3.1. Total Soluble Phenolic Content and Phenolic Profile
The high perishability and spoilage of vegetables is a major post-harvest challenge that hinders their effective integration as both fresh and processed food in health-focused food solution strategies. A major constrain is that in addition to high perishability occurring at storage stages, there is a deterioration of nutritional and other health relevant qualities due to the chemical changes of bioactive compounds in stored vegetables. Specifically, the composition and biological activity of phenolic compounds undergo significant changes during post-harvest handling and storage. Additionally, different food processing methods affect the content and bioactivity of phenolic compounds in processed foods. Therefore, advancing strategies to improve the stability and retention of phenolic metabolites and associated health protective food qualities of vegetables such as sweet potatoes is important for advancing high-value food applications. Fermentation using beneficial microorganism is a traditional food processing method that improves the human health relevant food qualities of plant-based food substrates and often enhances food quality and shelf-life.
In the current study, a beneficial LAB (L. plantarum) strain was rationally recruited to bio-transform aqueous extracts of different fleshed-colored sweet potatoes to improve their stability and retention of phenolic bioactive compounds and associated human health benefits. Three different flesh-colored sweet potato cultivars, which were previously screened and optimized, were selected for LAB-based liquid state fermentation (72 h). The total soluble phenolic (TSP) content of fermented and unfermented sweet potato was determined using the Folin-Ciocalteu reagent-based method. Additionally, the impact of changes in pH during fermentation on TSP content was also determined by comparing sweet potato extracts with natural pH and after adjusting to neutral pH (6.0) at all fermentation time points.
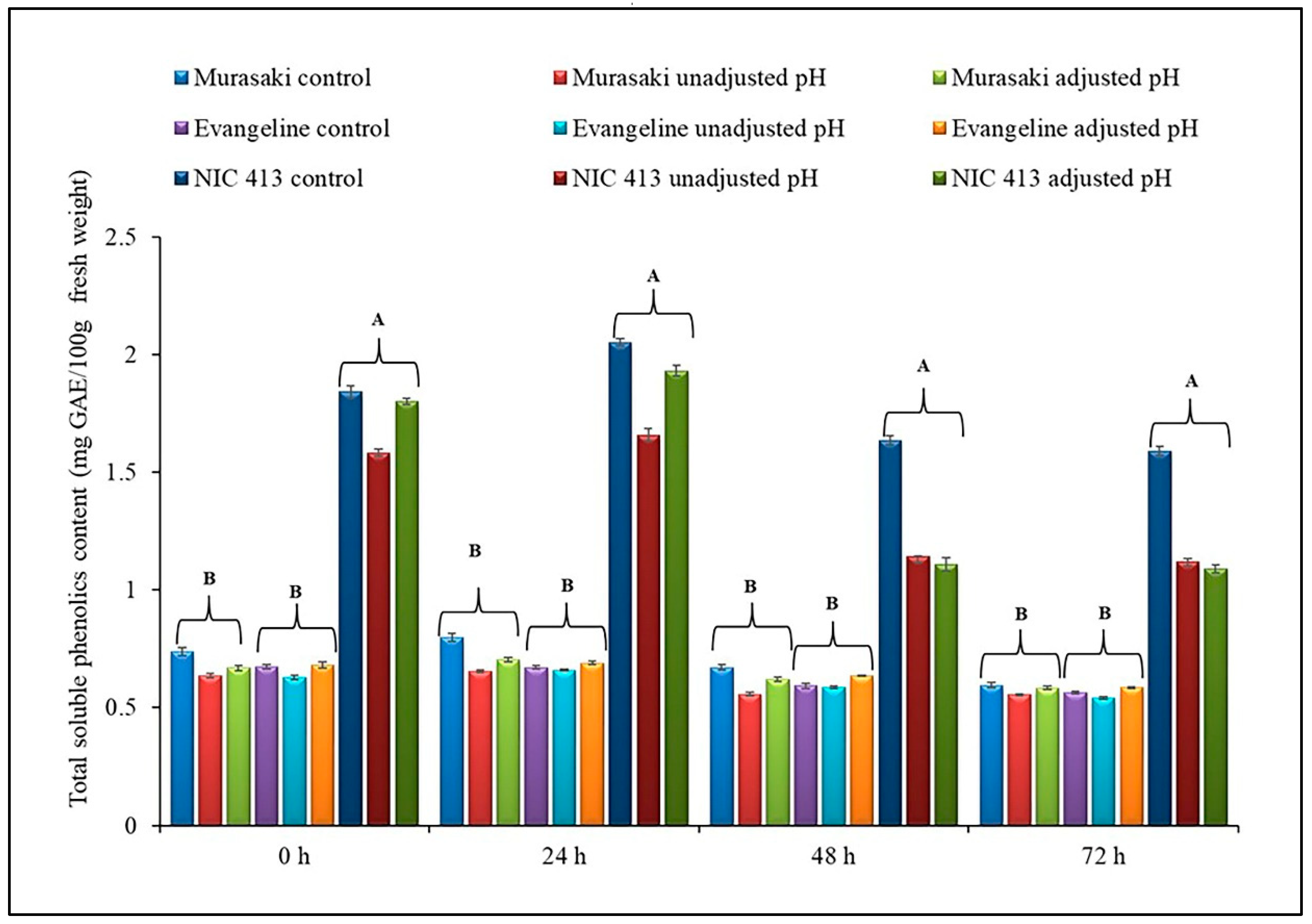
Overall, a higher retention of total soluble phenolic (TSP) content was observed among all three sweet potato cultivars during 72 h of fermentation (
Figure 1). Interestingly, the TSP content of fermented sweet potato was generally like the unfermented (control) sample even after adjusting to neutral pH. Analysis of variance showed statistically significant difference in TSP content among the three sweet potato cultivars, while the fermentation and interaction between cultivar × fermentation had no statistically significant effect. The differences in TSP content among the cultivars was possibly due to their different flesh colors, as observed earlier in our cultivar screening study.
Among the different sweet potato cultivars, the purple fleshed NIC-413 had a significantly (p < 0.05) higher TSP content across all fermentation time points. A slight improvement of TSP content was observed in purple-fleshed cultivar after 24 h fermentation and then it gradually reduced from 24–72 h. However, the reduction of TSP content was not statistically different when compared to the results of 0 h and unfermented sample.
In a previous study, Wu et al. [
9] reported an increased phenolic acid content in sweet potato (purple-fleshed) fermented with milk LAB cultures. In another study, an increase in phenolic content was reported for fermented sweet potato, when compared to the unfermented raw and boiled samples [
12]. However, the changes in phenolic content in these studies might be related to the changes in phenolic composition, specifically the release of some bound fractions after fermentation. The current TSP content results and those from previous studies suggest that LAB-based fermentation is an effective strategy to improve the retention and stability of phenolics and that this biotransformation strategy can be targeted to design sweet potato-based functional foods and beverages with probiotic benefits. In addition to TSP content, it is important to understand the effect of fermentation on the composition of phenolic compounds, as this biotransformation process changes the mobility and bioavailability of specific phenolics in food matrices.
In this study, catechin and gallic acid were found in all three sweet potato cultivars irrespective of fermentation treatment and time points (
Table 1 and
Supplementary File). In addition, protocatechuic acid was observed in purple-fleshed NIC-413, while chlorogenic acid was detected in orange-fleshed Evangeline and purple-fleshed NIC-413, specifically in the unfermented and fermented (with pH adjusted) samples. Previously, Ateea et al. [
20] and Padda and Picha [
21] also reported chlorogenic acid as major phenolic acid found in different sweet potato cultivars. In another study, Shen et al. [
12] reported ferulic acid and p-coumaric acid in LAB fermented sweet potato, while it was not present in unfermented sweet potato extracts. However, in the current study, ferulic acid and p-coumaric acid were not found, which might be due to use of food grade (cold water) extraction and the different HPLC analytical protocol. Additionally, dihydroxybenzoic acid was only found in the unfermented and 0 h fermented samples of off-white fleshed Murasaki and the 0 h unfermented sample of Evangeline. Significant changes in the contents of individual phenolic compounds during fermentation were also observed. Enhanced catechin content was found in fermented sample of all three sweet potato cultivars at 24, 48, and 72 h fermentation time points, and it was statistically significant when compared with unfermented samples. Similarly, the chlorogenic acid content of NIC-413 also increased at 24 and 48 h fermentation time points, specifically in fermented and pH-adjusted samples. Overall, a lower gallic acid content was found in fermented sweet potato samples when compared to unfermented extracts. The improvement in catechin and chlorogenic acid contents in select sweet potato cultivars with LAB fermentation has potential relevance in terms of its human health protective benefits, as both phenolic compounds are considered as potent dietary antioxidants with diverse functional benefits. Significant anti-inflammatory and redox protective functionalities of isolated and purified catechin [
22,
23] and chlorogenic acid [
24,
25] were previously reported. Additionally, anti-diabetic and anti-hypertensive properties were also found in plant foods that are rich in catechin and chlorogenic acid [
26,
27,
28].
Therefore, it is also important to investigate the effect of the LAB-based fermentation of sweet potatoes rich in different phenolics with human health relevant functional qualities such as antioxidant, anti-hyperglycemic, and anti-hypertensive properties.
3.2. Total Antioxidant Activity
The total antioxidant activity of unfermented and LAB-fermented sweet potato samples was determined using two different assays: 2,2-Dipheny-1-Picrylhydrazyl (DPPH) free radical scavenging assay and 2,2-Azino-bis-(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) radical cation assay.
Statistically significant (
p < 0.05) differences in antioxidant activity (DPPH assay) between sweet potato cultivars, fermentation time points, and interactions between cultivars × fermentation time points were observed (
Table 2). The highest total antioxidant activity (DPPH-based) was observed in the fermented and unadjusted pH sample of purple-fleshed (NIC-413) (83.82%) sweet potato followed by the unfermented (80.52%) and fermented with an adjusted pH (70.8%) samples of the same cultivar after 24 h of fermentation.
However, for the other two sweet potato cultivars, high antioxidant activity was observed in 0 h sample when compared to other fermentation time points. Interestingly, even after 72 h of fermentation, the fermented sweet potato sample of Murasaki and Evangeline showed similar antioxidant activity (DPPH-based) as the 24 h and 48 h time points. Therefore, the results of this study indicated that LAB-based fermentation is an effective strategy to retain and even improve the antioxidant activity of sweet potatoes.
Previously, many in vitro studies have found high antioxidant activities in anthocyanins rich purple sweet potatoes along with ameliorative effects and antihypertensive properties [
29,
30]. Wu et al. [
9] reported significantly high (
p < 0.01) free radical scavenging activity in the Chingshey purple sweet potato after fermentation with three different strains of
Lactobacillus such as
L. gasseri, L. acidophilus, and
L. delbrueckii subsp.
lactis. In this this study, improved antioxidant activities, higher GABA (gamma-aminobutyric acid) concentration, anthocyanins, and organic acids content were observed in fermented matrix developed from the Chingshey purple sweet potato. Similarly, Zhong-hue and Jie [
31] observed high superoxide anion radical scavenging activity and high Fe
+3 ions reducing capacity in wine derived from purple fleshed sweet potato.
Like the results of TSP content and DPPH-based antioxidant activity, statistically significant (
p < 0.05) differences in antioxidant activity (ABTS-based assay) were observed between sweet potato cultivars × fermentation time points (
Table 2). In general, even after 72 h fermentation, high antioxidant activity was observed in purple-fleshed (NIC-413) and orange-fleshed (Evangeline) sweet potato samples, while mean antioxidant activity was reduced in the off-white-fleshed (Murasaki) sweet potato sample. However, the fermented sample of Murasaki (pH adjusted) had significantly higher antioxidant activity when compared to the unfermented control sample after 72 h fermentation. Therefore, the results of this study indicated that LAB based fermentation has a positive effect on retaining and improving the antioxidant activity of sweet potatoes; thus, this strategy can be targeted for designing sweet potato-based functional foods and beverages that are particularly relevant for their antioxidant-linked anti-inflammatory benefits. Managing chronic oxidative stress is key for preventing and halting the progression of type 2 diabetes pathogenesis. Furthermore, it is also important to find and determine other functional benefits like anti-hyperglycemic and anti-hypertensive properties in order to design functional foods and ingredients from fermented and bioprocessed sweet potatoes targeting wider anti-diabetic benefits.
To understand the potential anti-hyperglycemic property of fermented sweet potato samples, α-amylase enzyme inhibitory activity was determined using an in vitro assay model. Moderate to high α-amylase enzyme inhibitory activity (ranging from 67.76% to 89.93%) was observed in the fermented and unfermented sweet potato samples of three cultivars with different flesh colors (
Table 3).
A significant (
p < 0.05) dose-dependent response [undiluted, half-diluted, and one-fifth diluted] in α-amylase enzyme inhibitory activity was also observed in fermented and unfermented samples [results of undiluted and half-diluted sample are presented in
Table 3 and a one-fifth dilution that had no inhibition was not included]. Overall, cultivar differences, fermentation, and the pH of the sample had statistically significant effect on α-amylase enzyme inhibitory activity (
p < 0.05); however, no statistically significant difference was observed between cultivars × fermentation time point interactions.
The α-amylase enzyme inhibitory activity gradually decreased from 0 to 72 h in most samples. The rate of reduction was significantly higher for the purple-fleshed sweet potato cultivar (NIC-413). The fermented sweet potato sample with natural pH had higher α-amylase enzyme inhibitory activity at most fermentation time points, and this was primarily due the interference of acidic condition. Among three sweet potato cultivars, the fermented sample of Murasaki had higher α-amylase enzyme inhibitory activity even after 48 and 72 h of fermentation. Therefore, for anti-hyperglycemic benefits, LAB-based fermentation can only be targeted for select sweet potato cultivars.
Previously, a reduction in α-amylase enzyme inhibitory activity was observed in fermented yam [
32]. Aligning these studies, the higher phenolic-linked antioxidant activity and moderate α-amylase enzyme inhibitory activity of fermented, purple-fleshed sweet potato (NIC-413) have potential relevance in health focused food applications, especially for improvements in overall dietary interventions to counter chronic oxidative stress and chronic hyperglycemia in the early stages of type 2 diabetes.
The potential inhibitory activity of another key anti-hyperglycemic relevant enzyme in fermented and unfermented sweet potato samples, α-glucosidase, was also determined using an in vitro assay model. In this study, low to moderate (13–55%) α-glucosidase enzyme inhibitory activity was observed in fermented and unfermented sweet potato samples (
Table 4). Additionally, a dose-dependent [undiluted, half-diluted, and one-fifth diluted] response in α-glucosidase enzyme inhibitory activity was also found in LAB-fermented and unfermented sweet potato sample. The results of undiluted and half diluted samples are presented in
Table 4 and the one-fifth dilution, which had no inhibition, was not included. Interestingly, the α-glucosidase enzyme inhibitory activity of Murasaki (off-white-fleshed) sweet potato improved after 72 h fermentation, while for orange-fleshed (Evangeline) and purple-fleshed (NIC-413) cultivars, it remained statistically at par between 0 to 72 h fermentation time points. Overall, the results of α-glucosidase enzyme inhibitory activity showed statistically significant (
p < 0.05) differences among cultivars, between fermentation time points, and cultivars × fermentation time points interactions in undiluted samples. The moderate α-amylase and α-glucosidase enzyme inhibitory activities in the fermented Murasaki sweet potato sample after 72 h of fermentation have potential relevance for using LAB fermentation to improve the anti-hyperglycemic functional benefits of off-white sweet potato cultivars. However, future studies with different white and off-white sweet potato cultivars are needed to further confirm the potential anti-hyperglycemic benefits of LAB-based fermentation strategy.
The higher retention of α-glucosidase enzyme inhibitory activity along with the high phenolic-linked antioxidant activity in select sweet potatoes after 48 and 72 h LAB-based fermentation are the key findings of this study. The higher retention and stability of phenolic content may be directly linked, and relevant to retaining and improving α-glucosidase enzyme inhibitory activity in fermented sweet potato samples. Previously, the presence of specific phenolic compounds such as 6-O-caffeoylsophorose in combination with the acylation of phenolics to sugar in fermented sweet potato were found to be associated with reduced postprandial glucose levels in rat models [
33]. Therefore, the total phenolic content and changes in the composition of phenolic acids during fermentation through the release of bound phenolics might have significant impact, potentially determining the anti-hyperglycemic and other human health relevant functional qualities of fermented plant-based foods.
Overall, the results of the present study suggest that LAB-based fermentation can be targeted to improve the stability and retention of phenolic bioactive linked antioxidant and anti-hyperglycemic functional qualities in select sweet potato cultivars, especially for the off-white-fleshed sweet potato cultivar (Murasaki).
3.4. Viable Cell Count of Lactiplantibacillus plantarum in Fermented Sweet Potatoes
The presence of active LAB counts in fermented food and beverage sample is important for their potential probiotic and gut health benefits. In this present study, viable cell counts of
L. plantarum in fermented sweet potato sample were determined at different time points (0, 24, 48, and 72 h during fermentation). Maximum counts of
L. plantarum were observed during the initial stages of fermentation (0 and 24 h) when compared to later periods (48 and 72 h), which could be due to the presence of more nutrients at the initial fermentation stage (
Table 6 and
Figure 2).
Overall, there was a reduction in colony forming units of LAB during fermentation from 24 to 72 h. The decrease in the viable cell counts might be due to the differences in pH or due to the declining growth of lactic acid bacteria after consuming all available nutrients that are required to sustain their growth. Significant differences in the viable cell counts of
L. plantarum due to different sweet potato substrate (cultivar differences) sources were observed, as Murasaki and NIC-413 supported higher growth (
Table 6) at the initial fermentation stages. Therefore, for probiotic application, 24 h fermentation of sweet potato and the selection of the right cultivar like Murasaki might be optimum, while a longer period of LAB fermentation can be targeted for other health-focused food and therapeutic applications. Furthermore, other plant-based foods with different fiber, oligosaccharide, and bioactive profiles like whole grains, beans, and root vegetables (parsnip, rutabaga) can also be integrated with fermented sweet potato to improve the growth of LAB and to enhance the probiotic and human gut health benefits. This fermentation strategy and food synergies can be targeted to design novel sweet potato-based fermented foods (bio-transformed flours for fries and whole fermented sweet potato foods and beverages) and as functional ingredients to add into breads, soups, and commercial baby foods for wider health benefits.


 ,
, 





{kind=link}
{kind=link}
{kind=link}