


Gut Microbiota in Patients with Prediabetes


 ,
,
Abstract
1. Introduction
2. Methods
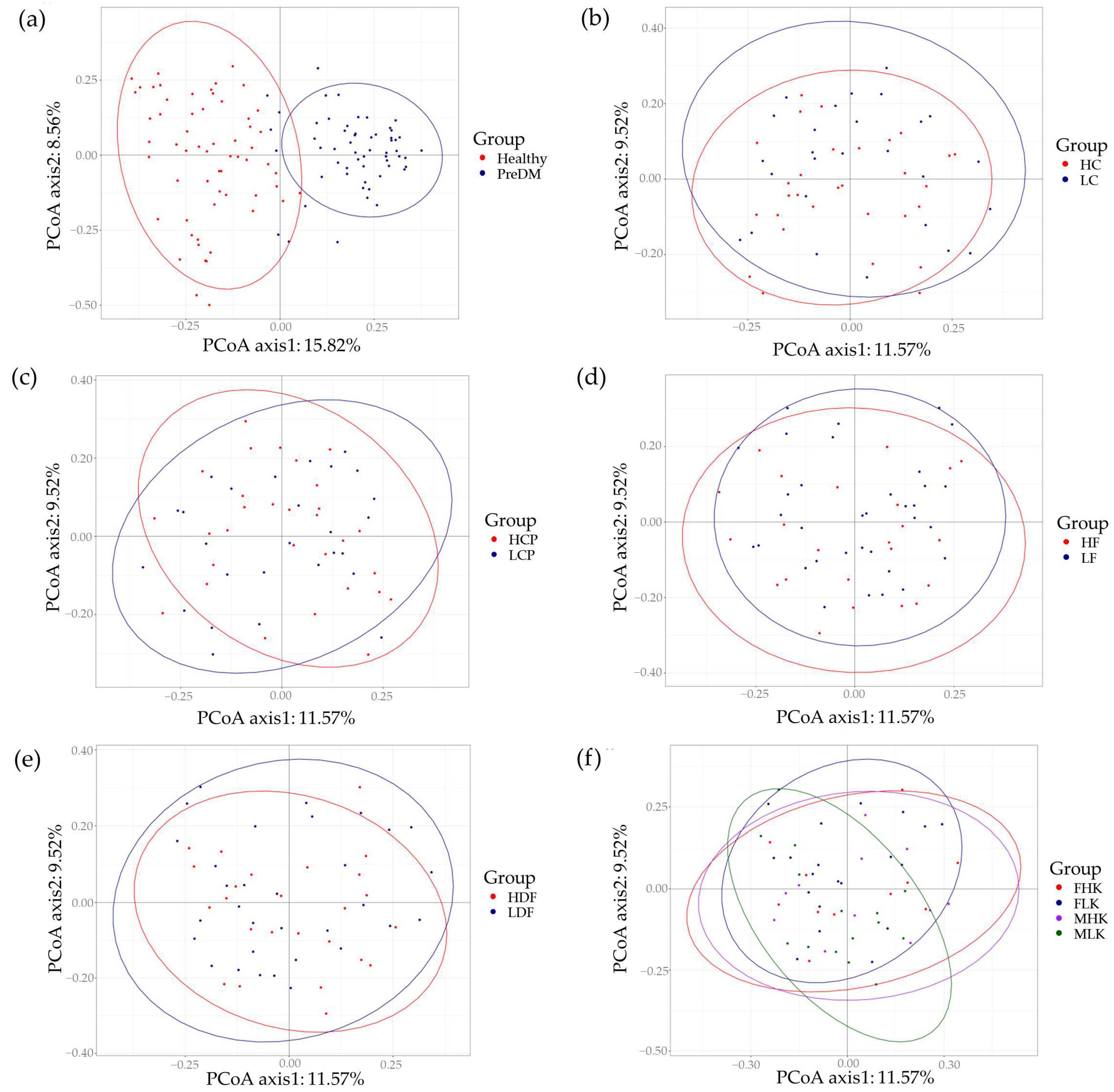
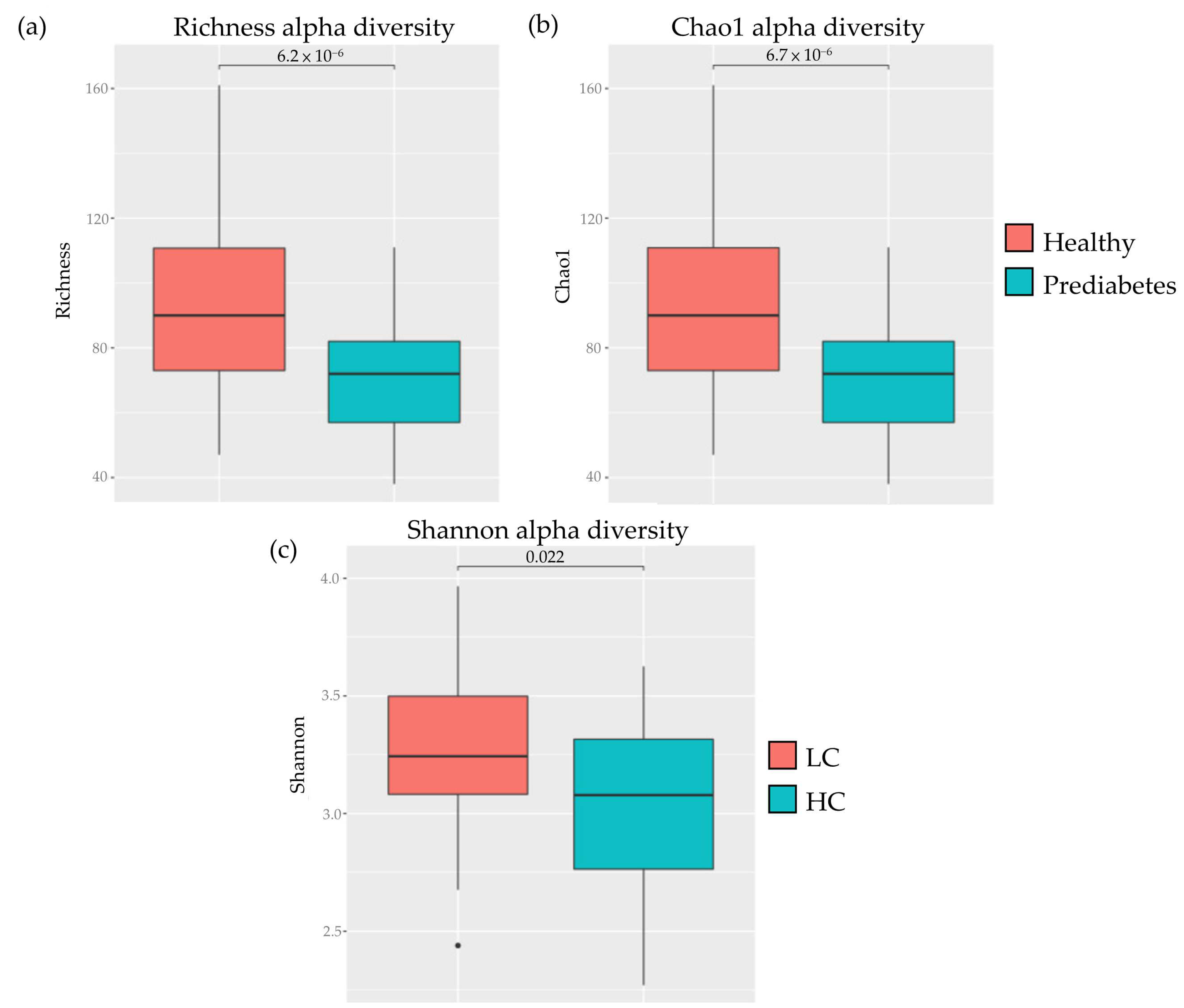
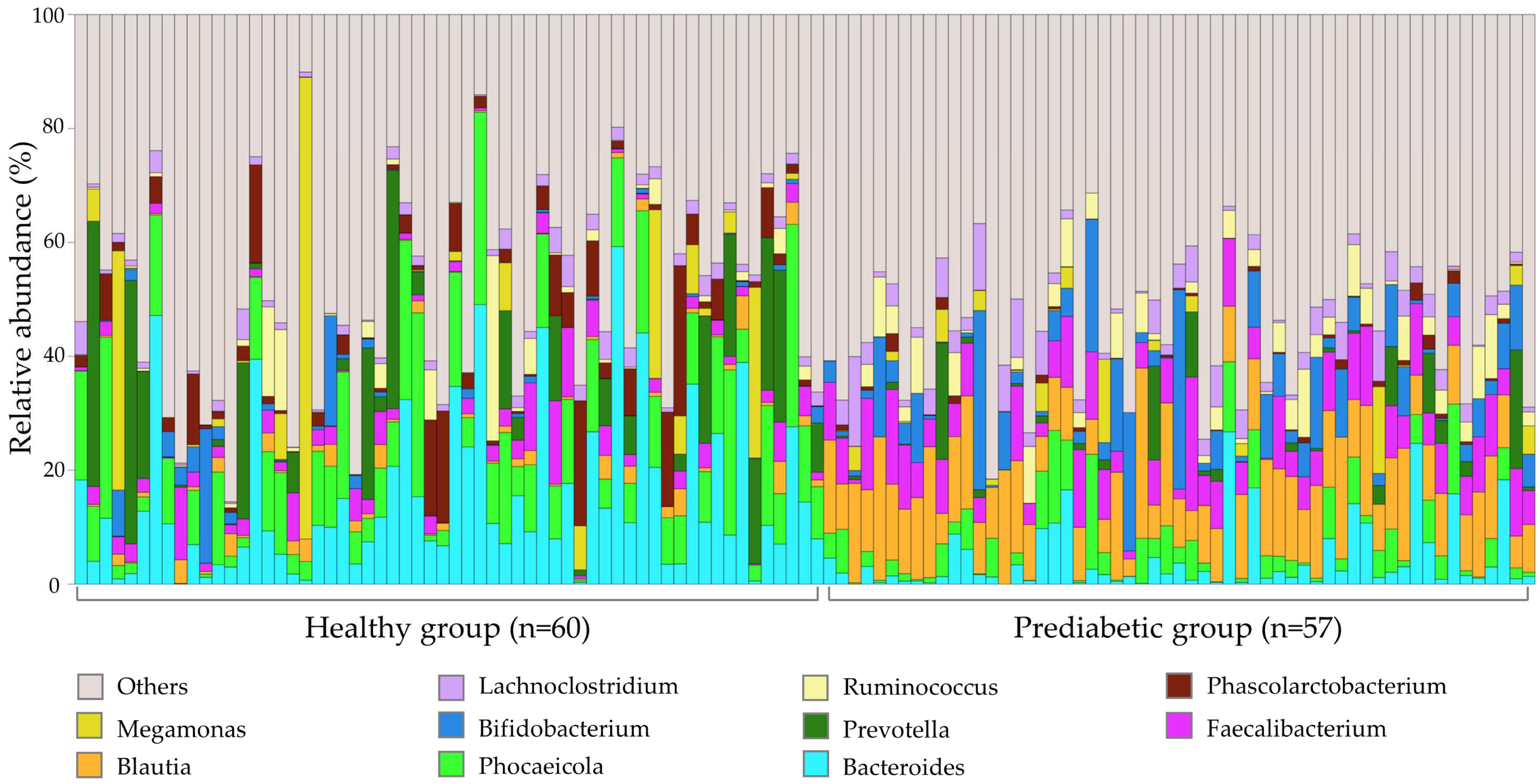
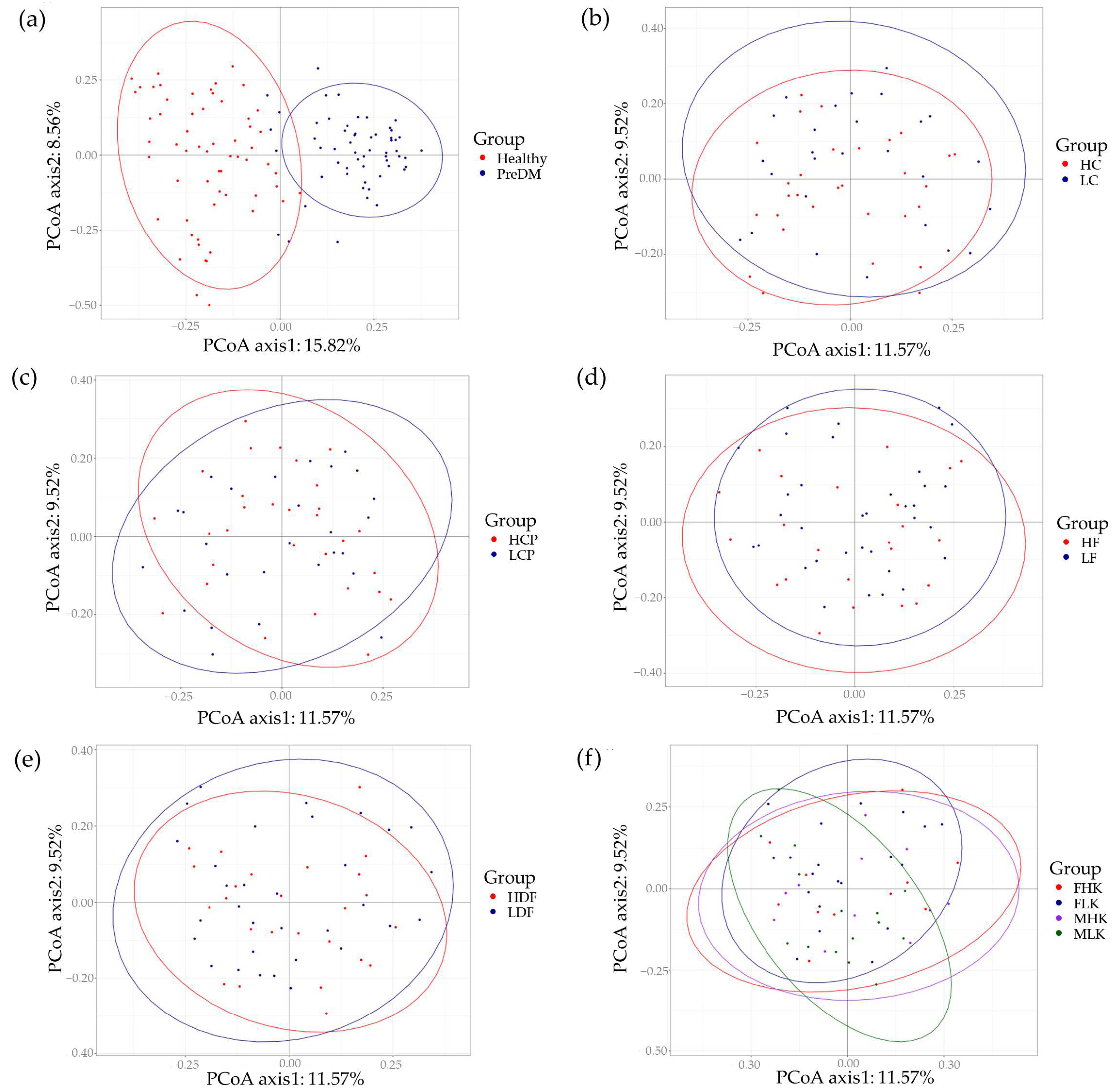
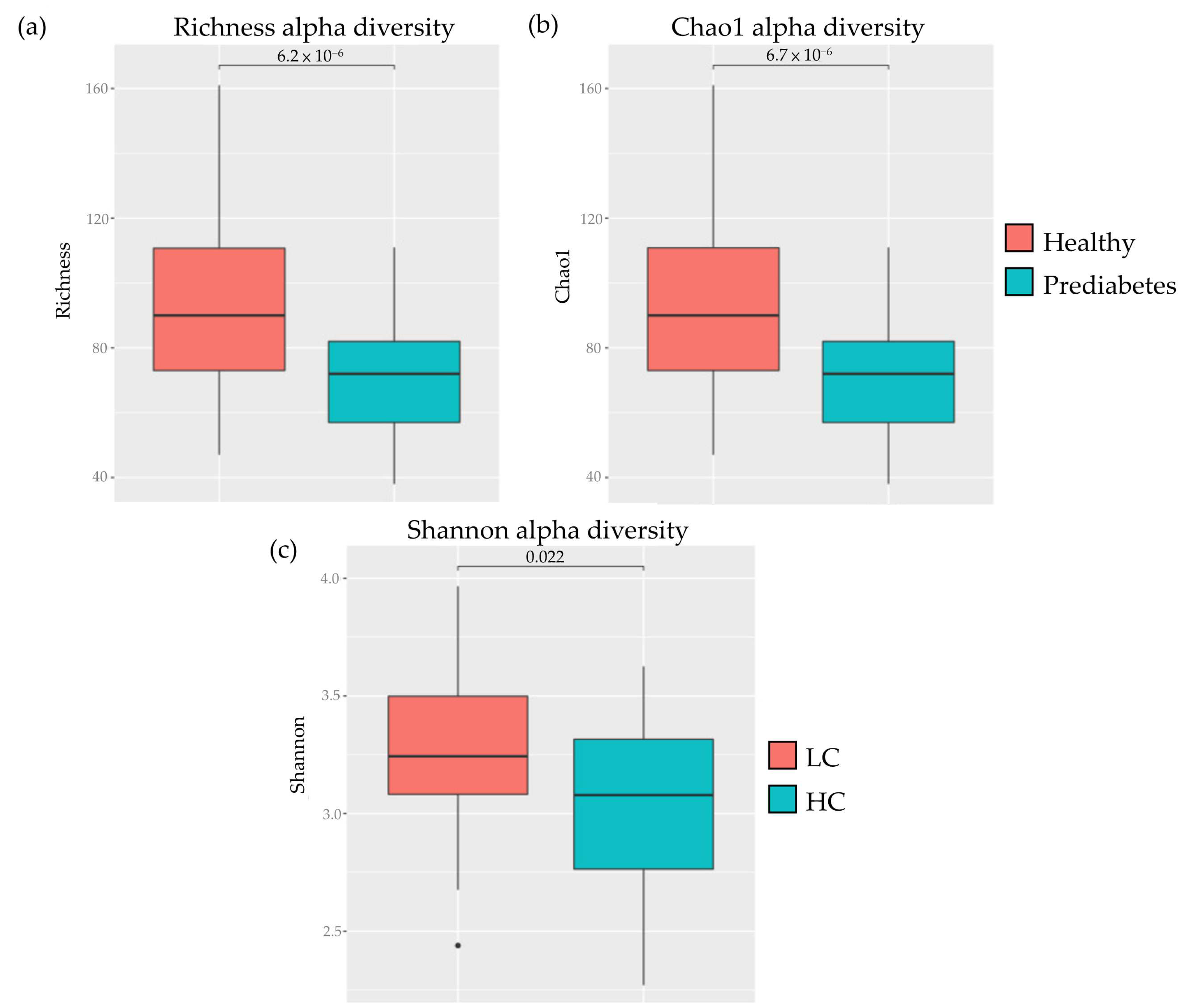
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tabák, A.G.; Herder, C.; Rathmann, W.; Brunner, E.J.; Kivimäki, M. Prediabetes: A high-risk state for diabetes development. Lancet 2012, 379, 2279–2290. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.; Karuranga, S.; Malanda, B.; Saeedi, P.; Basit, A.; Besançon, S.; Bommer, C.; Esteghamati, A.; Ogurtsova, K.; Zhang, P.; et al. Global and regional estimates and projections of diabetes-related health expenditure: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2020, 162, 108072. [Google Scholar] [CrossRef] [PubMed]
- Tuomilehto, J.; Lindström, J.; Eriksson, J.G.; Valle, T.T.; Hämäläinen, H.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Laakso, M.; Louheranta, A.; Rastas, M.; et al. Finnish Diabetes Prevention Study Group. Prevention of type 2 diabetes mellitus by changes in lifestyle among subjects with impaired glucose tolerance. N. Engl. J. Med. 2001, 344, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Knowler, W.C.; Barrett-Connor, E.; Fowler, S.E.; Hamman, R.F.; Lachin, J.M.; Walker, E.A.; Nathan, D.M.; Diabetes Prevention Program Research Group. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N. Engl. J. Med. 2002, 346, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, L.M.; Peltonen, M.; Ahlin, S.; Anveden, Å.; Bouchard, C.; Carlsson, B.; Jacobson, P.; Lönroth, H.; Maglio, C.; Näslund, I.; et al. Bariatric surgery and prevention of type 2 diabetes in Swedish obese subjects. N. Engl. J. Med. 2012, 367, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. EBiomedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.C.; Bueno, A.A.; de Souza, R.G.; Mota, J.F. Gut microbiota, probiotics and diabetes. Nutr. J. 2014, 13, 60. [Google Scholar] [CrossRef]
- Snelson, M.; de Pasquale, C.; Ekinci, E.I.; Coughlan, M.T. Gut microbiome, prebiotics, intestinal permeability and diabetes complications. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101507. [Google Scholar] [CrossRef] [PubMed]
- Tahapary, D.; Fatya, A.; Kurniawan, F.; Marcella, C.; Rinaldi, I.; Tarigan, T.; Harbuwono, D.S.; Yunir, E.; Soewondo, P.; Purnamasari, D. Increased intestinal-fatty acid binding protein in obesity-associated type 2 diabetes mellitus. PLoS ONE 2023, 18, e0279915. [Google Scholar] [CrossRef]
- Allam-Ndoul, B.; Castonguay-Paradis, S.; Veilleux, A. Gut Microbiota and Intestinal Trans-Epithelial Permeability. Int. J. Mol. Sci. 2020, 21, 6402. [Google Scholar] [CrossRef]
- Erdem, B.; Kaya, Y.; Kıran, T.R.; Yılmaz, S. An Association Between the Intestinal Permeability Biomarker Zonulin and the Development of Diabetic Retinopathy in Type II Diabetes Mellitus. Turk. J. Ophthalmol. 2023, 53, 91–96. [Google Scholar] [CrossRef]
- Tian, H.; Ge, X.; Nie, Y.; Yang, L.; Ding, C.; McFarland, L.V.; Zhang, X.; Chen, Q.; Gong, J.; Li, N. Fecal microbiota transplantation in patients with slow-transit constipation: A randomized, clinical trial. PLoS ONE 2017, 12, e0171308. [Google Scholar] [CrossRef] [PubMed]
- Vermeire, S.; Joossens, M.; Verbeke, K.; Wang, J.; Machiels, K.; Sabino, J.; Ferrante, M.; Van Assche, G.; Rutgeerts, P.; Raes, J. Donor Species Richness Determines Faecal Microbiota Transplantation Success in Inflammatory Bowel Disease. J. Crohn’s Colitis 2015, 10, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Liang, J.; Li, C. Effects of fecal microbiota transplantation in metabolic syndrome: A meta-analysis of randomized controlled trials. PLoS ONE 2023, 18, e0288718. [Google Scholar] [CrossRef] [PubMed]
- Apekey, T.A.; Maynard, M.J.; Kittana, M.; Kunutsor, S.K. Comparison of the Effectiveness of Low Carbohydrate Versus Low Fat Diets, in Type 2 Diabetes: Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2022, 14, 391. [Google Scholar] [CrossRef] [PubMed]
- Heiman, M.L.; Greenway, F.L. A healthy gastrointestinal microbiome is dependent on dietary diversity. Mol. Metab. 2016, 5, 317–320. [Google Scholar] [CrossRef]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.Z.; Abe, F.; Osawa, R. Age-related changes in gut microbiota composition from newborn to centenarian: A cross-sectional study. BMC Microbiol. 2016, 16, 90. [Google Scholar] [CrossRef]
- Rogers, M.A.M.; Aronoff, D.M. The influence of non-steroidal anti-inflammatory drugs on the gut microbiome. Clin. Microbiol. Infect. 2016, 22, e171–e179. [Google Scholar] [CrossRef]
- Zhang, N.; Ju, Z.; Zuo, T. Time for food: The impact of diet on gut microbiota and human health. Nutrition 2018, 51–52, 80–85. [Google Scholar] [CrossRef]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Tremaroli, V.; Schmidt, C.; Lundqvist, A.; Olsson, L.M.; Krämer, M.; Gummesson, A.; Perkins, R.; Bergström, G.; Bäckhed, F. The gut microbiota in prediabetes and diabetes: A population-based cross-sectional study. Cell Metab. 2020, 32, 379–390.e373. [Google Scholar] [CrossRef] [PubMed]
- Gaike, A.H.; Paul, D.; Bhute, S.S.; Dhotre, D.P.; Pande, P.; Upadhyaya, S.; Reddy, Y.; Sampath, R.; Ghosh, D.; Chandraprabha, D.; et al. The gut microbial diversity of newly diagnosed diabetics but not of prediabetics is significantly different from that of healthy nondiabetics. mSystems 2020, 5, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, A.; Lee, C.Y.; Lee, Y.C.; Liu, C.L.; Chen, H.K.; Li, Y.H.; Lai, L.C.; Tsai, M.H.; Ni, Y.H.; Chiu, H.M.; et al. Twnbiome: A public database of the healthy Taiwanese gut microbiome. BMC Bioinform. 2023, 24, 20231214. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.P.N.; Mannerås-Holm, L.; Puschmann, R.; Wu, H.; Troise, A.D.; Nijsse, B.; Boeren, S.; Bäckhed, F.; Fiedler, D.; deVos, W.M. Conversion of dietary inositol into propionate and acetate by commensal anaerostipes associates with host health. Nat. Commun. 2021, 12, 4798. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A Metagenome-Wide Association Study of Gut Microbiota in Type 2 Diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.R.; Varela-Nieto, I. Diabetes and the role of inositol-containing lipids in insulin signaling. Mol. Med. 1999, 5, 505–514. [Google Scholar] [CrossRef]
- Shettigar, K.; Murali, T.S. Virulence factors and clonal diversity of Staphylococcus aureus in colonization and wound infection with emphasis on diabetic foot infection. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 2235–2246. [Google Scholar] [CrossRef]
- Allin, K.H.; Tremaroli, V.; Caesar, R.; Jensen, B.A.H.; Damgaard, M.T.F.; Bahl, M.I.; Licht, T.R.; Hansen, T.H.; Nielsen, T.; Dantoft, T.M.; et al. Aberrant intestinal microbiota in individuals with prediabetes. Diabetologia 2018, 61, 810–820. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, M.; Wang, S.; Han, R.; Cao, Y.; Hua, W.; Mao, Y.; Zhang, X.; Pang, X.; Wei, C.; et al. Interactions between gut microbiota, host genetics and diet relevant to development of metabolic syndromes in mice. ISME J. 2010, 4, 232–241. [Google Scholar] [CrossRef]
- Yang, J.; Yu, J. The association of diet, gut microbiota and colorectal cancer: What we eat may imply what we get. Protein Cell 2018, 9, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Hui, H.; Cai, D. Differences in Fecal Bifidobacterium species Between Patients with Type 2 Diabetes and Healthy Individuals. Nan Fang Yi Ke Da Xue Xue Bao 2012, 32, 531–533, 564. [Google Scholar] [PubMed]
- Watanabe, M.; Sianoya, A.; Mishima, R.; Therdtatha, P.; Rodriguez, A.; Ramos, D.C.; Lee, Y.K.; Dalmacio, L.M.; Nakayama, J. Gut microbiome status of urban and rural Filipino adults in relation to diet and metabolic disorders. FEMS Microbiol. Lett. 2021, 368, fnab149. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Silva, C.; Kashani, A.; Hansen, T.H.; Pinna, N.K.; Anjana, R.M.; Dutta, A.; Saxena, S.; Støy, J.; Kampmann, U.; Nielsen, T.; et al. Trans-ethnic gut microbiota signatures of type 2 diabetes in Denmark and India. Genome Med. 2021, 13, 20210303. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Chung, S.K.; Vanamala, J.; Xu, B. Causal relationship between diet-induced gut microbiota changes and diabetes: A novel strategy to transplant Faecalibacterium prausnitzii in preventing diabetes. Int. J. Mol. Sci. 2018, 19, 3720. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Qi, Q.; Wang, Z.; Usyk, M.; Sotres-Alvarez, D.; Mattei, J.; Tamez, M.; Gellman, M.D.; Daviglus, M.; Hu, F.B.; et al. The gut microbiome modifies the association between a Mediterranean diet and diabetes in USA Hispanic/Latino population. J. Clin. Endocrinol. Metab. 2022, 107, e924–e934. [Google Scholar] [CrossRef] [PubMed]
- Baothman, O.A.; Zamzami, M.A.; Taher, I.; Abubaker, J.; Abu-Farha, M. The role of gut microbiota in the development of obesity and diabetes. Lipids Health Dis. 2016, 15, 108. [Google Scholar] [CrossRef] [PubMed]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, e917. [Google Scholar] [CrossRef] [PubMed]
- Bhute, S.S.; Suryavanshi, M.V.; Joshi, S.M.; Yajnik, C.S.; Shouche, Y.S.; Ghaskadbi, S.S. Gut microbial diversity assessment of indian type-2-diabetics reveals alterations in eubacteria, archaea, and eukaryotes. Front. Microbiol. 2017, 8, 214. [Google Scholar] [CrossRef]
- Jia, L.; Shan, K.; Pan, L.L.; Feng, N.; Lv, Z.; Sun, Y.; Li, J.; Wu, C.; Zhang, H.; Chen, W.; et al. Clostridium butyricum CGMCC0313.1 protects against autoimmune diabetes by modulating intestinal immune homeostasis and inducing pancreatic regulatory T cells. Front. Immunol. 2017, 8, 20171019. [Google Scholar] [CrossRef]
- Cohen, T.S.; Takahashi, V.; Bonnell, J.; Tovchigrechko, A.; Chaerkady, R.; Yu, W.; Jones-Nelson, O.; Lee, Y.; Raja, R.; Hess, S.; et al. Staphylococcus aureus drives expansion of low-density neutrophils in diabetic mice. J. Clin. Investig. 2019, 129, 2133–2144. [Google Scholar] [CrossRef] [PubMed]
- Fox, T.E.; Han, X.; Kelly, S.; Merrill, A.H.; Martin, R.E.; Anderson, R.E.; Gardner, T.W.; Kester, M. Diabetes alters sphingolipid metabolism in the retina: A potential mechanism of cell death in diabetic retinopathy. Diabetes 2006, 55, 3573–3580. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.; DeCoffe, D.; Molcan, E.; Gibson, D.L. Diet-induced dysbiosis of the intestinal microbiota and the effects on immunity and disease. Nutrients 2012, 4, 1095–1119. [Google Scholar] [CrossRef] [PubMed]
- Hippe, B.; Remely, M.; Aumueller, E.; Pointner, A.; Magnet, U.; Haslberger, A.G. Faecalibacterium prausnitzii phylotypes in type two diabetic, obese, and lean control subjects. Benef. Microbes 2016, 7, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Potrykus, M.; Czaja-Stolc, S.; Stankiewicz, M.; Kaska, Ł.; Małgorzewicz, S. Intestinal Microbiota as a Contributor to Chronic Inflammation and Its Potential Modifications. Nutrients 2021, 13, 3839. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, R.F.; Jobin, C. The microbiome and cancer. Nat. Rev. Cancer 2013, 13, 800–812. [Google Scholar] [CrossRef]
- Kort, R.; Notting, F.; Pirovano, W.; Sybesma, W. The butyrate-producing and spore-forming bacterial genus coprococcus as a potential biomarker for neurological disorders. Gut. Microbiome 2023, 2023, e16. [Google Scholar] [CrossRef]
- Di Vincenzo, F.; Del Gaudio, A.; Petito, V.; Lopetuso, L.R.; Scaldaferri, F. Gut microbiota, intestinal permeability, and systemic inflammation: A narrative review. Intern. Emerg. Med. 2024, 19, 275–293. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Category | Male | Female | Total |
|---|---|---|---|
| Healthy | 25 | 35 | 60 |
| Prediabetes | 26 | 31 | 57 |
| Prediabetes group | |||
| Carbohydrate group | |||
| LC | 14 | 13 | 27 |
| HC | 12 | 18 | 30 |
| Crude Protein group | |||
| LCP | 10 | 17 | 27 |
| HCP | 16 | 14 | 30 |
| Crude Fat group | |||
| LF | 14 | 20 | 34 |
| HF | 12 | 11 | 33 |
| Dietary Fiber group | |||
| LDF | 15 | 17 | 32 |
| HDF | 11 | 14 | 25 |
| Calories group | |||
| FHK | 0 | 11 | 11 |
| FLK | 0 | 20 | 20 |
| MHK | 11 | 0 | 11 |
| MLK | 15 | 0 | 15 |
| Healthy vs. Prediabetes Group | |||||
| Healthy | |||||
| Bacterial genera | Group | LDA score | p-value | Fold change (healthy/prediabetes) * | Reference |
| Bacteroides | Healthy | 4.70 | 6.92793 × 10−8 | 0.300 | [25] |
| Phocaeicola | Healthy | 4.55 | 8.30783 × 10−8 | 0.367 | |
| Alistipes | Healthy | 4.29 | 4.65658 × 10−8 | 0.074 | |
| Phascolarctobacterium | Healthy | 4.28 | 1.14771 × 10−7 | 0.121 | [26] |
| Parabacteroides | Healthy | 3.87 | 9.24002 × 10−11 | 0.190 | [27] |
| Paraprevotella | Healthy | 3.55 | 0.001328657 | 0.133 | [28] |
| Sutterella | Healthy | 3.25 | 1.29326 × 10−5 | 0.206 | [29] |
| Ruthenibacterium | Healthy | 3.19 | 2.69626 × 10−9 | 0.078 | [30] |
| Marseillibacter | Healthy | 3.01 | 6.86676 × 10−8 | 0.049 | [31] |
| Prediabetes | |||||
| Bacterial genera | Group | LDA score | p-value | Fold change (healthy/prediabetes) * | Reference |
| Blautia | Prediabetes | 4.75 | 2.94217 × 10−20 | 8.423 | [29,32] |
| Faecalibacterium | Prediabetes | 4.34 | 4.1797 × 10−7 | 2.335 | [33,34] |
| Bifidobacterium | Prediabetes | 4.32 | 3.62082 × 10−8 | 4.187 | [35] |
| Collinsella | Prediabetes | 4.22 | 1.76318 × 10−7 | 8.219 | |
| Fusicatenibacter | Prediabetes | 4.19 | 3.71865 × 10−10 | 9.128 | [36] |
| Streptococcus | Prediabetes | 4.11 | 1.00677 × 10−9 | 11.946 | [33,37] |
| Clostridium | Prediabetes | 4.01 | 1.40866 × 10−6 | 5.096 | [38] |
| Lachnoclostridium | Prediabetes | 3.94 | 0.001795935 | 1.985 | |
| Anaerostipes | Prediabetes | 3.92 | 3.32433 × 10−9 | 3.972 | [39] |
| Mediterraneibacter | Prediabetes | 3.89 | 2.233 × 10−7 | 5.735 | [40] |
| Butyricicoccus | Prediabetes | 3.57 | 4.19266 × 10−8 | 3.015 | [41] |
| Dorea | Prediabetes | 3.49 | 4.89717 × 10−11 | 6.086 | [37,42] |
| Clostridioides | Prediabetes | 3.18 | 8.49243 × 10−6 | 7.105 | |
| Eggerthella | Prediabetes | 3.02 | 1.11996 × 10−8 | 9.801 | [43] |
| Carbohydrate group | |||||
| Bacterial genera | Group | LDA score | p-value | Fold change (HC/LC) * | |
| Coprococcus | LC | 3.73 | 0.00920058 | 0.414 | [44] |
| Group | Kegg ID | Kegg Pathway Name | Healthy/Prediabetes * | p-Value |
|---|---|---|---|---|
| healthy/prediabetes | ko04975 | Fat digestion and absorption | 6.36 | 6.312 × 10−5 |
| ko00062 | Fatty acid elongation | 0.17 | 4.219 × 10−14 | |
| ko00563 | GPI-anchor biosynthesis | 64.97 | 1.528 × 10−2 | |
| ko00604 | Glycosphingolipid biosynthesis—ganglio series | 2.04 | 9.552 × 10−10 | |
| ko00072 | Synthesis and degradation of ketone bodies | 0.45 | 7.304 × 10−12 | |
| Kegg ID | Kegg pathway name | HC/LC * | p-value | |
| HC/LC | ko04975 | Fat digestion and absorption | 0.39 | 2.033 × 10−2 |
| Kegg ID | Kegg pathway name | HCP/LCP * | p-value | |
| HCP/LCP | ko04975 | Fat digestion and absorption | 2.31 | 4.899 × 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, W.-L.; Chen, Y.-E.; Tseng, H.-T.; Cheng, C.-F.; Wu, J.-H.; Hou, Y.-C. Gut Microbiota in Patients with Prediabetes. Nutrients 2024, 16, 1105. https://doi.org/10.3390/nu16081105
Chang W-L, Chen Y-E, Tseng H-T, Cheng C-F, Wu J-H, Hou Y-C. Gut Microbiota in Patients with Prediabetes. Nutrients. 2024; 16(8):1105. https://doi.org/10.3390/nu16081105
Chicago/Turabian StyleChang, Wei-Lin, Yu-En Chen, Hsiang-Tung Tseng, Ching-Feng Cheng, Jing-Hui Wu, and Yi-Cheng Hou. 2024. "Gut Microbiota in Patients with Prediabetes" Nutrients 16, no. 8: 1105. https://doi.org/10.3390/nu16081105
APA StyleChang, W.-L., Chen, Y.-E., Tseng, H.-T., Cheng, C.-F., Wu, J.-H., & Hou, Y.-C. (2024). Gut Microbiota in Patients with Prediabetes. Nutrients, 16(8), 1105. https://doi.org/10.3390/nu16081105