
Climate Change Impacts on Plant Phenology: Grapevine (Vitis vinifera) Bud Break in Wintertime in Southern Italy

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
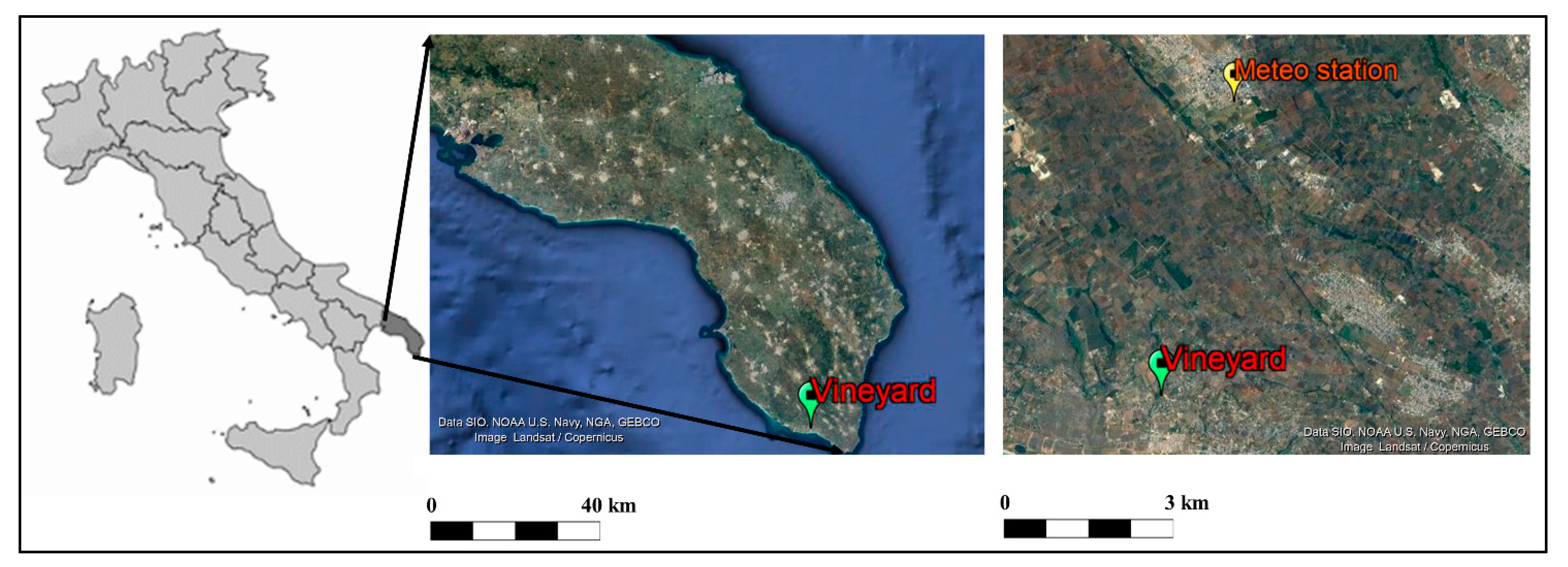
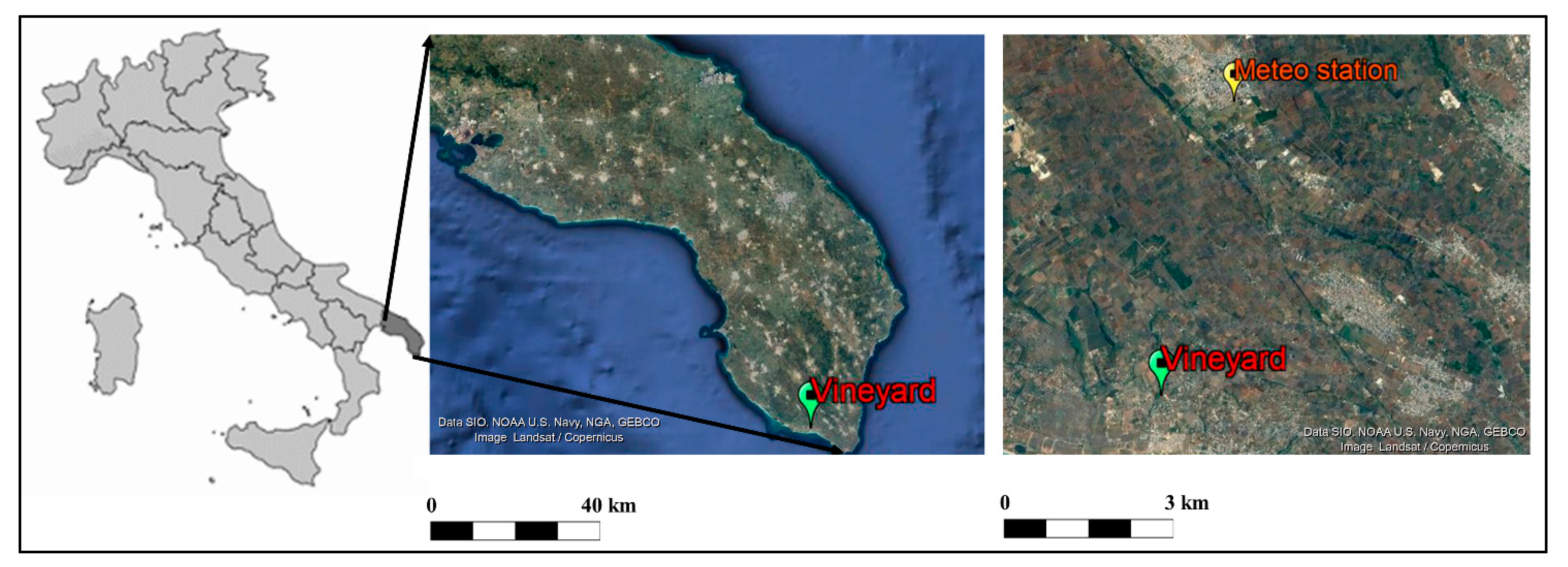
2.1. Plant Material and Experimental Design
2.2. Meteorological Data and Analysis
2.3. Plant Phenotyping
- Rx(w) = Reflectance (% calibrated on the reference blank)
- x = spectrum type: t0 = before reaction; Re = after Lugol reaction
- w = wavelength of interest (nm): 900 = normalization reference; 555 = starch–iodine complex absorption maximum.
2.4. SNP Genotyping
3. Results
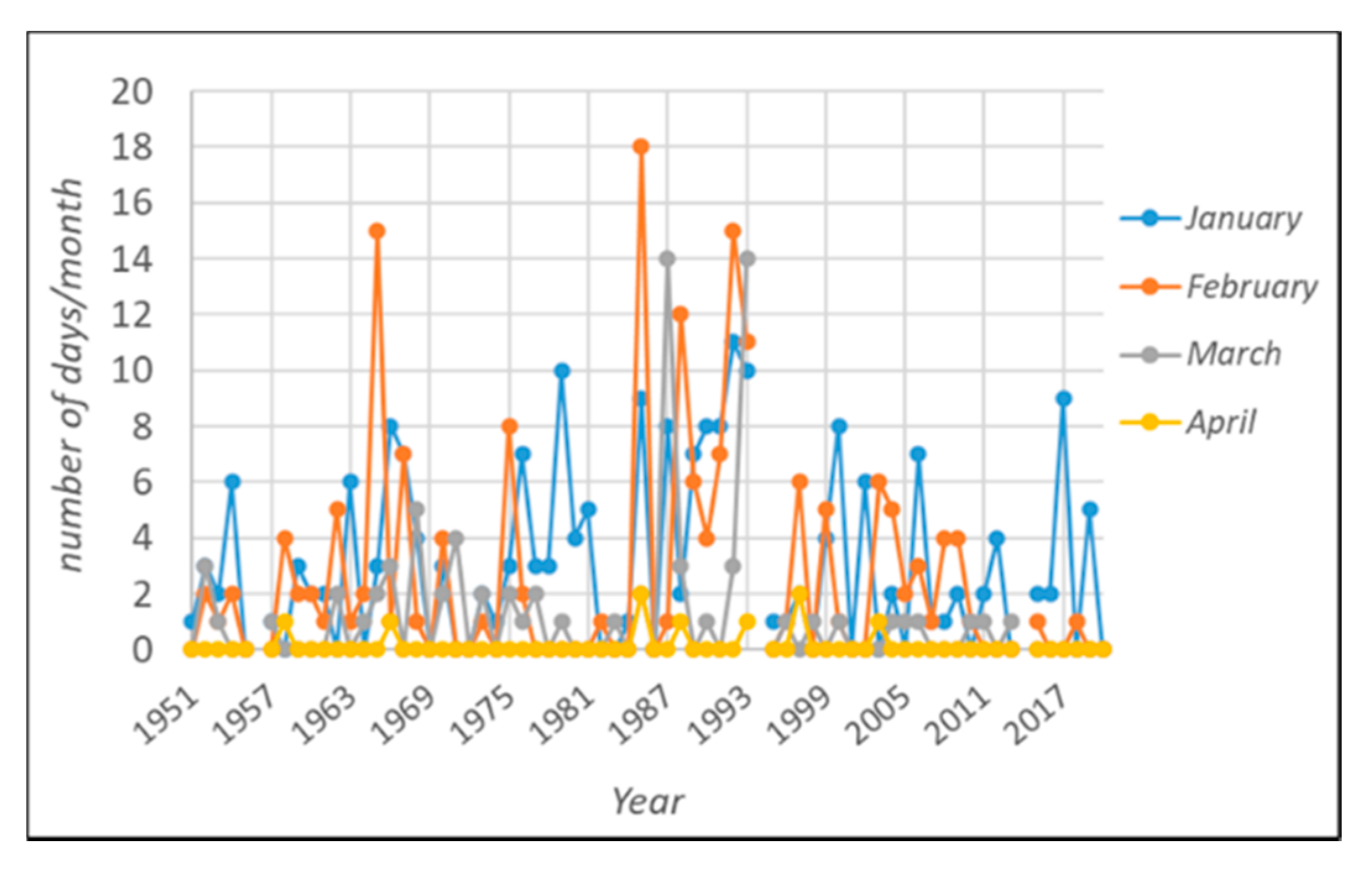
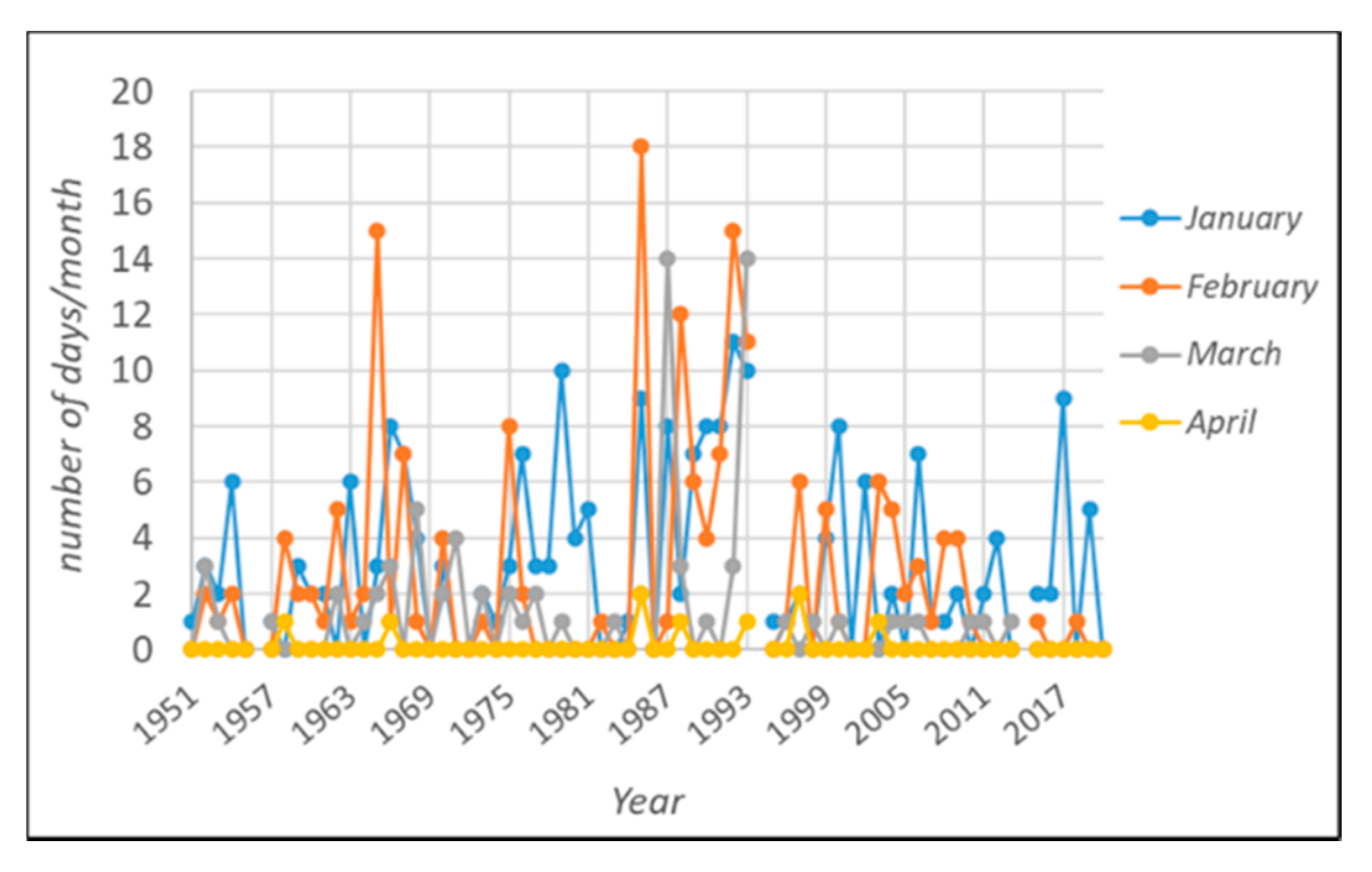
3.1. The Winter Warming
3.2. The Winter Budding Concerns the Entire Plant
3.3. Old Vineyards: A Source of Biodiversity
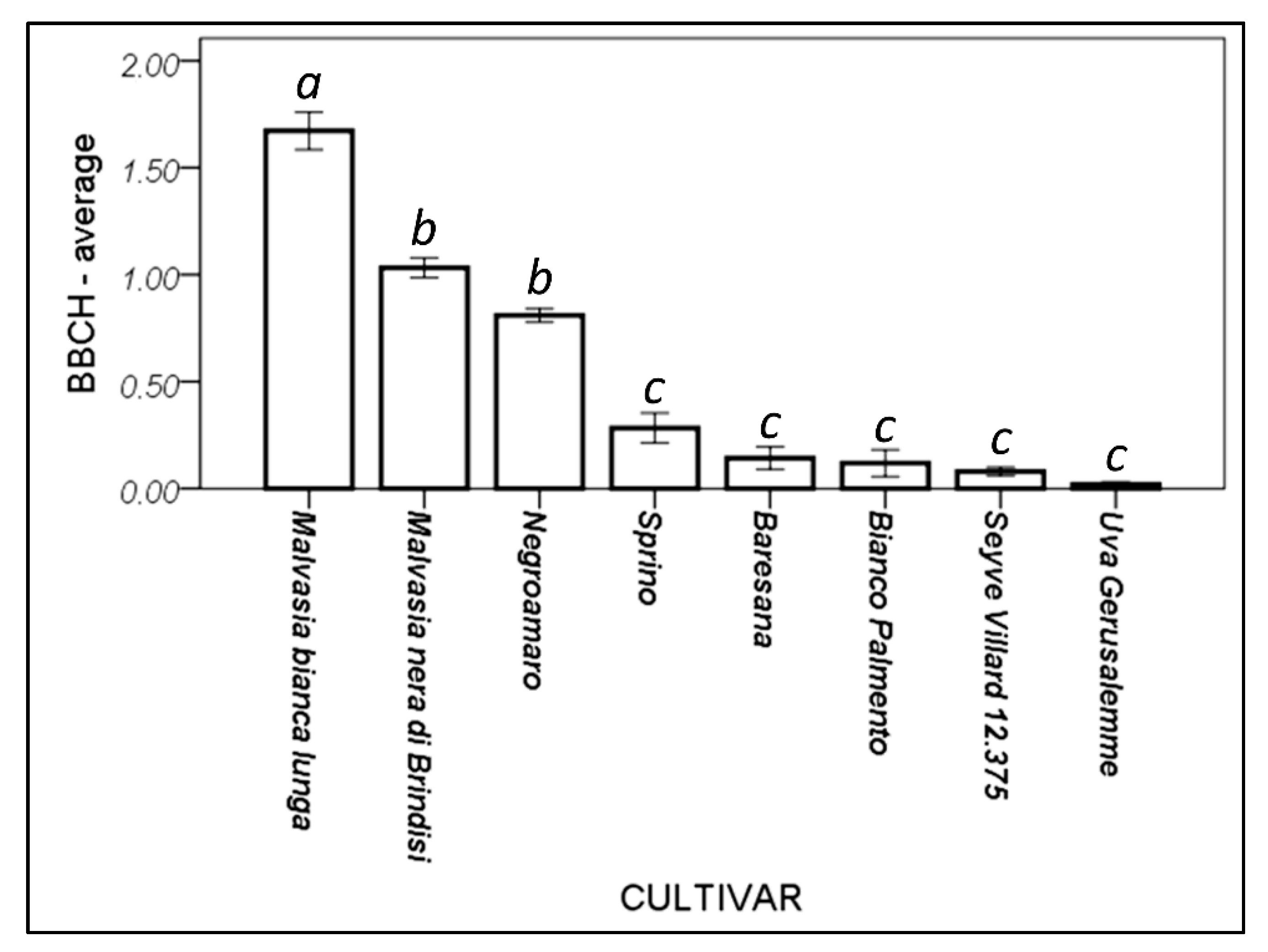
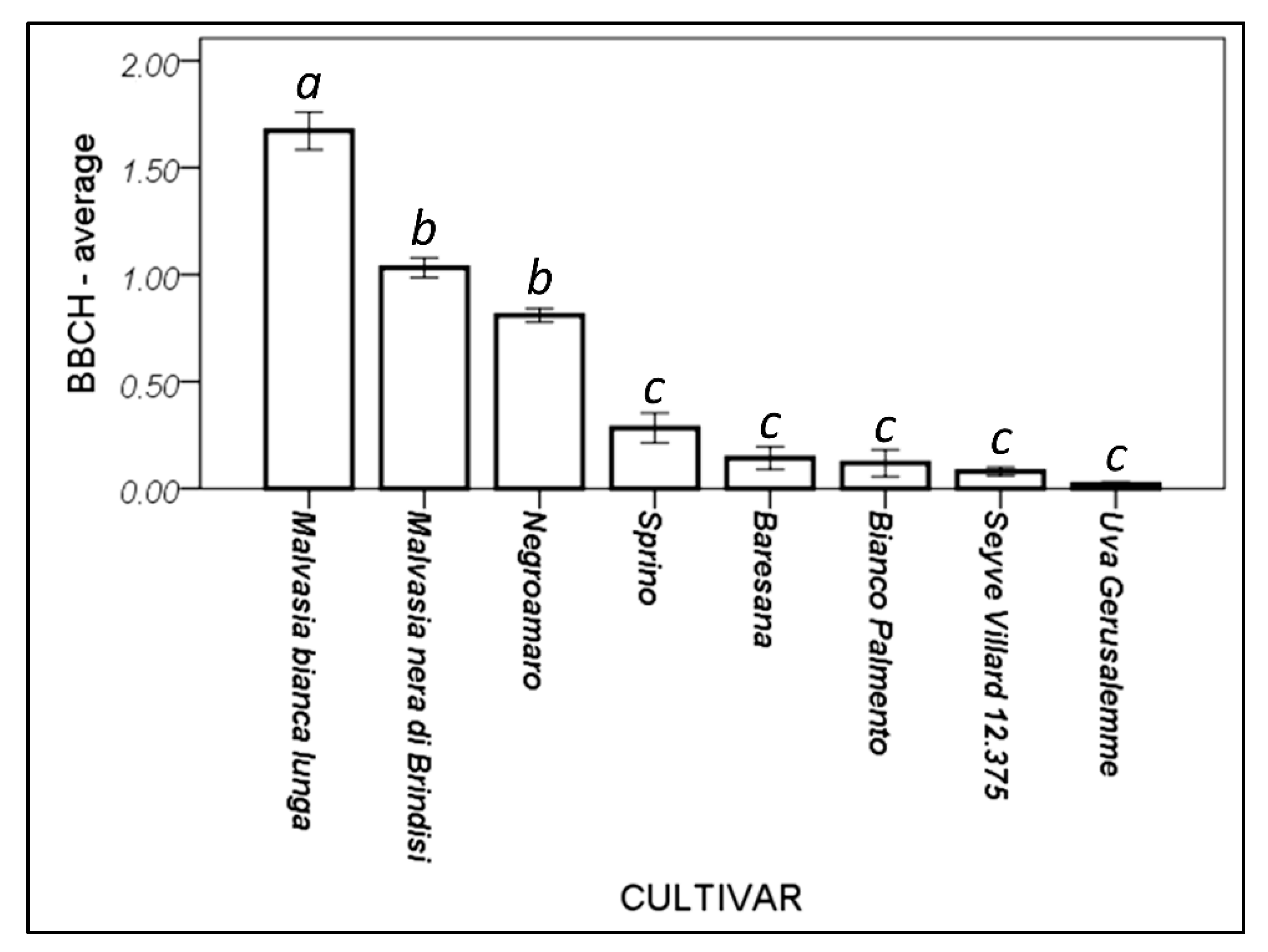
3.4. Different Cultivars Have Different Phenological Behavior
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reid, P.; Hari, R.; Beaugrand, G.; Livingstone, D.; Marty, C.; Straile, D.; Barichivich, J.; Goberville, E.; Adrian, R.; Aono, Y.; et al. Global impacts of the 1980s regime shift. Glob. Chang. Biol. 2016, 22, 682–703. [Google Scholar] [CrossRef] [Green Version]
- Sutton, R.T.; Dong, B. Atlantic Ocean influence on a shift in European climate in the 1990s. Nat. Geosci. 2012, 5, 788–792. [Google Scholar] [CrossRef]
- Mariani, L.; Parisi, S.G.; Cola, G.; Failla, O. Climate change in Europe and effects on thermal resources for crops. Int. J. Biometeorol. 2012, 56, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Koch, B.; Oehl, F. Climate change favors grapevine production in temperate zones. Agric. Sci. 2018, 9, 247–263. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.A.; Malheiro, A.C.; Pinto, J.G.; Jones, G.V. Macroclimate and viticultural zoning in Europe: Observed trends and atmospheric forcing. Clim. Res. 2012, 51, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Malheiro, A.; dos Santos, J.C.A.; Fraga, H.; Pinto, J.G. Climate change scenarios applied to viticultural zoning in Europe. Clim. Res. 2010, 43, 163–177. [Google Scholar] [CrossRef] [Green Version]
- van Leeuwen, C.; Darriet, P. The impact of climate change on viticulture and wine quality. J. Wine Econ. 2016, 11, 150–167. [Google Scholar] [CrossRef] [Green Version]
- Leolini, L.; Moriondo, M.; Fila, G.; Costafreda-Aumedes, S.; Ferrise, R.; Bindi, M. Late spring frost impacts on future grapevine distribution in Europe. Field Crop. Res. 2018, 222, 197–208. [Google Scholar] [CrossRef]
- Bucur, G.M.; Babes, A.C. Research on trends in extreme weather conditions and their effects on grapevine in Romanian viticulture. Bull. UASVM Hortic. 2016, 73, 126–134. [Google Scholar]
- Mosedale, J.R.; Wilson, R.; Maclean, I. Climate Change and Crop Exposure to Adverse Weather: Changes to Frost Risk and Grapevine Flowering Conditions. PLoS ONE 2015, 10, e0141218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wake, C.M.F.; Fennell, A. Morphological, physiological and dormancy responses of three Vitis genotypes to short photoperiod. Physiol. Plant. 2000, 109, 203–210. [Google Scholar] [CrossRef]
- Andreini, L.; Viti, R.; Scalabrelli, G. Study on the morphological evolution of bud break in Vitis vinifera L. Citeseer 2009, 48, 153–158. [Google Scholar]
- de Cortazar-Atauri, I.G.; Brisson, N.; Gaudillere, J.P. Performance of several models for predicting budburst date of grapevine (Vitis vinifera L.). Int. J. Biometeorol. 2009, 53, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Friend, A.; Trought, M.; Stushnoff, C.; Wells, G. Effect of delaying budburst on shoot development and yield of Vitis vinifera L. Chardonnay ‘Mendoza’ after a spring freeze event. Aust. J. Grape Wine Res. 2011, 17, 378–382. [Google Scholar] [CrossRef]
- Díaz-Riquelme, J.; Grimplet, J.; Martínez-Zapater, J.M.; Carmona, M.J. Transcriptome variation along bud development in grapevine (Vitis vinifera L.). BMC Plant Biol. 2012, 12, 181. [Google Scholar] [CrossRef] [Green Version]
- Charrier, G.; Ngao, J.; Saudreau, M.; Ameglio, T. Effects of environmental factors and management practices on microclimate, winter physiology, and frost resistance in trees. Front. Plant Sci. 2015, 6, 259. [Google Scholar] [CrossRef] [Green Version]
- Rubio, S.; Dantas, D.; Bressan-Smith, R.; Pérez, F.J. Relationship between Endodormancy and Cold Hardiness in Grapevine Buds. J. Plant Growth Regul. 2015, 35, 266–275. [Google Scholar] [CrossRef]
- Pellegrino, A.; Rogiers, S.; Deloire, A. Grapevine Latent Bud Dormancy and Shoot Development. IVES Tech. Rev. Vine Wine 2020. [Google Scholar] [CrossRef]
- Londo, J.P.; Martinson, T. Grapevine winter survival and prospects in an age of changing climate. Appell. Cornell 2016, 3, 1–7. [Google Scholar]
- Fuller, M.P.; Telli, G. An investigation of the frost hardiness of grapevine (Vitis vinfera L.) during bud break. Ann. Appl. Biol. 1999, 135, 135589–135595. [Google Scholar] [CrossRef]
- Filho, J.M.; Allebrandt, R.; de Bem, B.; Mudrei, P.; Macedo, T.; Schlemper, C.; Lerin, S.; Outemane, M.; Kretzschmar, A.; Rufato, L. Damage to ‘Cabernet Sauvignon’ after late frost in the southern Brazilian highlands. Acta Hortic. 2016, 1115, 211–216. [Google Scholar] [CrossRef]
- Friend, A.P.; Trought, M.C.T. Delayed winter spur-pruning in New Zealand can alter yield components of Mer-lot grapevines. Aust. J. Grape Wine Res. 2007, 13, 157–164. [Google Scholar] [CrossRef]
- Gatti, M.; Pirez, F.J.; Chiari, G.; Tombesi, S.; Palliotti, A.; Merli, M.C.; Poni, S. Phenology, Canopy Aging and Seasonal Carbon Balance as Related to Delayed Winter Pruning of Vitis vinifera L. cv. Sangiovese Grapevines. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, M.; Sadras, V.; Petrie, P. Late pruning and carry-over effects on phenology, yield components and berry traits in Shiraz. Aust. J. Grape Wine Res. 2017, 23, 390–398. [Google Scholar] [CrossRef]
- Brighenti, A.; Allebrandt, R.; Cipriani, R.; Malinovski, L.; de Bem, B.; Feldberg, N.; Silva, A. Using delayed winter pruning to prevent spring frost damage in ‘Chardonnay’ cultivar. Acta Hortic. 2017, 1157, 389–392. [Google Scholar] [CrossRef]
- Gatti, M.; Pirez, F.; Frioni, T.; Squeri, C.; Poni, S. Calibrated, delayed-cane winter pruning controls yield and significantly postpones berry ripening parameters in Vitis vinifera L. cv. Pinot Noir. Aust. J. Grape Wine Res. 2018, 24, 305–316. [Google Scholar] [CrossRef]
- Frioni, T.; Tombesi, S.; Silvestroni, O.; Lanari, V.; Bellincontro, A.; Sabbatini, P.; Gatti, M.; Poni, S.; Palliotti, A. Postbudburst Spur Pruning Reduces Yield and Delays Fruit Sugar Accumulation in Sangiovese in Central Italy. Am. J. Enol. Vitic. 2016, 67, 419–425. [Google Scholar] [CrossRef]
- Favero, A.C.; Angelucci de Amorim, D.; Vieira Da Mota, R.; Soares, A.M.; de Souza, C.R.; de Albuquerque Regina, M. Double-pruning of ‘Syrah’ grapevines: A management strategy to harvest wine grapes during the winter in the Brazilian Southeast. Vitis 2011, 50, 151–158. [Google Scholar]
- Palliotti, A.; Tombesi, S.; Silvestroni, O.; Lanari, V.; Gatti, M.; Poni, S. Changes in vineyard establishment and canopy management urged by earlier climate-related grape ripening: A review. Sci. Hortic. 2014, 178, 43–54. [Google Scholar] [CrossRef]
- Palliotti, A.; Frioni, T.; Tombesi, S.; Sabbatini, P.; Cruz-Castillo, J.G.; Lanari, V.; Silvestroni, O.; Gatti, M.; Poni, S. Double-Pruning Grapevines as a Management Tool to Delay Berry Ripening and Control Yield. Am. J. Enol. Vitic. 2017, 68, 412–421. [Google Scholar] [CrossRef]
- Silvestroni, O.; Lanari, V.; Lattanzi, T.; Palliotti, A. Delaying winter pruning, after pre-pruning, alters budburst, leaf area, photosynthesis, yield and berry composition in Sangiovese (Vitis vinifera L.). Aust. J. Grape Wine Res. 2018, 24, 478–486. [Google Scholar] [CrossRef]
- Martínez-Moreno, A.; Sanz, F.; Yeves, A.; Gil-Muñoz, R.; Martínez, V.; Intrigliolo, D.; Buesa, I. Forcing bud growth by double-pruning as a technique to improve grape composition of Vitis vinifera L. cv. Tempranillo in a semi-arid Mediterranean climate. Sci. Hortic. 2019, 256, 108614. [Google Scholar] [CrossRef]
- van Leeuwen, C.; Destrac-Irvine, A.; Dubernet, M.; Duchêne, E.; Gowdy, M.; Marguerit, E.; Pieri, P.; Parker, A.; de Rességuier, L.; Ollat, N. An Update on the Impact of Climate Change in Viticulture and Potential Adaptations. Agronomy 2019, 9, 514. [Google Scholar] [CrossRef] [Green Version]
- Maul, E.; Sudharma, K.N.; Kecke, S.; Marx, G.; Müller, C.; Audeguin, L.; Boselli, M.; Boursiquot, J.M.; Bucchetti, B.; Cabello, F.; et al. The European Vitis Database (www.eu-vitis.de)–a technical innovation through an online uploading and interactive modification system. Vitis 2012, 51, 79–85. [Google Scholar]
- Rustioni, L.; Cola, G.; Fiori, S.; Failla, O.; Bacilieri, R.; Maul, E.; Eiras Dias, J.E.; Brazão, J.; Kocsis, L.; Lorenzini, F.; et al. Application of standard methods for the grapevine (Vitis vinifera L.) phenotypic diversity exploration: Phenological traits. Acta Hortic. 2014, 1032, 253–260. [Google Scholar] [CrossRef]
- Rustioni, L.; Maghradze, D.; Popescu, C.F.; Cola, G.; Abashidze, E.; Aroutiounian, R.; Brazão, J.; Coletti, S.; Cornea, V.; Dejeu, L.; et al. First results of the European grapevine collections’ collaborative net-work: Validation of a standard enocarpological phenotyping method. Vitis 2014, 53, 219–226. [Google Scholar]
- Rustioni, L.; Cola, G.; Maghradze, D.; Abashidze, E.; Argiriou, A.; Aroutiounian, R.; Bacilieri, R. Description of the Vitis vinifera L. phenotypic variability in enocarpological traits by a Euro-Asiatic collaborative network among ampelo-graphic collections. Vitis 2019, 58, 37–46. [Google Scholar]
- Laucou, V.; Launay, A.; Bacilieri, R.; Lacombe, T.; Adam-Blondon, A.F.; Bérard, A.; Chauveau, A.; de Andrés, M.T.; Hausmann, L.; Ibáñez, J.; et al. Extended diversity analysis of cultivated grapevine Vitis vinifera with 10 K genome-wide SNPs. PLoS ONE 2018, 13, e0192540. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzis, G.; Mercati, F.; Bergamini, C.; Cardone, M.F.; Lupini, A.; Maucerim, A.; Caputo, A.R.; Abbate, L.; Barbagallo, M.G.; Antonacci, D.; et al. SNP genotyping elucidates the genetic diversity of Magna Graecia grapevine germplasm and its historical origin and dissemination. BMC Plant Biol. 2017, 19, 7. [Google Scholar]
- Cola, G.; Failla, O.; Maghradze, D.; Megrelidze, L.; Mariani, L. Grapevine phenology and climate change in Georgia. Int. J. Biometeorol. 2017, 61, 761–773. [Google Scholar] [CrossRef]
- Ferguson, J.C.; Tarara, J.M.; Mills, L.J.; Grove, G.G.; Keller, M. Dynamic thermal time model of cold hardiness for dormant grapevine buds. Ann. Bot. 2011, 107, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Nendel, C. Grapevine bud break prediction for cool winter climates. Int. J. Biometeorol. 2009, 54, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Meier, U. Growth stages of Mono- and Dicotyledonous plants BBCH. Monograph (2nd ed.). Germany: Federal Biological Research Centre of Agriculture. ISBN: 978-3-95547-071-5. Available online: https://www.julius-kuehn.de/media/Veroeffentlichungen/bbch%20epaper%20en/page.pdf (accessed on 7 November 2021). [CrossRef]
- Rustioni, L.; Ciacciulli, A.; Zulini, L.; Zuliani, E.; Sivilotti, P.; Herrera, J.C. Starch quantification in woody tissues by reflectance spectroscopy and on-solid iodine complexation. Sci. Hortic. 2017, 226, 117–121. [Google Scholar] [CrossRef]
- Rustioni, L.; Ciacciulli, A.; Grossi, D.; Brancadoro, L.; Failla, O. Stem xylem characterization for Vitis drought toler-ance. J. Agric. Food Chem. 2016, 64, 5317–5323. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Organero, G.; Gaforio, L.; García-Muñoz, S.; Cabello, F. Manual for Standarization of Vitis Descriptors; Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario (IMIDRA): Madrid, Spain, 2011; ISBN 978-84-7498-544-7. ISSN 1575-6092. [Google Scholar]
- de Lorenzis, G.; Chipashvili, R.; Failla, O.; Maghradze, D. Study of genetic variability in Vitis vinifera L. germplasm by high-throughput Vitis18kSNP array: The case of Georgian genetic resources. BMC Plant Biol. 2015, 15, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Onofrio, C.; Tumino, G.; Gardiman, M.; Crespan, M.; Bignami, C.; de Palma, L.; Barbagallo, M.G.; Muganu, M.; Morcia, C.; Novello, V.; et al. Parentage Atlas of Italian Grapevine Varieties as Inferred from SNP Genotyping. Front. Plant Sci. 2021, 11, 2265. [Google Scholar] [CrossRef] [PubMed]
- Kartschall, T.; Wodinski, M.; von Bloh, W.; Oesterle, H.; Rachimow, C.; Hoppmann, D. Changes in phenology and frost risks of Vitis vinifera (cv Riesling). Meteorol. Z. 2015, 24, 189–200. [Google Scholar] [CrossRef]
- Meier, M.; Fuhrer, J.; Holzkämper, A. Changing risk of spring frost damage in grapevines due to climate change? A case study in the Swiss Rhone Valley. Int. J. Biometeorol. 2018, 62, 991–1002. [Google Scholar] [CrossRef] [Green Version]
- Cola, G.; Mariani, L.; Maghradze, D.; Failla, O. Changes in thermal resources and limitations for Georgian viticulture. Aust. J. Grape Wine Res. 2020, 26, 29–40. [Google Scholar] [CrossRef]
- di Lena, B.; Farinelli, D.; Palliotti, A.; Poni, S.; DeJong, T.M.; Tombesi, S. Impact of climate change on the possible expansion of almond cultivation area pole-ward: A case study of Abruzzo, Italy. J. Hortic. Sci. Biotechnol. 2018, 93, 209–215. [Google Scholar] [CrossRef]
- Duchêne, E.; Schneider, C. Grapevine and climatic changes: A glance at the situation in Alsace. Agron. Sustain. Dev. 2005, 25, 93–99. [Google Scholar] [CrossRef]
- Tomasi, D.; Jones, G.V.; Giust, M.; Lovat, L.; Gaiotti, F. Grapevine phenology and climate change: Relationships and trends in the Veneto Region of Italy for 1964–2009. Am. J. Enol. Vitic. 2011, 62, 329–339. [Google Scholar] [CrossRef]
- Fraga, H.; Pinto, J.G.; Santos, J.A. Climate change projections for chilling and heat forcing conditions in European vineyards and olive orchards: A multi-model assessment. Clim. Chang. 2019, 152, 179–193. [Google Scholar] [CrossRef]
- Ma, Q.; Huang, J.; Hänninen, H.; Berninger, F. Divergent trends in the risk of spring frost damage to trees in Europe with recent warming. Glob. Chang. Biol. 2019, 25, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Dokoozlian, N.K. Chilling temperature and duration interact on the budbreak of ‘Perlette’ grapevine cuttings. HortScience 1999, 34, 1054–1056. [Google Scholar] [CrossRef] [Green Version]
- Zapata, C.; Deleens, E.; Chaillou, S.; Magné, C. Partitioning and mobilization of starch and N reserves in grapevine (Vitis vinifera L.). J. Plant Physiol. 2004, 161, 1031–1040. [Google Scholar] [CrossRef] [PubMed]
- Maghradze, D.; Rustioni, L.; Scienza, A.; Faillaa, O. Phenological diversity of Georgian grapevine cultivars in Northern Italy. J. Am. Pomol. Soc. 2012, 66, 56–67. [Google Scholar]
- Maghradze, D.; Vashakidze, L.; Abashidze, E.; Chipashvili, R.; Mdinaradze, I.; Failla, O.; Rustioni, L.; de Lorenzis, G.; Scienza, A.; Maul, E. Multidisciplinary study of traditional grape cultivars from kartli province of georgia (the caucasus region) and activities for their preservation. Acta Hortic. 2014, 235–241. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Minimum Temperatures | ||||||||||
| 2019/2020 | 1991–2020 | 1961–1990 | Significance of the Difference (1961–1990 vs. 1991–2020) | |||||||
| Aver. | Min. | Max. | St. Dev. | Aver. | Min. | Max. | St. Dev. | |||
| November | 12.31 | 10.0 | 7.1 | 12.5 | 1.6 | 8.6 | 4.8 | 11.5 | 1.6 | 0.007 ** |
| December | 8.96 | 6.3 | 1.0 | 9.7 | 1.9 | 5.7 | 0.9 | 9.6 | 1.6 | 0.184 |
| January | 5.75 | 4.7 | 1.9 | 7.8 | 1.8 | 4.3 | 1.8 | 7.5 | 1.5 | 0.576 |
| February | 6.10 | 5.1 | 0.4 | 9.0 | 2.0 | 4.6 | 0.3 | 7.8 | 1.7 | 0.295 |
| March | 7.52 | 7.3 | 2.6 | 10.6 | 1.7 | 6.2 | 1.7 | 8.4 | 1.5 | 0.007 ** |
| April | 9.87 | 10.1 | 5.7 | 12.8 | 1.6 | 8.8 | 7.2 | 10.8 | 0.9 | 0.000 *** |
| (b) Maximum Temperatures | ||||||||||
| 2019/2020 | 1991–2020 | 1961–1990 | Significance of the Difference (1961–1990 vs. 1991–2020) | |||||||
| Aver. | Min. | Max. | St. Dev. | Aver. | Min. | Max. | St. Dev. | |||
| November | 19.56 | 17.7 | 15.4 | 19.9 | 1.2 | 16.4 | 13.3 | 18.7 | 1.1 | 0.001 *** |
| December | 15.03 | 13.8 | 9.0 | 16.5 | 1.6 | 12.9 | 10.2 | 14.6 | 1.0 | 0.021 * |
| January | 13.57 | 12.6 | 10.1 | 14.8 | 1.2 | 11.7 | 7.7 | 14.2 | 1.3 | 0.035 * |
| February | 15.44 | 13.3 | 10.1 | 16.2 | 1.7 | 12.3 | 8.1 | 15.0 | 1.4 | 0.016 * |
| March | 16.19 | 15.6 | 12.3 | 19.0 | 1.3 | 14.5 | 9.8 | 17.9 | 1.9 | 0.012 * |
| April | 18.56 | 19.0 | 14.9 | 22.4 | 1.6 | 17.8 | 15.6 | 20.5 | 1.4 | 0.002 *** |
| (c) Cumulative Active Temperatures | ||||||||||
| 2019/2020 | 1991–2020 | 1961–1990 | Significance of the Difference (1961–1990 vs. 1991–2020) | |||||||
| Aver. | Min. | Max. | St. Dev. | Aver. | Min. | Max. | St. Dev. | |||
| November | 482.90 | 382.7 | 293.6 | 491.9 | 62.3 | 324.6 | 164.6 | 429.0 | 62.5 | 0.003 *** |
| December | 325.05 | 226.8 | 31.6 | 325.1 | 71.7 | 195.4 | 64.7 | 338.8 | 57.2 | 0.079 |
| January | 202.75 | 163.8 | 68.8 | 272.0 | 62.1 | 142.7 | 51.2 | 249.3 | 53.1 | 0.362 |
| February | 226.95 | 171.7 | 35.5 | 319.6 | 70.1 | 144.6 | 35.2 | 245.0 | 51.2 | 0.090 |
| March | 298.30 | 282.6 | 120.7 | 435.4 | 66.5 | 234.5 | 110.1 | 337.9 | 59.7 | 0.006 ** |
| April | 394.65 | 408.7 | 214.8 | 540.5 | 69.9 | 343.1 | 270.9 | 442.4 | 49.2 | 0.000 *** |
| (d) Occurrence of Frost Events (Number of Days with Temperatures Lower than 0) | ||||||||||
| 2019/2020 | 1991–2020 | 1961–1990 | Significance of the Difference (1961–1990 vs. 1991–2020) | |||||||
| Aver. | Min. | Max. | St. Dev. | Aver. | Min. | Max. | St. Dev. | |||
| November | 0 | 0.0 | 0 | 1 | 0.2 | 0.3 | 0 | 2 | 0.6 | 0.018 * |
| December | 0 | 2.0 | 0 | 11 | 3.1 | 2.2 | 0 | 15 | 3.3 | 0.811 |
| January | 0 | 3.1 | 0 | 11 | 3.5 | 3.5 | 0 | 10 | 3.2 | 0.758 |
| February | 0 | 2.6 | 0 | 15 | 3.7 | 3.0 | 0 | 18 | 4.7 | 0.820 |
| March | 0 | 0.9 | 0 | 14 | 2.7 | 1.6 | 0 | 14 | 2.7 | 0.377 |
| April | 0 | 0.1 | 0 | 2 | 0.4 | 0.1 | 0 | 2 | 0.4 | 0.901 |
| BBCH Maximum | BBCH Average | BBCH Standard Deviation | Shoot Humidity | Starch Content | |
|---|---|---|---|---|---|
| BBCH Maximum | 1 | 0.796 | 0.876 | 0.393 | −0.393 |
| BBCH Average | 0.796 | 1 | 0.888 | 0.562 | −0.5 |
| BBCH Standard Deviation | 0.876 | 0.888 | 1 | 0.401 | −0.457 |
| Shoot humidity | 0.393 | 0.562 | 0.401 | 1 | −0.457 |
| Starch content | −0.393 | −0.5 | −0.457 | −0.404 | −0.457 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dinu, D.G.; Ricciardi, V.; Demarco, C.; Zingarofalo, G.; De Lorenzis, G.; Buccolieri, R.; Cola, G.; Rustioni, L. Climate Change Impacts on Plant Phenology: Grapevine (Vitis vinifera) Bud Break in Wintertime in Southern Italy. Foods 2021, 10, 2769. https://doi.org/10.3390/foods10112769
Dinu DG, Ricciardi V, Demarco C, Zingarofalo G, De Lorenzis G, Buccolieri R, Cola G, Rustioni L. Climate Change Impacts on Plant Phenology: Grapevine (Vitis vinifera) Bud Break in Wintertime in Southern Italy. Foods. 2021; 10(11):2769. https://doi.org/10.3390/foods10112769
Chicago/Turabian StyleDinu, Daniel Grigorie, Valentina Ricciardi, Cosimo Demarco, Gianroberto Zingarofalo, Gabriella De Lorenzis, Riccardo Buccolieri, Gabriele Cola, and Laura Rustioni. 2021. "Climate Change Impacts on Plant Phenology: Grapevine (Vitis vinifera) Bud Break in Wintertime in Southern Italy" Foods 10, no. 11: 2769. https://doi.org/10.3390/foods10112769
APA StyleDinu, D. G., Ricciardi, V., Demarco, C., Zingarofalo, G., De Lorenzis, G., Buccolieri, R., Cola, G., & Rustioni, L. (2021). Climate Change Impacts on Plant Phenology: Grapevine (Vitis vinifera) Bud Break in Wintertime in Southern Italy. Foods, 10(11), 2769. https://doi.org/10.3390/foods10112769