Abstract
Food allergy (FA) is a growing public health concern, with an increasing prevalence in Western countries. Increasing evidence suggests that the balance of human gut microbiota and the integrity of our intestinal barrier may play roles in the development of FA. Environmental factors, including industrialization and consumption of highly processed food, can contribute to altering the gut microbiota and the intestinal barrier, increasing the susceptibility to allergic sensitization. Compositional and functional alterations to the gut microbiome have also been associated with FA. In addition, increased permeability of the gut barrier allows the translocation of allergenic molecules, triggering Th2 immune responses. Preclinical and clinical studies have highlighted the potential of probiotics, prebiotics, and postbiotics in the prevention and treatment of FA through enhancing gut barrier function and promoting the restoration of healthy gut microbiota. Finally, fecal microbiota transplantation (FMT) is now being explored as a promising therapeutic strategy to prevent FA in both experimental and clinical studies. In this review article, we aim to explore the complex interplay between intestinal permeability and gut microbiota in the development of FA, as well as depict potential therapeutic strategies.
1. Introduction to Leaky Gut and Gut Microbiota in Food Allergies
1.1. Gut Microbiota
The term “gut microbiota” refers to the complex community of microbes that inhabit the gut, which includes bacteria, fungi, viruses, and parasites [1]. Four major phyla, Firmicutes, Bacteroidetes, Actinobacteria and Proteobacteria, account for more than 90% of the whole bacteriome [2]. The composition of the gut microbiota varies from site to site, and it is extremely dynamic, depending on a huge number of factors, including age, diet, habits, drug use, and others [3,4,5,6]. Gut microbiota play a key role in human health and disease. As alterations to their composition and structure, generically defined as dysbiosis, may lead to diseases, a healthy microbiota helps in promoting and maintaining gut health, through a trophic effect on enterocytes, by maintaining intestinal barrier integrity, avoiding the adhesion of pathogenic bacteria, and producing vitamins [7,8,9]. Gut microbiota are also connected to other regions of the body, having complex metabolic systemic effects [10,11,12] and even interacting with the nervous system thanks to the so-called gut–brain axis [13,14].
Increasing evidence supports the role of gut microbiota in the pathogenesis of food allergies (FA) [15]. First, gut microbiota play a role in maintaining intestinal barrier integrity, as well as in modulating the gut inflammatory microenvironment. Moreover, modulation of the gut microbiota, from pre- and probiotics to fecal microbiota transplantation (FMT), is opening new and previously unknown possibilities for the treatment of conditions whose prevalence and social impact grows daily [16,17,18].
1.2. Intestinal Barrier
The primary function of the intestinal barrier is to facilitate the absorption of nutrients and fluids, while blocking the entry of harmful substances, such as toxins and pathogens, into the underlying tissue through the intestinal epithelium [19]. Moreover, it plays a vital role in ensuring the overall fitness and health of the body, protecting us from potentially harmful agents coming from the gut, and being the main controller of the substances that can be absorbed and the ones that should be blocked [20].
The intestinal barrier consists of multiple components. The outer layer includes various extracellular components, whose main actor is the mucus layer. It is composed of highly glycosylated mucin proteins, i.e., MUC2 [21]. The mucus layer contains an outer layer called the “stirred mucus layer” that acts as a first line of defense, due to the presence of secretory immunoglobulin A (sIgA) and antimicrobial products [22]. There is also an inner layer called the “non-stirred mucus layer” or “glycocalyx”, which is mainly devoted to nutrient absorption and epithelial protection, and cellular renewal and differentiation [23]. The intestinal barrier is also made of extracellular elements, including gut microbiota and digestive enzymes, such as proteases, lipases, amylases, and nucleases [24]. The inner layer of the intestinal barrier consists of epithelial cells that create a cohesive and polarized single layer that strictly controls the passage of molecules, through the aid of junctional complexes better described below in the text, carrying out both digestive and immune functions [25]. The intestinal barrier is mainly made up of enterocytes, but goblet cells, Paneth cells, enterochromaffin cells, and M cells can also be found. The pluripotent stem cells, which are important for the renewal of these elements, are located within the Lieberkühn crypts [26]. Underneath these two layers, there is the lamina propria, which contains innate and adaptive immune cells that build the gut-associated lymphoid tissue (GALT) [27], the connective tissue, and the enteric nervous system, composed by the myenteric (Auerbach’s) plexus and the submucosal (Meissner’s) plexus, which control motor and secretory functions [28]. Three main pathways allow molecules to cross the epithelial layer: the paracellular pathway, which allows the passage of substances through the intercellular space between cells; the carrier-mediated pathway, where substances are transported through the aid of carrier proteins; and the trans-cellular pathway, where substances pass through the cell [29].
Specialized areas of contact between the plasma membranes of neighboring cells are known as intercellular junctions. There are three main types of intercellular junctions: tight junctions, which are occluding junctions that enable the establishment and maintenance of concentration gradients across an epithelium [30]; anchoring junctions, which facilitate the adhesion of cells to either other cells or the substrate, enabling the distribution of local tensile forces across a tissue [31]; and GAP junctions, which facilitate communication between adjacent cells, enabling the exchange of ions or small molecules between closely connected cells [32].
1.3. Food Allergies
FAs are pathological immune reactions triggered by normally innocuous food protein antigens [33]. Their prevalence rises every year [34], and it is estimated to be 5% for adults and 8% for children [35]; nevertheless, the significative cost for healthcare systems [36], the impact on the personal and social life of the patients, and findings about new possible therapies make FA a relevant and current topic.
FAs must be distinct from food intolerances, as in those conditions the immune system is not involved, while coeliac disease is not typically classified as a food-allergic disease [33]. FAs may be classified according to their immunopathogenic pathway into three categories. The IgE-mediated FA group sees food antigens binding to preformed IgE, triggering a type-1 hypersensitivity reaction with degranulation of mast cells and basophils, which results in rapid appearance of symptoms, usually within minutes [37]. The not-IgE mediated group is characterized by the action of allergen-specific T helper 2 cells, with more delayed symptoms [38]. Eventually, mixed FA form the third group, in which IgE-dependent and IgE-independent pathways coexist; patients suffering from this kind of allergy are usually affected by eosinophilic gastrointestinal disorders, such as eosinophilic esophagitis [33].
Food allergic reactions are only possible if a previous sensitization, defined as the presence of food-specific IgE, has already happened [39]. In normal conditions, the antigens that reach the intestinal barrier are unable to pass through it directly. However, they cross it thanks to cell-mediated mechanisms, like the action of goblet cells or CX3CR1 macrophages, which transfer the antigens to dendritic cells and those to draining lymph nodes [40,41], promoting the differentiation of T-naïve cells into food-antigen specific T regulatory cells [42]. When the epithelial barrier is damaged, antigens, as well as pathogens, may cross it freely, and this leads to the release of pro-inflammatory molecules like IL-25, IL-33, and thymic stromal lymphopoietin (TSLP), which promote T-naïve cell differentiation into T H2 cells, IgE class-switching, and tissue accumulation of mast cells and eosinophils [43,44,45]. Once sensitization has occurred, allergic reactions to that specific food antigen may occur, with symptoms ranging from cutaneous rashes to persistent diarrhea and potentially deadly anaphylaxis [37].
Gut microbiota are one of the main actors in the complex mechanism of sensitization [46,47]. Low microbial diversity and an elevated Enterobacteriaceae/Bacteroidaceae ratio are associated with subsequent food sensitization in children [48], and children born through caesarean (who do not acquire the mother’s vaginal microbiota) are known to be at higher risk of allergic diseases [49]. Moreover, bacterial metabolites like short-chain fatty acids (SCFAs) are also involved. SCFAs are end products of bacterial fermentation, mostly produced by Firmicutes, the most relevant of which are butyrate and propionate [50]. These molecules exert anti-inflammatory effects that improve the epithelial barrier integrity, reducing the possible sensitization to food allergens [51,52].
At present, avoidance of the food allergen and treatment of the allergic reactions remain the milestone of FA management [53]. Nevertheless, immunotherapy has opened the possibility of a real “definitive” treatment [54,55]; and prevention strategies such as the early assumption of specific foods, like peanuts, are increasingly coming into practice [56,57]. Among these findings, the role of microbiota as a key-player in the gut barrier has emerged as a new area of research and possible treatment for these conditions. In the next chapters, we will more deeply explore its involvement in FA pathophysiology, and how its modulation may relieve these conditions.
1.4. Leaky Gut
The term “leaky gut” defines a status of weakening or disruption of the intestinal barrier, in which substances that are normally confined in the intestinal cavity pass through the intestinal wall and enter the bloodstream [58]. The disruption of the intestinal barrier and the increase in the blood concentration of toxins have both local and systemic consequences [59,60]. First, an increased immune activation in the lamina propria through the pathway of Toll-like Receptor (TLR4) has been reported, whose ligand is mainly LPS [61]. By binding LPS to its receptor TLR4, the NF-κB pathway is activated, raising the concentration of pro-inflammatory cytokines [62]. This pro-inflammatory environment increases the amount of immune cells in various organs such as the liver, adipose tissue, and muscles, enhancing insulin resistance [63] and fostering the process of atherosclerosis [64].
Many factors can lead to an increase in intestinal permeability, including diet, inflammatory conditions like inflammatory bowel diseases (IBD), cirrhosis and its complications, burn injuries [65], and alcohol [66] through different pathophysiological pathways [67]. Specific alterations in daily diet can alter gut permeability. Reduced presence of the vitamin D receptor causes downregulation of claudin-2, a protein involved in the formation of tight junctions, increasing gut leakiness [68].
Moreover, deficiency in dietary fibers lets the microbiota feed on mucus glycoproteins of the host, disrupting the muco-epithelial barrier and increasing intestinal permeability [69]. In addition, a diet high in saturated fats decreases the abundance of Lactobacilli, while increasing the number of Oscillospiraceae, which is negatively correlated with the mRNA expression of Zonulin-1 (ZO-1), a barrier-forming TJ protein, leading to the condition of leaky gut [70].
Because of the disruption of the gut barrier, IBDs are characterized by a considerably leaky gut [71,72]. Patients with IBD experience both a loss in the number of tight junctions and a qualitative difference in their structure [73], as well as presenting altered types of intestinal cells. First, a reduced number of goblet cells can be observed in the epithelium of patients with IBD, and these cells also produce a minor amount of mucus [74]. Moreover, Paneth cells can be found in colonic mucosa and not only in the Lieberkühn crypts, resulting in an increase in the secretion of protective proteins in the large intestine [75]. This condition of intestinal inflammation and the disruption of the gut barrier result in an altered passage of substances in blood circulation, leading to systemic proinflammatory conditions [76].
Intestinal permeability is also altered in liver disorders, mainly in cirrhosis [77]. Due to the close relationship between gut and liver, because of the portal circulation, a large amount of pathogen-associated molecular patterns (PAMPs) reach the liver and increase its inflammation, speeding up liver diseases [78]. Patients with cirrhosis present different microbial profiling, with an increase in the abundance of bacteria in the phyla Proteobacteria and Fusobacteria and a reduction in bacteria in the phylum Bacteroidetes [79].
These alterations to gut microbiome, in addition to a reduced intestinal permeability, boost liver inflammation, which worsens the illness and its complications, such as spontaneous bacterial peritonitis (SBP) [80] and hepatic encephalopathy [81].
Thus, leaky gut plays a key role in the pathogenesis of many diseases, many of them being autoimmune diseases or with a disimmune pathogenetical component, like IBD, coeliac disease, autoimmune hepatitis, and multiple sclerosis [71,82,83,84]. The release of proinflammatory molecules and epithelial damage are factors shared by essentially all of these diseases [85]. In particular, the inflammatory pathway of FA, differently from other disorders, is characterized by the overactivation of the Th2 pathway and relative cytokines, like IL-4 and IL-13, as further explained [86].
1.5. Leaky Gut, Gut Microbiome, and Implications in Food Allergies
Leaky gut has garnered significant interest due to its potential role in the onset of FA. The “epithelial barrier hypothesis” assumes that intestinal barrier dysfunction, which increases susceptibility to environmental factors, can contribute to the sensitization and development of allergic diseases [87]. This hypothesis suggests that alterations in the intestinal integrity, often associated with leaky gut, might play a role in the observed rise in FA prevalence [88]. Environmental and lifestyle factors, such as industrialization and the consumption of ultra-processed foods (UPFs), are believed to contribute to disrupting intestinal barrier integrity [87]. These external factors, which constitute the external exposome, can influence the gut microbiome and epithelial barriers, playing a significant role in the development of allergic diseases [89]. Food emulsifiers in UPFs (i.e., polysorbate 20 and 80) have a detrimental effect on intestinal epithelial integrity [90]. Reduced levels of antioxidants and vitamins in UPFs are thought to increase susceptibility to allergic diseases [89]. Furthermore, it has been hypothesized that increased exposure to advanced glycation end products (AGEs) may be linked to the increased prevalence of FA [91].
A compromised epithelial barrier can trigger immune responses that result in the release of inflammatory mediators, known as alarmins [92]. Alarmins are epithelial-derived cytokines, such as thymic stromal lymphopoietin (TSLP), interleukin 33 (IL-33), and IL-25, released in response to the cellular damage caused by cellular stress or infection [93,94,95]. While these cytokines have crucial roles in maintaining gut epithelial homeostasis, they can also promote a pro-allergic microenvironment by activating T helper 2 (Figure 1 [96,97,98,99,100,101,102,103]) and type 2 innate lymphoid cells [99,104,105,106,107,108,109,110].
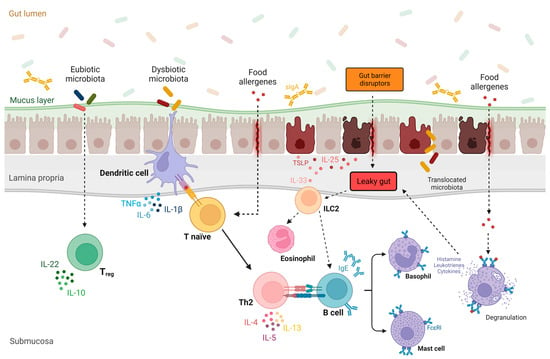
Figure 1.
The role of gut microbiota and leaky gut in the pathogenesis of food allergy. Under physiological conditions, a eubiotic gut microbiota promotes the differentiation of T lymphocytes into T regulatory cells, leading to immune tolerance of the gut [96]. Conversely, in the context of a leaky gut barrier and intestinal dysbiosis, epithelial-derived cytokines such as thymic stromal lymphopoietin (TSLP), interleukin 33 (IL-33), and IL-25 are released in response to various gut barrier disruptors [97,98]. These alarmins promote a pro-allergic microenvironment by activating T helper 2 (Th2) and type 2 innate lymphoid cells (ILC2), resulting in the release of proinflammatory cytokines (e.g., IL-4, IL-5, and IL-13) [95]. Furthermore, dysbiotic gut microbiota induce the differentiation of Th2 cells, promoting the IgE class-switching process in B cells [100]. After sensitization to a specific food allergen, which is also favored by leaky gut, allergen-specific IgE antibodies become immobilized on the surface of basophils and mast cells. Upon subsequent exposure to the allergen, these cells release histamine and other proinflammatory mediators (e.g., leukotrienes and type 2 cytokines) [101,102], worsening gut permeability and amplifying type 2 inflammation [103]. Abbreviations: FcεRI, high-affinity IgE receptor; IL, interleukin; IgE, immunoglobulin E; ILC2, innate lymphoid cell2; Th0, naive T cell; Th2, helper T cell 2; Treg, T regulatory cell; TLSP, thymic stromal lymphopoietin; sIgA, secretory IgA.
Several studies in mice and humans have linked increased intestinal barrier permeability to FA [111,112,113,114,115,116]. This increased permeability allows the translocation of allergenic molecules, such as dietary proteins, toxins, and microbial products, across the intestinal epithelium, where they can interact with immune cells in the gut-associated lymphoid tissue [117]. As a result, an aberrant immune response is triggered, leading to the production of allergen-specific immunoglobulin E (IgE) antibodies and subsequent activation of mast cells and basophils. Upon re-exposure to the allergen, IgE antibodies bind to the surface of mast cells and basophils, releasing proteases and inflammatory mediators, such as histamine [95,118]. This immunological response further impacts intestinal permeability, and the increase in allergen passage perpetuates the inflammatory response through immediate hypersensitivity reactions. Indeed, intestinal permeability may increase when intestinal mucosal barrier function is impaired during allergy [119,120,121].
A breach in the epithelial barrier may result from Th2 immune responses or exaggerated reactions caused by mast cell activation. Type-2 cytokines, such as IL-4 and IL-13, have been shown to directly affect the permeability of intestinal epithelial cells [86,122,123]. These cytokines reduce transepithelial resistance and increase the movement of macromolecules across the epithelium [86,122]. They also influence the expression of tight junction proteins, leading to enhanced permeability [124]. In sensitized mice, ovalbumin (OVA) downregulates the expression of zonulin (ZO)-1, which plays an important role in the regulation of tight junction permeability [125,126].
When the intestinal barrier becomes compromised and “leaky”, it allows increased permeability of the gut epithelial layer [127]. This means that allergenic proteins in ingested food have a higher chance of crossing this compromised barrier and entering the bloodstream. Several studies have demonstrated that individuals with FA have a distinctive transcellular facilitated transport route for the transportation of allergens [127]. This protein uptake has been demonstrated to be specific and exclusively transcellular, occurring within minutes of the allergen exposure and facilitated by the IgE/CD23 complex [128,129,130]. In contrast, healthy individuals primarily rely on transcytosis through enterocytes as the main pathway for protein uptake [131,132]. Moreover, it has been observed that, in FA individuals, the release of mediators from mast cells leads to an augmented allergen transport via the paracellular route [127].
Psychological stress and mast cell activity may also play a role in gut barrier defects and intestinal sensitization [133]. However, it is still unclear whether intestinal barrier dysfunction is the main initiating factor for FA sensitization [134]. A recent study indicates that a gastrointestinal bacterial infection can potentially disrupt oral tolerance to dietary antigens in mice [135]. This disruption can trigger an adaptive immune response against food antigens, resulting in increased intestinal permeability and abnormal pain signaling upon subsequent exposure to the same antigens [135]. Further research using experimental models and antigens with protease activity is needed to determine the order of events between permeability changes and allergic sensitization.
Emerging evidence suggests that compositional and functional changes to the gut microbiome, also known as dysbiosis, contribute to the development and progression of FA, as happens for other diseases [82,136,137,138]. Early studies using germ-free mice have provided initial insights into the significance of the gut microbiota in modulating FA. These studies demonstrated that germ-free mice were unable to achieve oral tolerance to food allergens, and successful induction of oral tolerance through intestinal microbiota reconstitution was only feasible in neonatal mice [139]. Recent studies have further shown that transferring gut microbiota from patients with FA to germ-free mice can transmit susceptibility to FA [140,141]. On the other hand, germ-free mice colonized with bacteria from healthy, but not FA, infants were protected against anaphylactic responses to a cow milk allergen [141]. Multiple microbial orders, including Clostridiales and Lactobacillales, have been implicated in FA, demonstrating beneficial effects. Similarly, Bacteroidales and Enterobacteriales have been described to have both beneficial and detrimental effects [142]. Observational cohort studies involving humans have also identified differences in gut microbiota composition between individuals with and without FA, suggesting that distinct microbiota profiles may have varying effects on food allergen tolerance [143,144,145]. Moreover, direct profiling of the gut microbiome in these studies has shown that dysbiosis, characterized by microbial imbalance, occurs before the onset of FA in affected individuals.
Microbiota–host interactions play a key role in regulating the immune system. The interplay between the gut microbiome and the development of FA is thought to occur through the immunomodulatory effects of microbes on food allergen tolerance [146]. The gut microbiome promotes tolerance by inducing the retinoic orphan receptor gamma T (ROR-γt)+ regulatory T-cells, which can be facilitated by the microbial production of short-chain fatty acids, including butyrate [147]. Furthermore, the intestinal microbiota may prevent allergic sensitization to food antigens through stimulating the production of IL-22 by immune cells in the gut, resulting in improved intestinal epithelial integrity and reduced interaction of the immune system with the allergen [148].
Early-life insults to the gut microbiota impair the differentiation of ROR-γt+ regulatory T (Treg) cells and disrupt the development of oral tolerance to food antigens. Notably, this dysregulated immune response is characterized by decreased IgA binding and increased IgE binding to the gut microbiota, indicating impaired anti-microbial antibody responses. Moreover, T follicular helper cell (Tfh) responses to food antigens develop, thereby facilitating the production of high-affinity IgE antibodies that can trigger anaphylactic reactions.
Understanding the intricate interactions between leaky gut, gut microbiome, and FA has significant implications for prevention and treatment strategies. Therapeutic interventions that restore gut barrier integrity and rebalance the gut microbiota composition and functionality hold promise in ameliorating FA symptoms. Approaches such as the use of specific probiotic strains, prebiotics, synbiotics, postbiotics, and dietary modifications have shown potential in preclinical and clinical studies by promoting a healthy gut microbiota, enhancing gut barrier function, and modulating immune responses, thereby reducing the risk of FA and improving the management of existing allergies.
2. Therapeutic Interventions to Restore Intestinal Barrier Integrity and Microbiome in Food Allergies
2.1. Diet, Prebiotics, and Short-Chain Fatty Acids
Prebiotics usually comprise oligosaccharides and short-chain polysaccharides, serving as nutritional substrates for beneficial gut microbial bacteria, promoting their expansion [149]. Fructans, specifically inulin and fructooligosaccharides (FOS), and galactans such as galactooligosaccharides (GOS) are the most studied prebiotics, since they are found in foods such as some vegetables (garlic, chicory, onion, artichoke, and asparagus), banana, and cereals such as rye and corn.
Human milk also comprises prebiotics named human milk oligosaccharides (HMOs). One- to two-week interventions with supplementation of inulin [150,151,152,153], FOS [154], or GOS [154,155,156,157] were significantly associated with an increase in abundance of Bifidobacterium, as well as Lactobacillus, Akkermansia, or Roseburia. Bifidobacterium spp. are the main commensals for HMO degradation [158]. Maternal prebiotic supplementation during pregnancy to prevent allergy in children seems to be the best window of opportunity to modulate gut microbiota composition and functions [159].
Gut bacteria ferment prebiotics and induce the production and release of short-chain fatty acids (SCFAs) in the lumen [50,160]. SCFAs can specifically interact with intestinal epithelial cells and innate/adaptive immune cells to influence cellular differentiation, proliferation, and apoptosis [161]. SCFAs induce signaling pathways through the stimulation of the production of protein kinases [162], mammalian target of rapamycin (mTOR) [163], and nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB) [164], while SCFAs also modulate the function of histone acetyltransferase (HATs) and histone deacetylases (HDACs) involved in the regulation of gene expression [165]. In preclinical studies, butyrate significantly reduced food allergic response, with induction of tolerogenic cytokines, inhibition of Th2 cytokine production, and modulation of oxidative stress [166]. A receptor for butyrate and niacin (GPR109A) may have a crucial role in maintaining epithelial function and may represent a negative regulator of immune responses [167]. Recently, Wang et al. showed that treatment with butyrate-releasing micelles increased the abundance of butyrate-producing taxa in Clostridium cluster XIVa and protected mice from an anaphylactic reaction to peanut challenge [168].
Interestingly, oral immunotherapy and FOS treatment in mice induce potential microbial alterations associated with increased butyrate levels in cecum content, representing promising strategies to improve oral immunotherapy efficacy in human studies to treat FA [169]. In humans, recent studies suggest that butyrate may directly affect mast cells by epigenetically regulating the FcεRI-mediated signaling molecules [170]. Thus, butyrate could have therapeutic benefits in human FA by inhibiting IgE-mediated mast cell degranulation and allergen-induced histamine release [171]. Moreover, in human enterocytes, butyrate has been shown to stimulate mucin production, tight junctions, and human beta-defensin-3 expression. In peripheral blood mononuclear cells from children with FA, butyrate enhanced IL-10, IFN-γ, and Forkhead box P3 (FOXP3) expression through epigenetic mechanisms [166]. Moreover, it promoted dendritic cells, regulatory T cells (Tregs), and the precursors of M2 macrophages [166].
All these recent findings highlight the critical role of prebiotics and SCFAs in promoting the integrity of the epithelial barrier, oral tolerance, and protection against FA. These observations could, at least in part, be explained by the inhibitory effects of SCFAs on HDACs in various immune cells, such as Tregs, B cells, and mast cells, as well as via stimulation of various SCFA receptors. However, recent studies [172,173] showed that prebiotic supplementation (especially inulin) at a high dose exacerbated the allergic inflammation in the context of excess accumulation of SCFAs and microbiota dysbiosis, emphasizing the importance of personalized use of prebiotics to safely alleviate FA. Indeed, interspecific interactions between gut microbial bacteria remain unclear and should be considered. The possible harmful effects of excessive proliferation of high-SCFA-producing bacteria should not be neglected.
2.2. Probiotics, Next-Generation Probiotics, and Postbiotics
Probiotics are live microorganisms that may benefit the host’s health when administered in adequate amounts [174]. Even if this term also includes yeasts, such as Saccharomyces boulardii [175], the most important probiotics studied in FA are bacteria falling into two genera, namely Lactobacillus and Bifidobacterium. The exact mechanism of action is still little understood; however, probiotic supplementation may prevent or reduce FA symptoms in several ways. First, they inhibit the blooming of pathogenic microorganisms competing with them for nutrients and producing antimicrobial metabolites; they adhere to intestinal epithelial cells through surface glycoprotein or polysaccharides, hampering the binding of pathogenic bacteria to epithelial cells. In addition, lactic acid bacteria (LAB) and Bifidobacteria contribute to maintaining a low oxygen tension and an acid gut environment, as an ideal habitat for beneficial species [176].
For this purpose, supplementation with L. rhamnosus GG (LGG) during a dietary intervention with extensively hydrolyzed casein formula (EHCF) in infants affected by cow milk allergy (CMA) was associated with the expansion of Blautia, Roseburia, and other butyrate-producing taxa and with an increase in fecal butyrate levels [177]. Probiotics enhance the gut barrier, producing both SCFAs as their main metabolites (see above) and increasing the expression of mucins (MUC2, MUC3, and MUC5AC) by intestinal cells [178]; hence, they enforce the expression of tight junction proteins such as occludins and zonulin protein (Zonula Occludens-1, ZO-1) [179]. In this way, they reduce epithelial barrier derangement, which is the main cause of bacterial and peptide translocation into the inner barrier, leading to microinflammation and immunization (the so-called “epithelial barrier hypothesis” of the onset of allergic diseases) [176].
As the basis of most food allergic reactions, there is an immune imbalance during the gut-associated lymphoid tissue (GALT) development toward a Th-2 phenotype. Whereas, in utero, the Th2 pattern is predominant to prevent maternal rejection, in the early phases of post-natal life, microbial stimulation, including vaginal bacteria such as Lactobacilli, reprograms the immune system towards a Th1 profile [16]. As demonstrated in a murine model of FA to ovalbumin (OVA), cesarean section (CS) yields a lack of Lactobacillus and Bifidobacterium strains in the feces and worsens the severity of allergic symptoms. Probiotic supplementation, based on a mixture of L. acidophilus, B. longum subsp. Infantis, E.faecalis, and B.cereus from birth to day 21 could rescue CS-related dysbiosis, improve allergic symptoms, reduce IgE and IgG production, and increase the expression of tight junction protein such as ZO-1, occluding, and claudin-1 [180]. These results are in line with those obtained in other animal studies, showing that new-born mice in antibiotic therapy and germ-free mice maintain a Th2 immune phenotype and an increased risk of allergic reactions to oral antigens. This risk is often corrected by probiotic supplementation in early life, which restores the Th1/Th2 balance, favoring a Th1 pattern [181,182,183].
Such results have been confirmed in the last two decades by several human studies, both retrospective and prospective (non-randomized and randomized trials), mostly in the management of CMA [176]. Most of the research has focused on L. rhamnosus GG, L. casei, L. plantarum, B.lactis, B. bifidum, B. breve, and included infants affected by CMA and orally supplemented with probiotics over a period ranging from at least 1 week [184] to 36 months [185]. Depending on the type of study, children were then followed for a period ranging from 72 h [184] to 5 years [186]. These studies had different endpoints, from reductions in daily vomiting and diarrhea in the early phases of supplementation [184] to the achievement of clinical food tolerance at 12 months [187]. Some of them used a unique strain such as L. rhamnosus GG [188] or a mixture of L. rhamnosus GG and B. lactis [187]. Others also included prebiotics like inulin and oligofructose [189]. Almost all these studies confirmed a significant impact of probiotic supplementation in ameliorating clinical features and reducing allergic reactions in infants affected by CMA, thus showing possible benefits. However, current evidence does still not support a clear recommendation on the use of probiotics in the prevention or treatment of FA by scientific societies, lacking clear information on the specific strain, dosage, and adequate duration of therapy [16].
A possible novel field of interest is oral food immunotherapy (OIT), based on the idea that progressive and continued oral or intestinal exposure to dietary antigens may lead to food tolerance [190]. In this concept, probiotics may figure as immune response modifiers (IMR), a class of compounds of microbial origin (bacteria themselves) that modulate immune response, together with antigen-presenting cells [16]. In a mice model of egg allergy, animals fed with L. casei variety rhamnosus (Lcr35) had a reduced anaphylactic response during OIT compared to controls (no OIT or only OIT) [191]. A randomized control trial in children affected by peanut allergy showed sustained unresponsiveness in patients treated with probiotics (L. rhamnosus CGMCC 1.3724) and peanut OIT (82.1%) compared to the placebo group (only peanut OIT) [192]. However, a recent multicenter randomized trial did not confirm these findings, showing similar rates of sustained unresponsiveness in both OIT and OIT + probiotic (L. rhamnosus ATCC 53103) supplementation [193].
Next-generation probiotics (NGP) are an emerging field of research in FA treatment. Differently from the traditional types, these microorganisms have been determined using next-generation sequencing techniques and bioinformatics methods. NGP may also include postbiotics, intended as “non-viable bacterial products or metabolic products from microorganisms that have biologic activity in the host” [194]. These microorganisms may also increase the shelf life of probiotic products and are generally safe when administered to immunocompromised individuals [195].
Akkermansia muciniphila is the prototype of NGP and the first approved by the European Food Safety Authority (EFSA) [196]. Its action is also exerted in non-vital cells through its surface protein (Amuc_1100) and has been extensively documented in diabetes mellitus type-2 and obesity [197]. Evidence shows that A. muciniphila is able to produce SCFAs and stimulate IL-10 synthesis and Treg cell proliferation, hence reinforcing the intestinal barrier [198]. In a murine model of OVA, the supplementation of viable A. muciniphila reduced serum levels of anti-OVA IgE and body weight loss (marker of illness) in allergic mice and reduced intestinal inflammation and leukocyte infiltration compared to controls. Furthermore, the inactivated bacteria significantly reduced anti-OVA IgE levels and eosinophil recruitment in allergic mice. This also confirmed the role of postbiotics in this setting [199]. Similar results were obtained in the same model through the supplementation of Bifidobacterium longum subsp. longum 51A (BL51A) only as viable cells, with the results depending on the dose and viability of the bacteria. In this case, an increase in the anti-inflammatory IL-10 was obtained. However, treatment with inactivated bacteria had no beneficial effects [200].
All these results confirm the potential role of probiotics and postbiotics in the management of FA (Table 1). However, further studies on the types of strain, dosage, and duration of treatment are still warranted to consider them as part of therapy.

Table 1.
Potential roles of prebiotics/probiotics in alleviating or preventing food allergies.
2.3. Fecal Microbiota Transplantation
Fecal microbiota transplantation (FMT) is defined as the infusion of stool from a healthy donor to the gastrointestinal (GI) tract of a recipient patient, with the aim to modifying the gut microbiota and restoring the condition of dysbiosis [201]. FMT can be performed via colonoscopy, oral capsules, or nasogastric tube [202]. Nowadays, FMT has proven its efficacy as a potential alternative to antibiotics in Clostridioides difficile infections, and it has also been integrated into international guidelines [203]. An increased body of evidence suggests the use of FMT in other disorders, including GI disorders (i.e., IBD and irritable bowel syndrome) [204], liver disease [205], metabolic disorders [206], and malignancies [207].
FMT has been proposed as a treatment in FA, due to its ability to modulate gut microbiota. This proposal stems from studies that have shed light on the role of different gut microbiota compositions in these patients [208].
Preliminary preclinical evidence suggests a potential role for FMT in FA. Two papers reported a decrease in the core body temperature in mice receiving FMT from FA-affected patients, while only a temporary drop was observed for mice receiving FMT from healthy donors, indicating a protective role against anaphylaxis after oral food challenge [140,141]. Moreover, higher fecal scores and increased GI symptoms (scratching, diarrhea, etc.) were observed in mice who were infused feces from FA when compared with the healthy control mice, when they underwent oral allergic sensitization [209]. Additionally, lower alpha and beta diversity; decreases in Bacteroidaceae and Lachnospiraceae families; and increases in Clostridiaceae, Enterobacteriaceae, and Bifidobacteriaceae were observed among mice receiving FMT from patients with FA and from healthy donors [210].
The emerging role of FMT in FA and the findings obtained in mice models have opened the road to new studies in humans. A Phase I open-label trial to evaluate the safety and tolerability of oral encapsulated FMT administered over 2 days for the treatment of peanut allergy in 10 adult subjects (18–40 years) is now closed, but the data have not been published yet [211]. Moreover, a phase II randomized double-blind placebo-controlled trial to evaluate the safety and tolerability of oral encapsulated FMT in 24 peanut allergic patients (12–17 years) is ongoing [212].
3. Conclusions
Increasing evidence, as reviewed in this manuscript, clearly supports a role for gut microbiota imbalance and disruption of the intestinal barrier in the pathogenesis of FA.
These pathogenic pathways are elicited by environmental factors, including industrialization or unbalanced diet (i.e., with the consumption of highly processed food), that may trigger Th2 immune responses and increase the susceptibility to allergic sensitization.
Preliminary evidence from preclinical and clinical experiences supports the potential use of various modulators of gut microbiome, including diet, prebiotics, probiotics, postbiotics, and FMT, as potential therapeutic tools for FA, although further well-designed studies are needed to confirm these insights.
Author Contributions
R.P., W.F. and G.I. conceived this review article. R.P., W.F., E.R., M.C., F.K., P.R., C.C., M.C.M., G.V., A.G., G.C. and G.I. wrote the initial draft of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The staff of the Fondazione Policlinico Gemelli IRCCS thank the Fondazione Roma for their invaluable support of their scientific research. This work was supported by the Ricerca Finalizzata Giovani Ricercatori 2018 of the Italian Ministry of Health (project GR-2018-12365734) to G.I.; by a BIOMIS grant of the Italian Ministry of Research to A.G., G.C., and G.I. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflicts of Interest
A.G. reports personal fees for consultancy for Eisai S.r.l., 3PSolutions, Real Time Meeting, Fondazione Istituto Danone, Sinergie S.r.l. Board MRGE and SanofiS.p.A; personal fees for acting as a speaker for Takeda S.p.A, AbbVie and Sandoz S.p.A; and personal fees for acting on advisory boards for VSL3 and Eisai. G.I. has received personal fees for acting as a speaker for Alfa Sigma, Biocodex, Illumina, Malesci, Sofar, and Tillotts Pharma, and for acting as a consultant/advisor for Biocodex, Malesci, and Tillots Pharma. The other authors have no potential competing interests to disclose.
References
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cell. Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Salazar, N.; González, S.; Nogacka, A.M.; Rios-Covián, D.; Arboleya, S.; Gueimonde, M.; de los Reyes-Gavilán, C.G. Microbiome: Effects of Ageing and Diet. Curr. Issues Mol. Biol. 2020, 36, 33–62. [Google Scholar] [CrossRef] [PubMed]
- Lange, K.; Buerger, M.; Stallmach, A.; Bruns, T. Effects of Antibiotics on Gut Microbiota. Dig. Dis. 2016, 34, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Bibbò, S.; Ianiro, G.; Giorgio, V.; Scaldaferri, F.; Masucci, L.; Gasbarrini, A.; Cammarota, G. The role of diet on gut microbiota composition. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4742–4749. [Google Scholar] [PubMed]
- Rinninella, E.; Tohumcu, E.; Raoul, P.; Fiorani, M.; Cintoni, M.; Mele, M.C.; Cammarota, G.; Gasbarrini, A.; Ianiro, G. The role of diet in shaping human gut microbiota. Best Pract. Res. Clin. Gastroenterol. 2023, 62–63, 101828. [Google Scholar] [CrossRef] [PubMed]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Di Tommaso, N.; Gasbarrini, A.; Ponziani, F.R. Intestinal Barrier in Human Health and Disease. Int. J. Environ. Res. Public Health 2021, 18, 12836. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Schoeler, M.; Caesar, R. Dietary lipids, gut microbiota and lipid metabolism. Rev. Endocr. Metab. Disord. 2019, 20, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Adolph, T.E.; Dudek, M.; Knolle, P. Non-alcoholic fatty liver disease: The interplay between metabolism, microbes and immunity. Nat. Metab. 2021, 3, 1596–1607. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Nance, K.; Chen, S. The Gut-Brain Axis. Annu. Rev. Med. 2022, 73, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.; Sandhu, K.V.; Bastiaanssen, T.F.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Kreft, L.; Hoffmann, C.; Ohnmacht, C. Therapeutic Potential of the Intestinal Microbiota for Immunomodulation of Food Allergies. Front. Immunol. 2020, 11, 1853. [Google Scholar] [CrossRef]
- Mennini, M.; Arasi, S.; Artesani, M.C.; Fiocchi, A.G. Probiotics in food allergy. Curr. Opin. Allergy Clin. Immunol. 2021, 21, 309–316. [Google Scholar] [CrossRef]
- Fiocchi, A.; Cabana, M.D.; Mennini, M. Current Use of Probiotics and Prebiotics in Allergy. J. Allergy Clin. Immunol. Pract. 2022, 10, 2219–2242. [Google Scholar] [CrossRef]
- Kang, Y.; Cai, Y. Future prospect of faecal microbiota transplantation as a potential therapy in asthma. Allergol. Immunopathol. 2018, 46, 307–309. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Salvo Romero, E.; Alonso Cotoner, C.; Pardo Camacho, C.; Casado Bedmar, M.; Vicario, M. The intestinal barrier function and its involvement in digestive disease. Rev. Esp. Enferm. Dig. 2015, 107, 686–696. [Google Scholar] [CrossRef]
- Pelaseyed, T.; Bergström, J.H.; Gustafsson, J.K.; Ermund, A.; Birchenough, G.M.H.; Schütte, A.; van der Post, S.; Svensson, F.; Rodríguez-Piñeiro, A.M.; Nyström, E.E.L.; et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol. Rev. 2014, 260, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Antoni, L.; Nuding, S.; Weller, D.; Gersemann, M.; Ott, G.; Wehkamp, J.; Stange, E.F. Human colonic mucus is a reservoir for antimicrobial peptides. J. Crohns Colitis 2013, 7, e652–e664. [Google Scholar] [CrossRef]
- Johansson, M.E.V.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.A.; Gyr, K. Non-immunological defence mechanisms of the gut. Gut 1992, 33, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Salim, S.Y.; Söderholm, J.D. Importance of disrupted intestinal barrier in inflammatory bowel diseases. Inflamm. Bowel Dis. 2011, 17, 362–381. [Google Scholar] [CrossRef] [PubMed]
- van der Flier, L.G.; Clevers, H. Stem cells, self-renewal, and differentiation in the intestinal epithelium. Annu. Rev. Physiol. 2009, 71, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Mörbe, U.M.; Jørgensen, P.B.; Fenton, T.M.; von Burg, N.; Riis, L.B.; Spencer, J.; Agace, W.W. Human gut-associated lymphoid tissues (GALT); diversity, structure, and function. Mucosal Immunol. 2021, 14, 793–802. [Google Scholar] [CrossRef]
- Furness, J.B. Types of neurons in the enteric nervous system. J. Auton. Nerv. Syst. 2000, 81, 87–96. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Anderson, J.M.; Van Itallie, C.M. Physiology and function of the tight junction. Cold Spring Harb. Perspect. Biol. 2009, 1, a002584. [Google Scholar] [CrossRef]
- Hartsock, A.; Nelson, W.J. Adherens and tight junctions: Structure, function and connections to the actin cytoskeleton. Biochim. Biophys. Acta 2008, 1778, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.H.; Martin, P.E.M. Gap junctions: Structure and function (Review). Mol. Membr. Biol. 2002, 19, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Freeland, D.M.H.; Nadeau, K.C. Food allergy: Immune mechanisms, diagnosis and immunotherapy. Nat. Rev. Immunol. 2016, 16, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Koplin, J.J.; Mills, E.N.C.; Allen, K.J. Epidemiology of food allergy and food-induced anaphylaxis: Is there really a Western world epidemic? Curr. Opin. Allergy Clin. Immunol. 2015, 15, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A.; Aceves, S.; Bock, S.A.; James, J.; Jones, S.; Lang, D.; Nadeau, K.; Nowak-Wegrzyn, A.; Oppenheimer, J.; Perry, T.T.; et al. Food allergy: A practice parameter update-2014. J. Allergy Clin. Immunol. 2014, 134, 1016–1025.e43. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Holdford, D.; Bilaver, L.; Dyer, A.; Holl, J.L.; Meltzer, D. The economic impact of childhood food allergy in the United States. JAMA Pediatr. 2013, 167, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Anvari, S.; Miller, J.; Yeh, C.Y.; Davis, C.M. IgE-Mediated Food Allergy. Clin. Rev. Allergy Immunol. 2019, 57, 244–260. [Google Scholar] [CrossRef]
- Nowak-Węgrzyn, A.; Katz, Y.; Mehr, S.S.; Koletzko, S. Non-IgE-mediated gastrointestinal food allergy. J. Allergy Clin. Immunol. 2015, 135, 1114–1124. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef]
- McDole, J.R.; Wheeler, L.W.; McDonald, K.G.; Wang, B.; Konjufca, V.; Knoop, K.A.; Newberry, R.D.; Miller, M.J. Goblet cells deliver luminal antigen to CD103+ dendritic cells in the small intestine. Nature 2012, 483, 345–349. [Google Scholar] [CrossRef]
- Mazzini, E.; Massimiliano, L.; Penna, G.; Rescigno, M. Oral tolerance can be established via gap junction transfer of fed antigens from CX3CR1+ macrophages to CD103+ dendritic cells. Immunity 2014, 40, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Hadis, U.; Wahl, B.; Schulz, O.; Hardtke-Wolenski, M.; Schippers, A.; Wagner, N.; Müller, W.; Sparwasser, T.; Förster, R.; Pabst, O. Intestinal tolerance requires gut homing and expansion of FoxP3+ regulatory T cells in the lamina propria. Immunity 2011, 34, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Divekar, R.; Kita, H. Recent advances in epithelium-derived cytokines (IL-33, IL-25, and thymic stromal lymphopoietin) and allergic inflammation. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Sehra, S.; Yao, W.; Nguyen, E.T.; Glosson-Byers, N.L.; Akhtar, N.; Zhou, B.; Kaplan, M.H. TH9 cells are required for tissue mast cell accumulation during allergic inflammation. J. Allergy Clin. Immunol. 2015, 136, 433–440.e1. [Google Scholar] [CrossRef] [PubMed]
- Stone, K.D.; Prussin, C.; Metcalfe, D.D. IgE, mast cells, basophils, and eosinophils. J. Allergy Clin. Immunol. 2010, 125 (Suppl. 2), S73–S80. [Google Scholar] [CrossRef] [PubMed]
- De Martinis, M.; Sirufo, M.M.; Suppa, M.; Ginaldi, L. New Perspectives in Food Allergy. Int. J. Mol. Sci. 2020, 21, 1474. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.A.; Yuen, A.W.T.; Woo, E.; Chu, K.-H.; Kwan, H.-S.; Yang, G.-X.; Yang, Y.; Leung, P.S.C. Microbiota and Food Allergy. Clin. Rev. Allergy Immunol. 2019, 57, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Konya, T.; Guttman, D.S.; Field, C.J.; Sears, M.R.; HayGlass, K.T.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; Becker, A.B.; et al. Infant gut microbiota and food sensitization: Associations in the first year of life. Clin. Exp. Allergy 2015, 45, 632–643. [Google Scholar] [CrossRef]
- Lee, E.; Kim, B.J.; Kang, M.J.; Choi, K.Y.; Cho, H.-J.; Kim, Y.; I Yang, S.; Jung, Y.-H.; Kim, H.Y.; Seo, J.-H.; et al. Dynamics of Gut Microbiota According to the Delivery Mode in Healthy Korean Infants. Allergy Asthma Immunol. Res. 2016, 8, 471–477. [Google Scholar] [CrossRef]
- Fusco, W.; Lorenzo, M.B.; Cintoni, M.; Porcari, S.; Rinninella, E.; Kaitsas, F.; Lener, E.; Mele, M.C.; Gasbarrini, A.; Collado, M.C.; et al. Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients 2023, 15, 2211. [Google Scholar] [CrossRef]
- Roduit, C.; Frei, R.; Ferstl, R.; Loeliger, S.; Westermann, P.; Rhyner, C.; Schiavi, E.; Barcik, W.; Rodriguez-Perez, N.; Wawrzyniak, M.; et al. High levels of butyrate and propionate in early life are associated with protection against atopy. Allergy 2019, 74, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.K.; Macia, L.; Mackay, C.R. Dietary fiber and SCFAs in the regulation of mucosal immunity. J. Allergy Clin. Immunol. 2023, 151, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Cosme-Blanco, W.; Arroyo-Flores, E.; Ale, H. Food Allergies. Pediatr. Rev. 2020, 41, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Schworer, S.A.; Kim, E.H. Sublingual immunotherapy for food allergy and its future directions. Immunotherapy 2020, 12, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Wood, R.A. Oral Immunotherapy for Food Allergy. J. Investig. Allergol. Clin. Immunol. 2017, 27, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Roberts, G.; Sayre, P.H.; Bahnson, H.T.; Radulovic, S.; Santos, A.F.; Brough, H.A.; Phippard, D.; Basting, M.; Feeney, M.; et al. Randomized trial of peanut consumption in infants at risk for peanut allergy. N. Engl. J. Med. 2015, 372, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Abrams, E.M.; Chan, E.S.; Sicherer, S. Peanut Allergy: New Advances and Ongoing Controversies. Pediatrics 2020, 145, e20192102. [Google Scholar] [CrossRef]
- Quigley, E.M.M. Leaky gut—Concept or clinical entity? Curr. Opin. Gastroenterol. 2016, 32, 74–79. [Google Scholar] [CrossRef]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Ghosh, S. Intestinal Barrier Dysfunction, LPS Translocation, and Disease Development. J. Endocr. Soc. 2020, 4, bvz039. [Google Scholar] [CrossRef]
- Poto, R.; Pecoraro, A.; Ferrara, A.L.; Punziano, A.; Lagnese, G.; Messuri, C.; Loffredo, S.; Spadaro, G.; Varricchi, G. Cytokine dysregulation despite immunoglobulin replacement therapy in common variable immunodeficiency (CVID). Front. Immunol. 2023, 14, 1257398. [Google Scholar] [CrossRef]
- Pålsson-McDermott, E.M.; O’Neill, L.A.J. Signal transduction by the lipopolysaccharide receptor, Toll-like receptor-4. Immunology 2004, 113, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [PubMed]
- Schumski, A.; Ortega-Gómez, A.; Wichapong, K.; Winter, C.; Lemnitzer, P.; Viola, J.R.; Pinilla-Vera, M.; Folco, E.; Solis-Mezarino, V.; Völker-Albert, M.; et al. Endotoxinemia Accelerates Atherosclerosis Through Electrostatic Charge-Mediated Monocyte Adhesion. Circulation 2021, 143, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, P.; Su, Q.; Wang, S.; Wang, F. Myosin light chain kinase mediates intestinal barrier disruption following burn injury. PLoS ONE 2012, 7, e34946. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Stärkel, P.; Turner, J.R.; Ho, S.B.; Schnabl, B. Dysbiosis-induced intestinal inflammation activates tumor necrosis factor receptor I and mediates alcoholic liver disease in mice. Hepatology 2015, 61, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Kirby, J.; Reilly, C.M.; Luo, X.M. Leaky Gut As a Danger Signal for Autoimmune Diseases. Front. Immunol. 2017, 8, 598. [Google Scholar] [CrossRef]
- Kong, J.; Zhang, Z.; Musch, M.W.; Ning, G.; Sun, J.; Hart, J.; Bissonnette, M.; Li, Y.C. Novel role of the vitamin D receptor in maintaining the integrity of the intestinal mucosal barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G208–G216. [Google Scholar] [CrossRef]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 2016, 167, 1339–1353.e21. [Google Scholar] [CrossRef]
- Lam, Y.Y.; Ha, C.W.Y.; Campbell, C.R.; Mitchell, A.J.; Dinudom, A.; Oscarsson, J.; Cook, D.I.; Hunt, N.H.; Caterson, I.D.; Holmes, A.J.; et al. Increased gut permeability and microbiota change associate with mesenteric fat inflammation and metabolic dysfunction in diet-induced obese mice. PLoS ONE 2012, 7, e34233. [Google Scholar] [CrossRef]
- Michielan, A.; D’Incà, R. Intestinal Permeability in Inflammatory Bowel Disease: Pathogenesis, Clinical Evaluation, and Therapy of Leaky Gut. Mediat. Inflamm. 2015, 2015, 628157. [Google Scholar] [CrossRef]
- Yu, S.; Sun, Y.; Shao, X.; Zhou, Y.; Yu, Y.; Kuai, X.; Zhou, C. Leaky Gut in IBD: Intestinal Barrier-Gut Microbiota Interaction. J. Microbiol. Biotechnol. 2022, 32, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Zeissig, S.; Bürgel, N.; Günzel, D.; Richter, J.; Mankertz, J.; Wahnschaffe, U.; Kroesen, A.J.; Zeitz, M.; Fromm, M.; Schulzke, J.-D. Changes in expression and distribution of claudin 2, 5 and 8 lead to discontinuous tight junctions and barrier dysfunction in active Crohn’s disease. Gut 2007, 56, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Pullan, R.D.; Thomas, G.A.; Rhodes, M.; Newcombe, R.G.; Williams, G.T.; Allen, A.; Rhodes, J. Thickness of adherent mucus gel on colonic mucosa in humans and its relevance to colitis. Gut 1994, 35, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Cunliffe, R.N.; Rose, F.R.; Keyte, J.; Abberley, L.; Chan, W.C.; Mahida, Y.R. Human defensin 5 is stored in precursor form in normal Paneth cells and is expressed by some villous epithelial cells and by metaplastic Paneth cells in the colon in inflammatory bowel disease. Gut 2001, 48, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Turpin, W.; Lee, S.H.; Raygoza Garay, J.A.; Madsen, K.L.; Meddings, J.B.; Bedrani, L.; Power, N.; Espin-Garcia, O.; Xu, W.; Smith, M.I.; et al. Increased Intestinal Permeability Is Associated With Later Development of Crohn’s Disease. Gastroenterology 2020, 159, 2092–2100.e5. [Google Scholar] [CrossRef] [PubMed]
- Chopyk, D.M.; Grakoui, A. Contribution of the Intestinal Microbiome and Gut Barrier to Hepatic Disorders. Gastroenterology 2020, 159, 849–863. [Google Scholar] [CrossRef]
- Tripathi, A.; Debelius, J.; Brenner, D.A.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut-liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar] [CrossRef]
- Oikonomou, T.; Papatheodoridis, G.V.; Samarkos, M.; Goulis, I.; Cholongitas, E. Clinical impact of microbiome in patients with decompensated cirrhosis. World J. Gastroenterol. 2018, 24, 3813–3820. [Google Scholar] [CrossRef]
- Cirera, I.; Bauer, T.M.; Navasa, M.; Vila, J.; Grande, L.; Taurá, P.; Fuster, J.; García-Valdecasas, J.C.; Lacy, A.; Suárez, M.J.; et al. Bacterial translocation of enteric organisms in patients with cirrhosis. J. Hepatol. 2001, 34, 32–37. [Google Scholar] [CrossRef]
- Lachar, J.; Bajaj, J.S. Changes in the Microbiome in Cirrhosis and Relationship to Complications: Hepatic Encephalopathy, Spontaneous Bacterial Peritonitis, and Sepsis. Semin. Liver Dis. 2016, 36, 327–330. [Google Scholar] [CrossRef]
- Caminero, A.; Verdu, E.F. Celiac disease: Should we care about microbes? Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G161–G170. [Google Scholar] [CrossRef]
- Paray, B.A.; Albeshr, M.F.; Jan, A.T.; Rather, I.A. Leaky Gut and Autoimmunity: An Intricate Balance in Individuals Health and the Diseased State. Int. J. Mol. Sci. 2020, 21, 9770. [Google Scholar] [CrossRef]
- Lin, R.; Zhou, L.; Zhang, J.; Wang, B. Abnormal intestinal permeability and microbiota in patients with autoimmune hepatitis. Int. J. Clin. Exp. Pathol. 2015, 8, 5153–5160. [Google Scholar]
- Fasano, A. Leaky gut and autoimmune diseases. Clin. Rev. Allergy Immunol. 2012, 42, 71–78. [Google Scholar] [CrossRef]
- Ceponis, P.J.; Botelho, F.; Richards, C.D.; McKay, D.M. Interleukins 4 and 13 increase intestinal epithelial permeability by a phosphatidylinositol 3-kinase pathway. Lack of evidence for STAT 6 involvement. J. Biol. Chem. 2000, 275, 29132–29137. [Google Scholar] [CrossRef]
- Akdis, C.A. Does the epithelial barrier hypothesis explain the increase in allergy, autoimmunity and other chronic conditions? Nat. Rev. Immunol. 2021, 21, 739–751. [Google Scholar] [CrossRef]
- Pothoven, K.L.; Schleimer, R.P. The barrier hypothesis and Oncostatin M: Restoration of epithelial barrier function as a novel therapeutic strategy for the treatment of type 2 inflammatory disease. Tissue Barriers 2017, 5, e1341367. [Google Scholar] [CrossRef]
- Celebi Sozener, Z.; Ozdel Ozturk, B.; Cerci, P.; Turk, M.; Akin, B.G.; Akdis, M.; Altiner, S.; Ozbey, U.; Ogulur, I.; Mitamura, Y.; et al. Epithelial barrier hypothesis: Effect of the external exposome on the microbiome and epithelial barriers in allergic disease. Allergy 2022, 77, 1418–1449. [Google Scholar] [CrossRef]
- Ogulur, I.; Yazici, D.; Pat, Y.; Bingöl, E.N.; Babayev, H.; Ardicli, S.; Heider, A.; Rückert, B.; Sampath, V.; Dhir, R.; et al. Mechanisms of gut epithelial barrier impairment caused by food emulsifiers polysorbate 20 and polysorbate 80. Allergy 2023, 78, 2441–2455. [Google Scholar] [CrossRef]
- Paparo, L.; Coppola, S.; Nocerino, R.; Pisapia, L.; Picariello, G.; Cortese, M.; Voto, L.; Maglio, M.; Miele, E.; Carucci, L.; et al. How advanced glycation end products could facilitate the occurrence of food allergy. J. Allergy Clin. Immunol. 2023. [Google Scholar] [CrossRef]
- Varricchi, G.; Ferri, S.; Pepys, J.; Poto, R.; Spadaro, G.; Nappi, E.; Paoletti, G.; Virchow, J.C.; Heffler, E.; Canonica, W.G. Biologics and airway remodeling in severe asthma. Allergy 2022, 77, 3538–3552. [Google Scholar] [CrossRef]
- Poto, R.; Gambardella, A.R.; Marone, G.; Schroeder, J.T.; Mattei, F.; Schiavoni, G.; Varricchi, G. Basophils from allergy to cancer. Front. Immunol. 2022, 13, 1056838. [Google Scholar] [CrossRef]
- Poto, R.; Loffredo, S.; Marone, G.; Di Salvatore, A.; de Paulis, A.; Schroeder, J.T.; Varricchi, G. Basophils beyond allergic and parasitic diseases. Front. Immunol. 2023, 14, 1190034. [Google Scholar] [CrossRef]
- Gambardella, A.R.; Poto, R.; Tirelli, V.; Schroeder, J.T.; Marone, G.; Mattei, F.; Varricchi, G.; Schiavoni, G. Differential Effects of Alarmins on Human and Mouse Basophils. Front. Immunol. 2022, 13, 894163. [Google Scholar] [CrossRef]
- Varricchi, G.; Bencivenga, L.; Poto, R.; Pecoraro, A.; Shamji, M.H.; Rengo, G. The emerging role of T follicular helper (TFH) cells in aging: Influence on the immune frailty. Ageing Res. Rev. 2020, 61, 101071. [Google Scholar] [CrossRef]
- Marcella, S.; Petraroli, A.; Canè, L.; Ferrara, A.L.; Poto, R.; Parente, R.; Palestra, F.; Cristinziano, L.; Modestino, L.; Galdiero, M.R.; et al. Thymic stromal lymphopoietin (TSLP) is a substrate for tryptase in patients with mastocytosis. Eur. J. Intern. Med. 2023, 117, 111–118. [Google Scholar] [CrossRef]
- Varricchi, G.; Pecoraro, A.; Marone, G.; Criscuolo, G.; Spadaro, G.; Genovese, A.; Marone, G. Thymic Stromal Lymphopoietin Isoforms, Inflammatory Disorders, and Cancer. Front. Immunol. 2018, 9, 1595. [Google Scholar] [CrossRef]
- Hammad, H.; Lambrecht, B.N. Barrier Epithelial Cells and the Control of Type 2 Immunity. Immunity 2015, 43, 29–40. [Google Scholar] [CrossRef]
- Campbell, E.; Hesser, L.A.; Nagler, C.R. B cells and the microbiota: A missing connection in food allergy. Mucosal Immunol. 2021, 14, 4–13. [Google Scholar] [CrossRef]
- Poto, R.; Quinti, I.; Marone, G.; Taglialatela, M.; de Paulis, A.; Casolaro, V.; Varricchi, G. IgG Autoantibodies Against IgE from Atopic Dermatitis Can Induce the Release of Cytokines and Proinflammatory Mediators from Basophils and Mast Cells. Front. Immunol. 2022, 13, 880412. [Google Scholar] [CrossRef]
- Poto, R.; Patella, V.; Criscuolo, G.; Marone, G.; Coscioni, E.; Varricchi, G. Autoantibodies to IgE can induce the release of proinflammatory and vasoactive mediators from human cardiac mast cells. Clin. Exp. Med. 2023, 23, 1265–1276. [Google Scholar] [CrossRef]
- Celebi Sözener, Z.; Cevhertas, L.; Nadeau, K.; Akdis, M.; Akdis, C.A. Environmental factors in epithelial barrier dysfunction. J. Allergy Clin. Immunol. 2020, 145, 1517–1528. [Google Scholar] [CrossRef]
- Sy, C.B.; Siracusa, M.C. The Therapeutic Potential of Targeting Cytokine Alarmins to Treat Allergic Airway Inflammation. Front. Physiol. 2016, 7, 214. [Google Scholar] [CrossRef]
- Schmitz, J.; Owyang, A.; Oldham, E.; Song, Y.; Murphy, E.; McClanahan, T.K.; Zurawski, G.; Moshrefi, M.; Qin, J.; Li, X.; et al. IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity 2005, 23, 479–490. [Google Scholar] [CrossRef]
- Kakkar, R.; Lee, R.T. The IL-33/ST2 pathway: Therapeutic target and novel biomarker. Nat. Rev. Drug Discov. 2008, 7, 827–840. [Google Scholar] [CrossRef]
- Fort, M.M.; Cheung, J.; Yen, D.; Li, J.; Zurawski, S.M.; Lo, S.; Menon, S.; Clifford, T.; Hunte, B.; Lesley, R.; et al. IL-25 induces IL-4, IL-5, and IL-13 and Th2-associated pathologies in vivo. Immunity 2001, 15, 985–995. [Google Scholar] [CrossRef]
- Pan, G.; French, D.; Mao, W.; Maruoka, M.; Risser, P.; Lee, J.; Foster, J.; Aggarwal, S.; Nicholes, K.; Guillet, S.; et al. Forced expression of murine IL-17E induces growth retardation, jaundice, a Th2-biased response, and multiorgan inflammation in mice. J. Immunol. 2001, 167, 6559–6567. [Google Scholar] [CrossRef]
- Lee, J.B.; Chen, C.Y.; Liu, B.; Mugge, L.; Angkasekwinai, P.; Facchinetti, V.; Dong, C.; Liu, Y.; Rothenberg, M.E.; Hogan, S.P.; et al. IL-25 and CD4(+) TH2 cells enhance type 2 innate lymphoid cell-derived IL-13 production, which promotes IgE-mediated experimental food allergy. J. Allergy Clin. Immunol. 2016, 137, 1216–1225.e5. [Google Scholar] [CrossRef]
- Moussion, C.; Ortega, N.; Girard, J.P. The IL-1-like cytokine IL-33 is constitutively expressed in the nucleus of endothelial cells and epithelial cells in vivo: A novel “alarmin”? PLoS ONE 2008, 3, e3331. [Google Scholar] [CrossRef]
- Chen, T.; Liu, X.; Li, M.; He, W.; Li, W.; Cao, Y.; Liu, Z. Food allergens affect the intestinal tight junction permeability in inducing intestinal food allergy in rats. Asian Pac. J. Allergy Immunol. 2014, 32, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Van Elburg, R.M.; Heymans, H.S.; De Monchy, J.G. Effect of disodiumcromoglycate on intestinal permeability changes and clinical response during cow’s milk challenge. Pediatr. Allergy Immunol. 1993, 4, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Schrander, J.J.; Unsalan-Hooyen, R.W.; Forget, P.P.; Jansen, J. [51Cr]EDTA intestinal permeability in children with cow’s milk intolerance. J. Pediatr. Gastroenterol. Nutr. 1990, 10, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Troncone, R.; Caputo, N.; Florio, G.; Finelli, E. Increased intestinal sugar permeability after challenge in children with cow’s milk allergy or intolerance. Allergy 1994, 49, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Andre, C.; Andre, F.; Colin, L.; Cavagna, S. Measurement of intestinal permeability to mannitol and lactulose as a means of diagnosing food allergy and evaluating therapeutic effectiveness of disodium cromoglycate. Ann. Allergy 1987, 59, 127–130. [Google Scholar]
- Jackson, P.G.; Lessof, M.H.; Baker, R.W.; Ferrett, J.; MacDonald, D.M. Intestinal permeability in patients with eczema and food allergy. Lancet 1981, 317, 1285–1286. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Poto, R.; Ianiro, G.; Punziano, A.; Marone, G.; Gasbarrini, A.; Spadaro, G. Gut Microbiome and Common Variable Immunodeficiency: Few Certainties and Many Outstanding Questions. Front. Immunol. 2021, 12, 712915. [Google Scholar] [CrossRef]
- Varricchi, G.; Poto, R.; Marone, G.; Schroeder, J.T. IL-3 in the development and function of basophils. Semin. Immunol. 2021, 54, 101510. [Google Scholar] [CrossRef]
- Secondulfo, M.; Iafusco, D.; Carratù, R.; Demagistris, L.; Sapone, A.; Generoso, M.; Mezzogiorno, A.; Sasso, F.; Cartenì, M.; De Rosa, R.; et al. Ultrastructural mucosal alterations and increased intestinal permeability in non-celiac, type I diabetic patients. Dig. Liver Dis. 2004, 36, 35–45. [Google Scholar] [CrossRef]
- Majamaa, H.; Isolauri, E. Evaluation of the gut mucosal barrier: Evidence for increased antigen transfer in children with atopic eczema. J. Allergy Clin. Immunol. 1996, 97, 985–990. [Google Scholar] [CrossRef]
- Caffarelli, C.; Cavagni, G.; Menzies, I.S.; Bertolini, P.; Atherton, D.J. Elimination diet and intestinal permeability in atopic eczema: A preliminary study. Clin. Exp. Allergy 1993, 23, 28–31. [Google Scholar] [CrossRef]
- Berin, M.C.; Yang, P.C.; Ciok, L.; Waserman, S.; Perdue, M.H. Role for IL-4 in macromolecular transport across human intestinal epithelium. Am. J. Physiol. 1999, 276, C1046–C1052. [Google Scholar] [CrossRef]
- Di Leo, V.; Yang, P.C.; Berin, M.C.; Perdue, M.H. Factors regulating the effect of IL-4 on intestinal epithelial barrier function. Int. Arch. Allergy Immunol. 2002, 129, 219–227. [Google Scholar] [CrossRef]
- Heller, F.; Florian, P.; Bojarski, C.; Richter, J.; Christ, M.; Hillenbrand, B.; Mankertz, J.; Gitter, A.H.; Bürgel, N.; Fromm, M.; et al. Interleukin-13 is the key effector Th2 cytokine in ulcerative colitis that affects epithelial tight junctions, apoptosis, and cell restitution. Gastroenterology 2005, 129, 550–564. [Google Scholar] [CrossRef]
- Pizzuti, D.; Bortolami, M.; Mazzon, E.; Buda, A.; Guariso, G.; D’Odorico, A.; Chiarelli, S.; D’Incà, R.; De Lazzari, F.; Martines, D.; et al. Transcriptional downregulation of tight junction protein ZO-1 in active coeliac disease is reversed after a gluten-free diet. Dig. Liver Dis. 2004, 36, 337–341. [Google Scholar] [CrossRef]
- Montalto, M.; Cuoco, L.; Ricci, R.; Maggiano, N.; Vecchio, F.M.; Gasbarrini, G. Immunohistochemical analysis of ZO-1 in the duodenal mucosa of patients with untreated and treated celiac disease. Digestion 2002, 65, 227–233. [Google Scholar] [CrossRef]
- Ballegaard, A.S.R.; Bøgh, K.L. Intestinal protein uptake and IgE-mediated food allergy. Food Res. Int. 2023, 163, 112150. [Google Scholar] [CrossRef]
- Berin, M.C.; Kiliaan, A.J.; Yang, P.C.; Groot, J.A.; Taminiau, J.A.; Perdue, M.H. Rapid transepithelial antigen transport in rat jejunum: Impact of sensitization and the hypersensitivity reaction. Gastroenterology 1997, 113, 856–864. [Google Scholar] [CrossRef]
- Yang, P.C.; Berin, M.C.; Yu, L.C.; Conrad, D.H.; Perdue, M.H. Enhanced intestinal transepithelial antigen transport in allergic rats is mediated by IgE and CD23 (FcepsilonRII). J. Clin. Investig. 2000, 106, 879–886. [Google Scholar] [CrossRef]
- Bevilacqua, C.; Montagnac, G.; Benmerah, A.; Candalh, C.; Brousse, N.; Cerf-Bensussan, N.; Perdue, M.H.; Heyman, M. Food allergens are protected from degradation during CD23-mediated transepithelial transport. Int. Arch. Allergy Immunol. 2004, 135, 108–116. [Google Scholar] [CrossRef]
- Laiping So, A.; Pelton-Henrion, K.; Small, G.; Becker, K.; Oei, E.; Tyorkin, M.; Sperber, K.; Mayer, L. Antigen uptake and trafficking in human intestinal epithelial cells. Dig. Dis. Sci. 2000, 45, 1451–1461. [Google Scholar]
- Warshaw, A.L.; Walker, W.A.; Isselbacher, K.J. Protein uptake by the intestine: Evidence for absorption of intact macromolecules. Gastroenterology 1974, 66, 987–992. [Google Scholar] [CrossRef]
- Ilchmann-Diounou, H.; Menard, S. Psychological Stress, Intestinal Barrier Dysfunctions, and Autoimmune Disorders: An Overview. Front. Immunol. 2020, 11, 1823. [Google Scholar] [CrossRef]
- Jalonen, T. Identical intestinal permeability changes in children with different clinical manifestations of cow’s milk allergy. J. Allergy Clin. Immunol. 1991, 88, 737–742. [Google Scholar] [CrossRef]
- Aguilera-Lizarraga, J.; Florens, M.V.; Viola, M.F.; Jain, P.; Decraecker, L.; Appeltans, I.; Cuende-Estevez, M.; Fabre, N.; Van Beek, K.; Perna, E.; et al. Local immune response to food antigens drives meal-induced abdominal pain. Nature 2021, 590, 151–156. [Google Scholar] [CrossRef]
- Parrish, A.; Boudaud, M.; Grant, E.T.; Willieme, S.; Neumann, M.; Wolter, M.; Craig, S.Z.; De Sciscio, A.; Cosma, A.; Hunewald, O.; et al. Akkermansia muciniphila exacerbates food allergy in fibre-deprived mice. Nat. Microbiol. 2023, 8, 1863–1879. [Google Scholar] [CrossRef]
- Hoskinson, C.; Dai, D.L.Y.; Del Bel, K.L.; Becker, A.B.; Moraes, T.J.; Mandhane, P.J.; Finlay, B.B.; Simons, E.; Kozyrskyj, A.L.; Azad, M.B.; et al. Delayed gut microbiota maturation in the first year of life is a hallmark of pediatric allergic disease. Nat. Commun. 2023, 14, 4785. [Google Scholar] [CrossRef]
- Bunyavanich, S.; Berin, M.C. Food allergy and the microbiome: Current understandings and future directions. J. Allergy Clin. Immunol. 2019, 144, 1468–1477. [Google Scholar] [CrossRef]
- Sudo, N.; Sawamura, S.; Tanaka, K.; Aiba, Y.; Kubo, C.; Koga, Y. The requirement of intestinal bacterial flora for the development of an IgE production system fully susceptible to oral tolerance induction. J. Immunol. 1997, 159, 1739–1745. [Google Scholar] [CrossRef]
- Abdel-Gadir, A.; Stephen-Victor, E.; Gerber, G.K.; Rivas, M.N.; Wang, S.; Harb, H.; Wang, L.; Li, N.; Crestani, E.; Spielman, S.; et al. Microbiota therapy acts via a regulatory T cell MyD88/RORγt pathway to suppress food allergy. Nat. Med. 2019, 25, 1164–1174. [Google Scholar] [CrossRef]
- Feehley, T.; Plunkett, C.H.; Bao, R.; Hong, S.M.C.; Culleen, E.; Belda-Ferre, P.; Campbell, E.; Aitoro, R.; Nocerino, R.; Paparo, L.; et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat. Med. 2019, 25, 448–453. [Google Scholar] [CrossRef]
- Huang, Y.J.; Marsland, B.J.; Bunyavanich, S.; O’Mahony, L.; Leung, D.Y.; Muraro, A.; Fleisher, T.A. The microbiome in allergic disease: Current understanding and future opportunities-2017 PRACTALL document of the American Academy of Allergy, Asthma & Immunology and the European Academy of Allergy and Clinical Immunology. J. Allergy Clin. Immunol. 2017, 139, 1099–1110. [Google Scholar]
- Joseph, C.L.; Sitarik, A.R.; Kim, H.; Huffnagle, G.; Fujimura, K.; Yong, G.J.M.; Levin, A.M.; Zoratti, E.; Lynch, S.; Ownby, D.R.; et al. Infant gut bacterial community composition and food-related manifestation of atopy in early childhood. Pediatr. Allergy Immunol. 2022, 33, e13704. [Google Scholar] [CrossRef]
- Fujimura, K.E.; Sitarik, A.R.; Havstad, S.; Lin, D.L.; Levan, S.; Fadrosh, D.; Panzer, A.R.; LaMere, B.; Rackaityte, E.; Lukacs, N.W.; et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 2016, 22, 1187–1191. [Google Scholar] [CrossRef]
- Bao, R.; Hesser, L.A.; He, Z.; Zhou, X.; Nadeau, K.C.; Nagler, C.R. Fecal microbiome and metabolome differ in healthy and food-allergic twins. J. Clin. Investig. 2021, 131, e141935. [Google Scholar] [CrossRef]
- Rachid, R.; Stephen-Victor, E.; Chatila, T.A. The microbial origins of food allergy. J. Allergy Clin. Immunol. 2021, 147, 808–813. [Google Scholar] [CrossRef]
- Iweala, O.I.; Nagler, C.R. The Microbiome and Food Allergy. Annu. Rev. Immunol. 2019, 37, 377–403. [Google Scholar] [CrossRef]
- Stefka, A.T.; Feehley, T.; Tripathi, P.; Qiu, J.; McCoy, K.; Mazmanian, S.K.; Tjota, M.Y.; Seo, G.-Y.; Cao, S.; Theriault, B.R.; et al. Commensal bacteria protect against food allergen sensitization. Proc. Natl. Acad. Sci. USA 2014, 111, 13145–13150. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef]
- Healey, G.; Murphy, R.; Butts, C.; Brough, L.; Whelan, K.; Coad, J. Habitual dietary fibre intake influences gut microbiota response to an inulin-type fructan prebiotic: A randomised, double-blind, placebo-controlled, cross-over, human intervention study. Br. J. Nutr. 2018, 119, 176–189. [Google Scholar] [CrossRef]
- Hiel, S.; Bindels, L.B.; Pachikian, B.D.; Kalala, G.; Broers, V.; Zamariola, G.; Chang, B.P.I.; Kambashi, B.; Rodriguez, J.; Cani, P.D.; et al. Effects of a diet based on inulin-rich vegetables on gut health and nutritional behavior in healthy humans. Am. J. Clin. Nutr. 2019, 109, 1683–1695. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.B.G.; Elderman, M.E.; El Aidy, S.; Burgerhof, J.G.M.; Visser, H.; Vaughan, E.E.; Faas, M.M.; de Vos, P. Flexibility of Gut Microbiota in Ageing Individuals during Dietary Fiber Long-Chain Inulin Intake. Mol. Nutr. Food Res. 2021, 65, e2000390. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Kolida, S.; Klinder, A.; Gietl, E.; Bäuerlein, M.; Frohberg, C.; Landschütze, V.; Gibson, G.R. A double-blind, placebo-controlled, cross-over study to establish the bifidogenic effect of a very-long-chain inulin extracted from globe artichoke (Cynara scolymus) in healthy human subjects. Br. J. Nutr. 2010, 104, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, P.; Chen, M.; Luo, Y.; Prabhakar, M.; Zheng, H.; He, Y.; Qi, Q.; Long, H.; Zhang, Y.; et al. Fructooligosaccharide (FOS) and Galactooligosaccharide (GOS) Increase Bifidobacterium but Reduce Butyrate Producing Bacteria with Adverse Glycemic Metabolism in healthy young population. Sci. Rep. 2017, 7, 11789. [Google Scholar] [CrossRef] [PubMed]
- Wilms, E.; An, R.; Smolinska, A.; Stevens, Y.; Weseler, A.R.; Elizalde, M.; Drittij, M.-J.; Ioannou, A.; van Schooten, F.J.; Smidt, H.; et al. Galacto-oligosaccharides supplementation in prefrail older and healthy adults increased faecal bifidobacteria, but did not impact immune function and oxidative stress. Clin. Nutr. Edinb. Scotl. 2021, 40, 3019–3031. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Lu, X.; Yang, X. Evaluation of clinical safety and beneficial effects of stachyose-enriched α-galacto-oligosaccharides on gut microbiota and bowel function in humans. Food Funct. 2017, 8, 262–269. [Google Scholar] [CrossRef]
- Vulevic, J.; Juric, A.; Walton, G.E.; Claus, S.P.; Tzortzis, G.; Toward, R.E.; Gibson, G.R. Influence of galacto-oligosaccharide mixture (B-GOS) on gut microbiota, immune parameters and metabonomics in elderly persons. Br. J. Nutr. 2015, 114, 586–595. [Google Scholar] [CrossRef]
- Masi, A.C.; Stewart, C.J. Untangling human milk oligosaccharides and infant gut microbiome. iScience 2022, 25, 103542. [Google Scholar] [CrossRef]
- Selle, A.; Brosseau, C.; Dijk, W.; Duval, A.; Bouchaud, G.; Rousseaux, A.; Bruneau, A.; Cherbuy, C.; Mariadassou, M.; Cariou, V.; et al. Prebiotic Supplementation During Gestation Induces a Tolerogenic Environment and a Protective Microbiota in Offspring Mitigating Food Allergy. Front. Immunol. 2021, 12, 745535. [Google Scholar] [CrossRef]
- Poto, R.; Laniro, G.; de Paulis, A.; Spadaro, G.; Marone, G.; Gasbarrini, A.; Varricchi, G. Is there a role for microbiome-based approach in common variable immunodeficiency? Clin. Exp. Med. 2023, 23, 1981–1998. [Google Scholar] [CrossRef]
- Kim, C.H.; Park, J.; Kim, M. Gut microbiota-derived short-chain Fatty acids, T cells, and inflammation. Immune Netw. 2014, 14, 277–288. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, F.; Wu, W.; Sun, M.; Bilotta, A.J.; Yao, S.; Xiao, Y.; Huang, X.; Eaves-Pyles, T.D.; Golovko, G.; et al. GPR43 mediates microbiota metabolite SCFA regulation of antimicrobial peptide expression in intestinal epithelial cells via activation of mTOR and STAT3. Mucosal Immunol. 2018, 11, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; van Esch, B.C.A.M.; Henricks, P.A.J.; Garssen, J.; Folkerts, G. Time and Concentration Dependent Effects of Short Chain Fatty Acids on Lipopolysaccharide- or Tumor Necrosis Factor α-Induced Endothelial Activation. Front. Pharmacol. 2018, 9, 233. [Google Scholar] [CrossRef] [PubMed]
- van der Hee, B.; Wells, J.M. Microbial Regulation of Host Physiology by Short-chain Fatty Acids. Trends Microbiol. 2021, 29, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Paparo, L.; Nocerino, R.; Ciaglia, E.; Di Scala, C.; De Caro, C.; Russo, R.; Trinchese, G.; Aitoro, R.; Amoroso, A.; Bruno, C.; et al. Butyrate as a bioactive human milk protective component against food allergy. Allergy 2021, 76, 1398–1415. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.L.; Ren, Z.; Tu, X.; Liu, H.; Tay, H.L.; Li, J.; Pan, X.; Dong, X.; Foster, P.S.; Sun, J. GPR109A deficiency promotes IL-33 overproduction and type 2 immune response in food allergy in mice. Allergy 2021, 76, 2613–2616. [Google Scholar] [CrossRef]
- Wang, R.; Cao, S.; Bashir, M.E.H.; Hesser, L.A.; Su, Y.; Hong, S.M.C.; Thompson, A.; Culleen, E.; Sabados, M.; Dylla, N.P.; et al. Treatment of peanut allergy and colitis in mice via the intestinal release of butyrate from polymeric micelles. Nat. Biomed. Eng. 2023, 7, 38–55. [Google Scholar] [CrossRef]
- Vonk, M.M.; Blokhuis, B.R.J.; Diks, M.A.P.; Wagenaar, L.; Smit, J.J.; Pieters, R.H.H.; Garssen, J.; Knippels, L.M.J.; van Esch, B.C.A.M. Butyrate Enhances Desensitization Induced by Oral Immunotherapy in Cow’s Milk Allergic Mice. Mediat. Inflamm. 2019, 2019, 9062537. [Google Scholar] [CrossRef]
- Folkerts, J.; Redegeld, F.; Folkerts, G.; Blokhuis, B.; van den Berg, M.P.M.; de Bruijn, M.J.W.; van IJcken, W.F.J.; Junt, T.; Tam, S.-Y.; Galli, S.J.; et al. Butyrate inhibits human mast cell activation via epigenetic regulation of FcεRI-mediated signaling. Allergy 2020, 75, 1966–1978. [Google Scholar] [CrossRef]
- Fiorani, M.; Del Vecchio, L.E.; Dargenio, P.; Kaitsas, F.; Rozera, T.; Porcari, S.; Gasbarrini, A.; Cammarota, G.; Ianiro, G. Histamine-producing bacteria and their role in gastrointestinal disorders. Expert Rev. Gastroenterol. Hepatol. 2023, 17, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Mu, K.; Chen, C.; Gu, S.; Luo, D.; Fu, W.; Xue, W. The high dose of inulin exacerbated food allergy through the excess accumulation of short-chain fatty acids in a BABL/c mouse model. Int. J. Biol. Macromol. 2023, 230, 123234. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-chain fatty acids activate GPR41 and GPR43 on intestinal epithelial cells to promote inflammatory responses in mice. Gastroenterology 2013, 145, 396–406.e10. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Ianiro, G.; Bruno, G.; Lopetuso, L.; Beghella, F.B.; Laterza, L.; D’Aversa, F.; Gigante, G.; Cammarota, G.; Gasbarrini, A. Role of yeasts in healthy and impaired gut microbiota: The gut mycome. Curr. Pharm. Des. 2014, 20, 4565–4569. [Google Scholar] [CrossRef] [PubMed]
- Cela, L.; Brindisi, G.; Gravina, A.; Pastore, F.; Semeraro, A.; Bringheli, I.; Marchetti, L.; Morelli, R.; Cinicola, B.; Capponi, M.; et al. Molecular Mechanism and Clinical Effects of Probiotics in the Management of Cow’s Milk Protein Allergy. Int. J. Mol. Sci. 2023, 24, 9781. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Sangwan, N.; Stefka, A.T.; Nocerino, R.; Paparo, L.; Aitoro, R.; Calignano, A.; A Khan, A.; A Gilbert, J.; Nagler, C.R. Lactobacillus rhamnosus GG-supplemented formula expands butyrate-producing bacterial strains in food allergic infants. ISME J. 2016, 10, 742–750. [Google Scholar] [CrossRef]
- Otte, J.M.; Podolsky, D.K. Functional modulation of enterocytes by gram-positive and gram-negative microorganisms. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G613–G626. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Jin, B.Y.; Li, Z.; Xia, Y.N.; Li, L.-X.; Zhao, Z.-X.; Li, X.-Y.; Li, B.; Zhou, R.-C.; Fu, S.-C.; Li, S.-Y.; et al. Probiotic Interventions Alleviate Food Allergy Symptoms Correlated With Cesarean Section: A Murine Model. Front. Immunol. 2021, 12, 741371. [Google Scholar] [CrossRef]
- Aitoro, R.; Simeoli, R.; Amoroso, A.; Paparo, L.; Nocerino, R.; Pirozzi, C.; di Costanzo, M.; Meli, R.; De Caro, C.; Picariello, G.; et al. Extensively hydrolyzed casein formula alone or with L. rhamnosus GG reduces β-lactoglobulin sensitization in mice. Pediatr. Allergy Immunol. Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2017, 28, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Schiavi, E.; Barletta, B.; Butteroni, C.; Corinti, S.; Boirivant, M.; Di Felice, G. Oral therapeutic administration of a probiotic mixture suppresses established Th2 responses and systemic anaphylaxis in a murine model of food allergy. Allergy 2011, 66, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Thang, C.L.; Baurhoo, B.; Boye, J.I.; Simpson, B.K.; Zhao, X. Effects of Lactobacillus rhamnosus GG supplementation on cow’s milk allergy in a mouse model. Allergy Asthma Clin. Immunol. 2011, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Ahanchian, H.; Nouri, Z.; Jafari, S.A.; Moghiman, T.; Amirian, M.-H.; Ezzati, A.; Kianifar, H.-R. Synbiotics in Children with Cow’s Milk Allergy: A Randomized Controlled Trial. Iran. J. Pediatr. 2014, 24, 29–34. [Google Scholar] [PubMed]
- Berni Canani, R.; Di Costanzo, M.; Bedogni, G.; Amoroso, A.; Cosenza, L.; Di Scala, C.; Granata, V.; Nocerino, R. Extensively hydrolyzed casein formula containing Lactobacillus rhamnosus GG reduces the occurrence of other allergic manifestations in children with cow’s milk allergy: 3-year randomized controlled trial. J. Allergy Clin. Immunol. 2017, 139, 1906–1913.e4. [Google Scholar] [CrossRef] [PubMed]
- Nocerino, R.; Di Costanzo, M.; Bedogni, G.; Cosenza, L.; Maddalena, Y.; Di Scala, C.; Della Gatta, G.; Carucci, L.; Voto, L.; Coppola, S.; et al. Dietary Treatment with Extensively Hydrolyzed Casein Formula Containing the Probiotic Lactobacillus rhamnosus GG Prevents the Occurrence of Functional Gastrointestinal Disorders in Children with Cow’s Milk Allergy. J. Pediatr. 2019, 213, 137–142.e2. [Google Scholar] [CrossRef] [PubMed]
- Vandenplas, Y.; Steenhout, P.; Planoudis, Y.; Grathwohl, D.; Althera Study Group. Treating cow’s milk protein allergy: A double-blind randomized trial comparing two extensively hydrolysed formulas with probiotics. Acta Paediatr. 2013, 102, 990–998. [Google Scholar] [CrossRef]
- Berni Canani, R.; Nocerino, R.; Terrin, G.; Coruzzo, A.; Cosenza, L.; Leone, L.; Troncone, R. Effect of Lactobacillus GG on tolerance acquisition in infants with cow’s milk allergy: A randomized trial. J. Allergy Clin. Immunol. 2012, 129, 580–582.e1–5. [Google Scholar] [CrossRef]
- Candy, D.C.A.; Van Ampting, M.T.J.; Oude Nijhuis, M.M.; Wopereis, H.; Butt, A.M.; Peroni, D.G.; Vandenplas, Y.; Fox, A.T.; Shah, N.; West, C.E.; et al. A synbiotic-containing amino-acid-based formula improves gut microbiota in non-IgE-mediated allergic infants. Pediatr. Res. 2018, 83, 677–686. [Google Scholar] [CrossRef]
- Berni Canani, R.; Paparo, L.; Nocerino, R.; Di Scala, C.; Della Gatta, G.; Maddalena, Y.; Buono, A.; Bruno, C.; Voto, L.; Ercolini, D. Gut Microbiome as Target for Innovative Strategies Against Food Allergy. Front. Immunol. 2019, 10, 191. [Google Scholar] [CrossRef]
- Kim, B.G.; Kim, J.N.; Jang, A.S.; Shin, M. Combined Effects of Lactobacillus rhamnosus and Egg Oral Immunotherapy in a Mouse Model of Egg Allergy. Allergy Asthma Immunol. Res. 2020, 12, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.L.K.; Ponsonby, A.L.; Orsini, F.; Tey, D.; Robinson, M.; Su, E.L.; Licciardi, P.; Burks, W.; Donath, S. Administration of a probiotic with peanut oral immunotherapy: A randomized trial. J. Allergy Clin. Immunol. 2015, 135, 737–744.e8. [Google Scholar] [CrossRef] [PubMed]
- Loke, P.; Orsini, F.; Lozinsky, A.C.; Gold, M.; O’Sullivan, M.D.; Quinn, P.; Lloyd, M.; E Ashley, S.; Pitkin, S.; Axelrad, C.; et al. Probiotic peanut oral immunotherapy versus oral immunotherapy and placebo in children with peanut allergy in Australia (PPOIT-003): A multicentre, randomised, phase 2b trial. Lancet Child. Adolesc. Health 2022, 6, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Mosca, A.; Abreu YAbreu, A.T.; Gwee, K.A.; Ianiro, G.; Tack, J.; Nguyen, T.V.H.; Hill, C. The clinical evidence for postbiotics as microbial therapeutics. Gut Microbes 2022, 14, 2117508. [Google Scholar] [CrossRef] [PubMed]
- Akter, S.; Park, J.H.; Jung, H.K. Potential Health-Promoting Benefits of Paraprobiotics, Inactivated Probiotic Cells. J. Microbiol. Biotechnol. 2020, 30, 477–481. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Nutrition; Novel Foods and Food Allergens (NDA); Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; et al. Safety of pasteurised Akkermansia muciniphila as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06780. [Google Scholar]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef]
- Miranda, V.C.; Souza, R.O.; Quintanilha, M.F.; Gallotti, B.; Assis, H.C.; Faria, A.M.C.; Nicoli, J.R.; Cara, D.C.; Martins, F.S. A Next-Generation Bacteria (Akkermansia muciniphila BAA-835) Presents Probiotic Potential Against Ovalbumin-Induced Food Allergy in Mice. Probiotics Antimicrob. Proteins 2023. [Google Scholar] [CrossRef]
- Santos, S.S.; Miranda, V.C.; Trindade, L.M.; Cardoso, V.N.; Reis, D.C.; Cassali, G.D.; Nicoli, J.R.; Cara, D.C.; Martins, F.S. Bifidobacterium longum subsp. longum 51A Attenuates Signs of Inflammation in a Murine Model of Food Allergy. Probiotics Antimicrob. Proteins 2023, 15, 63–73. [Google Scholar] [CrossRef]
- Porcari, S.; Benech, N.; Valles-Colomer, M.; Segata, N.; Gasbarrini, A.; Cammarota, G.; Sokol, H.; Ianiro, G. Key determinants of success in fecal microbiota transplantation: From microbiome to clinic. Cell Host Microbe 2023, 31, 712–733. [Google Scholar] [CrossRef] [PubMed]
- Bibbò, S.; Settanni, C.R.; Porcari, S.; Bocchino, E.; Ianiro, G.; Cammarota, G.; Gasbarrini, A. Fecal Microbiota Transplantation: Screening and Selection to Choose the Optimal Donor. J. Clin. Med. 2020, 9, 1757. [Google Scholar] [CrossRef] [PubMed]
- Baunwall, S.M.D.; Terveer, E.M.; Dahlerup, J.F.; Erikstrup, C.; Arkkila, P.; Vehreschild, M.J.; Ianiro, G.; Gasbarrini, A.; Sokol, H.; Kump, P.K.; et al. The use of Faecal Microbiota Transplantation (FMT) in Europe: A Europe-wide survey. Lancet Reg. Health Eur. 2021, 9, 100181. [Google Scholar] [CrossRef] [PubMed]
- Ianiro, G.; Punčochář, M.; Karcher, N.; Porcari, S.; Armanini, F.; Asnicar, F.; Beghini, F.; Blanco-Míguez, A.; Cumbo, F.; Manghi, P.; et al. Variability of strain engraftment and predictability of microbiome composition after fecal microbiota transplantation across different diseases. Nat. Med. 2022, 28, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Mullish, B.H.; Tohumcu, E.; Porcari, S.; Fiorani, M.; Di Tommaso, N.; Gasbarrini, A.; Cammarota, G.; Ponziani, F.R.; Ianiro, G. The role of faecal microbiota transplantation in chronic noncommunicable disorders. J. Autoimmun. 2023, 141, 103034. [Google Scholar] [CrossRef] [PubMed]
- Airola, C.; Severino, A.; Porcari, S.; Fusco, W.; Mullish, B.H.; Gasbarrini, A.; Cammarota, G.; Ponziani, F.R.; Ianiro, G. Future Modulation of Gut Microbiota: From Eubiotics to FMT, Engineered Bacteria, and Phage Therapy. Antibiotics 2023, 12, 868. [Google Scholar] [CrossRef] [PubMed]
- Ianiro, G.; Rossi, E.; Thomas, A.M.; Schinzari, G.; Masucci, L.; Quaranta, G.; Settanni, C.R.; Lopetuso, L.R.; Armanini, F.; Blanco-Miguez, A.; et al. Faecal microbiota transplantation for the treatment of diarrhoea induced by tyrosine-kinase inhibitors in patients with metastatic renal cell carcinoma. Nat. Commun. 2020, 11, 4333. [Google Scholar] [CrossRef]
- Nance, C.L.; Deniskin, R.; Diaz, V.C.; Paul, M.; Anvari, S.; Anagnostou, A. The Role of the Microbiome in Food Allergy: A Review. Children 2020, 7, 50. [Google Scholar] [CrossRef]
- Mauras, A.; Wopereis, H.; Yeop, I.; Esber, N.; Delannoy, J.; Labellie, C.; Reygner, J.; Kapel, N.; Slump, R.; van Eijndthoven, T.; et al. Gut microbiota from infant with cow’s milk allergy promotes clinical and immune features of atopy in a murine model. Allergy 2019, 74, 1790–1793. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, S.; Yang, X.; Huazeng, B.; Cheng, Q. Influences of non-IgE-mediated cow’s milk protein allergy-associated gut microbial dysbiosis on regulatory T cell-mediated intestinal immune tolerance and homeostasis. Microb. Pathog. 2021, 158, 105020. [Google Scholar] [CrossRef]
- Rachid, R. A Phase I Open Label Trial to Evaluate the Safety and Efficacy of Oral Encapsulated Fecal Microbiota Transplantation in Peanut Allergic Patients. clinicaltrials.gov. 2021. Available online: https://clinicaltrials.gov/study/NCT02960074 (accessed on 1 January 2023).
- Rachid, R. A Phase II Trial to Evaluate the Safety and Efficacy of Oral Encapsulated Microbiota Transplantation Therapy in Peanut Allergic Patients. clinicaltrials.gov. 2023. Available online: https://clinicaltrials.gov/study/NCT05695261 (accessed on 1 January 2023).
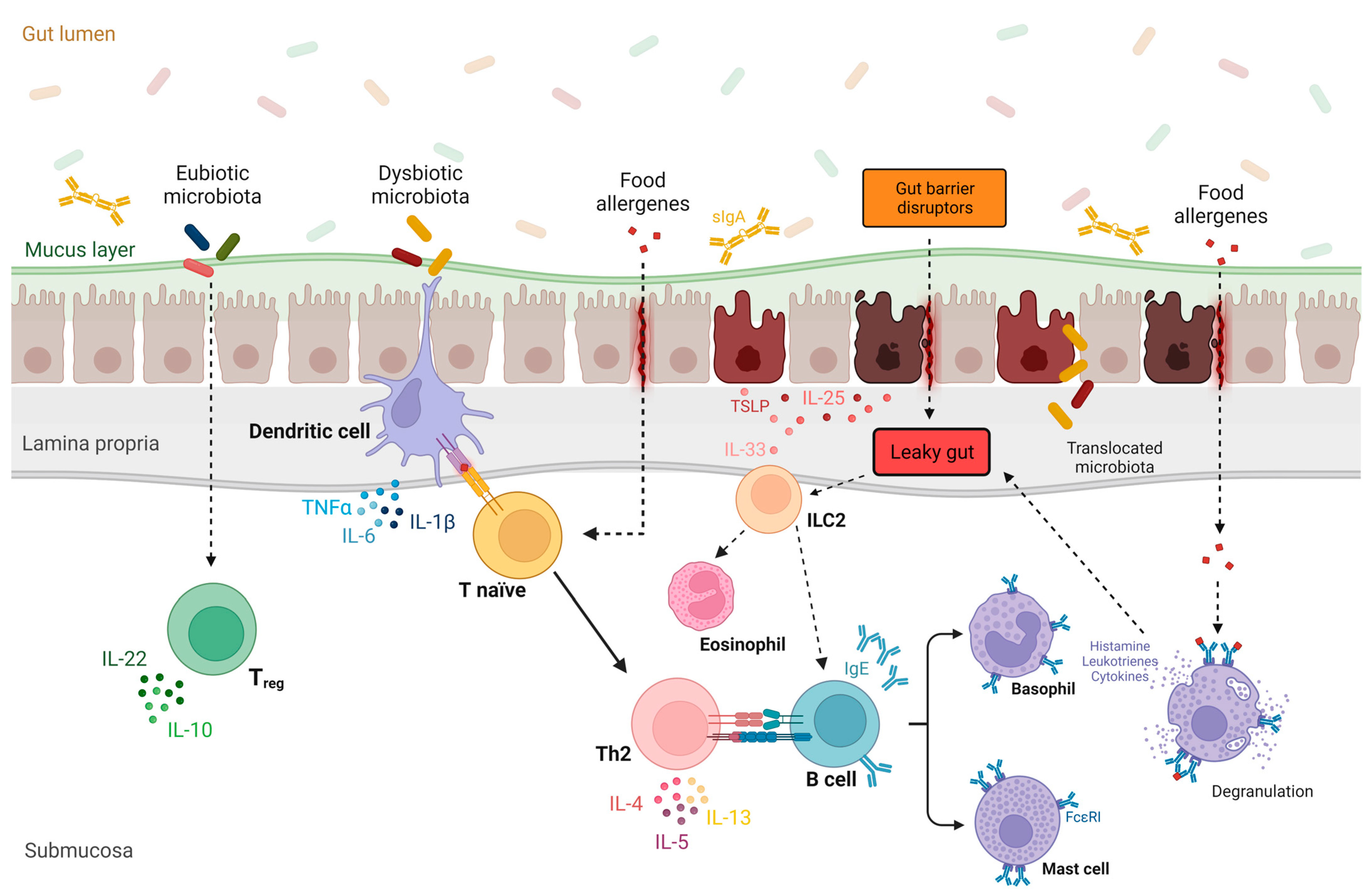
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).