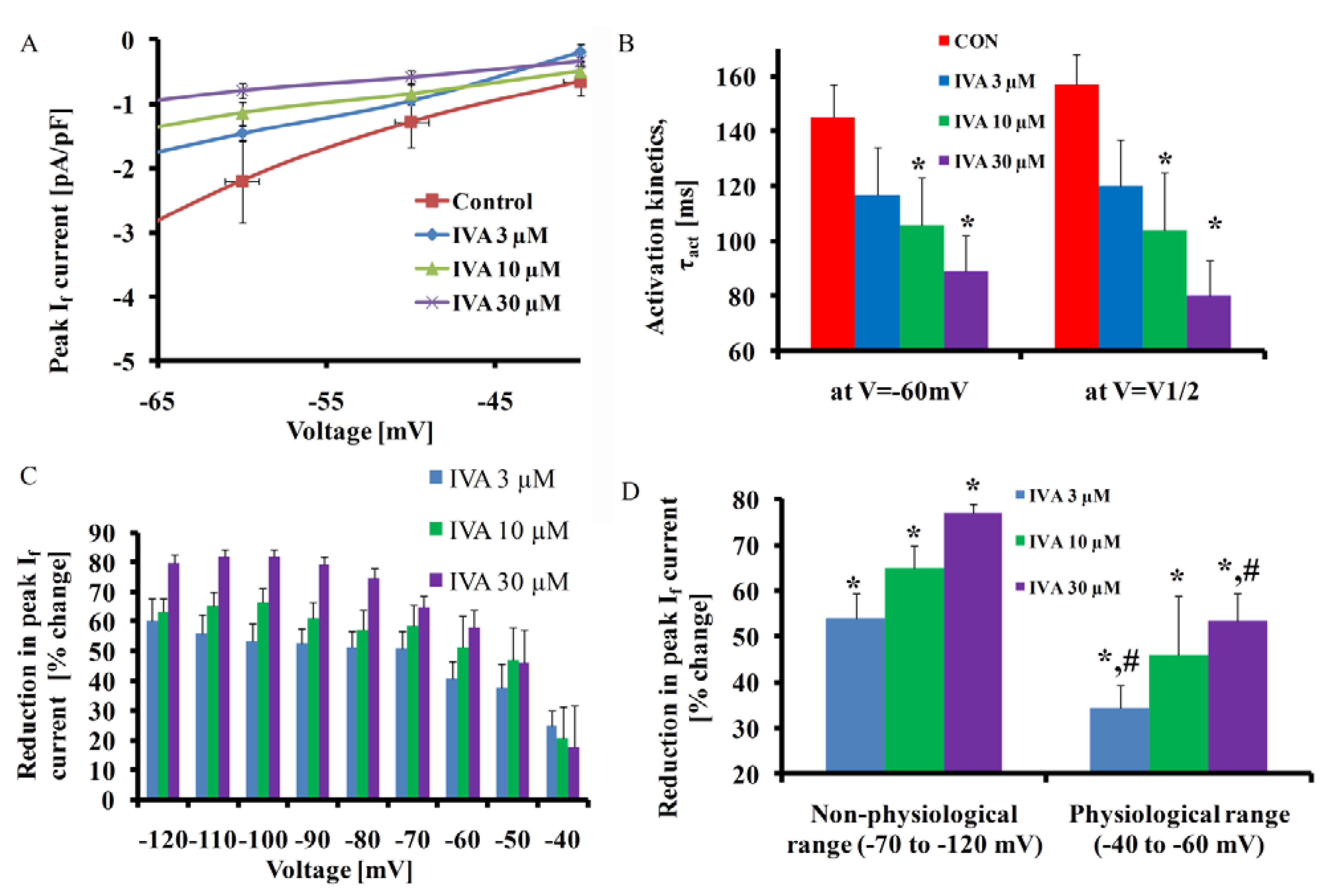
2.1. IVA Blocks If in SANC
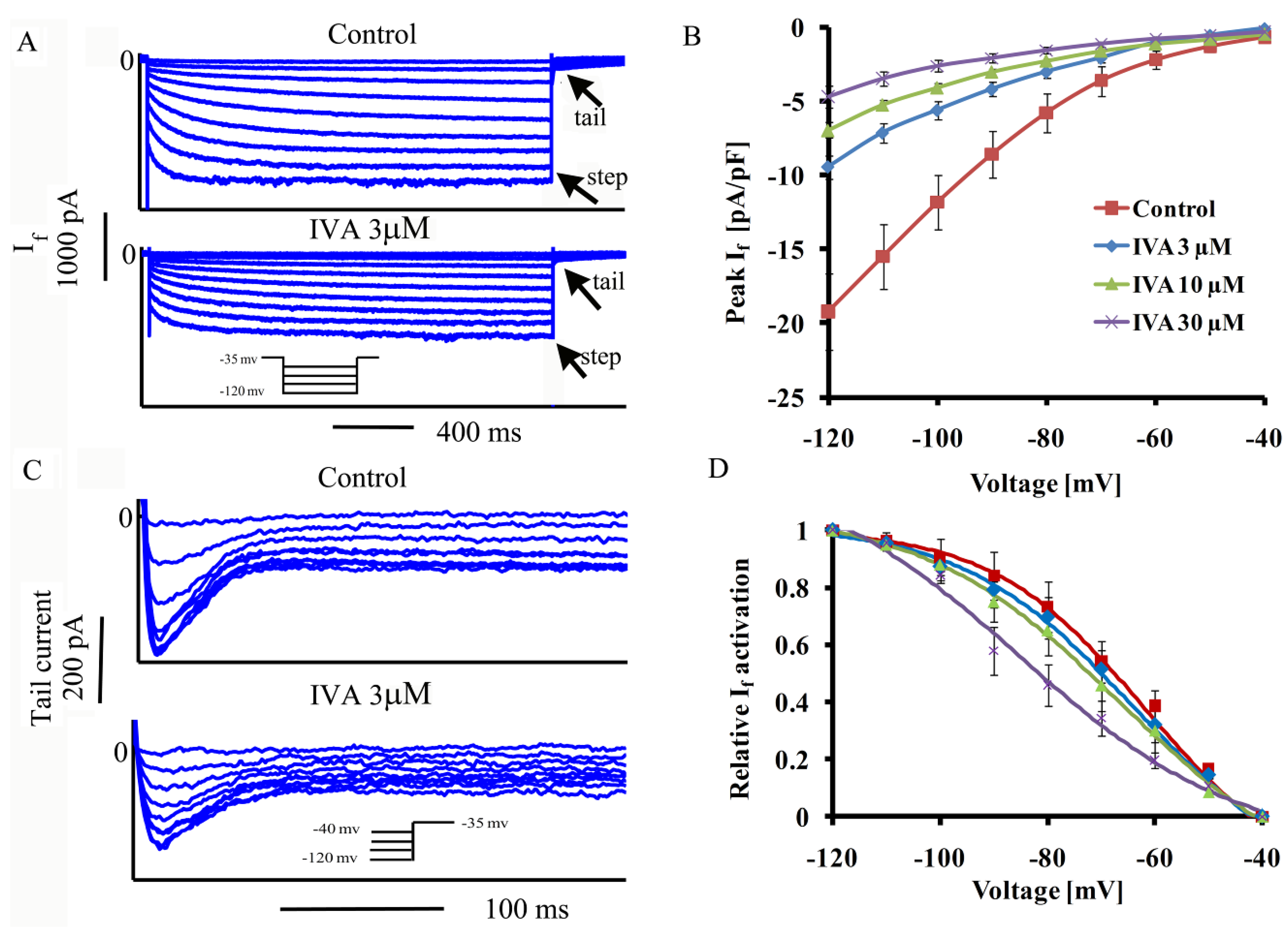

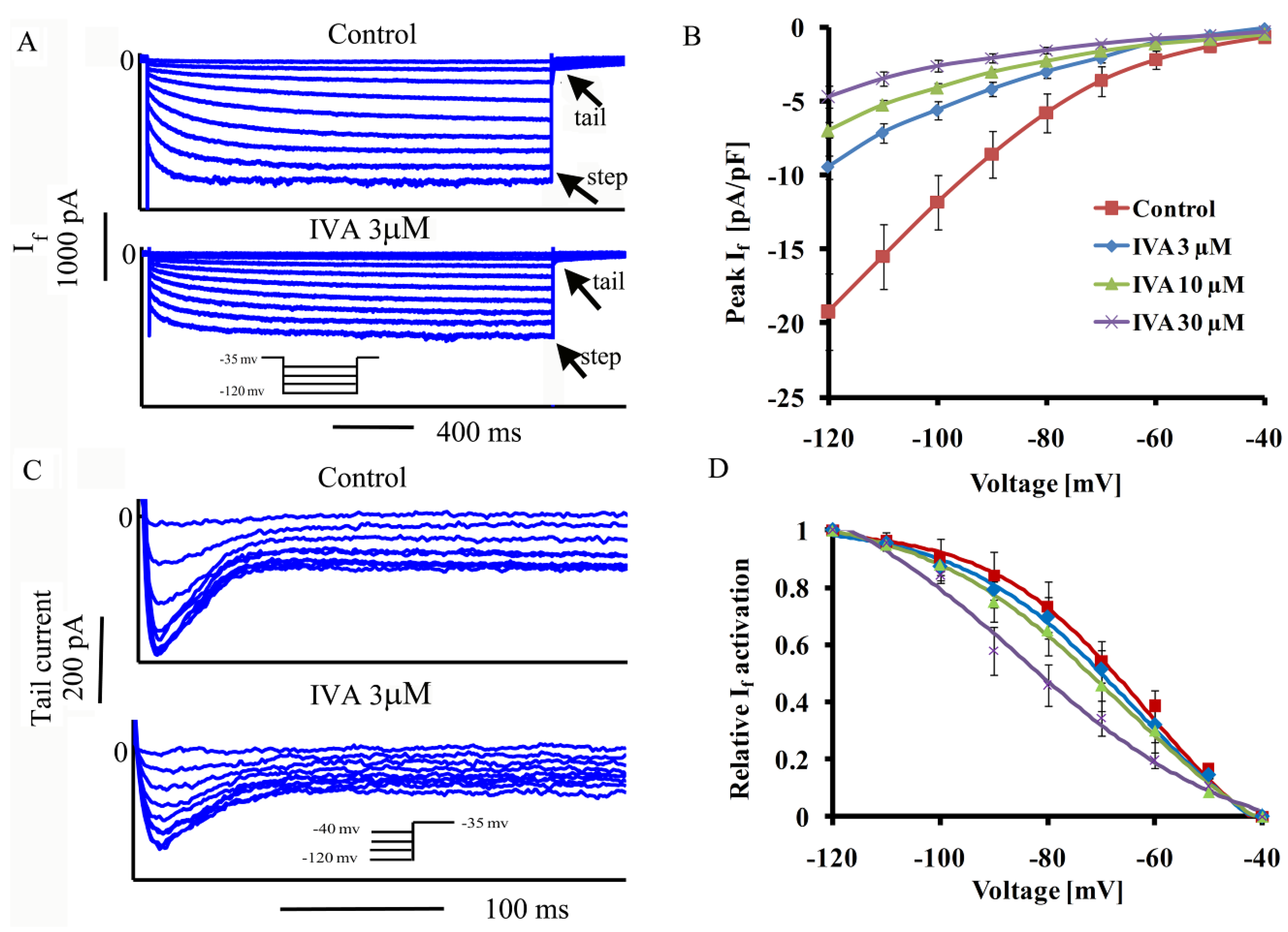{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Ivabradine | Ivabradine | Ivabradine | |
|---|---|---|---|---|
| 3 μM | 10 μM | 30 μM | ||
| (n = 9) | (n = 9) | (n = 9) | ||
| V1/2 activation (mV) | −68 ± 4 | −70 ± 4 | −74 ± 4 * | −77 ± 5 * |
| (p = 0.07) | (p = 0.02) | (p = 0.04) | ||
| Max τact | 157 ± 11 | 120 ± 17 | 106 ± 17 * | 80 ± 13 * |
| (ms) | (P = 0.5) | (P = 0.05) | (P = 0.02) | |
| If density (pA/pF) | 4.5 ± 0.7 | 2.1 ± 0.3 * | 1.6 ± 0.2 * | 1.1 ± 0.2 *,# |
| (V = control V1/2 activation) | (P = 0.04) | (P = 0.008) | (0.004) | |
| If (pA/pF) (V = MDP) | 3 ± 0.43 | 2 ± 0.3 * | 1.5 ± 0.2 * | 1.3 ± 0.2 * |
| (P = 0.045) | (P = 0.04) | (P = 0.01) |
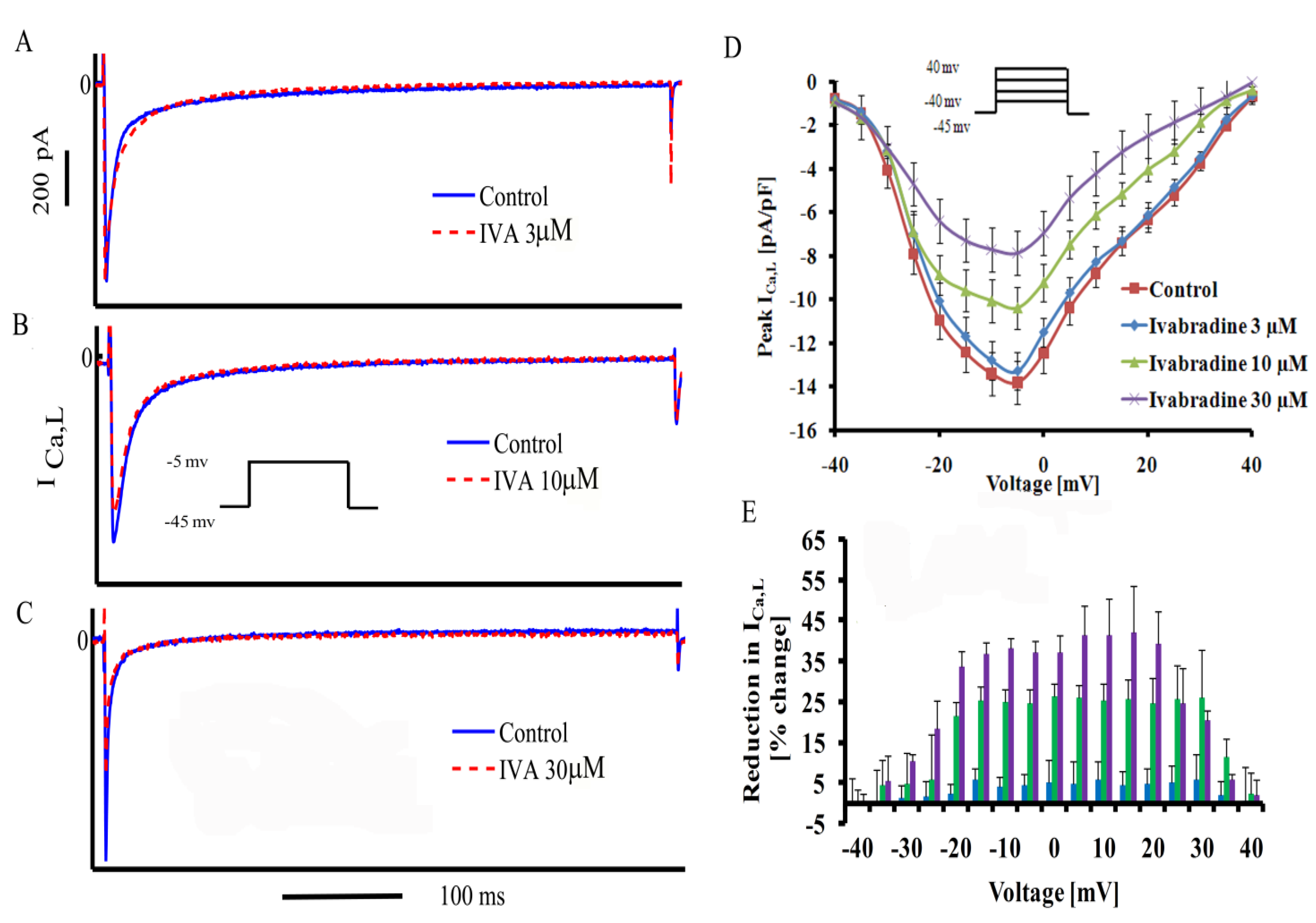
2.2. Concentration-Dependent Block of ICa,L by IVA in SANC

| Control | Ivabradine | Ivabradine | Ivabradine | |
|---|---|---|---|---|
| 3 μM | 10 μM | 30 μM | ||
| (n = 12) | (n = 9) | (n = 9) | ||
| Peak ICa,L density (pA/pF) | 13.9 ± 1 | 13.2 ± 1 | 9.85 ± 1.4 * | 8.1 ± 1 *,# |
| (p = 0.953) | (p = 0.0003) | (p < 0.001) | ||
| Voltage at peak ICa,L (mV) | −5.55 ± 1 | −5.4 ± 0.7 | −5 ± 0.1 | −6 ± 0.8 |
| (p = 0.84) | (p = 1) | (p = 0.98) |
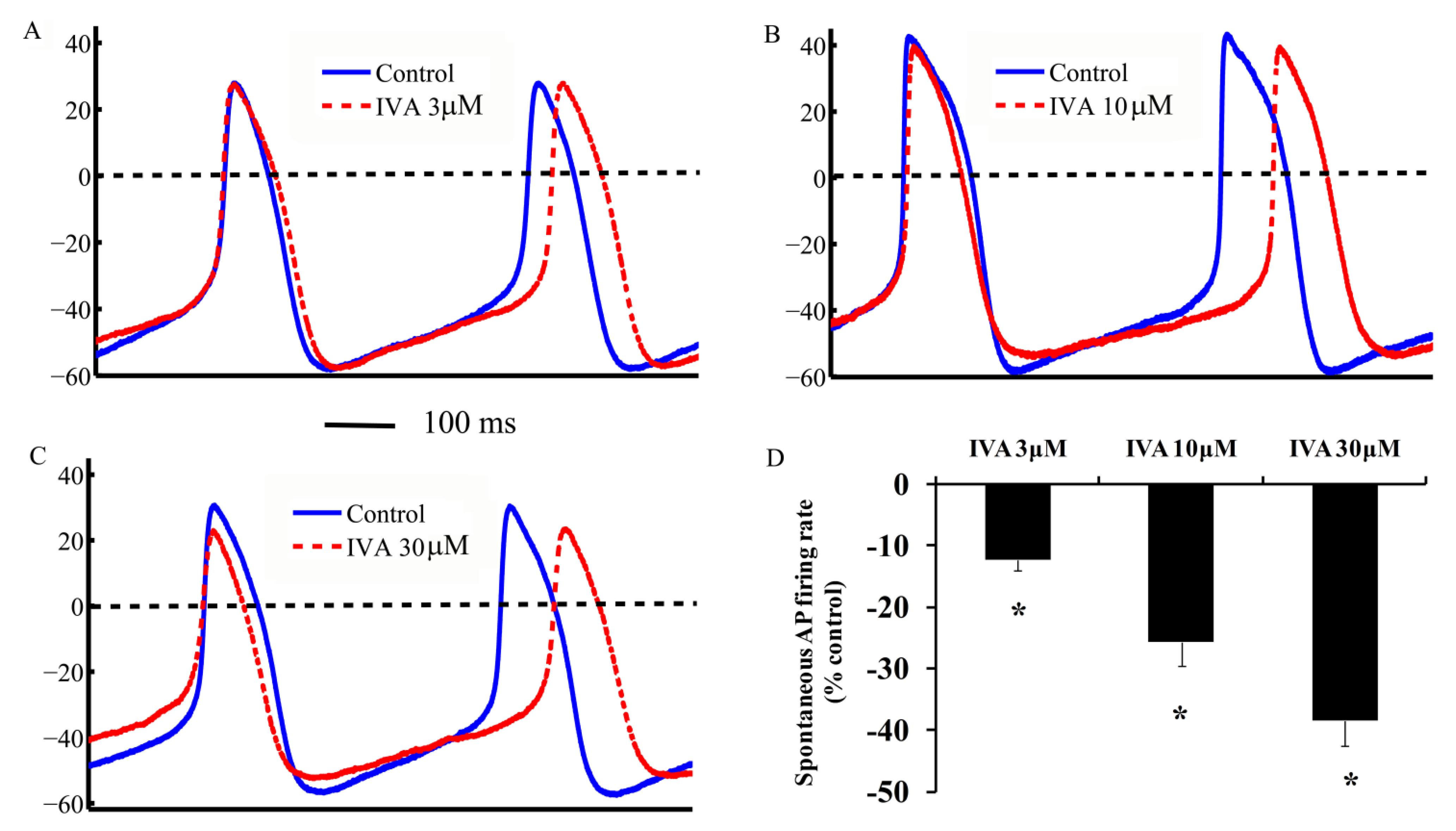
2.3. IVA Reduces the SANC Spontaneous AP Firing Rate
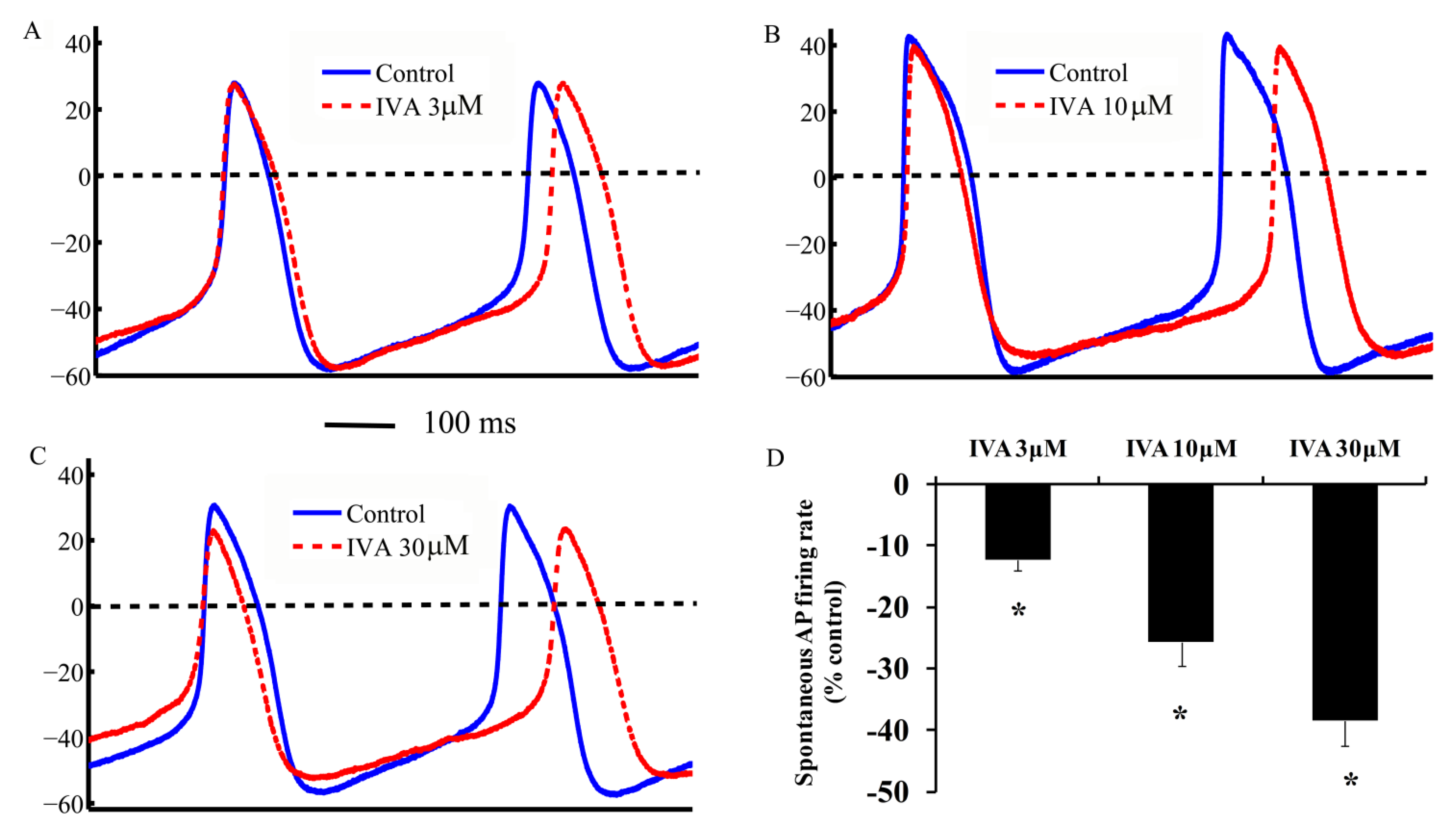
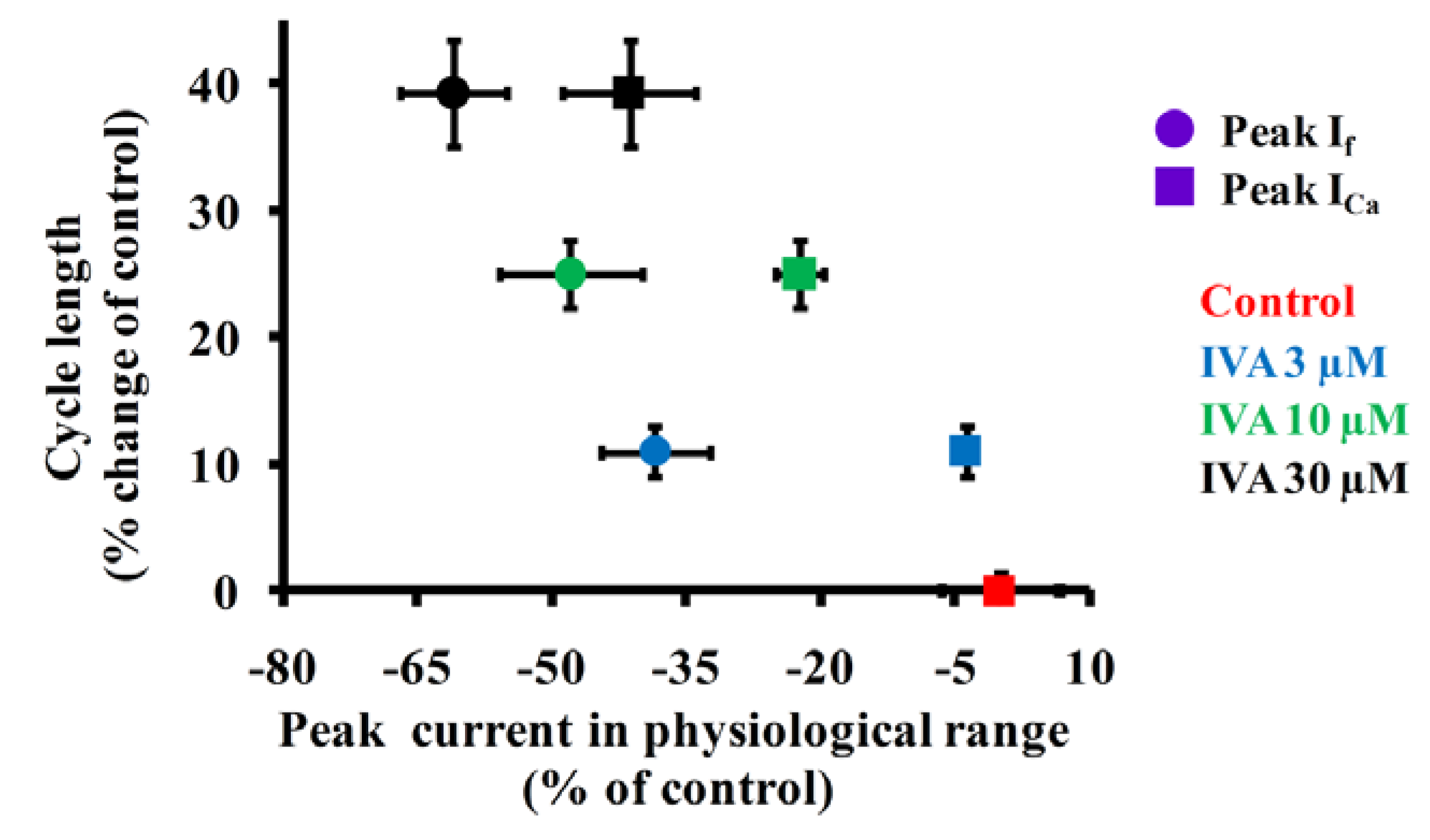
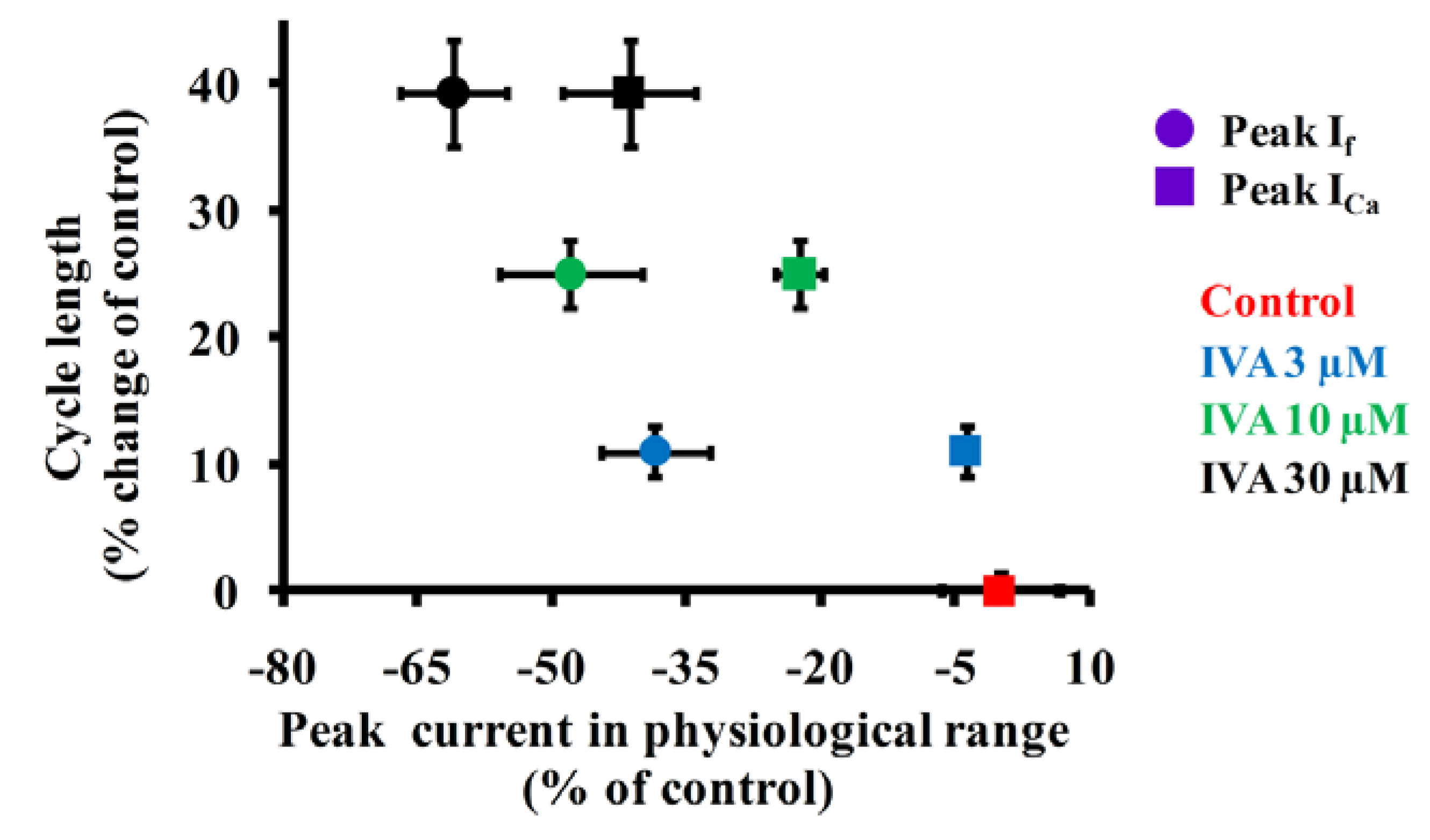
2.4. Correlations Among If, ICa,L and Spontaneous AP Cycle Length in the Absence and Presence of IVA
3. Discussion
| Species | MDP [mV] | Amplitude at MDP [pA/pF] | Maximal Current at −120 mV [pA/pF] | Reference |
|---|---|---|---|---|
| Rabbit | −60 | 8 | 16 | [7] |
| 10 | 30 | [8] | ||
| 1 | 10 | [9] | ||
| 1 | 5 | [10] | ||
| 1 | 6 | [11] | ||
| 1 | 18 | [12] | ||
| 1 | 22 | [13] | ||
| 4 | 20 | [14] | ||
| 4 | 16 | [15] | ||
| 3 | 19 | [16] | ||
| 1 | 6 | [17] | ||
| Mouse | −62 | 2 | 11 | [18] |
| 2 | 22 | [19] | ||
| 2 | 15 | [20] | ||
| 1 | 20 | [21] | ||
| 2 | 15 | [22] | ||
| Dog | −58 | 1 | 6 | [18] |
| 2 | 10 | [23] | ||
| 2 | 11 | [24] | ||
| Guinea pig | −61 | 1 | 12 | [25] |
| Rat | −58 | 5 | 15 | [26] |
| Cat | −68 | 3 | 6 | [27] |
| Human | −62 | 1 | 8 | [28] |
4. Experimental
4.1. Cell Preparation
4.2. Electrophysiology and Cell Contraction
4.3. If Measurements
4.4. ICa,L Measurements
4.5. Drugs
4.6. Statistical Analysis
5. Conclusions
Acknowledgment
Conflicts of Interest
References
- Mensink, G.B.; Hoffmeister, H. The relationship between resting heart rate and all-cause, cardiovascular and cancer mortality. Eur. Heart J. 1997, 18, 1404–1410. [Google Scholar] [CrossRef]
- Palatini, P. Heart rate as an independent risk factor for cardiovascular disease: Current evidence and basic mechanisms. Drugs 2007, 67 Suppl. 2, S3–S13. [Google Scholar] [CrossRef]
- Riccioni, G. Ivabradine: Recent and potential applications in clinical practice. Expert Opin. Pharmacother. 2011, 12, 443–450. [Google Scholar] [CrossRef]
- Bois, P.; Bescond, J.; Renaudon, B.; Lenfant, J. Mode of action of bradycardic agent, S 16257, on ionic currents of rabbit sinoatrial node cells. Br. J. Pharmacol. 1996, 118, 1051–1057. [Google Scholar] [CrossRef]
- Boldt, A.; Gergs, U.; Ponicke, K.; Simm, A.; Silber, R.E.; Neumann, J. Inotropic effects of ivabradine in the mammalian heart. Pharmacology 2010, 86, 249–258. [Google Scholar] [CrossRef]
- Bucchi, A.; Baruscotti, M.; DiFrancesco, D. Current-dependent block of rabbit sino-atrial node If channels by ivabradine. J. Gen. Physiol. 2002, 120, 1–13. [Google Scholar] [CrossRef]
- Accili, E.A.; Robinson, R.B.; DiFrancesco, D. Properties and modulation of If in newborn versus adult cardiac SA node. Am. J. Physiol. 1997, 272, H1549–H1552. [Google Scholar]
- Goethals, M.; Raes, A.; van Bogaert, P.P. Use-dependent block of the pacemaker current If in rabbit sinoatrial node cells by zatebradine (UL-FS 49). On the mode of action of sinus node inhibitors. Circulation 1993, 88, 2389–2401. [Google Scholar] [CrossRef]
- Honjo, H.; Boyett, M.R.; Kodama, I.; Toyama, J. Correlation between electrical activity and the size of rabbit sino-atrial node cells. J. Physiol. 1996, 496, 795–808. [Google Scholar]
- Lei, M.; Cooper, P.J.; Camelliti, P.; Kohl, P. Role of the 293b-sensitive, slowly activating delayed rectifier potassium current, iKs, in pacemaker activity of rabbit isolated sino-atrial node cells. Cardiovasc. Res. 2002, 53, 68–79. [Google Scholar] [CrossRef]
- Li, J.; Qu, J.; Nathan, R.D. Ionic basis of ryanodine’s negative chronotropic effect on pacemaker cells isolated from the sinoatrial node. Am. J. Physiol. 1997, 273, H2481–H2489. [Google Scholar]
- Lyashkov, A.E.; Vinogradova, T.M.; Zahanich, I.; Li, Y.; Younes, A.; Nuss, H.B.; Spurgeon, H.A.; Maltsev, V.A.; Lakatta, E.G. Cholinergic receptor signaling modulates spontaneous firing of sinoatrial nodal cells via integrated effects on PKA-dependent Ca2+ cycling and IKACh. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H949–H959. [Google Scholar] [CrossRef]
- Noble, D.; Denyer, J.C.; Brown, H.F.; DiFrancesco, D. Reciprocal role of the inward currents ib, Na and if in controlling and stabilizing pacemaker frequency of rabbit sino-atrial node cells. Proc. Biol. Sci. 1992, 250, 199–207. [Google Scholar] [CrossRef]
- Satoh, H. [Ca2+]i-dependent actions of taurine in spontaneously beating rabbit sino-atrial nodal cells. Eur. J. Pharmacol. 2001, 424, 19–25. [Google Scholar] [CrossRef]
- Verheijck, E.E.; van Ginneken, A.C.; Wilders, R.; Bouman, L.N. Contribution of L-type Ca2+ current to electrical activity in sinoatrial nodal myocytes of rabbits. Am. J. Physiol. 1999, 276, H1064–H1077. [Google Scholar]
- Verkerk, A.O.; den Ruijter, H.M.; Bourier, J.; Boukens, B.J.; Brouwer, I.A.; Wilders, R.; Coronel, R. Dietary fish oil reduces pacemaker current and heart rate in rabbit. Heart Rhythm. 2009, 6, 1485–1492. [Google Scholar] [CrossRef]
- Verkerk, A.O.; Wilders, R.; Zegers, J.G.; van Borren, M.M.; Ravesloot, J.H.; Verheijck, E.E. Ca2+-activated Cl− current in rabbit sinoatrial node cells. J. Physiol. 2002, 540, 105–117. [Google Scholar] [CrossRef]
- Gao, Z.; Chen, B.; Joiner, M.L.; Wu, Y.; Guan, X.; Koval, O.M.; Chaudhary, A.K.; Cunha, S.R.; Mohler, P.J.; Martins, J.B.; et al. If and SR Ca2+ release both contribute to pacemaker activity in canine sinoatrial node cells. J. Mol. Cell. Cardiol. 2010, 49, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Mangoni, M.E.; Nargeot, J. Properties of the hyperpolarization-activated current (If) in isolated mouse sino-atrial cells. Cardiovasc. Res. 2001, 52, 51–64. [Google Scholar] [CrossRef]
- Rose, R.A.; Kabir, M.G.; Backx, P.H. Altered heart rate and sinoatrial node function in mice lacking the cAMP regulator phosphoinositide 3-kinase-gamma. Circ. Res. 2007, 101, 1274–1282. [Google Scholar] [CrossRef]
- Rose, R.A.; Lomax, A.E.; Kondo, C.S.; Anand-Srivastava, M.B.; Giles, W.R. Effects of C-type natriuretic peptide on ionic currents in mouse sinoatrial node: A role for the NPR-C receptor. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H1970–H1977. [Google Scholar] [CrossRef] [Green Version]
- Springer, J.; Azer, J.; Hua, R.; Robbins, C.; Adamczyk, A.; McBoyle, S.; Bissell, M.B.; Rose, R.A. The natriuretic peptides BNP and CNP increase heart rate and electrical conduction by stimulating ionic currents in the sinoatrial node and atrial myocardium following activation of guanylyl cyclase-linked natriuretic peptide receptors. J. Mol. Cell. Cardiol. 2012, 52, 1122–1134. [Google Scholar] [CrossRef]
- Protas, L.; Oren, R.V.; Clancy, C.E.; Robinson, R.B. Age-dependent changes in Na current magnitude and TTX-sensitivity in the canine sinoatrial node. J. Mol. Cell. Cardiol. 2010, 48, 172–180. [Google Scholar] [CrossRef]
- Yeh, Y.H.; Burstein, B.; Qi, X.Y.; Sakabe, M.; Chartier, D.; Comtois, P.; Wang, Z.; Kuo, C.T.; Nattel, S. Funny current downregulation and sinus node dysfunction associated with atrial tachyarrhythmia: A molecular basis for tachycardia-bradycardia syndrome. Circulation 2009, 119, 1576–1585. [Google Scholar]
- Kojima, A.; Kitagawa, H.; Omatsu-Kanbe, M.; Matsuura, H.; Nosaka, S. Inhibitory effects of sevoflurane on pacemaking activity of sinoatrial node cells in guinea-pig heart. Br. J. Pharmacol. 2012. [Google Scholar]
- Satoh, H. Elecropharmacology of taurine on the hyperpolarization-activated inward current and the sustained inward current in spontaneously beating rat sino-atrial nodal cells. J. Pharmacol. Sci. 2003, 91, 229–238. [Google Scholar] [CrossRef]
- Zhou, Z.; Lipsius, S.L. Properties of the pacemaker current If in latent pacemaker cells isolated from cat right atrium. J. Physiol. 1992, 453, 503–523. [Google Scholar]
- Verkerk, A.O.; Wilders, R.; van Borren, M.M.; Peters, R.J.; Broekhuis, E.; Lam, K.; Coronel, R.; de Bakker, J.M.; Tan, H.L. Pacemaker current (If) in the human sinoatrial node. Eur. Heart J. 2007, 28, 2472–2478. [Google Scholar]
- Bucchi, A.; Baruscotti, M.; Robinson, R.B.; DiFrancesco, D. Modulation of rate by autonomic agonists in SAN cells involves changes in diastolic depolarization and the pacemaker current. J. Mol. Cell. Cardiol. 2007, 43, 39–48. [Google Scholar] [CrossRef]
- Borer, J.S.; Le Heuzey, J.Y. Characterization of the heart rate-lowering action of ivabradine, a selective I(f) current inhibitor. Am. J. Ther. 2008, 15, 461–473. [Google Scholar] [CrossRef]
- Camm, A.J.; Lau, C.P. Electrophysiological effects of a single intravenous administration of ivabradine (S 16257) in adult patients with normal baseline electrophysiology. Drugs R D 2003, 4, 83–89. [Google Scholar] [CrossRef]
- Simon, L.; Ghaleh, B.; Puybasset, L.; Giudicelli, J.F.; Berdeaux, A. Coronary and hemodynamic effects of S 16257, a new bradycardic agent, in resting and exercising conscious dogs. J. Pharmacol. Exp. Ther. 1995, 275, 659–666. [Google Scholar]
- Maltsev, V.A.; Lakatta, E.G. A novel quantitative explanation for the autonomic modulation of cardiac pacemaker cell automaticity via a dynamic system of sarcolemmal and intracellular proteins. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H2010–H2023. [Google Scholar] [CrossRef]
- Mangoni, M.E.; Nargeot, J. Genesis and regulation of the heart automaticity. Physiol. Rev. 2008, 88, 919–982. [Google Scholar] [CrossRef]
- Mangoni, M.E.; Couette, B.; Bourinet, E.; Platzer, J.; Reimer, D.; Striessnig, J.; Nargeot, J. Functional role of L-type Cav1.3 Ca2+ channels in cardiac pacemaker activity. Proc. Natl. Acad. Sci. USA 2003, 100, 5543–5548. [Google Scholar]
- Bogdanov, K.Y.; Vinogradova, T.M.; Lakatta, E.G. Sinoatrial nodal cell ryanodine receptor and Na+-Ca2+ exchanger: molecular partners in pacemaker regulation. Circ. Res. 2001, 88, 1254–1258. [Google Scholar] [CrossRef]
- Shinohara, T.; Park, H.W.; Joung, B.; Maruyama, M.; Chua, S.K.; Han, S.; Shen, M.J.; Chen, P.S.; Lin, S.F. Selective sinoatrial node optical mapping and the mechanism of sinus rate acceleration. Circ. J. 2012, 76, 309–316. [Google Scholar] [CrossRef]
- Bohm, M.; Swedberg, K.; Komajda, M.; Borer, J.S.; Ford, I.; Dubost-Brama, A.; Lerebours, G.; Tavazzi, L. Heart rate as a risk factor in chronic heart failure (SHIFT): The association between heart rate and outcomes in a randomised placebo-controlled trial. Lancet 2010, 376, 886–894. [Google Scholar]
- Amosova, E.; Andrejev, E.; Zaderey, I.; Rudenko, U.; Ceconi, C.; Ferrari, R. Efficacy of ivabradine in combination with Beta-blocker versus uptitration of Beta-blocker in patients with stable angina. Cardiovasc. Drugs Ther. 2011, 25, 531–537. [Google Scholar] [CrossRef]
- Vinogradova, T.M.; Sirenko, S.; Lyashkov, A.E.; Younes, A.; Li, Y.; Zhu, W.; Yang, D.; Ruknudin, A.M.; Spurgeon, H.; Lakatta, E.G. Constitutive phosphodiesterase activity restricts spontaneous beating rate of cardiac pacemaker cells by suppressing local Ca2+ releases. Circ. Res. 2008, 102, 761–769. [Google Scholar] [CrossRef]
- Yaniv, Y.; Juhaszova, M.; Lyashkov, A.E.; Spurgeon, H.; Sollott, S.J.; Lakatta, E.G. Ca2+-regulated-cAMP/PKA signaling in cardiac pacemaker cells links ATP supply to demand. J. Mol. Cell. Cardiol. 2011, 51, 740–748. [Google Scholar] [CrossRef]
- van Ginneken, A.C.; Giles, W. Voltage clamp measurements of the hyperpolarization-activated inward current If in single cells from rabbit sino-atrial node. J. Physiol. 1991, 434, 57–83. [Google Scholar]
- Sample Availability: Not available.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yaniv, Y.; Maltsev, V.A.; Ziman, B.D.; Spurgeon, H.A.; Lakatta, E.G. The “Funny” Current (If) Inhibition by Ivabradine at Membrane Potentials Encompassing Spontaneous Depolarization in Pacemaker Cells. Molecules 2012, 17, 8241-8254. https://doi.org/10.3390/molecules17078241
Yaniv Y, Maltsev VA, Ziman BD, Spurgeon HA, Lakatta EG. The “Funny” Current (If) Inhibition by Ivabradine at Membrane Potentials Encompassing Spontaneous Depolarization in Pacemaker Cells. Molecules. 2012; 17(7):8241-8254. https://doi.org/10.3390/molecules17078241
Chicago/Turabian StyleYaniv, Yael, Victor A. Maltsev, Bruce D. Ziman, Harold A. Spurgeon, and Edward G. Lakatta. 2012. "The “Funny” Current (If) Inhibition by Ivabradine at Membrane Potentials Encompassing Spontaneous Depolarization in Pacemaker Cells" Molecules 17, no. 7: 8241-8254. https://doi.org/10.3390/molecules17078241
APA StyleYaniv, Y., Maltsev, V. A., Ziman, B. D., Spurgeon, H. A., & Lakatta, E. G. (2012). The “Funny” Current (If) Inhibition by Ivabradine at Membrane Potentials Encompassing Spontaneous Depolarization in Pacemaker Cells. Molecules, 17(7), 8241-8254. https://doi.org/10.3390/molecules17078241