Paced Breathing Increases the Redundancy of Cardiorespiratory Control in Healthy Individuals and Chronic Heart Failure Patients

 ,
, 
 ,
,  , ,
, ,
Abstract
1. Introduction
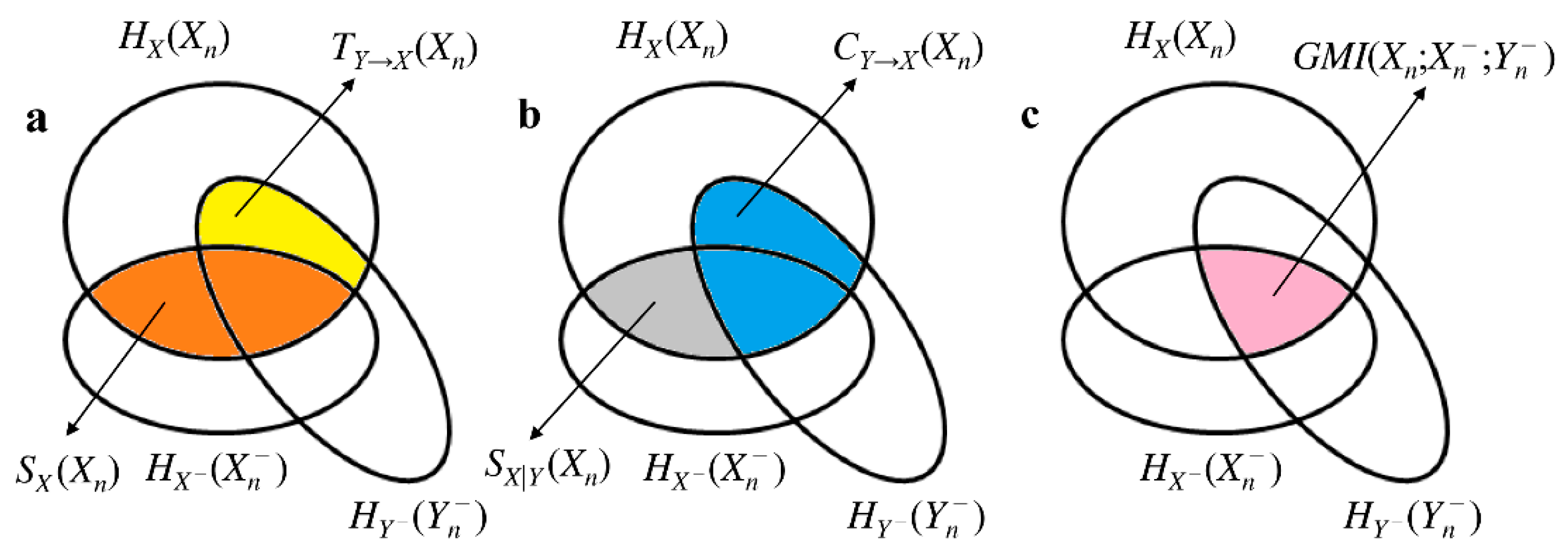
2. Quantifying Redundancy/Synergy in Bivariate Stochastic Systems
2.1. Notation and Preliminaries
2.2. Basic Information-Theoretic Quantities Contributing to Interaction Self-Information in Bivariate Stochastic Systems
2.3. Generalized MI and its Link with Interaction Self-Information in Bivariate Stochastic Systems
2.4. Separating the Contributions of Redundancy and Synergy in Bivariate Interaction Self-Information
2.5. Estimation of the Bivariate Interaction Self-Information
3. Experimental Protocols
3.1. Paced Breathing in Healthy Young Subjects
3.2. Paced Breathing in CHF Patients
3.3. Slow Breathing in CHF Patients
3.4. Extraction of Beat-to-Beat HP Variability, R Series and Breathing Rate
3.5. Computing Interaction Self-Information from Beat-to-Beat HP Variability and R Series
3.6. Computation of Traditional Markers Describing Cardiorespiratory Interactions
3.7. Statistical Analysis
4. Results
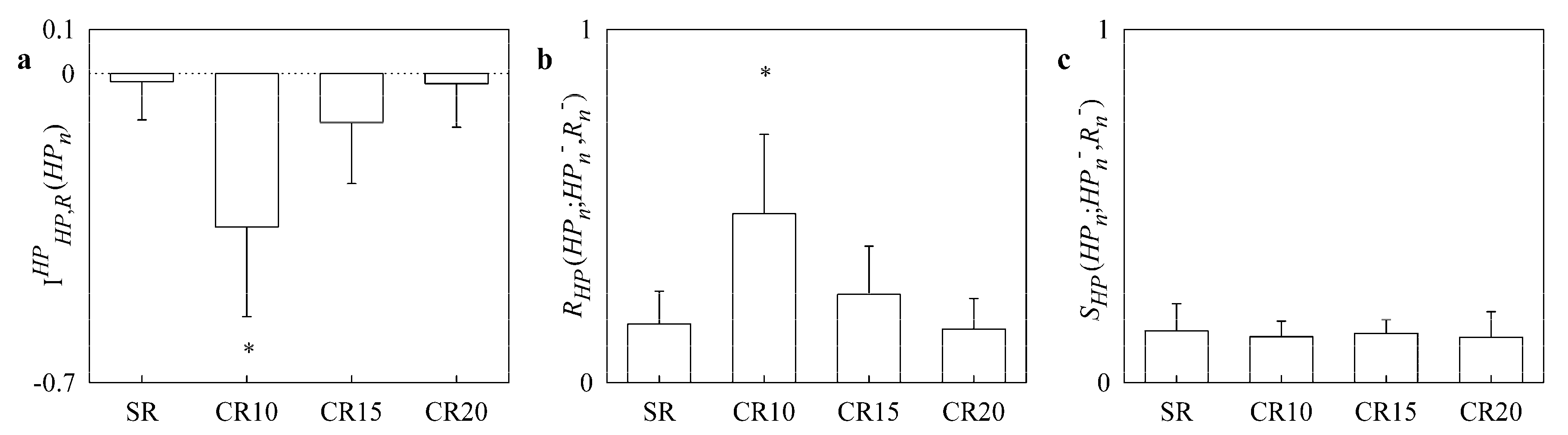
4.1. Results of the Paced Breathing Protocol in Healthy Young Subjects
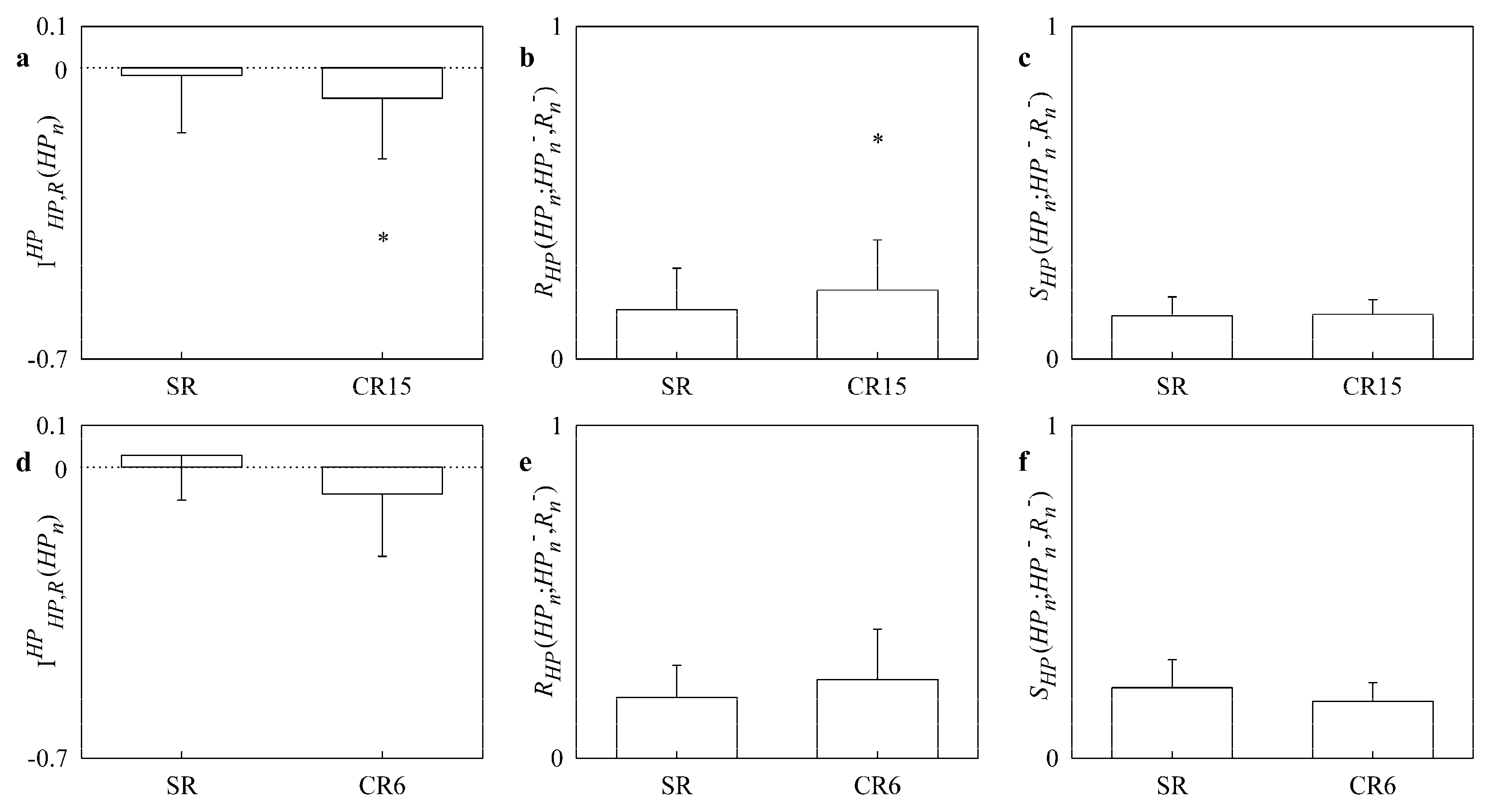
4.2. Results of the Paced and Slow Breathing Protocols in CHF Patients
5. Discussion
- applly two approaches framed in the field of information dynamics for the assessment of redundancy/synergy in bivariate stochastic systems;
- the two approaches were made operational in Gaussian bivariate stochastic systems describing the interactions between heart and respiration;
- in healthy young subjects pacing respiration at a frequency slower than the spontaneous one increased redundancy of the cardiorespiratory control;
- this effect was visible in CHF population as well even though the increase of redundancy might be limited by factors related to the efficiency of the cardiac pump;
- redundancy/synergy parameters provided information complementary to the respiratory sinus arrhythmia and strength of the HP-R linear coupling at the respiratory rate.
5.1. Assessing Redundancy/Synergy in Bivariate Stochastic Systems
5.2. On the Relevance of Assessing Redundancy/Synergy of the Cardiorespiratory Interactions
5.3. Paced Breathing Increases Redundancy of Cardiorespiratory Control in Healthy Young Subjects and CHF Patients
5.4. Correlation of Redundancy/Synergy Markers with Traditional Cardiorespiratory Indexes
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Prokopenko, M.; Boschetti, F.; Ryan, A.J. An information-theoretic primer on complexity, self-Organization, and emergence. Complexity 2008, 15, 11. [Google Scholar] [CrossRef]
- Angelini, L.; de Tommaso, M.; Marinazzo, D.; Nitti, L.; Pellicoro, M.; Stramaglia, S. Redundant variables and Granger causality. Phys. Rev. E 2010, 81, 037201. [Google Scholar] [CrossRef] [PubMed]
- Stramaglia, S.; Wu, G.-R.; Pellicoro, M.; Marinazzo, D. Expanding the transfer entropy to identify information circuits in complex systems. Phys. Rev. E 2012, 86, 066211. [Google Scholar] [CrossRef] [PubMed]
- Porta, A.; Bari, V.; De Maria, B.; Perseguini, N.M.; Milan, J.; Rehder-Santos, P.; Minatel, V.; Takahashi, A.C.M.; Catai, A.M. Assessing the evolution of redundancy/synergy of spontaneous variability regulation with age. Physiol. Meas. 2017, 38, 940–958. [Google Scholar] [CrossRef] [PubMed]
- Porta, A.; Bari, V.; De Maria, B.; Takahashi, A.C.M.; Guzzetti, S.; Colombo, R.; Catai, A.M.; Raimondi, F.; Faes, L. Quantifying net synergy/redundancy of spontaneous variability regulation via predictability and transfer entropy decomposition frameworks. IEEE Trans. Biomed. Eng. 2017, 64, 2628–2638. [Google Scholar]
- Faes, L.; Porta, A.; Nollo, G. Redundant and synergistic information transfer in cardiovascular and cardiorespiratory variability. In Proceedings of the 37th Annual International Conference of the IEEE EMBS, Milan, Italy, 25–29 August 2015. [Google Scholar]
- Marinazzo, D.; Gosseries, O.; Boly, M.; Ledoux, D.; Rosanova, M.; Massimini, M.; Noirhomme, Q.; Laureys, S. Directed information transfer in scalp electroencephalographic recordings: Insights on disorders of consciousness. Clin. EEG Neurosci. 2014, 45, 33–39. [Google Scholar] [CrossRef]
- Stramaglia, S.; Angelini, L.; Wu, G.; Cortes, J.M.; Faes, L.; Marinazzo, D. Synergetic and Redundant Information Flow Detected by Unnormalized Granger Causality: Application to Resting State fMRI. IEEE Trans. Biomed. Eng. 2016, 63, 2518–2524. [Google Scholar]
- Barrett, A.B. Exploration of synergistic and redundant information sharing in static and dynamical Gaussian systems. Phys. Rev. E 2015, 91, 052802. [Google Scholar] [CrossRef]
- Harder, M.; Salge, C.; Polani, D. Bivariate measure of redundant information. Phys. Rev. E 2013, 87, 012130. [Google Scholar] [CrossRef]
- Williams, P.L.; Beer, R.D. Nonnegative decomposition of multivariate information. arXiv, 2010; arXiv:1004.2515v1. [Google Scholar]
- Camino-Pontes, B.; Diez, I.; Jimenez-Marin, A.; Rasero, J.; Erramuzpe, A.; Bonifazi, P.; Stramaglia, S.; Swinnen, S.; Cortes, J.M. Interaction information along lifespan of the resting brain dynamics reveals a major redundant role of the default mode network. Entropy 2018, 20, 742. [Google Scholar] [CrossRef]
- Schneidman, E.; Bialek, W.; Berry, M. Synergy, redundancy, and independence in population codes. J. Neurosci. 2003, 23, 11539–11553. [Google Scholar] [CrossRef] [PubMed]
- Porta, A.; Faes, L.; Marchi, A.; Bari, V.; De Maria, B.; Guzzetti, S.; Colombo, R.; Raimondi, F. Disentangling cardiovascular control mechanisms during head-down tilt via joint transfer entropy and self-entropy decompositions. Front. Physiol. 2015, 6, 301. [Google Scholar] [CrossRef] [PubMed]
- Faes, L.; Porta, A.; Nollo, G.; Javorka, M. Information decomposition multivariate systems: Definitions, implementation and application to cardiovascular networks. Entropy 2017, 19, 5. [Google Scholar] [CrossRef]
- Eckberg, D.L. The human respiratory gate J. Physiol. 2003, 548, 339–352. [Google Scholar] [CrossRef]
- Hirsch, J.A.; Bishop, B. Respiratory sinus arrhythmia in humans: How breathing pattern modulates heart rate. Am. J. Physiol. 1981, 241, H620–H629. [Google Scholar] [CrossRef]
- Widjaja, D.; Montalto, A.; Vlemincx, E.; Marinazzo, D.; Van Huffel, S.; Faes, L. Cardiorespiratory information dynamics during mental arithmetic and sustained attention. PLoS ONE 2015, 10, e0129112. [Google Scholar] [CrossRef]
- Iatsenko, D.; Bernjak, A.; Stankovski, T.; Shiogai, Y.; Owen-Lynch, P.J.; Clarkson, P.B.M.; McClintock, P.V.E.; Stefanovska, A. Evolution of cardiorespiratory interactions with age. Phil. Trans. R. Soc. A 2013, 371, 20110622. [Google Scholar] [CrossRef]
- Saul, J.P.; Berger, R.D.; Chen, M.H.; Cohen, R.J. Transfer function analysis of autonomic regulation II. Respiratory sinus arrhythmia. Am. J. Physiol. 1989, 256, H153–H161. [Google Scholar] [CrossRef]
- Schafer, C.; Rosenblum, M.G.; Kurths, J.; Abel, H.-H. Heartbeat synchronized with ventilation. Nature 1998, 392, 239–240. [Google Scholar]
- Bracic Lotric, M.; Stefanovska, A. Synchronization and modulation in the human cardiorespiratory system. Physica A 2000, 283, 451–461. [Google Scholar] [CrossRef]
- Bartsch, R.; Kartelhardt, J.W.; Penzel, T.; Havlin, S. Experimental evidence for phase synchronization transitions in the human cardiorespiratory system. Phys. Rev. Lett. 2007, 98, 054102. [Google Scholar] [CrossRef]
- Riedl, M.; Muller, A.; Kraemer, J.F.; Penzel, T.; Kurths, J.; Wessel, N. Cardio-respiratory coordination increases during sleep apnea. PLoS ONE 2014, 9, e93866. [Google Scholar] [CrossRef] [PubMed]
- Penzel, T.; Kantelhardt, J.W.; Bartsch, R.P.; Riedl, M.; Kramer, J.; Wessel, N.; Garcia, C.; Glos, M.; Fietze, I.; Schobel, C. Modulations of heart rate, ECG, and cardio-respiratory coupling observed in polysomnography. Front. Physiol. 2016, 7, 460. [Google Scholar] [CrossRef] [PubMed]
- Mazzucco, C.E.; Marchi, A.; Bari, V.; De Maria, B.; Guzzetti, S.; Raimondi, F.; Catena, E.; Ottolina, D.; Amadio, C.; Cravero, S.; Fossali, T.; Colombo, R.; Porta, A. Mechanical ventilatory modes and cardioventilatory phase synchronization in acute respiratory failure patients. Physiol. Meas. 2017, 38, 895–911. [Google Scholar] [CrossRef] [PubMed]
- Porta, A.; Guzzetti, S.; Montano, N.; Pagani, M.; Somers, V.; Malliani, A.; Baselli, G.; Cerutti, S. Information domain analysis of cardiovascular variability signals: Evaluation of regularity, synchronisation and co-ordination. Med. Biol. Eng. Comput. 2000, 38, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Pinna, G.D.; Maestri, R.; La Rovere, M.T.; Gobbi, E.; Fanfulla, F. Effect of paced breathing on ventilatory and cardiovascular variability parameters during short-term investigations of autonomic function. Am. J. Physiol. 2006, 290, H424–H433. [Google Scholar] [CrossRef] [PubMed]
- Faes, L.; Porta, A.; Nollo, G. Information decomposition in bivariate systems: Theory and application to cardiorespiratory dynamics. Entropy 2015, 17, 277–303. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell System Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Lizier, J.T.; Prokopenko, M.; Zomaya, A.Y. Local measures of information storage in complex distributed computation. Inform. Sci. 2012, 208, 39–54. [Google Scholar] [CrossRef]
- Wibral, M.; Lizier, J.T.; Vogler, S.; Priesemann, V.; Galuske, R. Local active information storage as a tool to understand distributed neural information processing. Front. Neuroinf. 2014, 8, 1. [Google Scholar] [CrossRef]
- Porta, A.; Faes, L.; Nollo, G.; Bari, V.; Marchi, A.; De Maria, B.; Takahashi, A.C.M.; Catai, A.M. Conditional self-entropy and conditional joint transfer entropy in heart period variability during graded postural challenge. PLoS ONE 2015, 10, e0132851. [Google Scholar] [CrossRef]
- Porta, A.; Baselli, G.; Lombardi, F.; Montano, N.; Malliani, A.; Cerutti, S. Conditional entropy approach for the evaluation of the coupling strength. Biol. Cybern. 1999, 81, 119–129. [Google Scholar] [CrossRef]
- Schreiber, T. Measuring information transfer. Phys. Rev. Lett. 2000, 85, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Barnett, L.; Barrett, A.B.; Seth, A.K. Granger causality and transfer entropy are equivalent for Gaussian variables. Phys. Rev. Lett. 2009, 103, 238701. [Google Scholar] [CrossRef]
- Kay, S.M.; Marple, S.L. Spectrum analysis: A modern perspective. Proc. IEEE 1981, 69, 1380–1418. [Google Scholar] [CrossRef]
- Eckberg, D.L. Temporal response patterns of the human sinus node to brief carotid baroreceptor stimuli. J. Physiol. 1976, 258, 769–782. [Google Scholar] [CrossRef]
- Porta, A.; Bassani, T.; Bari, V.; Pinna, G.D.; Maestri, R.; Guzzetti, S. Accounting for respiration is necessary to reliably infer Granger causality from cardiovascular variability series. IEEE Trans. Biomed. Eng. 2012, 59, 832–841. [Google Scholar] [CrossRef]
- Soderstrom, T.; Stoica, P. System Identification; Prentice Hall: Englewood Cliffs, NJ, USA, 1988. [Google Scholar]
- Baselli, G.; Porta, A.; Rimoldi, O.; Pagani, M.; Cerutti, S. Spectral decomposition in multichannel recordings based on multivariate parametric identification. IEEE Trans. Biomed. Eng. 1997, 44, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. A new look at the statistical novel identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Porta, A.; Castiglioni, P.; Di Rienzo, M.; Bassani, T.; Bari, V.; Faes, L.; Nollo, G.; Cividjan, A.; Quintin, L. Cardiovascular control and time domain Granger causality: Insights from selective autonomic blockade. Phil. Trans. R. Soc. A 2013, 371, 20120161. [Google Scholar] [CrossRef] [PubMed]
- Porta, A.; Faes, L.; Bari, V.; Marchi, A.; Bassani, T.; Nollo, G.; Perseguini, N.M.; Milan, J.; Minatel, V.; Borghi-Silva, A.; et al. Effect of age on complexity and causality of the cardiovascular control: Comparison between model-based and model-free approaches. PLoS ONE 2014, 9, e89463. [Google Scholar] [CrossRef]
- Malik, M. Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology. Heart rate variability-Standards of measurement, physiological interpretation and clinical use. Circulation 1996, 93, 1043–1065. [Google Scholar]
- Russo, M.A.; Santarelli, D.M.; O’Rourke, D. The physiological effects of slow breathing in the healthy human. Breathe 2017, 13, 298–309. [Google Scholar]
- Toska, K.; Eriksen, M. Respiration-synchronous fluctuations in stroke volume, heart rate and arterial pressure in humans. J. Physiol. 1993, 472, 501–512. [Google Scholar] [CrossRef] [PubMed]
- De Boer, R.W.; Karemaker, J.M.; Strackee, J. Hemodynamic fluctuations and baroreflex sensitivity in humans: A beat-to-beat model. Am. J. Physiol. 1987, 253, H680–H689. [Google Scholar]
- Eckberg, D.L.; Karemaker, J.M. Point:Counterpoint: Respiratory sinus arrhythmia is due to a central mechanism vs. respiratory sinus arrhythmia is due to the baroreflex mechanism. J. Appl. Physiol. 2009, 106, 1740–1744. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.A.; Eckberg, D.L. Fundamental relations between short-term RR interval and arterial pressure oscillations in humans. Circulation 1996, 93, 1527–1532. [Google Scholar] [CrossRef]
- Porta, A.; Catai, A.M.; Takahashi, A.C.M.; Magagnin, V.; Bassani, T.; Tobaldini, E.; van de Borne, P.; Montano, N. Causal relationships between heart period and systolic arterial pressure during graded head-up tilt. Am. J. Physiol. 2011, 300, R378–R386. [Google Scholar] [CrossRef]
- Spyer, K.M. Central nervous mechanisms responsible for cardio-respiratory homeostasis. Adv. Exp. Med. Biol. 1995, 381, 73–79. [Google Scholar]
- Crystal, G.J.; Salem, M.R. The Bainbridge and the “reverse” Bainbridge reflexes: History, physiology, and clinical relevance. Anesth. Analg. 2012, 114, 520–532. [Google Scholar] [CrossRef]
- Taha, B.H.; Simon, P.M.; Dempsey, J.A.; Skatrud, J.B.; Iber, C. Respiratory sinus arrhythmia in humans: An obligatory role for vagal feedback from the lungs. J. Appl. Physiol. 1995, 78, 638–645. [Google Scholar] [CrossRef]
- Seals, D.R.; Suwarno, N.O.; Dempsey, J.A. Influence of lung volume on sympathetic nerve discharge in normal subjects. Circ. Res. 1990, 67, 130–141. [Google Scholar] [CrossRef]
- Sundlof, G.; Wallin, B.G. Human muscle nerve sympathetic activity at rest. Relationship to blood pressure and age. J. Physiol. 1978, 274, 621–637. [Google Scholar] [CrossRef]
- Marchi, A.; Bari, V.; De Maria, B.; Esler, M.; Lambert, E.; Baumert, M.; Porta, A. Simultaneous characterization of sympathetic and cardiac arms of the baroreflex through sequence techniques during incremental head-up tilt. Front. Physiol. 2016, 7, 438. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, L.; Gabutti, A.; Porta, C.; Spicuzza, L. Slow breathing reduces chemoreflex response to hypoxia and hypercapnia, and increases baroreflex sensitivity. J. Hypertens. 2001, 19, 2221–2229. [Google Scholar] [CrossRef]
- Radaelli, A.; Raco, R.; Perfetti, P.; Viola, A.; Azzellino, A.; Signorini, M.G.; Ferrari, A.U. Effects of slow, controlled breathing on baroreceptor control of heart rate and blood pressure in healthy men. J. Hypertens. 2004, 22, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.E.; Beightol, L.A.; Kob, J.; Eckberg, D.L. Important influence of respiration on human RR interval power spectra is largely ignored. J. Appl. Physiol. 1993, 75, 2310–2317. [Google Scholar] [CrossRef] [PubMed]
- Angelone, A.; Coulter, N.A. Respiratory sinus arrhythmia: A frequency dependent phenomenon. J. Appl. Physiol. 1964, 19, 479–482. [Google Scholar] [CrossRef]
- Lopes, T.C.; Beda, A.; Granja-Filho, P.C.N.; Jandre, F.C.; Giannella-Neto, A. Cardio-respiratory interactions and relocation of heartbeats within the respiratory cycle during spontaneous and paced breathing. Physiol. Meas. 2011, 32, 1389–1401. [Google Scholar] [CrossRef]
- Porta, A.; Baselli, G.; Lombardi, F.; Cerutti, S.; Antolini, R.; Del Greco, M.; Ravelli, F.; Nollo, G. Performance assessment of standard algorithms for dynamic RT interval measurement: Comparison between RTapex and RTend approach. Med. Biol. Eng. Comput. 1998, 36, 35–42. [Google Scholar] [CrossRef]
- Cohen, M.A.; Taylor, J.A. Short-term cardiovascular oscillations in man: Measuring and modeling the physiologies. J. Physiol. 2002, 542, 669–683. [Google Scholar] [CrossRef]
- Baselli, G.; Cerutti, S.; Badilini, F.; Biancardi, L.; Porta, A.; Pagani, M.; Lombardi, F.; Rimoldi, O.; Furlan, R.; Malliani, A. Model for the assessment of heart period and arterial pressure variability interactions and of respiration influences. Med. Biol. Eng. Comput. 1994, 32, 143–152. [Google Scholar] [CrossRef]





{kind=link}
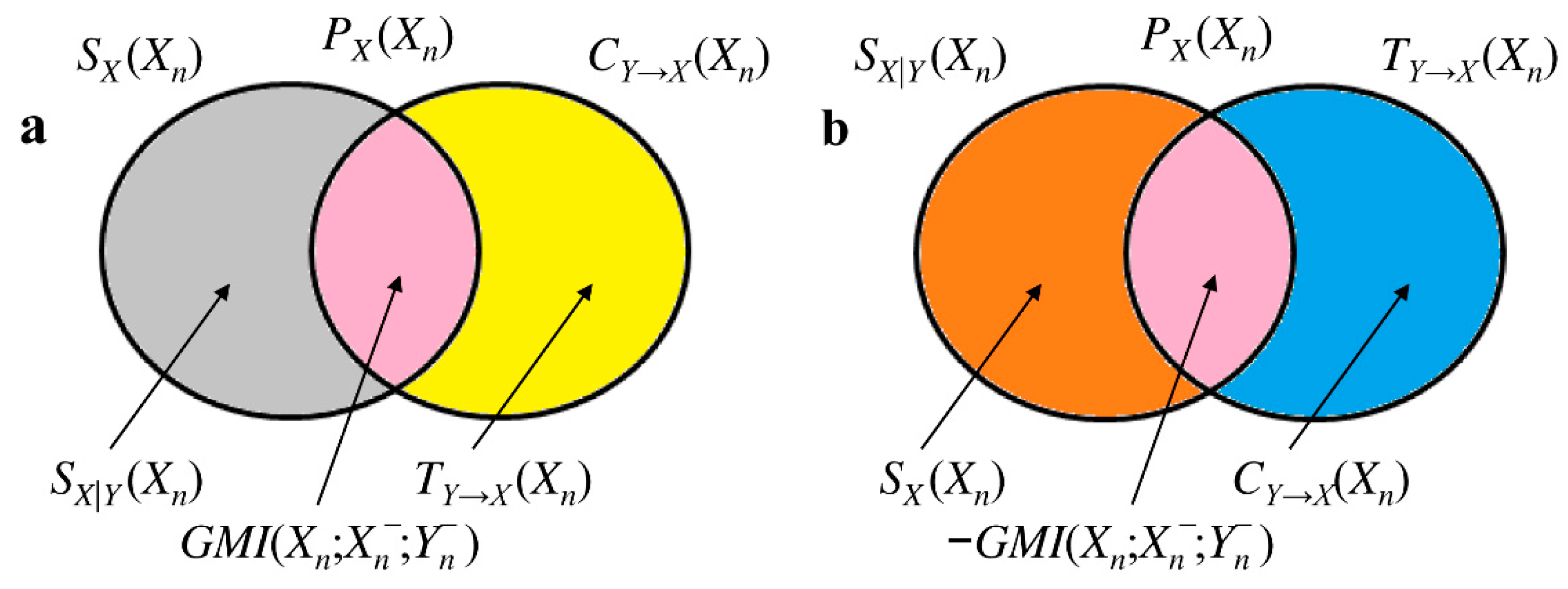{kind=link}
{kind=link}
{kind=link}
| SR | CR10 | CR15 | CR20 | |
|---|---|---|---|---|
| µHP [ms] | 1009.86 ± 167.92 | 989.33 ± 156.82 | 1022.53 ± 161.94 | 1027.95 ± 162.37 |
| σ2HP [ms2] | 3368.30 ± 2622.13 | 4784.17 ± 3355.86 * | 3704.77 ± 3090.59 | 2812.66 ± 2158.02 |
| fR [Hz] | 0.260 ± 0.031 | 0.210 ± 0.049 * | 0.250 ± 0.026 | 0.310 ± 0.024 * |
| HFHP [ms2] | 1405.41 ± 1555.21 | 2856.10 ± 2969.00 * | 1891.23 ± 1906.98 | 1140.68 ± 1205.75 |
| K2HP-R(HF) | 0.85 ± 0.11 | 0.93 ± 0.08 * | 0.96 ± 0.04 * | 0.92 ± 0.05 * |
| Marker | HFHP | K2HP-R(HF) | |
|---|---|---|---|
| −0.438; 6.04 × 10−2 # | −0.142; 5.61 × 10−1 | ||
| SR | 0.556; 1.34 × 10−2 * | 0.450; 5.32 × 10−2 # | |
| 0.191; 4.34 × 10−1 | 0.395; 9.41 × 10−2 | ||
| −0.099; 6.86 × 10−1 | −0.415; 7.72 × 10−2 | ||
| CR10 | 0.041; 8.68 × 10−1 | 0.453; 5.15 × 10−2 # | |
| −0.243; 3.16 × 10−1 | 0.398; 9.17 × 10−2 | ||
| −0.556; 1.35 × 10−2 * | −0.431; 6.52 × 10−2 # | ||
| CR15 | 0.452; 5.20 × 10−2 # | 0.584; 8.69 × 10−3 * | |
| −0.339; 9.02 × 10−2 | 0.497; 3.04 × 10−2 * | ||
| −0.521; 2.23 × 10−2 * | 0.213; 3.81 × 10−1 | ||
| CR20 | 0.328; 1.70 × 10−1 | 0.064; 7.93 × 10−1 | |
| −0.315; 1.89 × 10−1 | 0.365; 1.25 × 10−1 |
| SR | CR15 | |
|---|---|---|
| µHP [ms] | 934.73 ± 121.12 | 931.85 ± 125.90 |
| σ2HP [ms2] | 710.46 ± 762.30 | 626.69 ± 538.63 |
| fR [Hz] | 0.289 ± 0.053 | 0.239 ± 0.024 * |
| HFHP [ms2] | 162.74 ± 204.04 | 203.38 ± 162.52 |
| K2HP-R (HF) | 0.89 ± 0.09 | 0.97 ± 0.03 * |
| SR | CR6 | |
|---|---|---|
| µHP [ms] | 816.36 ± 169.53 | 818.83 ± 155.94 |
| σ2HP [ms2] | 184.44 ± 179.38 | 339.90 ± 415.33 |
| fR [Hz] | 0.292 ± 0.065 | 0.123 ± 0.018 * |
| HFHP [ms2] | 24.89 ± 19.59 | 105.71 ± 168.31 |
| K2HP-R (HF) | 0.88 ± 0.14 | 0.94 ± 0.07 |
| Marker | HFHP | K2HP-R(HF) | |
|---|---|---|---|
| −0.454; 4.42 × 10−2 * | −0.012; 9.58 × 10−1 | ||
| SR | 0.484; 3.06 × 10−2 * | 0.245; 2.97 × 10−1 | |
| −0.047; 8.45 × 10−1 | 0.516; 1.98 × 10−2 * | ||
| −0.345; 1.37 × 10−1 | −0.401; 7.95 × 10−2 | ||
| CR15 | 0.348; 1.33 × 10−1 | 0.518; 1.93 × 10−2 * | |
| 0.051; 8.32 × 10−1 | 0.441; 5.16 × 10−2 # |
| Marker | HFHP | K2HP-R(HF) | |
|---|---|---|---|
| −0.212; 5.83 × 10−1 | 0.223; 5.64 × 10−1 | ||
| SR | 0.306; 4.23 × 10−1 | 0.339; 2.97 × 10−1 | |
| 0.074; 8.49 × 10−1 | 0.740; 2.271 × 10−2 * | ||
| −0.258; 5.03 × 10−1 | −0.275; 4.74 × 10−1 | ||
| CR6 | 0.303; 4.29 × 10−1 | 0.458; 2.15 × 10−1 | |
| 0.130; 7.38 × 10−1 | 0.504; 1.66 × 10−1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porta, A.; Maestri, R.; Bari, V.; De Maria, B.; Cairo, B.; Vaini, E.; La Rovere, M.T.; Pinna, G.D. Paced Breathing Increases the Redundancy of Cardiorespiratory Control in Healthy Individuals and Chronic Heart Failure Patients. Entropy 2018, 20, 949. https://doi.org/10.3390/e20120949
Porta A, Maestri R, Bari V, De Maria B, Cairo B, Vaini E, La Rovere MT, Pinna GD. Paced Breathing Increases the Redundancy of Cardiorespiratory Control in Healthy Individuals and Chronic Heart Failure Patients. Entropy. 2018; 20(12):949. https://doi.org/10.3390/e20120949
Chicago/Turabian StylePorta, Alberto, Roberto Maestri, Vlasta Bari, Beatrice De Maria, Beatrice Cairo, Emanuele Vaini, Maria Teresa La Rovere, and Gian Domenico Pinna. 2018. "Paced Breathing Increases the Redundancy of Cardiorespiratory Control in Healthy Individuals and Chronic Heart Failure Patients" Entropy 20, no. 12: 949. https://doi.org/10.3390/e20120949
APA StylePorta, A., Maestri, R., Bari, V., De Maria, B., Cairo, B., Vaini, E., La Rovere, M. T., & Pinna, G. D. (2018). Paced Breathing Increases the Redundancy of Cardiorespiratory Control in Healthy Individuals and Chronic Heart Failure Patients. Entropy, 20(12), 949. https://doi.org/10.3390/e20120949