Information Theoretical Study of Cross-Talk Mediated Signal Transduction in MAPK Pathways


Abstract
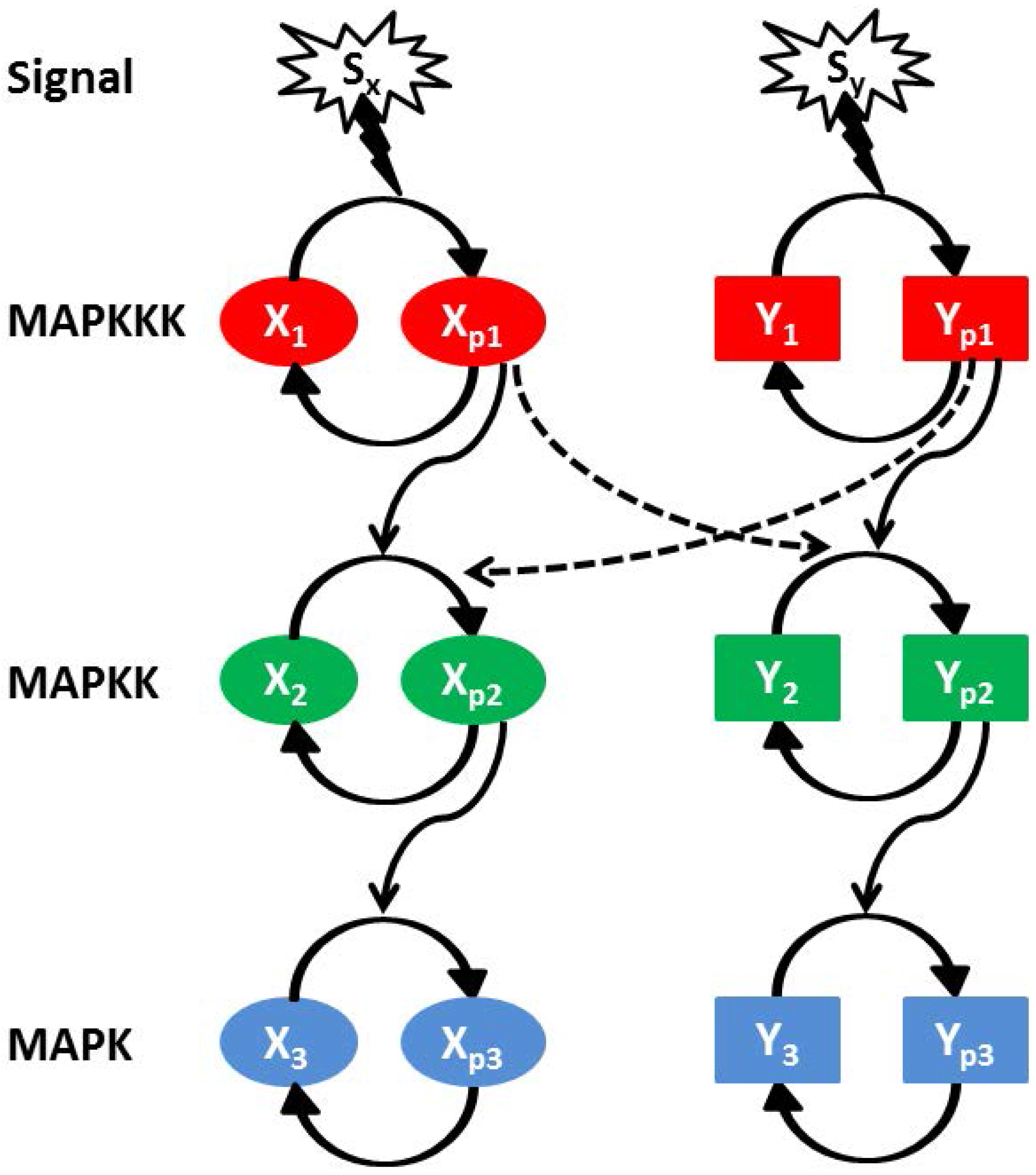
1. Introduction
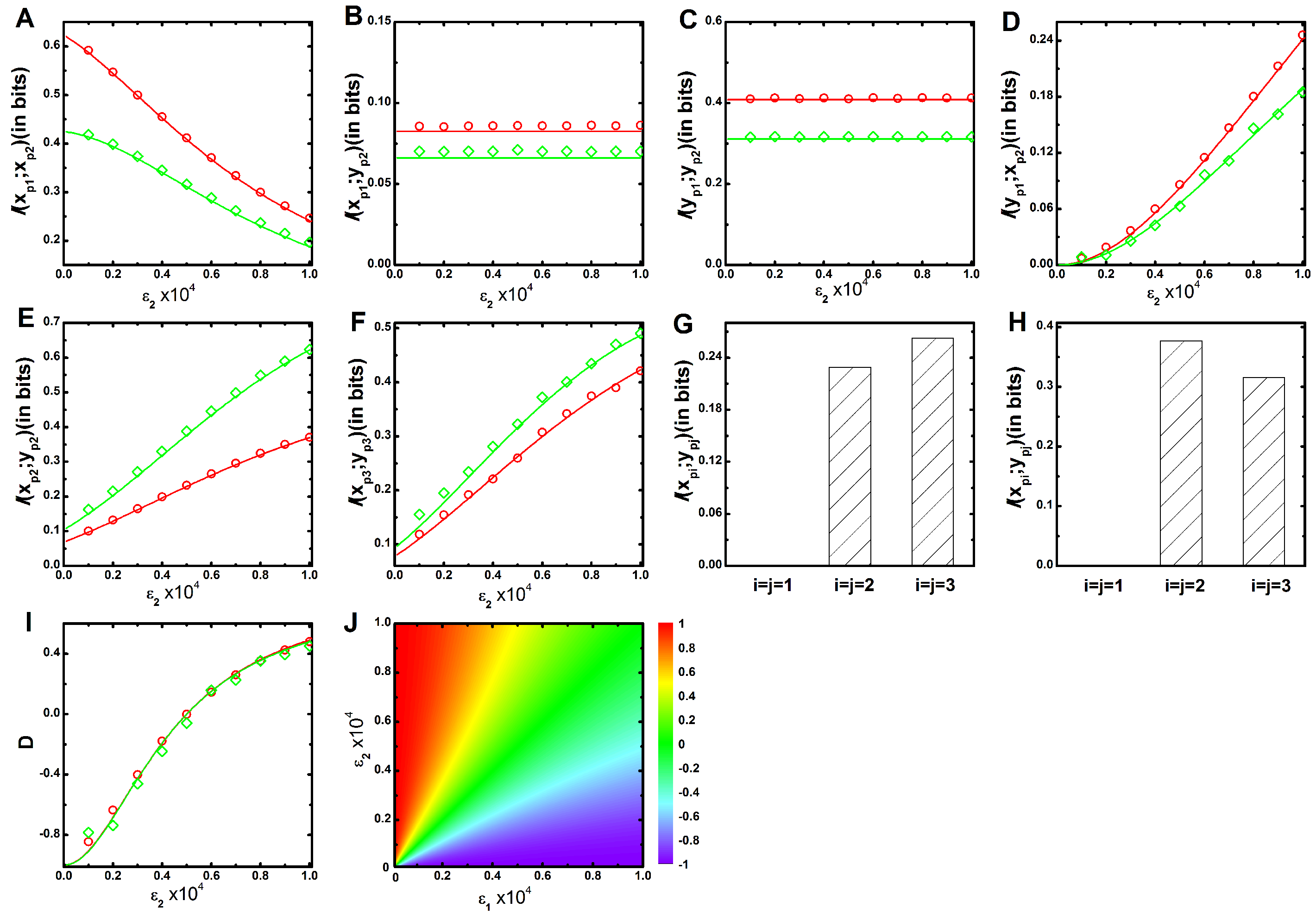
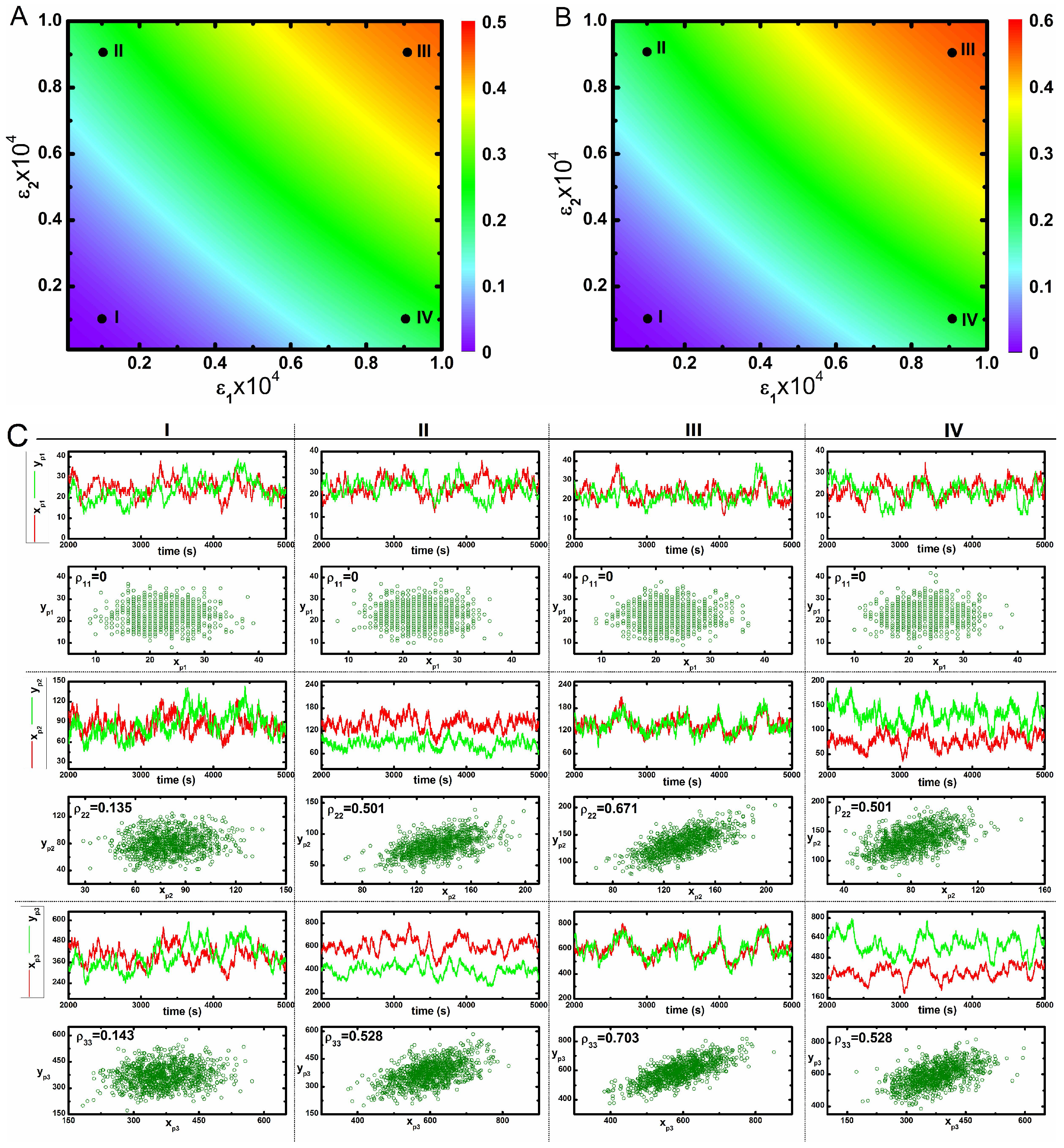
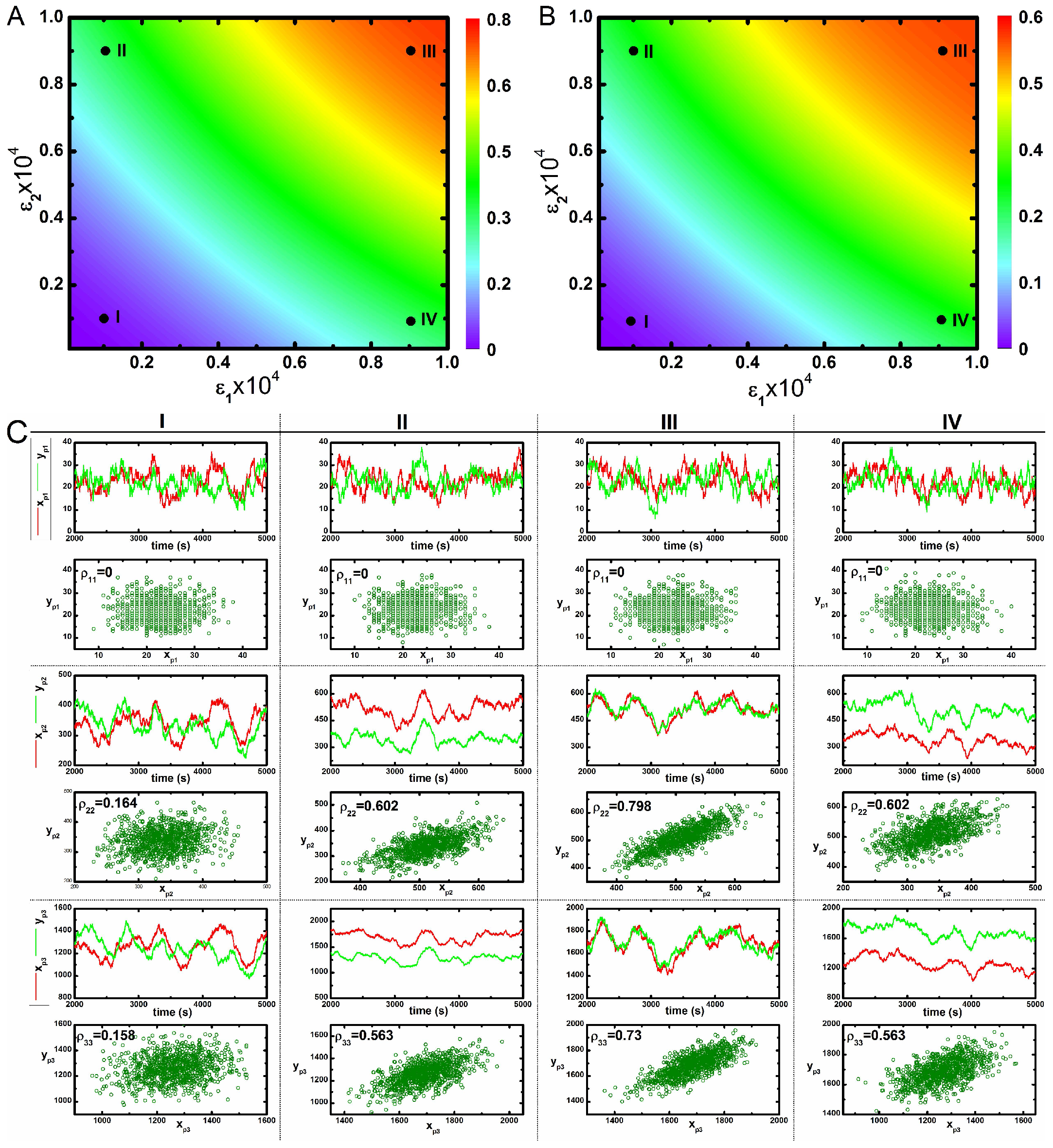
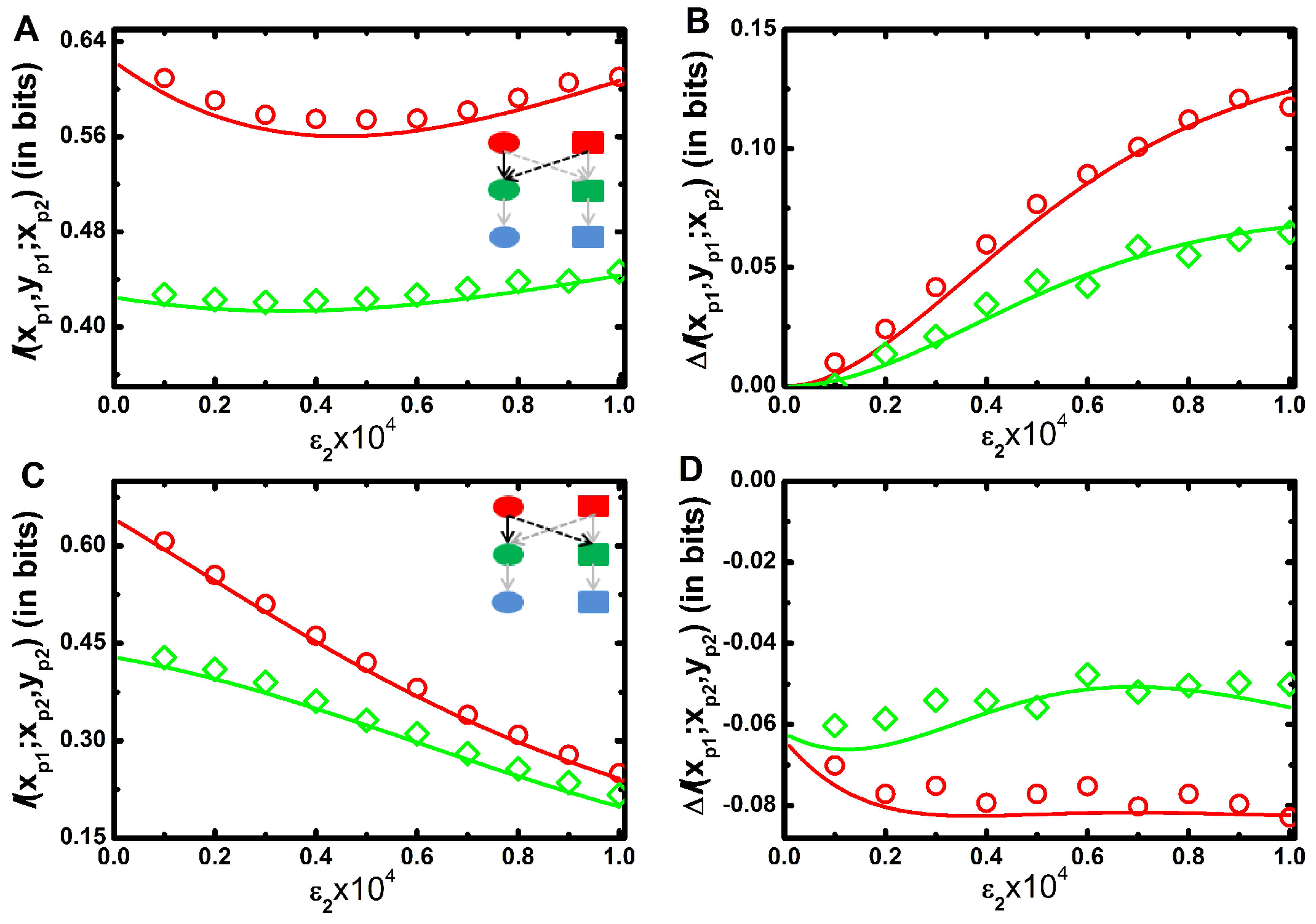
2. Results and Discussion
2.1. Two Variable Mutual Information
2.2. Three Variable Mutual Information
3. Materials and Methods
3.1. Two Variable Mutual Information
3.2. Three Variable Mutual Information
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MAPK | Mitogen activated protein kinase |
| LNA | Linear noise approximation |
| PID | Partial information decomposition |
Appendix A. Calculation of variance and covariance
References
- Tyson, J.J.; Chen, K.C.; Novak, B. Sniffers, buzzers, toggles and blinkers: dynamics of regulatory and signaling pathways in the cell. Curr. Opin. Cell Biol. 2003, 15, 221–231. [Google Scholar] [CrossRef]
- Alon, U. An Introduction to Systems Biology: Design Principles of Biological Circuits; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Lyons, S.M.; Prasad, A. Cross-Talk and Information Transfer in Mammalian and Bacterial Signaling. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Tareen, A.; Wingreen, N.S.; Mukhopadhyay, R. Modeling Evolution of Crosstalk in Noisy Signal Transduction Networks. arXiv, 2017; arXiv:physics.bio-ph/1707.01467. [Google Scholar]
- Bardwell, L. Mechanisms of MAPK signalling specificity. Biochem. Soc. Trans. 2006, 34, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Ferrell, J.E. Tripping the switch fantastic: How a protein kinase cascade can convert graded inputs into switch-like outputs. Trends Biochem. Sci. 1996, 21, 460–466. [Google Scholar] [CrossRef]
- Huang, C.Y.F.; Ferrell, J.E. Ultrasensitivity in the mitogen-activated protein kinase cascade. Proc. Natl. Acad. Sci. USA 1996, 93, 10078–10083. [Google Scholar] [CrossRef] [PubMed]
- Saito, H. Regulation of cross-talk in yeast MAPK signaling pathways. Curr. Opin. Microbiol. 2010, 13, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Voliotis, M.; Perrett, R.M.; McWilliams, C.; McArdle, C.A.; Bowsher, C.G. Information transfer by leaky, heterogeneous, protein kinase signaling systems. Proc. Natl. Acad. Sci. USA 2014, 111, E326–E333. [Google Scholar] [CrossRef] [PubMed]
- Laub, M.T.; Goulian, M. Specificity in two-component signal transduction pathways. Annu. Rev. Genet. 2007, 41, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Posas, F.; WurglerMurphy, S.M.; Maeda, T.; Witten, E.A.; Thai, T.C.; Saito, H. Yeast HOG1 MAP kinase cascade is regulated by a multistep phosphorelay mechanism in the SLN1-YPD1-SSK1 ”two-component” osmosensor. Cell 1996, 86, 865–875. [Google Scholar] [CrossRef]
- Rowland, M.A.; Deeds, E.J. Crosstalk and the evolution of specificity in two-component signaling. Proc. Natl. Acad. Sci. USA 2014, 111. [Google Scholar] [CrossRef] [PubMed]
- Siryaporn, A.; Perchuk, B.S.; Laub, M.T.; Goulian, M. Evolving a robust signal transduction pathway from weak cross-talk. Mol. Syst. Biol. 2010, 6. [Google Scholar] [CrossRef] [PubMed]
- Trach, K.A.; Hoch, J.A. Multisensory Activation of the Phosphorelay Initiating Sporulation in Bacillus Subtilis—Identification and Sequence of the Protein-Kinase of the Alternate Pathway. Mol. Microbiol. 1993, 8, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Gustin, M.C.; Albertyn, J.; Alexander, M.; Davenport, K. MAP kinase pathways in the yeast Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 1998, 62, 1264–1300. [Google Scholar] [PubMed]
- Kunkel, B.N.; Brooks, D.M. Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol. 2002, 5, 325–331. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Hayden, M.S.; Ghosh, S. Crosstalk in NF-κB signaling pathways. Nat. Immunol. 2011, 12, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Iborra, F.J.; Escargueil, A.E.; Kwek, K.Y.; Akoulitchev, A.; Cook, P.R. Molecular cross-talk between the transcription, translation, and nonsense-mediated decay machineries. J. Cell Sci. 2004, 117, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Mather, W.H.; Hasty, J.; Tsimring, L.S.; Williams, R.J. Translational Cross Talk in Gene Networks. Biophys. J. 2013, 104, 2564–2572. [Google Scholar] [CrossRef] [PubMed]
- Mauri, M.; Klumpp, S. A Model for Sigma Factor Competition in Bacterial Cells. PLoS Comput. Biol. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Riba, A.; Bosia, C.; El Baroudi, M.; Ollino, L.; Caselle, M. A Combination of Transcriptional and MicroRNA Regulation Improves the Stability of the Relative Concentrations of Target Genes. PLoS Comput. Biol. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Tsimring, L.S. Noise in biology. Rep. Prog. Phys. 2014, 77. [Google Scholar] [CrossRef] [PubMed]
- Palus, M.; Komarek, V.; Hrncir, Z.; Sterbova, K. Synchronization as adjustment of information rates: Detection from bivariate time series. Phys. Rev. E 2001, 63. [Google Scholar] [CrossRef] [PubMed]
- Cookson, N.A.; Mather, W.H.; Danino, T.; Mondragon-Palomino, O.; Williams, R.J.; Tsimring, L.S.; Hasty, J. Queueing up for enzymatic processing: correlated signaling through coupled degradation. Mol. Syst. Biol. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Komili, S.; Silver, P.A. Coupling and coordination in gene expression processes: a systems biology view. Nat. Rev. Genet. 2008, 9, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Bardwell, L.; Zou, X.F.; Nie, Q.; Komarova, N.L. Mathematical models of specifcity in cell signaling. Biophys. J. 2007, 92, 3425–3441. [Google Scholar] [CrossRef] [PubMed]
- Komarova, N.L.; Zou, X.F.; Nie, Q.; Bardwell, L. A theoretical framework for specificity in cell signaling. Mol. Syst. Biol. 2005, 1. [Google Scholar] [CrossRef] [PubMed]
- McClean, M.N.; Mody, A.; Broach, J.R.; Ramanathan, S. Cross-talk and decision making in MAP kinase pathways. Nat. Genet. 2007, 39, 567. [Google Scholar] [CrossRef]
- Ubersax, J.A.; Ferrell, J.E. Mechanisms of specificity in protein phosphorylation. Nat. Rev. Mol. Cell Biol. 2007, 8, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Van Kampen, N.G. Stochastic Processes in Physics and Chemistry; North-Holland: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Gardiner, C.W. Stochastic Methods, 4th ed.; Springer: Berlin, Germany, 2009. [Google Scholar]
- Grima, R. Linear-noise approximation and the chemical master equation agree up to second-order moments for a class of chemical systems. Phys. Rev. E 2015, 92. [Google Scholar] [CrossRef] [PubMed]
- Elf, J.; Ehrenberg, M. Fast evaluation of fluctuations in biochemical networks with the linear noise approximation. Genome Res. 2003, 13, 2475–2484. [Google Scholar] [CrossRef] [PubMed]
- Maity, A.K.; Bandyopadhyay, A.; Chaudhury, P.; Banik, S.K. Role of functionality in two-component signal transduction: a stochastic study. Phys. Rev. E 2014, 89. [Google Scholar] [CrossRef] [PubMed]
- Maity, A.K.; Chaudhury, P.; Banik, S.K. Role of relaxation time scale in noisy signal transduction. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Paulsson, J. Summing up the noise in gene networks. Nature 2004, 427, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Paulsson, J. Models of stochastic gene expression. Phys. Life Rev. 2005, 2, 157–175. [Google Scholar] [CrossRef]
- Grima, R.; Thomas, P.; Straube, A.V. How accurate are the nonlinear chemical Fokker-Planck and chemical Langevin equations? J. Chem. Phys. 2011, 135. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Matuschek, H.; Grima, R. How reliable is the linear noise approximation of gene regulatory networks? BMC Genom. 2013, 14 (Suppl. S4). [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Straube, A.V.; Timmer, J.; Fleck, C.; Grima, R. Signatures of nonlinearity in single cell noise-induced oscillations. J. Theor. Biol. 2013, 335, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.M.; Thomas, J.A. Elements of Information Theory; Wiley Interscience: Hoboken, NJ, USA, 2012. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 623–656. [Google Scholar] [CrossRef]
- Gillespie, D.T. A general method for numerically simulating the stochastic time evolution of coupled chemical reactions. J. Comput. Phys. 1976, 22, 403–434. [Google Scholar] [CrossRef]
- Granger, C.W.J. Investigating Causal Relations by Econometric Models and Cross-Spectral Methods. Econometrica 1969, 37. [Google Scholar] [CrossRef]
- Schreiber, T. Measuring Information Transfer. Phys. Rev. Lett. 2000, 85, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Faes, L.; Kugiumtzis, D.; Nollo, G.; Jurysta, F.; Marinazzo, D. Estimating the decomposition of predictive information in multivariate systems. Phys. Rev. E 2015, 91. [Google Scholar] [CrossRef] [PubMed]
- Faes, L.; Porta, A.; Nollo, G. Information Decomposition in Bivariate Systems: Theory and Application to Cardiorespiratory Dynamics. Entropy 2015, 17, 277–303. [Google Scholar] [CrossRef]
- Spinney, R.E.; Lizier, J.T.; Prokopenko, M. Transfer entropy in physical systems and the arrow of time. Phys. Rev. E 2016, 94. [Google Scholar] [CrossRef] [PubMed]
- Wibral, M.; Priesemann, V.; Kay, J.W.; Lizier, J.T.; Phillips, W.A. Partial information decomposition as a unified approach to the specification of neural goal functions. Brain Cognit. 2017, 112, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Schneidman, E.; Bialek, W.; Berry, M.J. Synergy, redundancy, and independence in population codes. J. Neurosci. 2003, 23, 11539–11553. [Google Scholar] [PubMed]
- Williams, P.L.; Beer, R.D. Nonnegative decomposition of Multivariate Information. arXiv, 2010; arXiv:cs.IT/1004.2515. [Google Scholar]
- Harder, M.; Salge, C.; Polani, D. Bivariate measure of redundant information. Phys. Rev. E 2013, 87. [Google Scholar] [CrossRef] [PubMed]
- Bertschinger, N.; Rauh, J.; Olbrich, E.; Jost, J.; Ay, N. Quantifying Unique Information. Entropy 2014, 16, 2161–2183. [Google Scholar] [CrossRef]
- Griffith, V.; Koch, C. Quantifying synergistic mutual information. In Guided Self-Organization: Inception, Emergence, Complexity and Computation; Prokopenko, M., Ed.; Springer: Berlin, Germany, 2014; Volume 9, pp. 159–190. [Google Scholar]
- Barrett, A.B. Exploration of synergistic and redundant information sharing in static and dynamical Gaussian systems. Phys. Rev. E 2015, 91. [Google Scholar] [CrossRef] [PubMed]
- Faes, L.; Porta, A.; Nollo, G.; Javorka, M. Information Decomposition in Multivariate Systems: Definitions, Implementation and Application to Cardiovascular Networks. Entropy 2017, 19, 5. [Google Scholar] [CrossRef]
- Dunlop, M.J.; Cox, R.S.; Levine, J.H.; Murray, R.M.; Elowitz, M.B. Regulatory activity revealed by dynamic correlations in gene expression noise. Nat. Genet. 2008, 40, 1493–1498. [Google Scholar] [CrossRef] [PubMed]
- Munsky, B.; Neuert, G.; van Oudenaarden, A. Using Gene Expression Noise to Understand Gene Regulation. Science 2012, 336, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Elowitz, M.B.; Levine, A.J.; Siggia, E.D.; Swain, P.S. Stochastic gene expression in a single cell. Science 2002, 297, 1183–1186. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Ann. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; Goyal, S.; Long, T.; Bassler, B.L.; Wingreen, N.S. Information processing and signal integration in bacterial quorum sensing. Mol. Syst. Biol. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Armitage, J.P. Bacterial tactic responses. Adv. Microb. Physiol. 1999, 41, 229–289. [Google Scholar] [PubMed]
- Heinrich, R.; Neel, B.G.; Rapoport, T.A. Mathematical models of protein kinase signal transduction. Mol. Cell 2002, 9, 957–970. [Google Scholar] [CrossRef]
- Suderman, R.; Deeds, E.J. Machines vs. Ensembles: Effective MAPK Signaling through Heterogeneous Sets of Protein Complexes. PLoS Comput. Biol. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Tănase-Nicola, S.; Warren, P.B.; ten Wolde, P.R. Signal detection, modularity, and the correlation between extrinsic and intrinsic noise in biochemical networks. Phys. Rev. Lett. 2006, 97. [Google Scholar] [CrossRef] [PubMed]
- Bardwell, L.; Cook, J.G.; Chang, E.C.; Cairns, B.R.; Thorner, J. Signaling in the yeast pheromone response pathway: specific and high-affinity interaction of the mitogen-activated protein (MAP) kinases Kss1 and Fus3 with the upstream MAP kinase kinase Ste7. Mol. Cell Biol. 1996, 16, 3637–3650. [Google Scholar] [CrossRef] [PubMed]
- Atay, O.; Skotheim, J.M. Spatial and temporal signal processing and decision making by MAPK pathways. J. Cell Biol. 2017, 216, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, M.C.; Schluter, D. The Analysis of Biological Data; Roberts and Company Publishers: Greenwood Village, CO, USA, 2009. [Google Scholar]
- White, R.J.; Sharrocks, A.D. Coordinated control of the gene expression machinery. Trends Genet. 2010, 26, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Stewart-Ornstein, J.; Weissman, J.S.; El-Samad, H. Cellular Noise Regulons Underlie Fluctuations in Saccharomyces cerevisiae. Mol. Cell 2012, 45, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Junttila, M.R.; Li, S.P.; Westermarck, J. Phosphatase-mediated crosstalk between MAPK signalling pathways in the regulation of cell survival. FASEB J. 2008, 22, 954–965. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Stock, A.M. Probing kinase and phosphatase activities of two-component systems in vivo with concentration-dependent phosphorylation profiling. Proc. Natl. Acad. Sci. USA 2013, 110, 672–677. [Google Scholar] [CrossRef] [PubMed]
- Schulz, K.R.; Danna, E.A.; Krutzik, P.O.; Nolan, G.P. Single-cell phospho-protein analysis by flow cytometry. Curr. Protoc. Immunol. 2012, Chapter 8, 1–20. [Google Scholar] [CrossRef]
- Yaginuma, H.; Kawai, S.; Tabata, K.V.; Tomiyama, K.; Kakizuka, A.; Komatsuzaki, T.; Noji, H.; Imamura, H. Diversity in ATP concentrations in a single bacterial cell population revealed by quantitative single-cell imaging. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- De Ronde, W.H.; Tostevin, F.; ten Wolde, P.R. Feed-forward loops and diamond motifs lead to tunable transmission of information in the frequency domain. Phys. Rev. E 2012, 86. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; Goyal, S.; Wingreen, N.S. A quantitative comparison of sRNA-based and protein-based gene regulation. Mol. Syst. Biol. 2008, 4. [Google Scholar] [CrossRef] [PubMed]
- Thattai, M.; van Oudenaarden, A. Stochastic gene expression in fluctuating environments. Genetics 2004, 167, 523–530. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Reaction | Propensity Function | Rate Constant | |
|---|---|---|---|---|
| Activation of | molecules s | |||
| Deactivation of | s | |||
| Activation of | molecules s | |||
| Deactivation of | s | |||
| Activation of | molecules s | |||
| Activation of | molecules s | |||
| Deactivation of | s | |||
| Activation of | molecules s | |||
| Activation of | molecules s | |||
| Deactivation of | s | |||
| Activation of | molecules s | |||
| Deactivation of | s | |||
| Activation of | molecules s | |||
| Deactivation of | s |
| Description | Reaction | Propensity Function | Rate Constant | |
|---|---|---|---|---|
| Activation of | molecules s | |||
| Deactivation of | s | |||
| Activation of | molecules s | |||
| Deactivation of | s | |||
| Activation of | molecules s | |||
| Activation of | molecules s | |||
| Deactivation of | s | |||
| Activation of | molecules s | |||
| Activation of | molecules s | |||
| Deactivation of | s | |||
| Activation of | molecules s | |||
| Deactivation of | s | |||
| Activation of | molecules s | |||
| Deactivation of | s |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maity, A.K.; Chaudhury, P.; Banik, S.K. Information Theoretical Study of Cross-Talk Mediated Signal Transduction in MAPK Pathways. Entropy 2017, 19, 469. https://doi.org/10.3390/e19090469
Maity AK, Chaudhury P, Banik SK. Information Theoretical Study of Cross-Talk Mediated Signal Transduction in MAPK Pathways. Entropy. 2017; 19(9):469. https://doi.org/10.3390/e19090469
Chicago/Turabian StyleMaity, Alok Kumar, Pinaki Chaudhury, and Suman K. Banik. 2017. "Information Theoretical Study of Cross-Talk Mediated Signal Transduction in MAPK Pathways" Entropy 19, no. 9: 469. https://doi.org/10.3390/e19090469
APA StyleMaity, A. K., Chaudhury, P., & Banik, S. K. (2017). Information Theoretical Study of Cross-Talk Mediated Signal Transduction in MAPK Pathways. Entropy, 19(9), 469. https://doi.org/10.3390/e19090469