

The Gut Microbiome in Early Life Stress: A Systematic Review

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
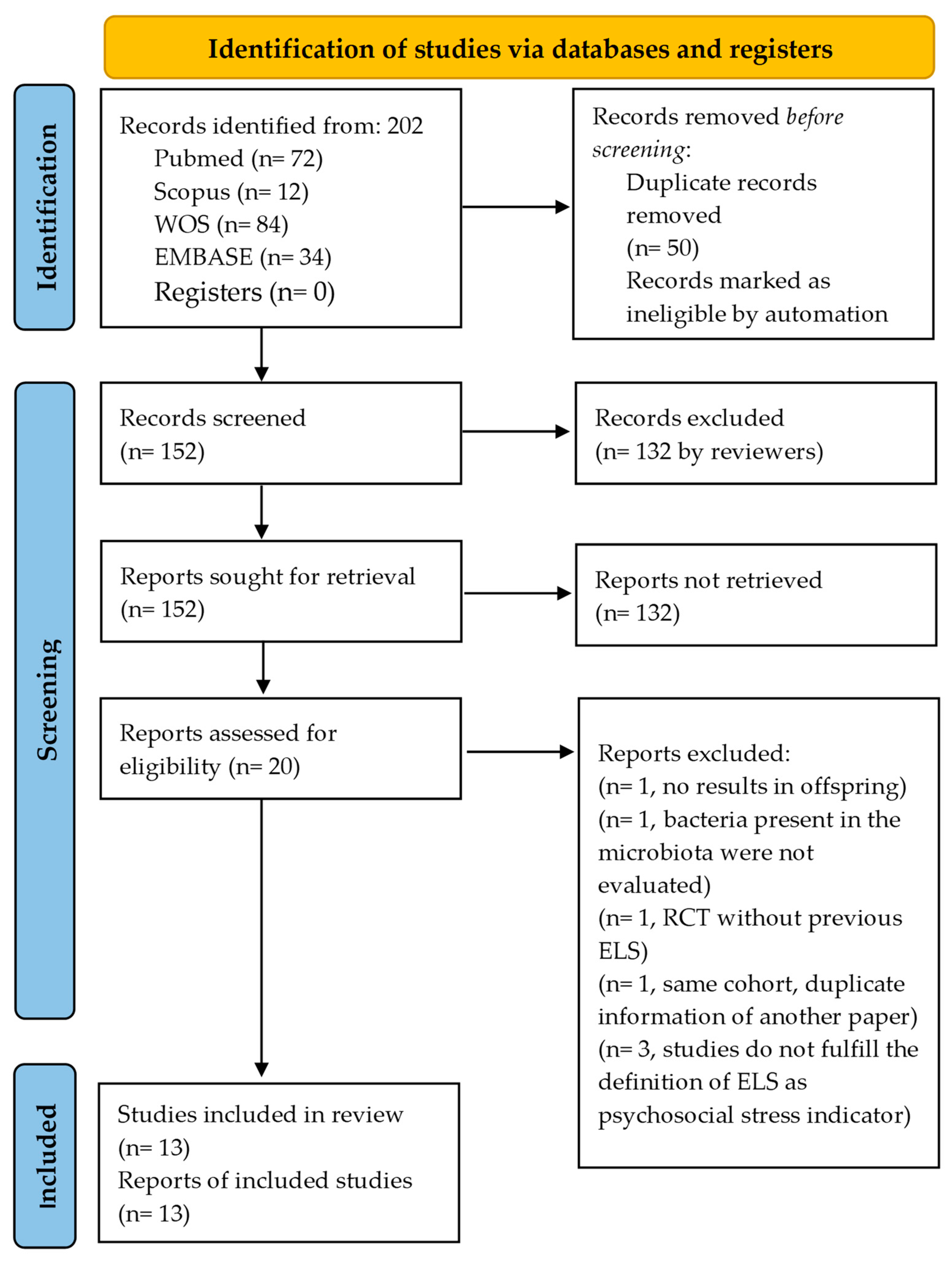
2.1. Search Strategy and Inclusion/Exclusion Criteria
2.2. Data Extraction
2.3. Assessment of Risk of Bias
2.4. Strategy for Data Synthesis and Analysis of Subgroups
3. Results
3.1. Overview
3.2. Risk of Bias
3.2.1. Quality Assessment of Longitudinal Cohort Studies Based on the Newcastle-Ottawa Scale
3.2.2. Quality Assessment of Case-Control Studies Based on the Newcastle-Ottawa Scale
3.2.3. Quality Assessment of Cross-Sectional Studies Based on the Newcastle-Ottawa Scale
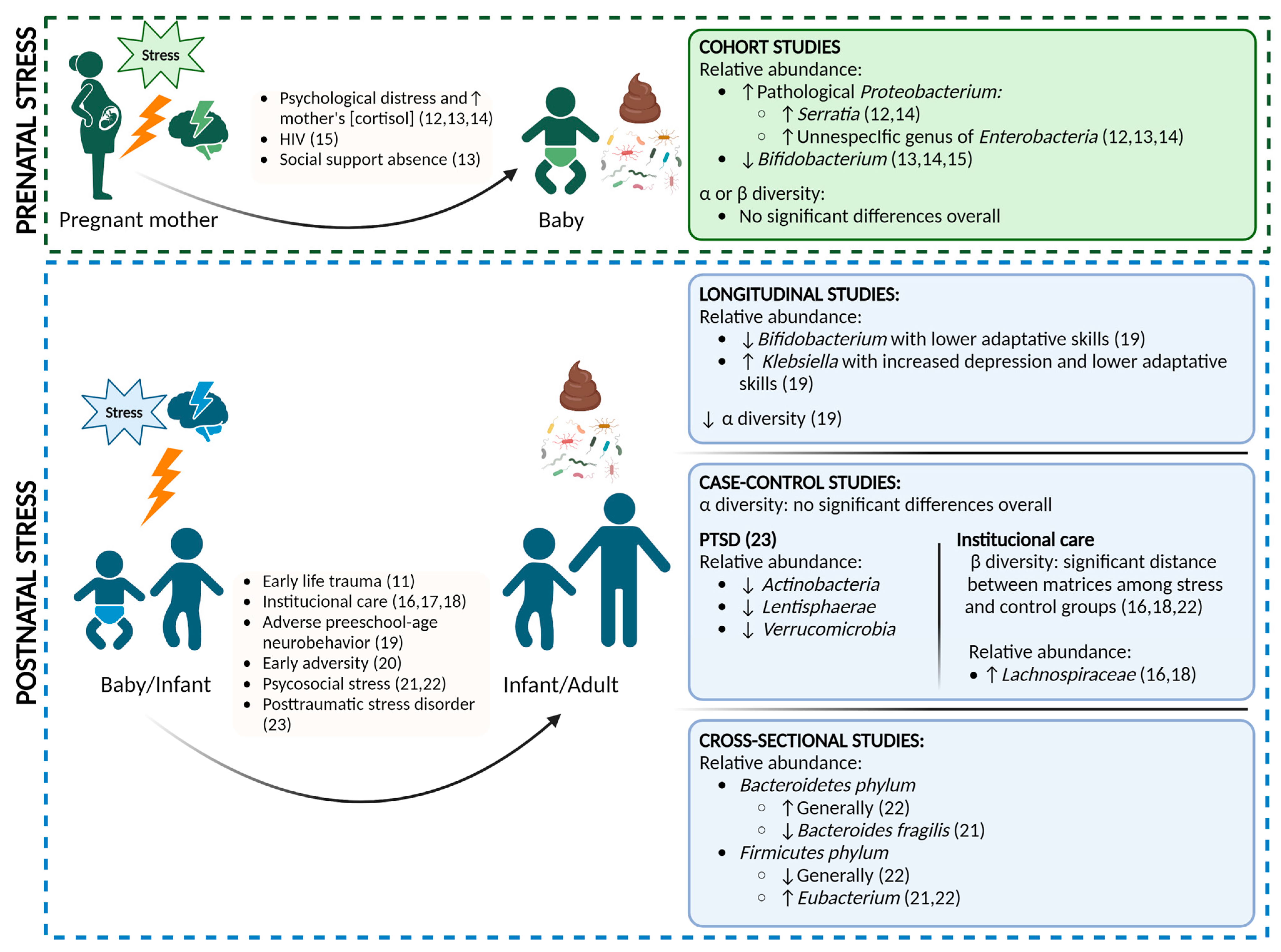
3.3. Prenatal Stress
Longitudinal Studies
3.4. Postnatal Stress
3.4.1. Longitudinal Studies
3.4.2. Case-Control Studies
3.4.3. Cross-Sectional Studies
4. Discussion
5. Limitations
6. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coussons-Read, M.E. Effects of prenatal stress on pregnancy and human development: Mechanisms and pathways. Obstet. Med. 2013, 6, 52–57. [Google Scholar] [CrossRef]
- Leeb, R.T. Child Maltreatment Surveillance: Uniform Definitions for Public Health and Recommended Data Elements; Centers for Disease Control and Prevention, National Center for Injury Prevention and Control: Atlanta, GA, USA, 2008; pp. 131–135. [Google Scholar]
- Vogel, S.C.; Brito, N.H.; Callaghan, B.L. Early Life Stress and the Development of the Infant Gut Microbiota: Implications for Mental Health and Neurocognitive Development. Curr. Psychiatry Rep. 2020, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Green, J.G.; McLaughlin, K.A.; Berglund, P.A.; Gruber, M.J.; Sampson, N.A.; Zaslavsky, A.M.; Kessler, R.C. Childhood adversities and adult psychiatric disorders in the national comorbidity survey replication I: Associations with first onset of DSM-IV disorders. Arch. Gen. Psychiatry 2010, 67, 113–123. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, K.A.; Green, J.G.; Gruber, M.J.; Sampson, N.A.; Zaslavsky, A.M.; Kessler, R.C. Childhood adversities and adult psychiatric disorders in the national comorbidity survey replication II: Associations with persistence of DSM-IV disorders. Arch. Gen. Psychiatry 2010, 67, 124–132. [Google Scholar] [CrossRef]
- Knowles, S.R.; Nelson, E.A.; Palombo, E. Investigating the role of perceived stress on bacterial flora activity and salivary cortisol secretion: A possible mechanism underlying susceptibility to illness. Biol. Psychol. 2008, 77, 132–137. [Google Scholar] [CrossRef]
- Kato-Kataoka, A.; Nishida, K.; Takada, M.; Kawai, M.; Kikuchi-Hayakawa, H.; Suda, K.; Ishikawa, H.; Gondo, Y.; Shimizu, K.; Matsuki, T.; et al. Fermented milk con taining lactobacillus casei strain shirota preserves the diversity of the gut microbiota and relieves abdominal dysfunction in healthy medical students exposed to academic stress. Appl. Environ. Microbiol. 2016, 82, 3649–3658. [Google Scholar] [CrossRef]
- Lupien, S.; Maheu, F.; Tu, M.; Fiocco, A.; Schramek, T. The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition. Brain Cogn. 2007, 65, 209–237. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Physiology and neurobiology of stress and adaptation: Central role of the brain. Physiol. Rev. 2007, 87, 873–904. [Google Scholar] [CrossRef] [PubMed]
- Mariani, N.; Borsini, A.; Cecil, C.A.M.; Felix, J.F.; Sebert, S.; Cattaneo, A.; Walton, E.; Milaneschi, Y.; Cochrane, G.; Amid, C.; et al. Identifying causative mechanisms linking early-life stress to psycho-cardio-metabolic multi-morbidity: The EarlyCause project. PLoS ONE 2021, 16, e0245475. [Google Scholar] [CrossRef]
- Coley, E.J.; Mayer, E.A.; Osadchiy, V.; Chen, Z.; Subramanyam, V.; Zhang, Y.; Hsiao, E.Y.; Gao, K.; Bhatt, R.; Dong, T.; et al. Early life adversity predicts brain-gut alterations associated with increased stress and mood. Neurobiol. Stress 2021, 15, 100348. [Google Scholar] [CrossRef]
- Aatsinki, A.-K.; Keskitalo, A.; Laitinen, V.; Munukka, E.; Uusitupa, H.-M.; Lahti, L.; Kortesluoma, S.; Mustonen, P.; Rodrigues, A.J.; Coimbra, B.; et al. Maternal prenatal psychological distress and hair cortisol levels associate with infant fecal microbiota composition at 2.5 months of age. Psychoneuroendocrinology 2020, 119, 104754. [Google Scholar] [CrossRef] [PubMed]
- Jahnke, J.R.; Roach, J.; Azcarate-Peril, M.A.; Thompson, A.L. Maternal precarity and HPA axis functioning shape infant gut microbiota and HPA axis development in humans. PLoS ONE 2021, 16, e0251782. [Google Scholar] [CrossRef]
- Zijlmans, M.A.; Korpela, K.; Riksen-Walraven, J.M.; de Vos, W.M.; de Weerth, C. Maternal prenatal stress is associated with the infant intestinal microbiota. Psychoneuroendocrinology 2015, 53, 233–245. [Google Scholar] [CrossRef]
- Grant-Beurmann, S.; Jumare, J.; Ndembi, N.; Matthew, O.; Shutt, A.; Omoigberale, A.; Martin, O.A.; Fraser, C.M.; Charurat, M. Dynamics of the infant gut microbiota in the first 18 months of life: The impact of maternal HIV infection and breastfeeding. Microbiome 2022, 12, 61. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, B.L.; Fields, A.; Gee, D.G.; Gabard-Durnam, L.; Caldera, C.; Humphreys, K.L.; Goff, B.; Flannery, J.; Telzer, E.H.; Shapiro, M.; et al. Mind and gut: Associations between mood and gastrointestinal distress in children exposed to adversity. Dev. Psychopathol. 2020, 32, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Hermes, G.D.A.; Eckermann, H.A.; de Vos, W.M.; de Weerth, C. Does entry to center-based childcare affect gut microbial colonization in young infants? Sci. Rep. 2020, 24, 10235. [Google Scholar] [CrossRef] [PubMed]
- Reid, B.M.; Horne, R.; Donzella, B.; Szamosi, J.C.; Coe, C.L.; Foster, J.A.; Gunnar, M.R. Microbiota-immune alterations in adolescents following early life adversity: A proof of concept study. Dev. Psychobiol. 2021, 63, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Laue, H.E.; Karagas, M.R.; Coker, M.O.; Bellinger, D.C.; Baker, E.R.; Korrick, S.A.; Madan, J.C. Sex-specific relationships of the infant microbiome and early-childhood behavioral outcomes. Pediatr. Res. 2022, 92, 580–591. [Google Scholar] [CrossRef]
- Keskitalo, A.; Aatsinki, A.-K.; Kortesluoma, S.; Pelto, J.; Korhonen, L.; Lahti, L.; Lukkarinen, M.; Munukka, E.; Karlsson, H.; Karlsson, L. Gut microbiota diversity but not composition is related to saliva cortisol stress response at the age of 2.5 months. Stress 2021, 24, 551–560. [Google Scholar] [CrossRef]
- Flannery, J.E.; Stagaman, K.; Burns, A.R.; Hickey, R.J.; Roos, L.E.; Giuliano, R.J.; Fisher, P.A.; Sharpton, T.J. Gut Feelings Begin in Childhood: The Gut Metagenome Correlates with Early Environment, Caregiving, and Behavior. mBio 2020, 11, e02780-19. [Google Scholar] [CrossRef]
- Michels, N.; Van de Wiele, T.; Fouhy, F.; O’Mahony, S.; Clarke, G.; Keane, J. Gut microbiome patterns depending on children’s psychosocial stress: Reports versus biomarkers. Brain, Behav. Immun. 2019, 80, 751–762. [Google Scholar] [CrossRef]
- Malan-Muller, S.; Valles-Colomer, M.; Foxx, C.L.; Vieira-Silva, S.; Heuvel, L.L.v.D.; Raes, J.; Seedat, S.; Lowry, C.A.; Hemmings, S.M. Exploring the relationship between the gut microbiome and mental health outcomes in a posttraumatic stress disorder cohort relative to trauma-exposed controls. Eur. Neuropsychopharmacol. 2022, 56, 24–38. [Google Scholar] [CrossRef]
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [PubMed]
- Carroll, I.M.; Threadgill, D.; Threadgill, D.S. The gastrointestinal microbiome: A malleable, third genome of mammals. Mamm. Genome 2009, 20, 395–403. [Google Scholar] [CrossRef]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef] [PubMed]
- Agustí, A.; Campillo, I.; Balzano, T.; Benítez-Páez, A.; López-Almela, I.; Romaní-Pérez, M.; Forteza, J.; Felipo, V.; Avena, N.M.; Sanz, Y. Bacteroides uniformis CECT 7771 Modulates the Brain Reward Response to Reduce Binge Eating and Anxiety-Like Behavior in Rat. Mol. Neurobiol. 2021, 58, 4959–4979. [Google Scholar] [CrossRef]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, G.; Siopi, E.; Guenin-Macé, L.; Pascal, M.; Laval, T.; Rifflet, A.; Boneca, I.G.; Demangel, C.; Colsch, B.; Pruvost, A.; et al. Effect of gut microbiota on depressive-like behaviors in mice is mediated by the endocannabinoid system. Nat. Commun. 2020, 11, 6363. [Google Scholar] [CrossRef]
- Doll, J.P.; Vázquez-Castellanos, J.F.; Schaub, A.C.; Schweinfurth, N.; Kettelhack, C.; Schneider, E.; Yamanbaeva, G.; Mählmann, L.; Brand, S.; Beglinger, C.; et al. Fecal Microbiota Transplantation (FMT) as an Adjunctive Therapy for Depression-Case Report. Front. Psychiatry 2022, 17, 815422. [Google Scholar] [CrossRef] [PubMed]
- Backhed, F. Programming of host metabolism by the gut microbiota. Ann. Nutr. Metab. 2011, 58 (Suppl. 2), 44–52. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, K.-A.M.; Luczynski, P.; Dinan, T.G.; Cryan, J.F. Reframing the Teenage Wasteland: Adolescent Microbiota-Gut-Brain Axis. Can. J. Psychiatry 2016, 61, 214–221. [Google Scholar] [CrossRef]
- Monk, C.; Lugo-Candelas, C.; Trumpff, C. Prenatal Developmental Origins of Future Psychopathology: Mechanisms and Pathways. Annu. Rev. Clin. Psychol. 2019, 15, 317–344. [Google Scholar] [CrossRef]
- Weinstock, M. The long-term behavioural consequences of prenatal stress. Neurosci. Biobehav. Rev. 2008, 32, 1073–1086. [Google Scholar] [CrossRef]
- van de Loo, K.F.; van Gelder, M.M.; Roukema, J.; Roeleveld, N.; Merkus, P.J.; Verhaak, C.M. Prenatal maternal psychological stress and childhood asthma and wheezing: A meta-analysis. Eur. Respir. J. 2016, 47, 133–146. [Google Scholar] [CrossRef]
- Soares-Cunha, C.; Coimbra, B.; Borges, S.; Domingues, A.V.; Silva, D.; Sousa, N.; Rodrigues, A.J. Mild prenatal stress causes emotional and brain structuralmodifications in rats of both sexes. Front. Behav. Neurosci. 2018, 12, 129. [Google Scholar] [CrossRef] [PubMed]
- Jašarević, E.; Howard, C.D.; Misic, A.M.; Beiting, D.P.; Bale, T.L. Stress during pregnancy alters temporal and spatial dynamics of the maternal and offspring microbiome in a sex-specific manner. Sci. Rep. 2017, 7, 44182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, N.; Chen, R.; Lee, T.; Gao, Y.; Yuan, Z.; Nie, Y.; Sun, T. Prenatal stress leads to deficits in brain development, mood related behaviors and gut microbiota in offspring. Neurobiol. Stress 2021, 15, 100333. [Google Scholar] [CrossRef] [PubMed]
- Hantsoo, L.; Zemel, B.S. Stress gets into the belly: Early life stress and the gut microbiome. Behav. Brain Res. 2021, 414, 113474. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Wells, G.A.; Shea, B.; O’Connell, D.; Peterson, J.; Welch, V.; Losos, M.; Tugwell, P. The Newcastle–Ottawa Scale (NOS) for Assessing the Quality of Non-Randomized Studies in Meta-Analysis; The Ottawa Hospital Research Institute: Ottawa, ON, Canada, 2011. [Google Scholar]
- Herzog, R.; Álvarez-Pasquin, M.J.; Díaz, C.; Del Barrio, J.L.; Estrada, J.M.; Gil, Á. Are healthcare workers’ intentions to vaccinate related to their knowledge, beliefs and attitudes? a systematic review. BMC Public Health 2013, 19, 154. [Google Scholar] [CrossRef] [PubMed]
- Warner, B.B.; Deych, E.; Zhou, Y.; Hall-Moore, C.; Weinstock, G.M.; Sodergren, E.; Shaikh, N.; Hoffmann, J.A.; Linneman, L.A.; Hamvas, A.; et al. Gut bacteria dysbiosis and necrotising enterocolitis in very low birthweight infants: A prospective case-control study. Lancet 2016, 387, 1928–1936. [Google Scholar] [CrossRef]
- Picard, C.; Fioramonti, J.; Francois, A.; Robinson, T.; Neant, F.; Matuchansky, C. Review article: Bifidobacteria as probiotic agents—Physiological effects and clinical benefits. Aliment. Pharmacol. Ther. 2005, 22, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Chen, Y.; Zhu, H.; Wang, L.; Qian, X.; Zou, R.; Zhao, J.; Zhang, H.; Qian, L.; Wang, Q.; et al. Bifidobacterium breve CCFM1025 attenuates major depression disorder via regulating gut microbiome and tryptophan metabolism: A randomized clinical trial. Brain Behav. Immun. 2022, 100, 233–241. [Google Scholar] [CrossRef]
- Pinto-Sanchez, M.I.; Hall, G.B.; Ghajar, K.; Nardelli, A.; Bolino, C.; Lau, J.T.; Martin, F.P.; Cominetti, O.; Welsh, C.; Rieder, A.; et al. Probiotic Bifidobacterium longum NCC3001 Reduces Depression Scores and Alters Brain Activity: A Pilot Study in Patients with Irritable Bowel Syndrome. Gastroenterology 2017, 153, 448–459.e8. [Google Scholar] [CrossRef] [PubMed]
- Hemmings, S.M.; Malan-Muller, S.; van den Heuvel, L.L.; Demmitt, B.A.; Stanislawski, M.A.; Smith, D.G.; Bohr, A.D.; Stamper, C.E.; Hyde, E.R.; Morton, J.T.; et al. The Microbiome in Posttraumatic Stress Disorder and Trauma-Exposed Controls: An Exploratory Study. Psychosom. Med. 2017, 79, 936–946. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef] [PubMed]
- Rosin, S.; Xia, K.; Azcarate-Peril, M.A.; Carlson, A.L.; Propper, C.B.; Thompson, A.L.; Grewen, K.; Knickmeyer, R.C. A preliminary study of gut microbiome variation and HPA axis reactivity in healthy infants. Psychoneuroendocrinology 2021, 124, 105046. [Google Scholar] [CrossRef]
- Huang, Y.; Shi, X.; Li, Z.; Shen, Y.; Shi, X.; Wang, L.; Li, G.; Yuan, Y.; Wang, J.; Zhang, Y.; et al. Possible association of Firmicutes in the gut microbiota of patients with major depressive disorder. Neuropsychiatr. Dis. Treat. 2018, 14, 3329–3337. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Reference | Selection | Comparability | Outcome | Total | |||||
|---|---|---|---|---|---|---|---|---|---|
| Representativeness of Exposed Cohort | Selection Non-Exposed Cohort | Ascertainment of Exposure | Outcome of Interest Not Present at Start of Study | Comparability of Cohort | Assessment of the Outcome | Follow-Up Long Enough for Outcomes | Adequacy of Follow-Up of Cohorts | ||
| Zijlmans et al., 2015 [14]. | * | * | * | ** | * | * | 7 | ||
| Aatsinki et al., 2020 [12]. | * | * | ** | * | * | * | 7 | ||
| Grant-Beurmann et al., 2022 [15]. | * | * | * | * | * | * | * | 7 | |
| Jahnke et al., 2021 [13]. | * | * | * | ** | * | * | 7 | ||
| Laue et al., 2021 [19]. | * | * | * | * | ** | * | * | 8 | |
| Reference | Selection | Comparability | Exposure | Total | |||||
|---|---|---|---|---|---|---|---|---|---|
| Case Definition | Representativeness of the Cases | Selection of Controls | Definition of Controls | Comparability of Cases and Controls | Ascertainment of the Exposure | Same Method of Ascertainment | Non-Response Rate | ||
| Malan-Muller et al., 2022 [23]. | * | * | * | * | ** | * | * | 8 | |
| Hermes et al., 2020 [17]. | * | * | * | * | ** | * | * | 8 | |
| Keskitalo et al., 2021 [20]. | * | * | * | * | * | * | * | 7 | |
| Callahan et al., 2020 [16]. | * | * | * | ** | * | * | 7 | ||
| Reid et al., 2021 [18]. | * | * | * | ** | * | * | 7 | ||
| Reference | Selection | Comparability | Outcome | Total | ||||
|---|---|---|---|---|---|---|---|---|
| Representativeness of the Sampler | Selection of the Control Group | Ascertainment of Exposure | Non-Respondents | The Subjects in Different Outcome Groups are Comparable | Ascertainment of the Outcome | Statistical Test | ||
| Coley et al., 2021 [11]. | * | * | ** | ** | ** | * | 9 | |
| Michels et al., 2019 [22]. | * | ** | ** | ** | * | 8 | ||
| Flannery et al., 2020 [21]. | * | * | * | ** | * | 6 | ||
| Study | Population | Design and Aim | Measures | Results/Outcomes | ||
|---|---|---|---|---|---|---|
| References | N | Characteristics | Microbiota | Stress | ||
| Aatsinki et al., 2020 [12] | N = 446 Mother–infant pairs |
| Large prospective longitudinal study (FinnBrain Birth Cohort 2011–2015) Aim: to investigate the role of early life exposures (PPD and HCC) specifically on infant fecal microbiota | Infant fecal sample collection: at 2.5 mths Analysis of fecal samples:
| Maternal hair samples cortisol measurement at 24 gwk (Cortisol Saliva kit): HCC Maternal prenatal psychological distress questionnaires, self-reported at 14, 24, and 34 gwk:
| Chronically elevated * maternal PPD symptoms were associated with proteobacteria in the infant:
Cortisol (hair) in mothers at 24 gwk was associated:
|
| Grant-Beurmann et al., 2022 [15] | N total = 272 mother–infant pairs N (HIV/HUU) = 131 N(HIV+/HEU) = 141 |
31–37 yrs (HEU)
| Prospective cohort study of mother–infant pairs 2015–2018 Aim: to assess potential differences in the gut microbiota in infants born to HIV-positive and -negative mothers during the first 18 mths of life | Mother samples:
Analysis of fecal and vaginal samples:
| Clinical assessment: Standardized questionnaires (medication and comorbidity history), general physical examination, anthropometric assessment | Maternal vaginal and infant fecal microbiota showed increased the diversity over time. Breastfeeding was associated with differences in gut microbiota of infants (HEU and HUU). The relative abundance association was:
Low Bifidobacterium abundance associated with low weight Breast milk composition differed by time point and HIV infection status
|
| Jahnke et al., 2021 [13] | N total = 25 mother–infant dyads |
Data collected over 12 mths Country: Ecuador (Galápagos, San Cristobal island) | Longitudinal cohort dyad study. Aim: to assess relationships among maternal precarity and HPA axis dysregulation during the peripartum period, infant gut microbiome composition, and infant HPA axis functioning | Infant fecal collection at 2 mths postpartum Analysis of fecal samples:
| Measures of maternal precarity taken during and after pregnancy:
scales Saliva sample (cortisol):
| Gut microbiome alpha diversity:
|
| Zijlmans et al., 2015 [14] | N total = 56 children | Children followed from the 3rd trimester of pregnancy until 110 days after birth Children:
| Prospective longitudinal study Aim: to investigate the development of the gut microbiota as a potential pathway linking maternal prenatal stress and infant health | Infant fecal sample collection: 5 time points (from birth until ± 110 days of life): T1–T5 Analysis of fecal samples:
| Questionnaires (3rd Trimester):
Mothers report infant gastrointestinal symptoms and allergic reactions monthly using the ICPC | Sum of: stress questionnaire scores + cortisol concentration (12 a.m.) were most strongly associated with the infant microbiota Infants with high cumulative stress (high reported stress + high cortisol saliva measure) showed:
|
| Study | Population | Design | Measures | Results/Effect on Outcomes | ||
|---|---|---|---|---|---|---|
| References | N | Characteristics | Microbiota | Stress | ||
| Longitudinal Studies | ||||||
| Laue et al., 2022 [19] | N = 260 children from mothers in NHBCS cohort with complete stool and BASC data | Male = 144 Female = 116 Mean gestational age 39.5 wks C-section n = 78 Country: U.S.A | NHBCS longitudinal pregnancy cohort Aim: to identify sex-specific prospective associations between the early life gut microbiome and preschool-age neurobehavior | Stool sampling in children at 6 wks, 1 yr and 2 yr postpartum Analysis of fecal samples:
| Questionnaires: Behavioral development assessed by parent using BASC-2 at age 3 with 10 subscales scores:
| Alpha diversity:
Boys at 6 wks: Adaptive skill composite score linked to higher abundance of:
Worse hyperactivity scores linked to relative abundance of Faecalitalea. Overall, stool sample at 2 yrs: 4 Blautia ASVs linked to worse measures of Hyperactivity. Metagenomic species related to BASC-2 scores: 6 wks:
No associations significant at the FDR level observed at 1 yr, except for lower Depression and Internalizing problems score in girls with abundance of Streptococcus peroris. Bacterial functional pathways related to BASC-2 scores:
|
| Case-control studies | ||||||
| Callahan et al., 2020 [16]. (2nd study) | N total = 16. N Controls = 8 N children with EA n = 8 | Controls:
| Aim: to study association between early caregiving adversity, the gastrointestinal (GI) microbiome, and brain reactivity to threat stimuli (fear faces) | Single fecal sample collection. Analysis of fecal samples:
| EA group, i.e., institutional or foster care followed by international adoption vs. controls | Alpha Diversity:
|
| Hermes et al., 2020 [17] | N = 98 infants CC infants HOME infants | CC group:
| Case-control study. CC entry at 3 mths vs. HOME Aim: to investigate whether CC, as compared with being cared for by the parents at home, alters the composition of the gut microbiota | Stool samples collected at 10 wks post-partum and 4 wks after CC entrance. Analysis of fecal samples:
| CC vs. HOME | No significant effect of CC entry or the number of half-days in CC compared with staying at home, on the microbiota using Redundancy analysis, Random Forest, and Bayesian linear models. |
| Keskitalo et al., 2021 [20] | N = 193 2.5-mth-old infants with both fecal sample and salivary stress response measurement | Males = 102 Females = 91 Vaginal birth = 154 Mean age = 10.4 wks Mean birth weight = 3616 g Gestational age mean = 40 wks Country: Finland | Nested case-control study with infants exposed to different types of maternal prenatal psychological stress vs. non-exposed controls (Subset of FinnBrain Focus cohort study) Aim: to identify the potential link between the cortisol stress response and the gut microbiota at the age of 2.5 mths | Single fecal sample collected (2.5 mths) Analysis of fecal samples:
| Infants exposed to maternal prenatal psychological stress vs. non-exposed controls Stress test:
| Post-stressor salivary cortisol AUCi negatively associated with alpha diversity but not beta diversity. No evident associations between cortisol stress response and bacterial taxa when ALDEx2 analyses were adjusted for the selected covariates. |
| Malan-Muller et al., 2022 [23] | N total = 137 N PTSD = 79 PTSD N TECs = 58 | Age PTSD = 32–52 yrs Age TECs = 38–58 yrs Female PTSD = 63 Female TECs = 47 Country: South Africa | Case-control study to investigate associations between the gut microbiome and mental health outcomes PTSD and TECs | Stool samples collected the same wk as the clinical assessment. Analysis of fecal samples:
| PTSD vs. TE To evaluate PTSD:
| Mental outcome
↑ Psychotropic medication use; ↑ Prevalence of MDD and anxiety disorders. Alpha diversity:
Associations between traumatic experiences, psychiatric diagnosis and taxonomic abundance:
|
| Reid et al., 2021 [18] | N total = 38 N PI = 17 N COMP = 18 | PI Mean age = 16.8 yrs Female = 13 CMV Seropositive = 14 COMP Mean age = 15.9 yrs Female = 11 CMV seropositive = 5 Country: U.S.A | Case-control study to assess the relation between ELS in PI participants: gut microbiome and inflammatory markers PI participants spent 70% of preadoption life in institutional care COMP participants were from families with comparable education and incomes Parent/youth questionnaire report of health and diet Stool and blood samples were collected once | Fecal samples were collected at home Analysis of fecal samples:
| PI vs. COMP There was no questionnaire for evaluating stress However, PI participants spent 70% of preadoption life in institutional care, considered as an early life stress factor in the study | Alpha diversity:
|
| Cross-sectional studies | ||||||
| Coley et al., 2021 [11] | Ntotal = 128 healthy adults | Male n = 43 Female n = 85 Age females ≤ 45 years and premenopausal groups based on ETI-SR score
| Case-control based on ETI score (ETI-SR total > 4) Aim: to test whether ELS-related alterations in gut microbial metabolites are associated with alterations in brain connectivity, disordered mood, and increased vulnerability to stress in adulthood | Single fecal sample collected Analysis of fecal samples:
| ETI score > 4 vs. ETI score ≤ 4 Questionnaires:
| No significant relationships between history of ELS exposure and microbial alpha diversity, microbial beta diversity, or relative taxonomic abundance, at either phylum or genus level. Symptoms of anxiety, depression and body mass index correlated significantly with feces metabolites: urate, glutamate gamma-methyl ester, and 5-oxoproline. |
| Flannery et al., 2020 [21] | N = 40 children from a larger study | Mean age = 6.12 yrs (range 5–7). Female = 23 Country: U.S.A | Subsample from a midsize city in the Pacific Northwest of the U.S.A. already participating in a larger study of families, asked to participate in FU to collect stool samples (Cross-sectional study) Aim: determine how both the microbial taxa and the specific genetic functions they encode associate with subclinical child behavioral dysregulation symptoms (hereafter “behavioral dysregulation”), socioeconomic risk, and caregiver behavior | Single fecal sample collection Analysis of fecal samples:
| Questionnaires:
Other questionnaires: Gut history questionnaire filled out by parent | Significant association between functional composition and impulsivity. Bacteroides fragilis associated with reduction in:
|
| Michels et al., 2019 [22] | N total = 93 children/adolescents | Age = 8–16 yrs Country: Belgium | Cross-sectional study to investigate associations between gut microbiome and psychosocial stress in children/adolescents Children and adolescents recruited for the longitudinal ChiBS study (Michels et al., 2012) | Unique fecal sample: collected at home Analysis of fecal samples:
| Stress markers:
| Stress biomarkers: High stress: low pnn50 and more negative events; High pnn50: positively related to happiness. Alpha diversity:
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agusti, A.; Lamers, F.; Tamayo, M.; Benito-Amat, C.; Molina-Mendoza, G.V.; Penninx, B.W.J.H.; Sanz, Y. The Gut Microbiome in Early Life Stress: A Systematic Review. Nutrients 2023, 15, 2566. https://doi.org/10.3390/nu15112566
Agusti A, Lamers F, Tamayo M, Benito-Amat C, Molina-Mendoza GV, Penninx BWJH, Sanz Y. The Gut Microbiome in Early Life Stress: A Systematic Review. Nutrients. 2023; 15(11):2566. https://doi.org/10.3390/nu15112566
Chicago/Turabian StyleAgusti, Ana, Femke Lamers, Maria Tamayo, Carlos Benito-Amat, Gara V. Molina-Mendoza, Brenda W. J. H. Penninx, and Yolanda Sanz. 2023. "The Gut Microbiome in Early Life Stress: A Systematic Review" Nutrients 15, no. 11: 2566. https://doi.org/10.3390/nu15112566
APA StyleAgusti, A., Lamers, F., Tamayo, M., Benito-Amat, C., Molina-Mendoza, G. V., Penninx, B. W. J. H., & Sanz, Y. (2023). The Gut Microbiome in Early Life Stress: A Systematic Review. Nutrients, 15(11), 2566. https://doi.org/10.3390/nu15112566