Connective Core Structures in Cognitive Networks: The Role of Hubs

Abstract
1. Introduction
2. Results
2.1. Net Entropy Individual Contribution
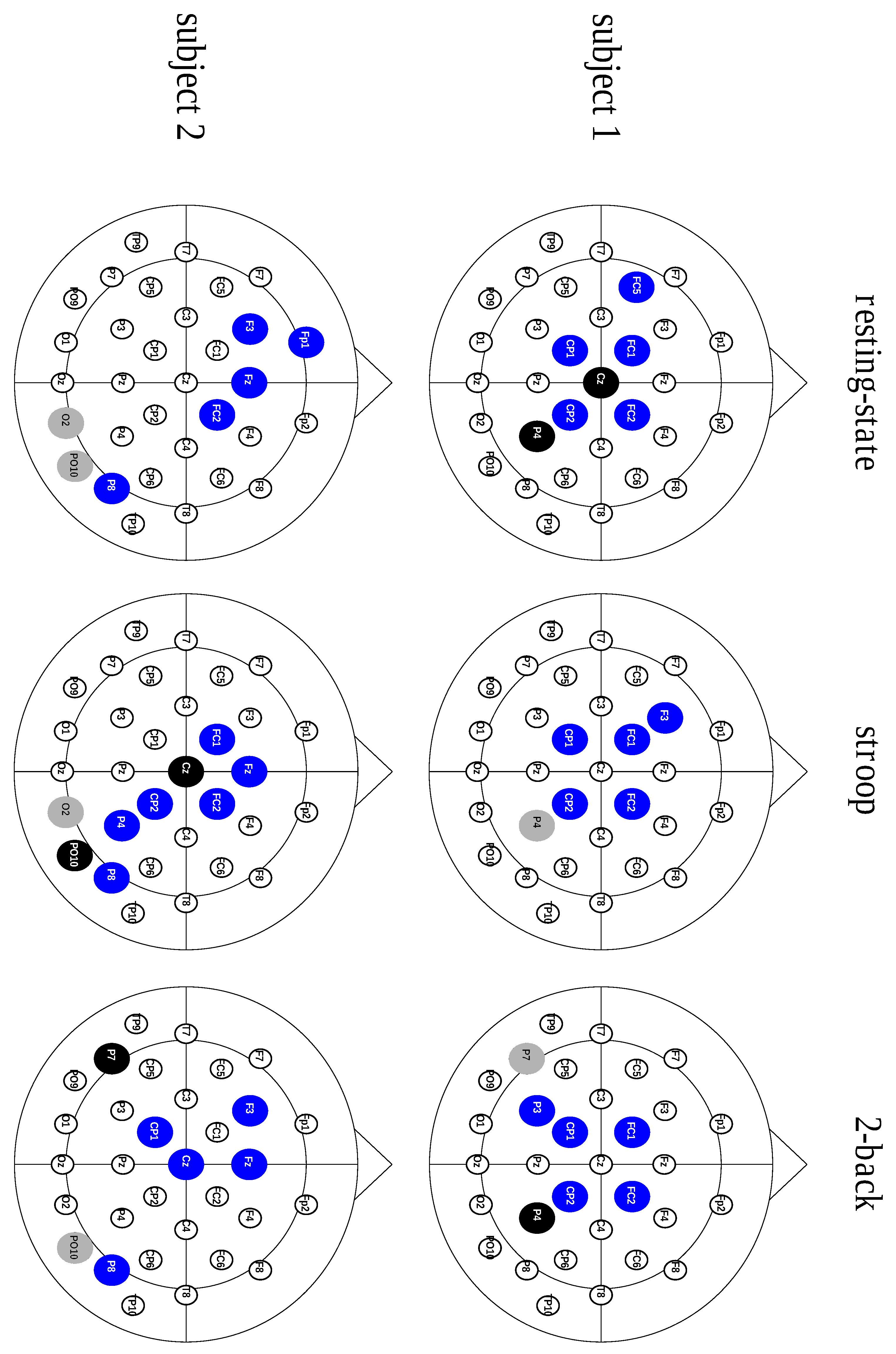
2.2. Divergent Hubs Play a Role in the Brain Network
2.3. The Connective Core Is Predominantly Formed by Divergent Hubs
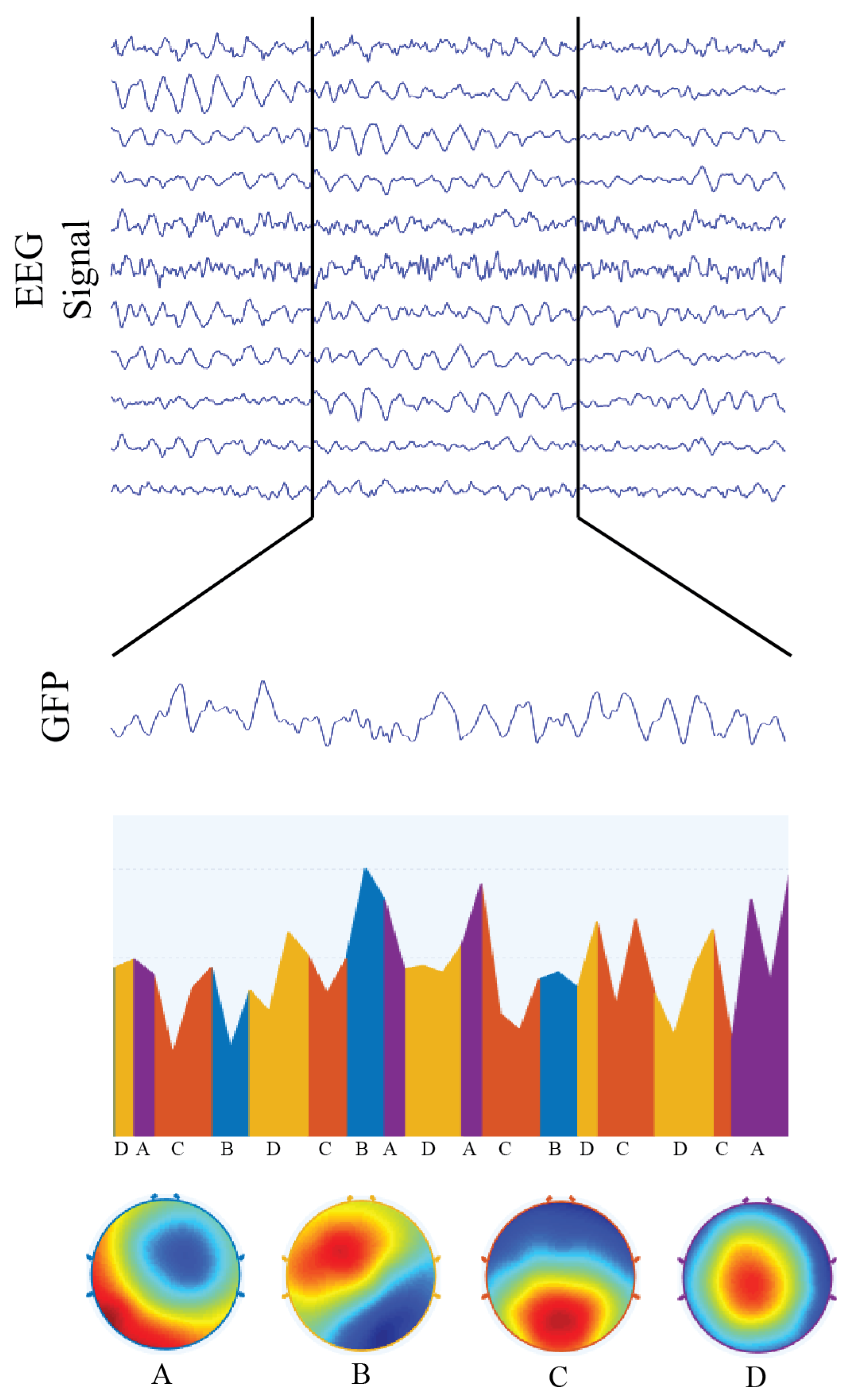
2.4. Microstates & Hubs
3. Discussion
4. Materials and Methods
4.1. EEG Data and Tasks
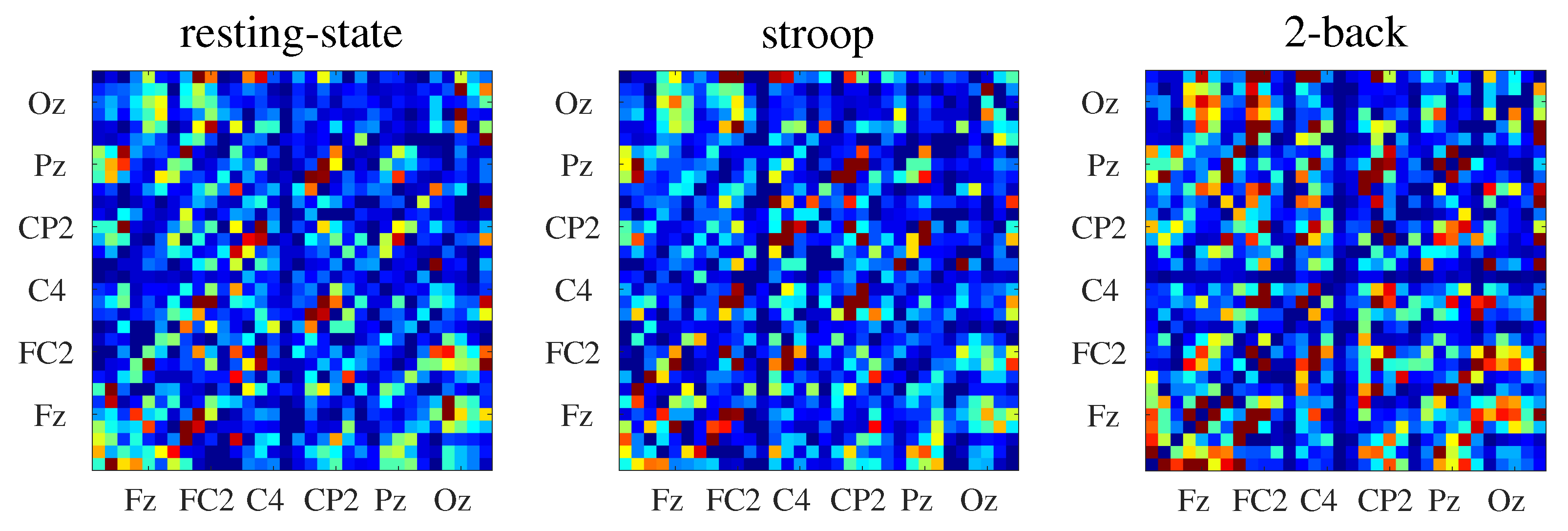
4.2. Normalized Transfer Entropy
4.3. Order Parameters
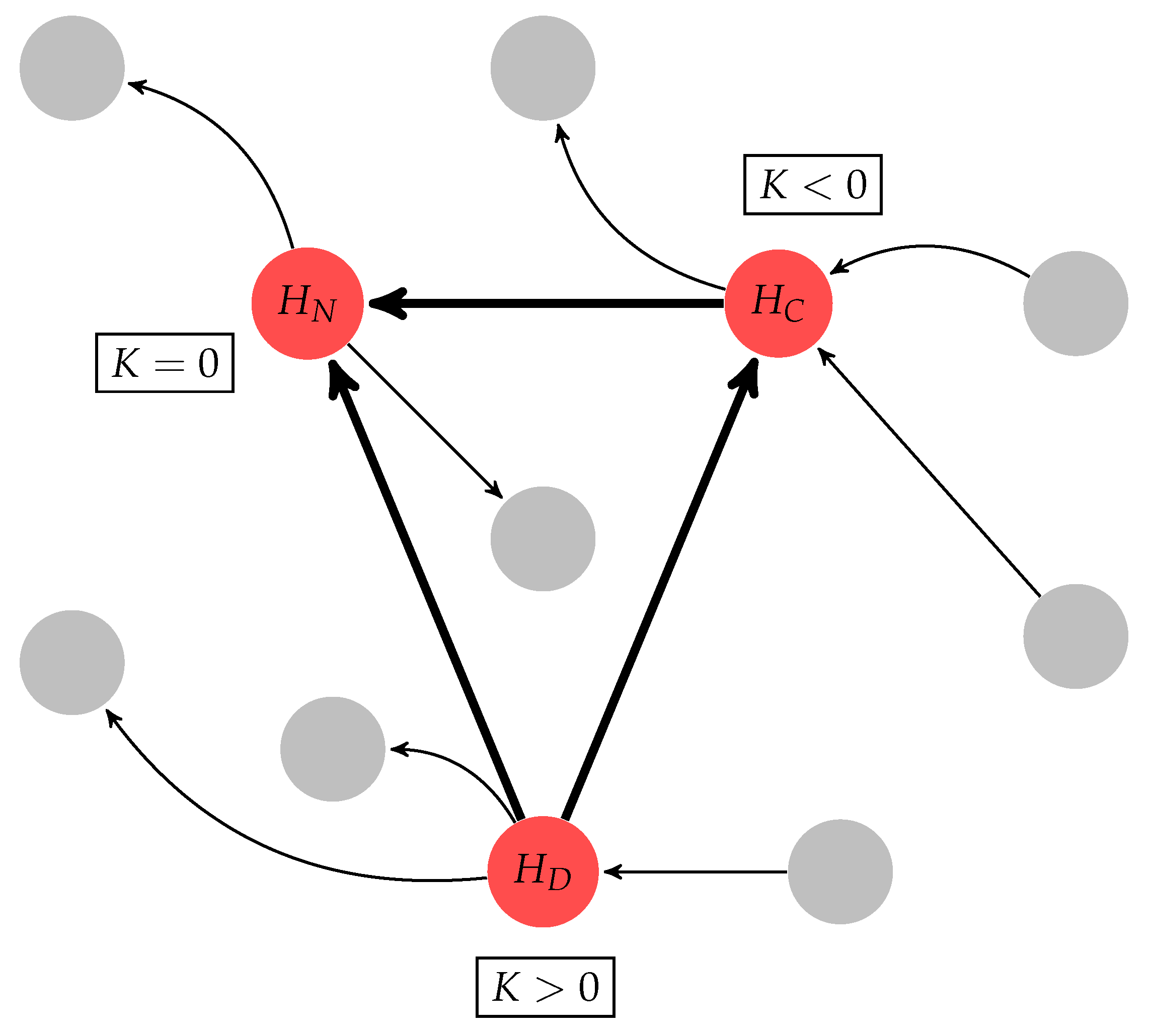
4.4. Divergent, Convergent and Neutral Hubs
4.5. Homogeneity and Heterogeneity Measures
4.6. Brain Microstates and Hubs
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 2-back working memory test | |
| Betweenness centrality | |
| BOLD | Blood-oxygen-level-dependent |
| DMN | Default-mode-network |
| EEG | Electroencephalography |
| fMRI | Functional magnetic resonance imaging |
| Global field potential | |
| GW | Global neuronal workspace |
| Convergent hubs | |
| Divergent hubs | |
| Neutral hubs | |
| Input degree | |
| Normalized transfer entropy | |
| Output degree | |
| p-value of Kruskal–Wallis test | |
| p-value of Friedman test | |
| Resting-state | |
| rsfMRI | Resting-state functional magnetic resonance imaging |
| Stroop color–word test |
References
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O. Networks of the Brain; MIT Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Moutoussis, K.; Zeki, S. Functional segregation and temporal hierarchy of the visual perceptive systems. Proc. R. Soc. B Biol. Sci. 1997, 264, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, M.P.; Sporns, O. Network hubs in the human brain. Trends Cogn. Sci. 2013, 17, 693–696. [Google Scholar] [CrossRef] [PubMed]
- Dehaene, S.; Sergent, C.; Changeux, J. A neuronal network model linking subjective reports and objective physiological data during conscious perception. Proc. Natl. Acad. Sci. USA 2003, 100, 1–6. [Google Scholar] [CrossRef]
- Baar, B.J. Theater of Consciousness. J. Conscious. Stud. 1997, 4, 292–309. [Google Scholar]
- Dehaene, S.; Naccache, L. Towards a cognitive neuroscience of consciousness: Basic evidence and a workspace framework. Cognition 2001, 79, 1–37. [Google Scholar] [CrossRef]
- Bullmore, E.; Bassett, D.S.; Duke, T.; Meyer-Lindenberg, A.; Achard, S. Adaptive reconfiguration of fractal small-world human brain functional networks. Proc. Natl. Acad. Sci. USA 2006, 103, 19518–19523. [Google Scholar]
- Cohen, R.; Erez, K.; Ben-Avraham, D.; Havlin, S. Resilience of the internet to random breakdown. Phys. Rev. Lett. 2000, 85, 4626–4628. [Google Scholar] [CrossRef]
- Gao, J.; Barzel, B.; Barabási, A.L. Universal resilience patterns in complex networks. Nature 2016, 536, 307–312. [Google Scholar] [CrossRef]
- Petersen, S.E.; Posner, M.I. The attention system of the human brain: 20 years after. Annu. Rev. Neurosci. 2012, 35, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Owen, A.M.; McMillan, K.M.; Laird, A.R.; Bullmore, E. N-back working memory paradigm: A meta-analysis of normative functional neuroimaging studies. Hum. Brain Mapp. 2005, 25, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Damoiseaux, J.S.; Rombouts, S.A.R.B.; Barkhof, F.; Scheltens, P.; Stam, C.J.; Smith, S.M.; Beckmann, C.F. Consistent resting-state networks across healthy subjects. Proc. Natl. Acad. Sci. USA 2006, 103, 13848–13853. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, M.; Martin, R.; Andras, P.; Young, M.P. Simulation of robustness against lesions of cortical networks. Eur. J. Neurosci. 2007, 25, 3185–3192. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O. Network attributes for segregation and integration in the human brain. Curr. Opin. Neurobiol. 2013, 23, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.; Damasio, A. Convergence and divergence in a neural architecture for recognition and memory. Trends Neurosci. 2009, 32, 376–382. [Google Scholar] [CrossRef]
- Van Den Heuvel, M.P.; Pol, H.E.H. Exploring the brain network: A review on resting-state fMRI functional connectivity. Eur. Neuropsychopharmacol. 2010, 20, 519–534. [Google Scholar] [CrossRef]
- Fair, D.A.; Dosenbach, N.U.; Church, J.A.; Cohen, A.L.; Brahmbhatt, S.; Miezin, F.M.; Barch, D.M.; Raichle, M.E.; Petersen, S.E.; Schlaggar, B.L. Development of distinct control networks through segregation and integration. Proc. Natl. Acad. Sci. USA 2007, 104, 13507–13512. [Google Scholar] [CrossRef]
- Dehaene, S.; Changeux, J.P. Experimental and theoretical approaches to conscious processing. Neuron 2011, 70, 200–227. [Google Scholar] [CrossRef]
- Dehaene, S.; Kerszberg, M.; Changeux, J.P. A neuronal model of a global workspace in effortful cognitive tasks. Proc. Natl. Acad. Sci. USA 1998, 95, 14529–14534. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A. Timing in cognition and EEG brain dynamics: Discreteness versus continuity. Cogn. Process. 2006, 7, 135–162. [Google Scholar] [CrossRef]
- Grossberg, S. The complementary brain: Unifying brain dynamics and modularity. Trends Cogn. Sci. 2000, 4, 233–246. [Google Scholar] [CrossRef]
- Schroeder, C.E.; Lakatos, P. Low-frequency neuronal oscillations as instruments of sensory selection. Trends Cogn. Sci. 2009, 32, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Britz, J.; Van De Ville, D.; Michel, C.M. BOLD correlates of EEG topography reveal rapid resting-state network dynamics. Neuroimage 2010, 52, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Mantini, D.; Perrucci, M.G.; Del Gratta, C.; Romani, G.L.; Corbetta, M. Electrophysiological signatures of resting state networks in the human brain. Proc. Natl. Acad. Sci. USA 2007, 104, 13170–13175. [Google Scholar] [CrossRef] [PubMed]
- Bressler, S.L. Large-scale cortical networks and cognition. Brain Res. Rev. 1995, 20, 288–304. [Google Scholar] [CrossRef]
- Fuster, J.M. The cognit: A network model of cortical representation. Int. J. Psychophysiol. 2006, 60, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Seeley, W.W.; Crawford, R.K.; Zhou, J.; Miller, B.L.; Greicius, M.D. Neurodegenerative diseases target large-scale human brain networks. Neuron 2009, 62, 42–52. [Google Scholar] [CrossRef]
- Harrison, B.J.; Shaw, M.; Yücel, M.; Purcell, R.; Brewer, W.J.; Strother, S.C.; Egan, G.F.; Olver, J.S.; Nathan, P.J.; Pantelis, C. Functional connectivity during Stroop task performance. Neuroimage 2005, 24, 181–191. [Google Scholar] [CrossRef]
- Shovon, M.H.I.; Nandagopal, D.N.; Vijayalakshmi, R.; Du, J.T.; Cocks, B. Transfer entropy and information flow patterns in functional brain networks during cognitive activity. In International Conference on Neural Information Processing; Springer: Cham, Switzerland, 2014; pp. 1–10. [Google Scholar]
- Schreiber, T. Measuring information transfer. Phys. Rev. Lett. 2000, 85, 461. [Google Scholar] [CrossRef]
- Da Silva, L.M.U.; Baltazar, C.A.; Silva, C.A.; Ribeiro, M.W.; de Aratanha, M.A.A.; Deolindo, C.S.; Rodrigues, A.C.; Machado, B.S. Measures for brain connectivity analysis: Nodes centrality and their invariant patterns. Eur. Phys. J. Spec. Top. 2017, 226, 2235–2245. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, H.G. The neural origins of human event-related potentials. Ann. N. Y. Acad. Sci. 1982, 388, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, D.; Ozaki, H.; Pal, I. EEG alpha map series: Brain micro-states by space-oriented adaptive segmentation. Electroencephalogr. Clin. Neurophysiol. 1987, 67, 271–288. [Google Scholar] [CrossRef]
- Lempel, A.; Ziv, J. On the complexity of finite sequences. IEEE Trans. Inf. Theory 1976, 22, 75–81. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hub Classification | Resting-State | Stroop | 2-Back | ||
|---|---|---|---|---|---|
| 0.2986 | 0.3490 | 0.3160 | 16.8788% | 5.8272% | |
| −0.7293 | −0.2440 | −0.2390 | 66.5433% | 67.2288% | |
| 0.0171 | 0.0210 | 0.0170 | 22.8070% | −0.5848% |
| Hub Classification | Resting-State | Stroop | 2-Back |
|---|---|---|---|
| Network minus core | 39.65% | 40.18% | 39.36% |
| Network minus | 27.13% | 23.86% | 27.15% |
| Network minus | 4.53% | 5.97% | 6.07% |
| Network minus | 6.04% | 9.60% | 4.21% |
| Hub Classification | Resting-State | Stroop | 2-Back |
|---|---|---|---|
| 64.61% | 65.82% | 72.91% | |
| 13.47% | 14.68% | 14.40% | |
| 21.92% | 19.51% | 12.69% |
| Connective Core | Hub Change | |||
|---|---|---|---|---|
| Invariant structure | 32.94% | 34.03% | 35.64% | |
| 01.14% | 02.31% | 02.27% | ||
| 05.84% | 04.16% | 00.96% | ||
| Total preserved | 39.92% | 42.18% | 38.87% | |
| Variant structure | 02.61% | 01.21% | 02.81% | |
| 00.00% | 01.30% | 00.00% | ||
| 26.01% | 25.92% | 28.39% | ||
| 01.30% | 03.93% | 01.65% | ||
| 01.14% | 00.00% | 01.40% | ||
| 10.57% | 06.99% | 09.66% | ||
| 08.82% | 10.52% | 05.86% | ||
| 01.40% | 03.32% | 03.34% | ||
| 08.24% | 06.31% | 08.02% | ||
| Total | 100.00% | 100.00% | 100.00% | |
| Correlation | S | ||||
|---|---|---|---|---|---|
| −0.3323 | −0.3576 | −0.3022 | −7.6113% | 9.0586% | |
| −0.2135 | −0.1747 | −0.2026 | 18.1746% | 5.1016% | |
| −0.0987 | −0.1281 | −0.2013 | −29.7501% | −103.8972% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baltazar, C.A.; Guinle, M.I.B.; Caron, C.J.; Amaro Jr., E.; Machado, B.S. Connective Core Structures in Cognitive Networks: The Role of Hubs. Entropy 2019, 21, 961. https://doi.org/10.3390/e21100961
Baltazar CA, Guinle MIB, Caron CJ, Amaro Jr. E, Machado BS. Connective Core Structures in Cognitive Networks: The Role of Hubs. Entropy. 2019; 21(10):961. https://doi.org/10.3390/e21100961
Chicago/Turabian StyleBaltazar, Carlos Arruda, Maria Isabel Barros Guinle, Cora Jirschik Caron, Edson Amaro Jr., and Birajara Soares Machado. 2019. "Connective Core Structures in Cognitive Networks: The Role of Hubs" Entropy 21, no. 10: 961. https://doi.org/10.3390/e21100961
APA StyleBaltazar, C. A., Guinle, M. I. B., Caron, C. J., Amaro Jr., E., & Machado, B. S. (2019). Connective Core Structures in Cognitive Networks: The Role of Hubs. Entropy, 21(10), 961. https://doi.org/10.3390/e21100961