
A Protective Role for Arachidonic Acid Metabolites against Advanced Colorectal Adenoma in a Phase III Trial of Selenium

 , ,
, ,  and
and
Abstract
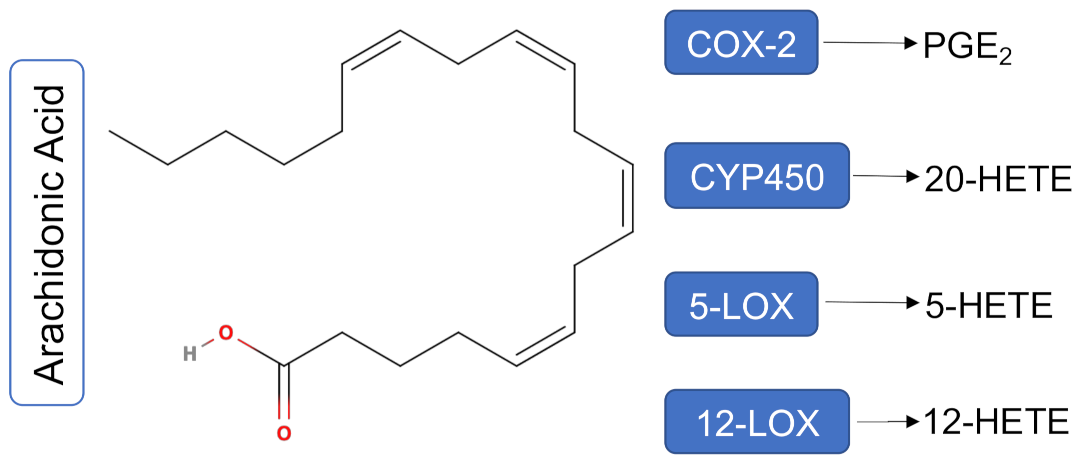
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Plasma Sample Collection and Preparation
2.3. Reverse Phase Chromatography with HPLC-MS
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nakanishi, M.; Rosenberg, D.W. Multifaceted roles of PGE2 in inflammation and cancer. Semin Immunopathol. 2013, 35, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.L.; Shu, L.; Zheng, P.F.; Zhang, X.Y.; Si, C.J.; Yu, X.L.; Gao, W.; Zhang, L. Dietary patterns and colorectal cancer risk: A meta-analysis. Eur. J. Cancer Prev. 2017, 26, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Randi, G.; Edefonti, V.; Ferraroni, M.; La Vecchia, C.; Decarli, A. Dietary patterns and the risk of colorectal cancer and adenomas. Nutr. Rev. 2010, 68, 389–408. [Google Scholar] [CrossRef]
- Wang, D.; DuBois, R.N. An inflammatory mediator, prostaglandin E2, in colorectal cancer. Cancer J. 2013, 19, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Shrubsole, M.J.; Cai, Q.; Wen, W.; Milne, G.; Smalley, W.E.; Chen, Z.; Ness, R.M.; Zheng, W. Urinary prostaglandin E2 metabolite and risk for colorectal adenoma. Cancer Prev. Res. 2012, 5, 336–342. [Google Scholar] [CrossRef]
- Bezawada, N.; Song, M.; Wu, K.; Mehta, R.S.; Milne, G.L.; Ogino, S.; Fuchs, C.S.; Giovannucci, E.L.; Chan, A.T. Urinary PGE-M levels are associated with risk of colorectal adenomas and chemopreventive response to anti-inflammatory drugs. Cancer Prev. Res. 2014, 7, 758–765. [Google Scholar] [CrossRef]
- Cai, Q.; Gao, Y.T.; Chow, W.H.; Shu, X.O.; Yang, G.; Ji, B.T.; Wen, W.; Rothman, N.; Li, H.L.; Morrow, J.D.; et al. Prospective study of urinary prostaglandin E2 metabolite and colorectal cancer risk. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2006, 24, 5010–5016. [Google Scholar] [CrossRef]
- Greenhough, A.; Smartt, H.J.; Moore, A.E.; Roberts, H.R.; Williams, A.C.; Paraskeva, C.; Kaidi, A. The COX-2/PGE2 pathway: Key roles in the hallmarks of cancer and adaptation to the tumour microenvironment. Carcinogenesis 2009, 30, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Armaghany, T.; Wilson, J.D.; Chu, Q.; Mills, G. Genetic alterations in colorectal cancer. Gastrointest Cancer Res. 2012, 5, 19–27. [Google Scholar]
- U.S. Preventive Services Task Force; Bibbins-Domingo, K.; Grossman, D.C.; Curry, S.J.; Davidson, K.W.; Epling, J.W., Jr.; Garcia, F.A.R.; Gillman, M.W.; Harper, D.M.; Kemper, A.R.; et al. Screening for Colorectal Cancer: US Preventive Services Task Force Recommendation Statement. JAMA 2016, 315, 2564–2575. [Google Scholar] [CrossRef]
- Pereira, C.; Queiros, S.; Galaghar, A.; Sousa, H.; Marcos-Pinto, R.; Pimentel-Nunes, P.; Brandao, C.; Medeiros, R.; Dinis-Ribeiro, M. Influence of Genetic Polymorphisms in Prostaglandin E2 Pathway (COX-2/HPGD/SLCO2A1/ABCC4) on the Risk for Colorectal Adenoma Development and Recurrence after Polypectomy. Clin. Transl. Gastroenterol. 2016, 7, e191. [Google Scholar] [CrossRef] [PubMed]
- Kleinstein, S.E.; Heath, L.; Makar, K.W.; Poole, E.M.; Seufert, B.L.; Slattery, M.L.; Xiao, L.; Duggan, D.J.; Hsu, L.; Curtin, K.; et al. Genetic variation in the lipoxygenase pathway and risk of colorectal neoplasia. Genes Chromosomes Cancer 2013, 52, 437–449. [Google Scholar] [CrossRef]
- Alexanian, A.; Sorokin, A. Targeting 20-HETE producing enzymes in cancer—rationale, pharmacology, and clinical potential. Onco Targets Ther. 2013, 6, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.A.; Ashbeck, E.L.; Roe, D.J.; Fales, L.; Buckmeier, J.; Wang, F.; Bhattacharyya, A.; Hsu, C.H.; Chow, H.H.; Ahnen, D.J.; et al. Selenium Supplementation for Prevention of Colorectal Adenomas and Risk of Associated Type 2 Diabetes. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.A.; Ashbeck, E.L.; Roe, D.J.; Fales, L.; Buckmeier, J.; Wang, F.; Bhattacharyya, A.; Hsu, C.H.; Chow, S.H.; Ahnen, D.J.; et al. Celecoxib for the Prevention of Colorectal Adenomas: Results of a Suspended Randomized Controlled Trial. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef]
- Thompson, P.; Roe, D.J.; Fales, L.; Buckmeier, J.; Wang, F.; Hamilton, S.R.; Bhattacharyya, A.; Green, S.; Hsu, C.H.; Chow, H.H.; et al. Design and baseline characteristics of participants in a phase III randomized trial of celecoxib and selenium for colorectal adenoma prevention. Cancer Prev. Res. 2012, 5, 1381–1393. [Google Scholar] [CrossRef]
- Bresalier, R.S.; Sandler, R.S.; Quan, H.; Bolognese, J.A.; Oxenius, B.; Horgan, K.; Lines, C.; Riddell, R.; Morton, D.; Lanas, A.; et al. Cardiovascular events associated with rofecoxib in a colorectal adenoma chemoprevention trial. N. Engl. J. Med. 2005, 352, 1092–1102. [Google Scholar] [CrossRef]
- Solomon, S.D.; Wittes, J.; Finn, P.V.; Fowler, R.; Viner, J.; Bertagnolli, M.M.; Arber, N.; Levin, B.; Meinert, C.L.; Martin, B.; et al. Cardiovascular risk of celecoxib in 6 randomized placebo-controlled trials: The cross trial safety analysis. Circulation 2008, 117, 2104–2113. [Google Scholar] [CrossRef]
- Yang, J.; Schmelzer, K.; Georgi, K.; Hammock, B.D. Quantitative profiling method for oxylipin metabolome by liquid chromatography electrospray ionization tandem mass spectrometry. Anal. Chem. 2009, 81, 8085–8093. [Google Scholar] [CrossRef]
- Zivkovic, A.M.; Yang, J.; Georgi, K.; Hegedus, C.; Nording, M.L.; O’Sullivan, A.; German, J.B.; Hogg, R.J.; Weiss, R.H.; Bay, C.; et al. Serum oxylipin profiles in IgA nephropathy patients reflect kidney functional alterations. Metabolomics 2012, 8, 1102–1113. [Google Scholar] [CrossRef][Green Version]
- Keizer, R.J.; Jansen, R.S.; Rosing, H.; Thijssen, B.; Beijnen, J.H.; Schellens, J.H.; Huitema, A.D. Incorporation of concentration data below the limit of quantification in population pharmacokinetic analyses. Pharmacol. Res. Perspect. 2015, 3, e00131. [Google Scholar] [CrossRef]
- Whitcomb, B.W.; Schisterman, E.F. Assays with lower detection limits: Implications for epidemiological investigations. Paediatr. Perinat. Epidemiol. 2008, 22, 597–602. [Google Scholar] [CrossRef]
- Baines, A.; Taylor-Parker, M.; Goulet, A.C.; Renaud, C.; Gerner, E.W.; Nelson, M.A. Selenomethionine inhibits growth and suppresses cyclooxygenase-2 (COX-2) protein expression in human colon cancer cell lines. Cancer Biol. Ther. 2002, 1, 370–374. [Google Scholar] [CrossRef]
- Johnson, J.C.; Schmidt, C.R.; Shrubsole, M.J.; Billheimer, D.D.; Joshi, P.R.; Morrow, J.D.; Heslin, M.J.; Washington, M.K.; Ness, R.M.; Zheng, W.; et al. Urine PGE-M: A metabolite of prostaglandin E2 as a potential biomarker of advanced colorectal neoplasia. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2006, 4, 1358–1365. [Google Scholar] [CrossRef] [PubMed]
- Davenport, J.R.; Cai, Q.; Ness, R.M.; Milne, G.; Zhao, Z.; Smalley, W.E.; Zheng, W.; Shrubsole, M.J. Evaluation of pro-inflammatory markers plasma C-reactive protein and urinary prostaglandin-E2 metabolite in colorectal adenoma risk. Mol. Carcinog 2016, 55, 1251–1261. [Google Scholar] [CrossRef]
- Ghadimi, R.; Kuriki, K.; Tsuge, S.; Takeda, E.; Imaeda, N.; Suzuki, S.; Sawai, A.; Takekuma, K.; Hosono, A.; Tokudome, Y.; et al. Serum concentrations of fatty acids and colorectal adenoma risk: A case-control study in Japan. Asian Pac. J. Cancer Prev. 2008, 9, 111–118. [Google Scholar] [PubMed]
- Hall, M.N.; Campos, H.; Li, H.; Sesso, H.D.; Stampfer, M.J.; Willett, W.C.; Ma, J. Blood levels of long-chain polyunsaturated fatty acids, aspirin, and the risk of colorectal cancer. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 314–321. [Google Scholar] [CrossRef]
- Kojima, M.; Wakai, K.; Tokudome, S.; Suzuki, K.; Tamakoshi, K.; Watanabe, Y.; Kawado, M.; Hashimoto, S.; Hayakawa, N.; Ozasa, K.; et al. Serum levels of polyunsaturated fatty acids and risk of colorectal cancer: A prospective study. Am. J. Epidemiol. 2005, 161, 462–471. [Google Scholar] [CrossRef]
- Pot, G.K.; Geelen, A.; van Heijningen, E.M.; Siezen, C.L.; van Kranen, H.J.; Kampman, E. Opposing associations of serum n-3 and n-6 polyunsaturated fatty acids with colorectal adenoma risk: An endoscopy-based case-control study. Int. J. Cancer 2008, 123, 1974–1977. [Google Scholar] [CrossRef] [PubMed]
- Rifkin, S.B.; Shrubsole, M.J.; Cai, Q.; Smalley, W.E.; Ness, R.M.; Swift, L.L.; Zheng, W.; Murff, H.J. PUFA levels in erythrocyte membrane phospholipids are differentially associated with colorectal adenoma risk. Br. J. Nutr. 2017, 117, 1615–1622. [Google Scholar] [CrossRef]
- Austin Pickens, C.; Yin, Z.; Sordillo, L.M.; Fenton, J.I. Arachidonic acid-derived hydroxyeicosatetraenoic acids are positively associated with colon polyps in adult males: A cross-sectional study. Sci. Rep. 2019, 9, 12033. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.T.; Kim, Y.M.; Surh, Y.J.; Baik, H.W.; Lee, S.K.; Ha, J.; Park, O.J. Selenium regulates cyclooxygenase-2 and extracellular signal-regulated kinase signaling pathways by activating AMP-activated protein kinase in colon cancer cells. Cancer Res. 2006, 66, 10057–10063. [Google Scholar] [CrossRef] [PubMed]
- Vunta, H.; Davis, F.; Palempalli, U.D.; Bhat, D.; Arner, R.J.; Thompson, J.T.; Peterson, D.G.; Reddy, C.C.; Prabhu, K.S. The anti-inflammatory effects of selenium are mediated through 15-deoxy-Delta12,14-prostaglandin J2 in macrophages. J. Biol. Chem. 2007, 282, 17964–17973. [Google Scholar] [CrossRef]
- Duntas, L.H. Selenium and inflammation: Underlying anti-inflammatory mechanisms. Horm. Metab. Res. 2009, 41, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Stahle, J.A.; Vunta, H.; Channa Reddy, C.; Sandeep Prabhu, K. Regulation of expression of apolipoprotein A-I by selenium status in human liver hepatoblastoma cells. Eur. J. Nutr. 2009, 48, 283–290. [Google Scholar] [CrossRef]
- Rao, C.V.; Cooma, I.; Rodriguez, J.G.; Simi, B.; El-Bayoumy, K.; Reddy, B.S. Chemoprevention of familial adenomatous polyposis development in the APC(min) mouse model by 1,4-phenylene bis(methylene)selenocyanate. Carcinogenesis 2000, 21, 617–621. [Google Scholar] [CrossRef]
- Qian, F.; Misra, S.; Prabhu, K.S. Selenium and selenoproteins in prostanoid metabolism and immunity. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 484–516. [Google Scholar] [CrossRef]
- Ghosh, J. Rapid induction of apoptosis in prostate cancer cells by selenium: Reversal by metabolites of arachidonate 5-lipoxygenase. Biochem. Biophys. Res. Commun 2004, 315, 624–635. [Google Scholar] [CrossRef]


{kind=link}
| Non-Advanced | Advanced | p-Value | |
|---|---|---|---|
| n = 130 | n = 126 | ||
| Age (years) a | 63.3 (8.7) | 61.6 (8.3) | 0.12 |
| Sex b | 0.15 | ||
| Female | 48 (36.9%) | 36 (28.6%) | |
| Male | 82 (63.1%) | 90 (71.4%) | |
| Highest level of education b | 0.94 | ||
| High School or less | 25 (19.2%) | 22 (17.5%) | |
| Some College | 37 (28.5%) | 40 (31.7%) | |
| Bachelor’s Degree | 30 (23.1%) | 29 (23.0%) | |
| Graduate or Professional Degree | 38 (29.2%) | 35 (27.8%) | |
| Baseline Body Mass Index (kg/m2) a | 28.8 (5.2) | 28.7 (4.4) | 0.78 |
| Smoke cigarettes (≥100) b | 0.30 | ||
| Current | 8 (6.3%) | 13 (10.7%) | |
| Never | 58 (45.7%) | 46 (38.0%) | |
| Past | 61 (48.0%) | 62 (51.2%) | |
| Routine NSAID use b | 6 (4.6%) | 4 (3.2%) | 0.55 |
| Routine Aspirin use b | 60 (46.2%) | 49 (38.9%) | 0.24 |
| CRC in a first-degree relative b | 0.18 | ||
| None | 96 (76.8%) | 102 (83.6%) | |
| One or more | 29 (23.2%) | 20 (16.4%) | |
| History of polyps b | 49 (38.6%) | 39 (31.2%) | 0.22 |
| History of cancer b,d | 8 (6.2%) | 5 (4.0%) | 0.43 |
| Randomization c | 0.62 | ||
| Placebo | 69 (53.1%) | 62 (49.2%) | |
| Selenium | 61 (46.9%) | 64 (50.8%) | |
| Serum Selenium Concentration (ng/mL) a | 140.0 (26.2) | 132.5 (22.3) | 0.014 |
| Dietary Omega 6 (mg/day) a | 14.9 (9.1) | 13.6 (9.9) | 0.28 |
| Dietary Omega 3 (mg/day) a | 1.5 (0.7) | 1.3 (0.7) | 0.16 |
| Dietary Omega6/Omega3 Ratio) a | 9.9 (1.8) | 9.9 (2.4) | 0.97 |
| Category of Oxylipin Metabolite | Advanced n = 126 | Size ≥1 cm n = 115 | Villous Histology n = 47 | Multiplicity n = 50 |
|---|---|---|---|---|
| PGE2 (pg/mL) | OR (95% CI) | |||
| 1 (0.00–18.0) | Ref | Ref | Ref | Ref |
| 2 (18.1–277.5) | 0.55 (0.33–0.92) | 0.52 (0.31–0.87) | 0.90 (0.47–1.72) | 0.78 (0.41–1.48) |
| 20-HETE (pg/mL) | ||||
| 1 (0.0–5.0) | Ref | Ref | Ref | Ref |
| 2 (5.1–20.3) | 0.95 (0.57–1.57) | 0.97 (0.58–1.61) | 1.15 (0.60–2.19) | 0.92 (0.49–1.74) |
| 12-HETE (pg/mL) | ||||
| 1 (0.0–24.5) | Ref | Ref | Ref | Ref |
| 2 (24.6–718.7) | 0.66 (0.40–1.09) | 0. 84 (0.51–1.40) | 0.71 (0.37–1.36) | 0.80 (0.43–1.51) |
| 5-HETE | ||||
| 1 (0.0–46.6) | Ref | Ref | Ref | Ref |
| 2 (46.7–301.2) | 0.53 (0.33–0.94) | 0.61 (0.37–1.02) | 0.37 (0.19–0.75) | 0.88 (0.47–1.67) |
| Oxylipin | Overall a n = 256 (OR, 95% CI) d | Selenium b n = 125 (OR, 95% CI) | Placebo c n = 131 (OR, 95% CI) | |||
|---|---|---|---|---|---|---|
| Category of Metabolite Concentration | Non-Advanced | Advanced | Non-Advanced | Advanced | Non-Advanced | Advanced |
| PGE2 (pg/mL) | ||||||
| 1 (−2.8 to −0.0002) | Ref | Ref | Ref | Ref | Ref | Ref |
| 2 (−0.0003 to 14.2) | 1.41 (0.85–2.00) | 1.46 (0.54–3.97) | 1.54 (0.67–3.49) | 0.64 (0.20–2.04) | 1.24 (0.55–2.80) | 0.72 (0.22–2.28) |
| 20-HETE (pg/mL) | ||||||
| 1 (−0.004 to 0.00) | Ref | Ref | Ref | Ref | Ref | Ref |
| 2 (0.01 to 11.9) | 0.98 (0.54–1.77) | 1.54 (0.69–3.45) | 1.01 (0.43–2.36) | 1.95 (0.61–6.25) | 0.95 (0.40–2.19) | 1.16 (0.36–2.49) |
| 12-HETE (pg/mL) | ||||||
| 1 (−57.1 to 0.0) | Ref | Ref | Ref | Ref | Ref | Ref |
| 2 (0.01 to 80.3) | 1.40 (0.74–2.63) | 1.30 (0.45–3.13) | 1.09 (0.46–2.58) | 0.83 (0.25–2.81) | 1.55 (0.58–4.14) | 2.29 (0.59–8.90) |
| 5-HETE (pg/mL) | ||||||
| 1 (−300.8 to 0.00) | Ref | Ref | Ref | Ref | Ref | Ref |
| 2 (0.01 to 2583.5) | 1.21 (0.66–2.21) | 1.33 (0.58–3.03) | 0.97 (0.40–2.37) | 1.67 (0.53–5.3) | 1.62 (0.32–3.67) | 1.09 (0.32–3.69) |
| Mean ± SD | Baselinen n = 256 | Follow-Up a n = 253 | Differencen n = 253 | p-Value b |
|---|---|---|---|---|
| PGE2 c,d | 0.08 ± 0.32 | 0.48 ± 1.4 | 0.39 ± 1.38 | <0.001 |
| 20-HETE | 0.05 ± 0.04 | 0.05 ± 0.04 | 0.00 ± 0.03 | 0.68 |
| 12-HETE | 1.4 ± 5.86 | 3.90 ± 12.0 | 2.48 ± 12.13 | 0.001 |
| 5-HETE | 3.82 ± 25.5 | 64.17 ± 285.16 | 60.32 ± 282.31 | <0.001 |
| Mean ± SD | Placebo a n = 123 | Selenium a n = 130 | p-Value a |
|---|---|---|---|
| PGE2 b,c | 0.46 ± 1.6 | 0.32 ± 1.07 | 0.43 |
| 20-HETE | 0.00 ± 0.04 | 0.00 ± 0.03 | 0.66 |
| 12-HETE | 3.30 ± 1.2 | 1.70 ± 10.7 | 0.30 |
| 5-HETE | 99.1 ± 381.9 | 19.3 ± 84.1 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez, J.A.; Skiba, M.B.; Chow, H.-H.S.; Chew, W.M.; Saboda, K.; Lance, P.; Ellis, N.A.; Jacobs, E.T. A Protective Role for Arachidonic Acid Metabolites against Advanced Colorectal Adenoma in a Phase III Trial of Selenium. Nutrients 2021, 13, 3877. https://doi.org/10.3390/nu13113877
Martinez JA, Skiba MB, Chow H-HS, Chew WM, Saboda K, Lance P, Ellis NA, Jacobs ET. A Protective Role for Arachidonic Acid Metabolites against Advanced Colorectal Adenoma in a Phase III Trial of Selenium. Nutrients. 2021; 13(11):3877. https://doi.org/10.3390/nu13113877
Chicago/Turabian StyleMartinez, Jessica A., Meghan B. Skiba, H-H. Sherry Chow, Wade M. Chew, Kathylynn Saboda, Peter Lance, Nathan A. Ellis, and Elizabeth T. Jacobs. 2021. "A Protective Role for Arachidonic Acid Metabolites against Advanced Colorectal Adenoma in a Phase III Trial of Selenium" Nutrients 13, no. 11: 3877. https://doi.org/10.3390/nu13113877
APA StyleMartinez, J. A., Skiba, M. B., Chow, H.-H. S., Chew, W. M., Saboda, K., Lance, P., Ellis, N. A., & Jacobs, E. T. (2021). A Protective Role for Arachidonic Acid Metabolites against Advanced Colorectal Adenoma in a Phase III Trial of Selenium. Nutrients, 13(11), 3877. https://doi.org/10.3390/nu13113877