
Increasing Natural Killer Cell Activity of Mineral Nanomaterial ALP1018 in Healthy Adults: A Randomized, Double-Blind, Placebo Comparative Clinical Trial


Abstract
:1. Introduction
2. Materials and Methods
2.1. Trial Supplements
2.2. Participants
2.3. Study Design, Screening, and Randomization
2.4. Evaluation of Diet and Physical Activity
2.5. Outcome Measures
2.5.1. NK Cell Activity
2.5.2. Cytokine Levels
2.6. Safety Outcome Measurements
2.7. Statistical Analysis
3. Results
3.1. Participant Demographic Characteristics
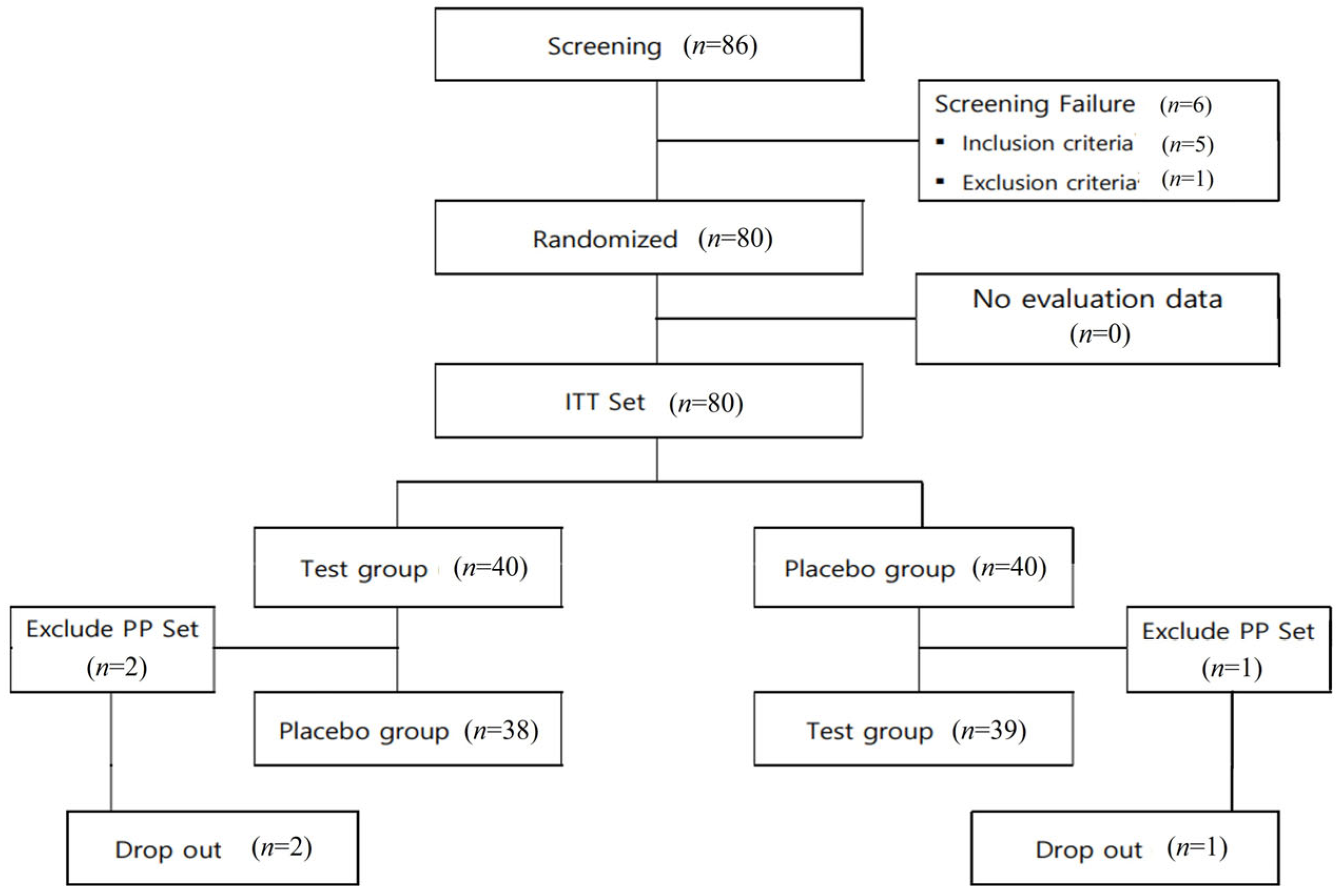
3.2. Trial Procedure and Follow-Up
3.3. Dietary Intake and Physical Activity Evaluation
3.4. Efficacy and Safety Evaluation
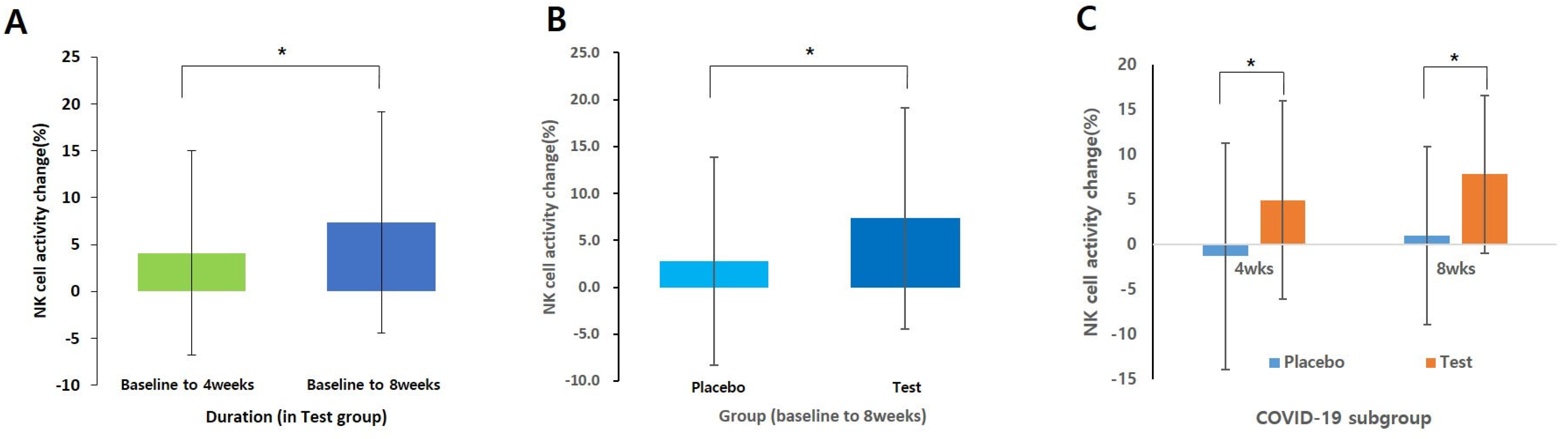
3.4.1. NK Cell Activity
3.4.2. Cytokine Levels
3.4.3. Safety Outcome Monitoring
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, R.; Kupper, T. Old Meets New: The Interaction Between Innate and Adaptive Immunity. J. Investig. Dermatol. 2005, 125, 629–637. [Google Scholar] [CrossRef]
- Gaforio, J.J.; Ortega, E.; Algarra, I.; Serrano, M.J.; Alvarez de Cienfuegos, G. NK Cells Mediate Increase of Phagocytic Activity but Not of Proinflammatory Cytokine (Interleukin-6 [IL-6], Tumor Necrosis Factor Alpha, and IL-12) Production Elicited in Splenic Macrophages by Tilorone Treatment of Mice during Acute Systemic Candidiasis. Clin. Diagn. Lab. Immunol. 2002, 9, 1282. [Google Scholar] [CrossRef] [PubMed]
- Topham, N.J.; Hewitt, E.W. Natural Killer Cell Cytotoxicity: How Do They Pull the Trigger? Immunology 2009, 128, 7. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of Natural Killer Cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Cichocki, F.; Sitnicka, E.; Bryceson, Y.T. NK Cell Development and Function—Plasticity and Redundancy Unleashed. Semin. Immunol. 2014, 26, 114–126. [Google Scholar] [CrossRef]
- Sun, J.C.; Lanier, L.L. NK Cell Development, Homeostasis and Function: Parallels with CD8+ T Cells. Nat. Rev. Immunol. 2011, 11, 645. [Google Scholar] [CrossRef] [PubMed]
- Cronin, S.J.F.; Woolf, C.J.; Weiss, G.; Penninger, J.M. The Role of Iron Regulation in Immunometabolism and Immune-Related Disease. Front. Mol. Biosci. 2019, 6, 116. [Google Scholar] [CrossRef]
- Shankar, A.H.; Prasad, A.S. Zinc and Immune Function: The Biological Basis of Altered Resistance to Infection. Am. J. Clin. Nutr. 1998, 68, 447S–463S. [Google Scholar] [CrossRef]
- Mahoro, P.; Kwon, E.; Kim, Y.S.; Bae, J.M.; Kim, S.A.; Cho, J.; Cha, Y.S.; Kim, S. In Vitro and in Vivo Genotoxicity Evaluation of ALP1018, a Nanomineral Food Supplement. Food Chem. Toxicol. 2024, 184, 114437. [Google Scholar] [CrossRef]
- Björkström, N.K.; Strunz, B.; Ljunggren, H.G. Natural Killer Cells in Antiviral Immunity. Nat. Rev. Immunol. 2021, 22, 112–123. [Google Scholar] [CrossRef]
- Kwon, H.J.; Kim, N.; Kim, H.S. Molecular Checkpoints Controlling Natural Killer Cell Activation and Their Modulation for Cancer Immunotherapy. Exp. Mol. Med. 2017, 49, e311. [Google Scholar] [CrossRef]
- Bryceson, Y.T.; Long, E.O. Line of Attack: NK Cell Specificity and Integration of Signals. Curr. Opin. Immunol. 2008, 20, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Wolf, N.K.; Kissiov, D.U.; Raulet, D.H. Roles of Natural Killer Cells in Immunity to Cancer, and Applications to Immunotherapy. Nat. Rev. Immunol. 2022, 23, 90–105. [Google Scholar] [CrossRef]
- Herberman, R.B.; Ortaldo, J.R. Natural Killer Cells: Their Roles in Defenses against Disease. Science 1981, 214, 24–30. [Google Scholar] [CrossRef]
- Li, Q.; Morimoto, K.; Nakadai, A.; Qu, T.; Matsushima, H.; Katsumata, M.; Shimizu, T.; Inagaki, H.; Hirata, Y.; Hirata, K.; et al. Healthy Lifestyles Are Associated with Higher Levels of Perforin, Granulysin and Granzymes A/B-Expressing Cells in Peripheral Blood Lymphocytes. Prev. Med. 2007, 44, 117–123. [Google Scholar] [CrossRef]
- Morimoto, K.; Takeshita, T.; Inoue-Sakurai, C.; Maruyama, S. Lifestyles and Mental Health Status Are Associated with Natural Killer Cell and Lymphokine-Activated Killer Cell Activities. Sci. Total. Environ. 2001, 270, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Kusaka, Y.; Kondou, H.; Morimoto, K. Healthy Lifestyles Are Associated with Higher Natural Killer Cell Activity. Prev. Med. 1992, 21, 602–615. [Google Scholar] [CrossRef]
- Wang, D.; Uyemura, B.; Hashemi, E.; Bjorgaard, S.; Riese, M.; Verbsky, J.; Thakar, M.S.; Malarkannan, S. Role of GATA2 in Human NK Cell Development. Crit. Rev. Immunol. 2021, 41, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Gineau, L.; Cognet, C.; Kara, N.; Lach, F.P.; Dunne, J.; Veturi, U.; Picard, C.; Trouillet, C.; Eidenschenk, C.; Aoufouchi, S.; et al. Partial MCM4 Deficiency in Patients with Growth Retardation, Adrenal Insufficiency, and Natural Killer Cell Deficiency. J. Clin. Investig. 2012, 122, 821–832. [Google Scholar] [CrossRef]
- Mace, E.M.; Paust, S.; Conte, M.I.; Baxley, R.M.; Schmit, M.M.; Patil, S.L.; Guilz, N.C.; Mukherjee, M.; Pezzi, A.E.; Chmielowiec, J.; et al. Human NK Cell Deficiency as a Result of Biallelic Mutations in MCM10. J. Clin. Investig. 2020, 130, 5272–5286. [Google Scholar] [CrossRef]
- Mace, E.M.; Bigley, V.; Gunesch, J.T.; Chinn, I.K.; Angelo, L.S.; Care, M.A.; Maisuria, S.; Keller, M.D.; Togi, S.; Watkin, L.B.; et al. Biallelic Mutations in IRF8 Impair Human NK Cell Maturation and Function. J. Clin. Investig. 2017, 127, 306–320. [Google Scholar] [CrossRef] [PubMed]
- De Vries, E.; Koene, H.R.; Vossen, J.M.; Gratama, J.W.; Von Dem Borne, A.E.G.K.; Waaijer, J.L.M.; Haraldsson, A.; De Haas, M.; Van Tol, M.J.D. Identification of an Unusual Fcγ Receptor IIIa (CD16) on Natural Killer Cells in a Patient with Recurrent Infections. Blood 1996, 88, 3022–3027. [Google Scholar] [CrossRef]
- Grier, J.T.; Forbes, L.R.; Monaco-Shawver, L.; Oshinsky, J.; Atkinson, T.P.; Moody, C.; Pandey, R.; Campbell, K.S.; Orange, J.S. Human Immunodeficiency-Causing Mutation Defines CD16 in Spontaneous NK Cell Cytotoxicity. J. Clin. Investig. 2012, 122, 3769–3780. [Google Scholar] [CrossRef]
- Mace, E.M.; Orange, J.S. Emerging Insights into Human Health and NK Cell Biology from the Study of NK Cell Deficiencies. Immunol. Rev. 2019, 287, 202–225. [Google Scholar] [CrossRef] [PubMed]
- Biron, C.A.; Byron, K.S.; Sullivan, J.L. Severe Herpesvirus Infections in an Adolescent without Natural Killer Cells. N. Engl. J. Med. 1989, 320, 1731–1735. [Google Scholar] [CrossRef]
- Pegram, H.J.; Andrews, D.M.; Smyth, M.J.; Darcy, P.K.; Kershaw, M.H. Activating and Inhibitory Receptors of Natural Killer Cells. Immunol. Cell. Biol. 2011, 89, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.S. Biology of Natural Killer Cells in Cancer and Infection. Cancer Investig. 2002, 20, 405–419. [Google Scholar] [CrossRef]
- Wiley, S.R.; Schooley, K.; Smolak, P.J.; Din, W.S.; Huang, C.P.; Nicholl, J.K.; Sutherland, G.R.; Smith, T.D.; Rauch, C.; Smith, C.A.; et al. Identification and Characterization of a New Member of the TNF Family That Induces Apoptosis. Immunity 1995, 3, 673–682. [Google Scholar] [CrossRef]
- Smyth, M.J.; Cretney, E.; Kelly, J.M.; Westwood, J.A.; Street, S.E.A.; Yagita, H.; Takeda, K.; Dommelen, S.L.H.V.; Degli-Esposti, M.A.; Hayakawa, Y. Activation of NK Cell Cytotoxicity. Mol. Immunol. 2005, 42, 501–510. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Guan, W.; Ni, Z.; Hu, Y.; Liang, W.; Ou, C.; He, J.; Liu, L.; Shan, H.; Lei, C.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Van Eeden, C.; Khan, L.; Osman, M.S.; Tervaert, J.W.C. Natural Killer Cell Dysfunction and Its Role in COVID-19. Int. J. Mol. Sci. 2020, 21, 6351. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, Q.; Cai, S.; Peng, H.; Huyan, T.; Yang, H. The Role of NK Cells in Fighting the Virus Infection and Sepsis. Int. J. Med. Sci. 2021, 18, 3236. [Google Scholar] [CrossRef] [PubMed]
- Di Vito, C.; Calcaterra, F.; Coianiz, N.; Terzoli, S.; Voza, A.; Mikulak, J.; Della Bella, S.; Mavilio, D. Natural Killer Cells in SARS-CoV-2 Infection: Pathophysiology and Therapeutic Implications. Front. Immunol. 2022, 13, 888248. [Google Scholar] [CrossRef] [PubMed]
- Sherman, A.R. Zinc, Copper, and Iron Nutriture and Immunity. J. Nutr. 1992, 122, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Sherman, A.R.; Lockwood, J.F. Impaired Natural Killer Cell Activity in Iron-Deficient Rat Pups. J. Nutr. 1987, 117, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Hallquist, N.A.; Sherman, A.R. Effect of Iron Deficiency on the Stimulation of Natural Killer Cells by Macrophage-Produced Interferon. Nutr. Res. 1989, 9, 283–292. [Google Scholar] [CrossRef]
- Spear, A.T.; Sherman, A.R. Iron Deficiency Alters DMBA-Induced Tumor Burden and Natural Killer Cell Cytotoxicity in Rats. J. Nutr. 1992, 122, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, G.; Erbas, D.; i̇mi̇r, T.; Bor, N.M. Decreased Natural Killer (NK) Cell Activity in Zinc-Deficient Rats. Gen. Pharmacol. Vasc. Syst. 1994, 25, 1499–1503. [Google Scholar] [CrossRef]
- Rolles, B.; Maywald, M.; Rink, L. Influence of Zinc Deficiency and Supplementation on NK Cell Cytotoxicity. J. Funct. Foods 2018, 48, 322–328. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Inclusion Criteria |
|---|
|
| Exclusion Criteria |
|
| Variables | Test Group | Placebo Group | Total | p- Value 1 | ||||
|---|---|---|---|---|---|---|---|---|
| n | % | n | % | n | % | |||
| Sex | Male | 13 | 32.5 | 8 | 20.0 | 21 | 26.3 | 0.204 |
| Female | 27 | 67.5 | 32 | 80.0 | 59 | 73.8 | ||
| Age (year) | Mean ± SD | 40.85 ± 9.68 | 39.50 ± 10.19 | 40.18 ± 9.90 | 0.545 | |||
| Min~Max | 23~60 | 22~55 | 22~60 | |||||
| Height (cm) | Mean ± SD | 164.87 ± 8.03 | 163.87 ± 7.12 | 164.37 ± 7.56 | 0.558 | |||
| Min~Max | 151.3~182.3 | 151.0~183.0 | 151.0~183.0 | |||||
| Weight (kg) | Mean ± SD | 62.73 ± 13.51 | 59.82 ± 12.47 | 61.27 ± 13.01 | 0.319 | |||
| Min~Max | 40.0~93.5 | 44.0~94.0 | 40.0~94.0 | |||||
| BMI (kg/m2) | Mean ± SD | 22.82 ± 3.06 | 22.13 ± 3.26 | 22.48 ± 3.16 | 0.329 | |||
| Min~Max | 16.2~29.7 | 17.9~29.5 | 16.2~29.7 | |||||
| SBP (mmHg) | Mean ± SD | 118.58 ± 13.24 | 117.60 ± 11.42 | 118.09 ± 12.30 | 0.725 | |||
| Min~Max | 91.0~159.0 | 95.0~141.0 | 91.0~159.0 | |||||
| DBP (mmHg) | Mean ± SD | 73.18 ± 9.96 | 72.68 ± 9.72 | 72.93 ± 9.78 | 0.821 | |||
| Min~Max | 56.0~90.0 | 49.0~97.0 | 49.0~97.0 | |||||
| Pulse (ppm) | Mean ± SD | 76.48 ± 9.14 | 77.15 ± 8.16 | 76.81 ± 8.62 | 0.729 | |||
| Min~Max | 55.0~92.0 | 60.0~96.0 | 55.0~96.0 | |||||
| Alcohol | Drinker | 9 | 22.5 | 10 | 25.0 | 19 | 23.8 | - |
| Amount (units/Week) | 12.22 ± 7.40 | 11.04 ± 15.05 | 11.60 ± 11.75 | 0.834 | ||||
| Smoke | Smoker | 2 | 5.0 | 2 | 5.0 | 4 | 5.0 | - |
| Amount (ea/day) | 6.00 ± 1.41 | 10.50 ± 0.71 | 8.25 ± 2.75 | 0.057 | ||||
| Duration (Year) | 37.50 ± 3.54 | 32.50 ± 17.68 | 35.00 ± 10.80 | 0.733 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-A.; Kim, S.; Chai, H.; Cho, J.; Paek, Y.-J. Increasing Natural Killer Cell Activity of Mineral Nanomaterial ALP1018 in Healthy Adults: A Randomized, Double-Blind, Placebo Comparative Clinical Trial. Nutrients 2024, 16, 850. https://doi.org/10.3390/nu16060850
Kim S-A, Kim S, Chai H, Cho J, Paek Y-J. Increasing Natural Killer Cell Activity of Mineral Nanomaterial ALP1018 in Healthy Adults: A Randomized, Double-Blind, Placebo Comparative Clinical Trial. Nutrients. 2024; 16(6):850. https://doi.org/10.3390/nu16060850
Chicago/Turabian StyleKim, Soon-Ae, Seyl Kim, Hyungyung Chai, Junlae Cho, and Yu-Jin Paek. 2024. "Increasing Natural Killer Cell Activity of Mineral Nanomaterial ALP1018 in Healthy Adults: A Randomized, Double-Blind, Placebo Comparative Clinical Trial" Nutrients 16, no. 6: 850. https://doi.org/10.3390/nu16060850