Single Nucleotide Polymorphisms in Vitamin D Receptor Gene Affect Birth Weight and the Risk of Preterm Birth: Results From the “Mamma & Bambino” Cohort and A Meta-Analysis

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.1.1. VDR Genotyping
2.1.2. Assessment of Vitamin D Intake
2.1.3. Statistical Analyses
2.2. Systematic Review and Meta-Analysis
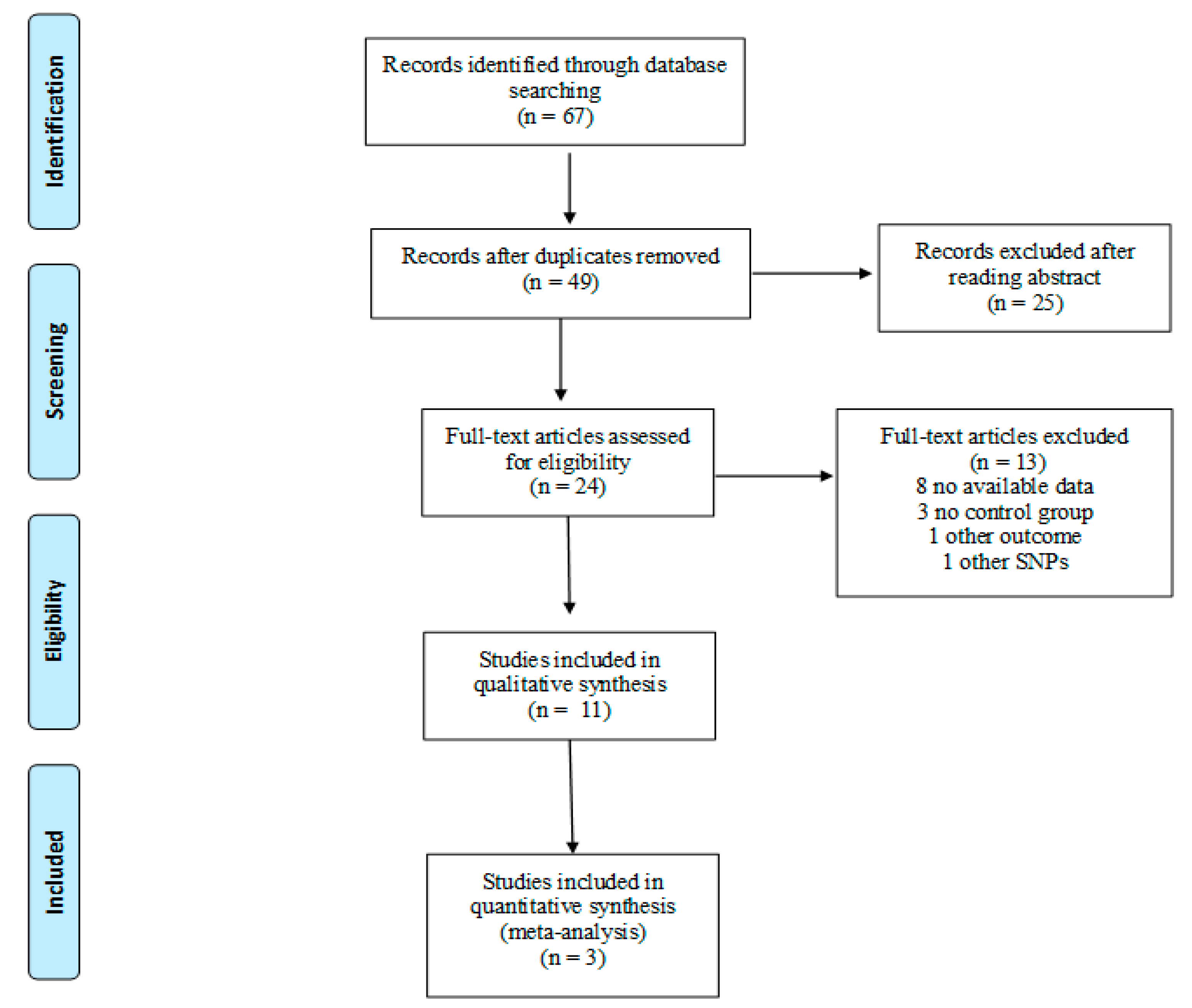
2.2.1. Search Strategy
2.2.2. Selection Criteria
2.2.3. Study Selection and Data Extraction
2.2.4. Procedures of Meta-Analysis
3. Results
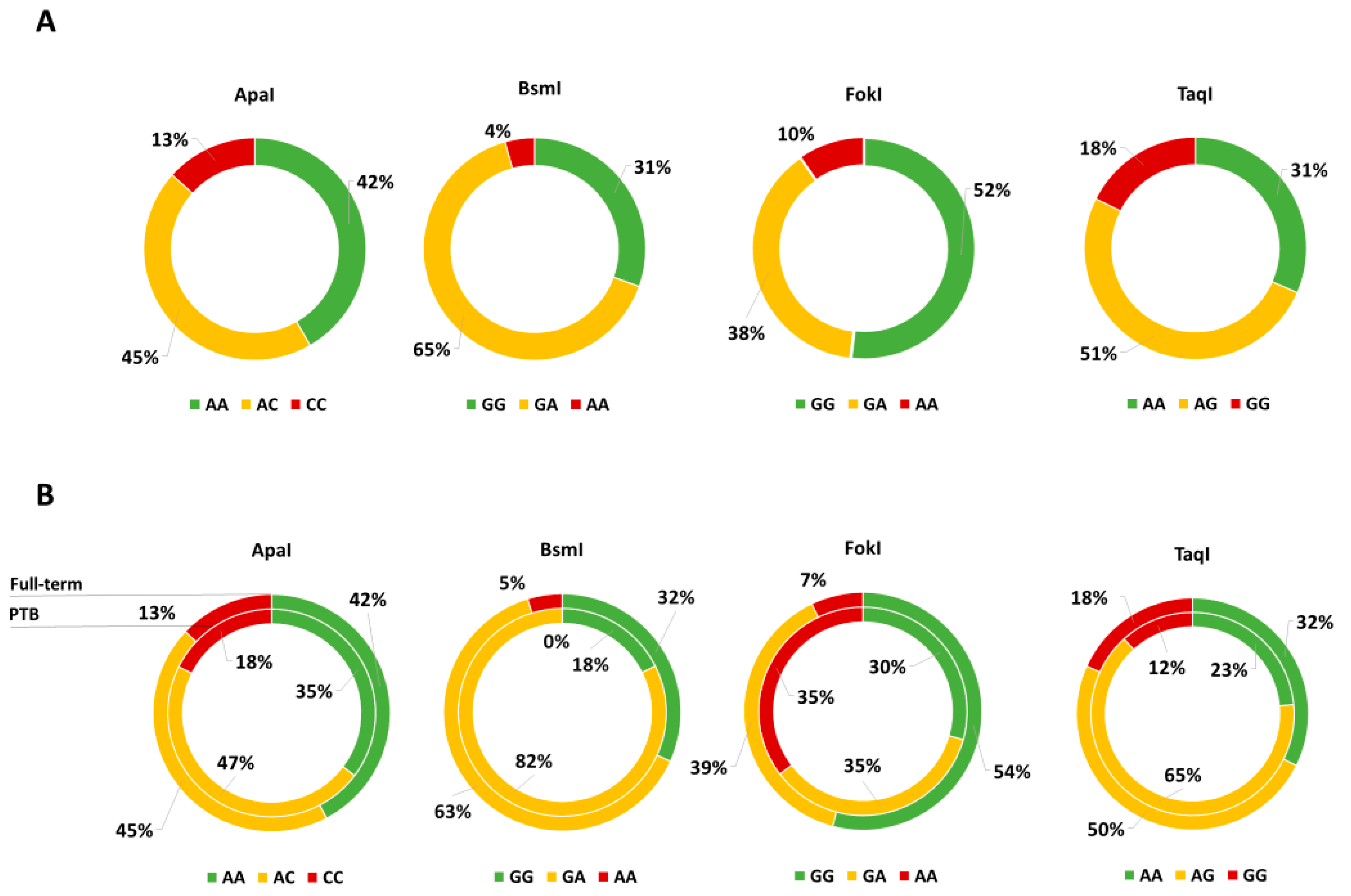
3.1. “Mamma & Bambino” Cohort
3.2. Systematic Review
3.2.1. Study Characteristics
3.2.2. VDR Polymorphisms and Neonatal Anthropometric Measures
3.2.3. VDR Polymorphisms and PTB Risk
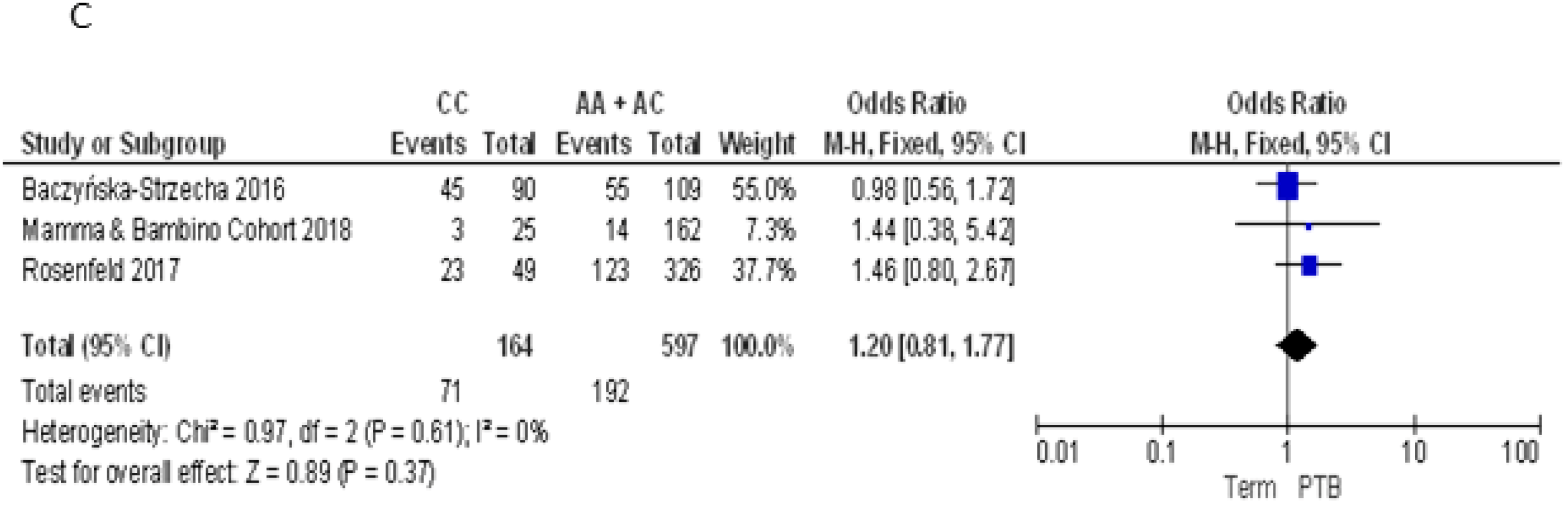
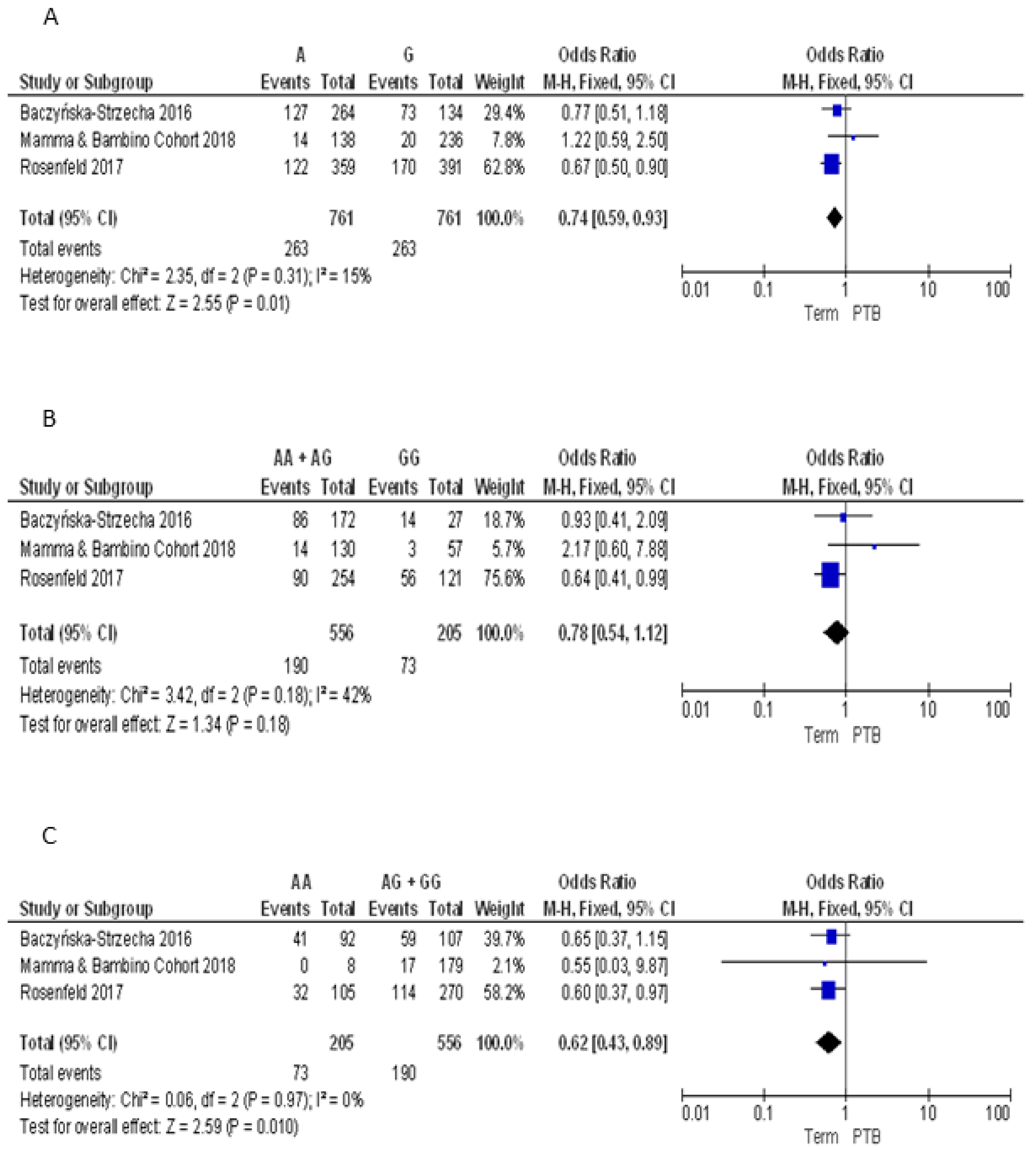
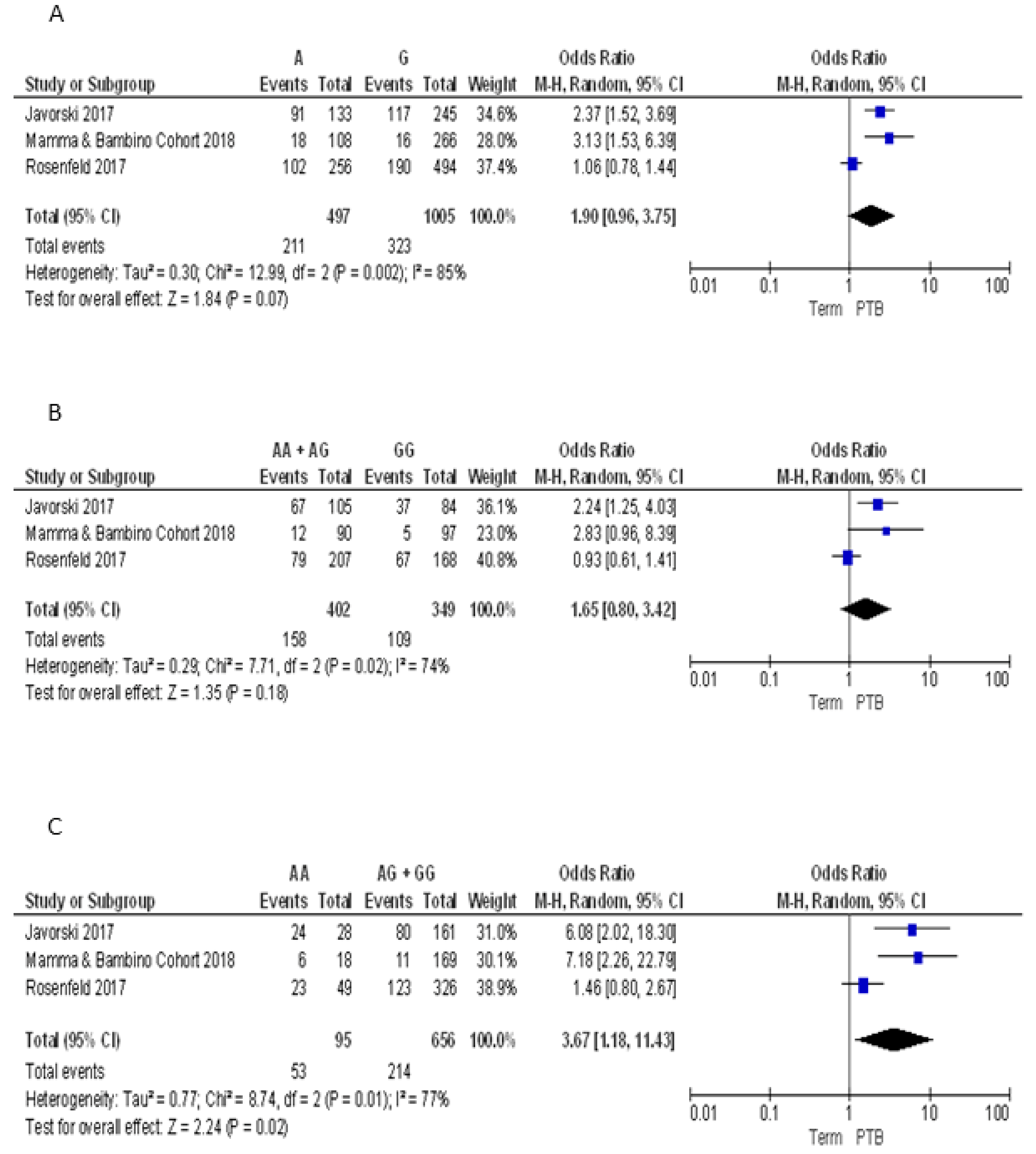
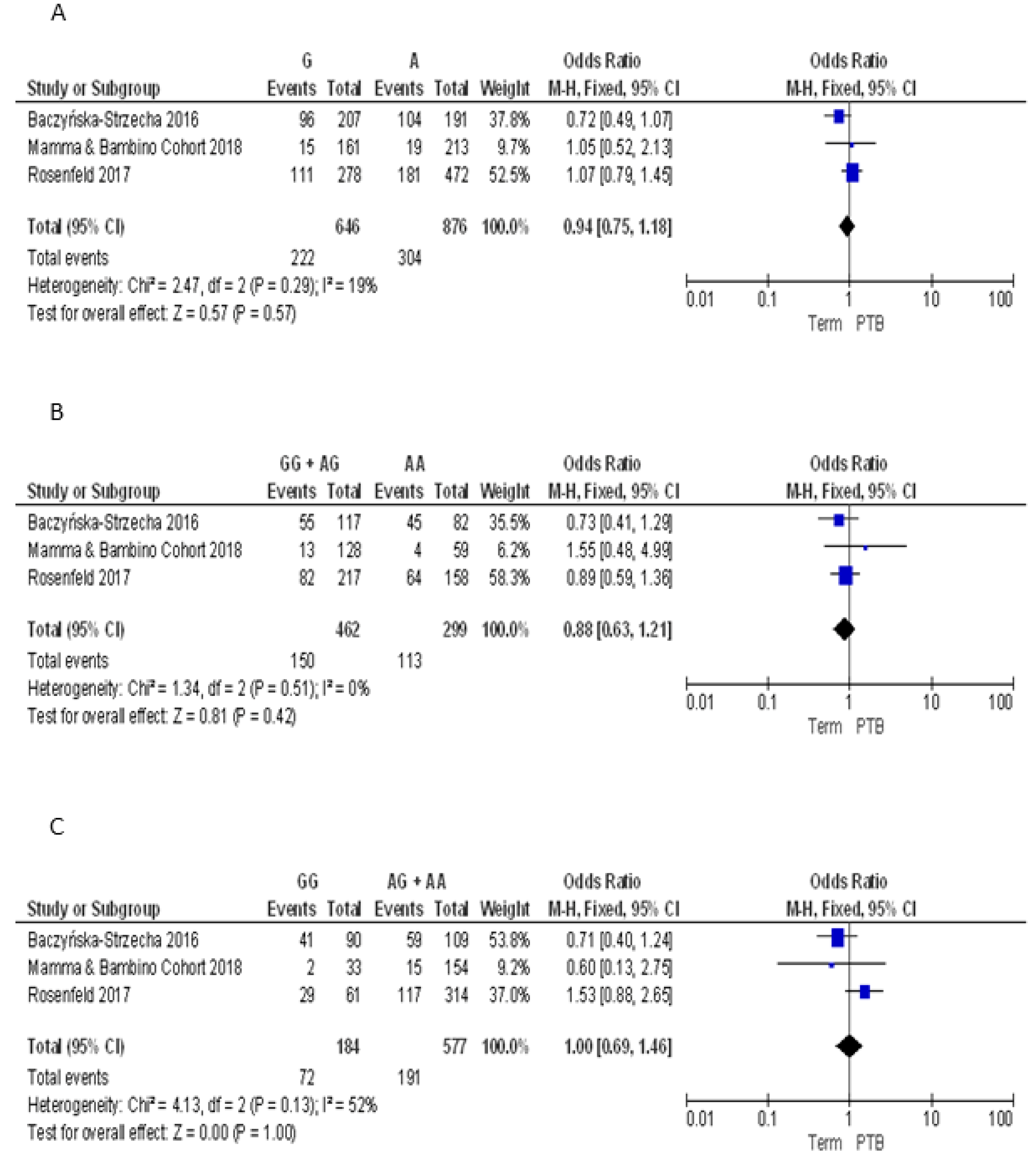
3.2.4. Meta-Analyses of the Association between VDR Polymorphisms and PTB Risk
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chiavaroli, V.; Castorani, V.; Guidone, P.; Derraik, J.G.; Liberati, M.; Chiarelli, F.; Mohn, A. Incidence of infants born small- and large-for-gestational-age in an italian cohort over a 20-year period and associated risk factors. Ital. J. Pediatr. 2016, 42, 42. [Google Scholar] [CrossRef] [PubMed]
- Lawn, J.E.; Blencowe, H.; Pattinson, R.; Cousens, S.; Kumar, R.; Ibiebele, I.; Gardosi, J.; Day, L.T.; Stanton, C. Stillbirths: Where? When? Why? How to make the data count? Lancet 2011, 377, 1448–1463. [Google Scholar] [CrossRef]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef]
- Howson, C.P.; Kinney, M.V.; McDougall, L.; Lawn, J.E. Born too soon: Preterm birth matters. Reprod. Health 2013, 10 (Suppl. 1), S1. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.P.; Chawanpaiboon, S.; Moller, A.B.; Watananirun, K.; Bonet, M.; Lumbiganon, P. The global epidemiology of preterm birth. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Vitale, S.G.; Marilli, I.; Rapisarda, A.M.; Rossetti, D.; Belluomo, G.; Iapichino, V.; Stancanelli, F.; Cianci, A. Cellular and biochemical mechanisms, risk factors and management of preterm birth: State of the art. Minerva Ginecol. 2014, 66, 589–595. [Google Scholar] [PubMed]
- Giunta, G.; Giuffrida, L.; Mangano, K.; Fagone, P.; Cianci, A. Influence of lactoferrin in preventing preterm delivery: A pilot study. Mol. Med. Rep. 2012, 5, 162–166. [Google Scholar] [PubMed]
- Pino, A.; Giunta, G.; Randazzo, C.L.; Caruso, S.; Caggia, C.; Cianci, A. Bacterial biota of women with bacterial vaginosis treated with lactoferrin: An open prospective randomized trial. Microb. Ecol. Health Dis. 2017, 28, 1357417. [Google Scholar] [CrossRef] [PubMed]
- Baczyńska-Strzecha, M.; Kalinka, J. Influence of apa1 (rs7975232), taq1 (rs731236) and bsm1 (rs154410) polymorphisms of vitamin d receptor on preterm birth risk in the polish population. Ginekol. Pol. 2016, 87, 763–768. [Google Scholar] [CrossRef] [PubMed]
- York, T.P.; Eaves, L.J.; Neale, M.C.; Strauss, J.F. The contribution of genetic and environmental factors to the duration of pregnancy. Am. J. Obstet. Gynecol. 2014, 210, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Black, R.E. Global prevalence of small for gestational age births. Nestle Nutr. Inst. Workshop Ser. 2015, 81, 1–7. [Google Scholar] [PubMed]
- Chavarro, J.E.; Rich-Edwards, J.W.; Rosner, B.A.; Willett, W.C. Iron intake and risk of ovulatory infertility. Obstet. Gynecol. 2006, 108, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, E.; Gremy, I.; Lang, J.M.; Cohen, A.P. Low birthweight at term and the timing of fetal exposure to maternal smoking. Am. J. Public Health 1994, 84, 1127–1131. [Google Scholar] [CrossRef] [PubMed]
- Pietrantoni, M.; Knuppel, R.A. Alcohol use in pregnancy. Clin. Perinatol. 1991, 18, 93–111. [Google Scholar] [CrossRef]
- Fowles, E.R. Prenatal nutrition and birth outcomes. J. Obstet. Gynecol. Neonatal. Nurs. 2004, 33, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.L.; Muyayalo, K.P.; Zhang, Y.H.; Hu, X.H.; Liao, A.H. Immunological function of vitamin D during human pregnancy. Am. J. Reprod. Immunol. 2017, 78. [Google Scholar] [CrossRef] [PubMed]
- Kiely, M.; Hemmingway, A.; O’Callaghan, K.M. Vitamin D in pregnancy: Current perspectives and future directions. Ther. Adv. Musculoskelet. Dis. 2017, 9, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D: A D-lightful solution for health. J. Investig. Med. 2011, 59, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine. Dietary Reference Intakes for Calcium and Vitamin D; The National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- U.S. Department of Agriculture, Agricultural Research Service (ARS). USDA National Nutrient Database for Standard Reference. Available online: https://www.ars.usda.gov/northeast-area/beltsville-md-bhnrc/beltsville-human-nutrition-research-center/nutrient-data-laboratory/docs/usda-national-nutrient-database-for-standard-reference/ (accessed on 1 May 2018).
- Ovesen, L.; Brot, C.; Jakobsen, J. Food contents and biological activity of 25-hydroxyvitamin D: A vitamin D metabolite to be reckoned with? Ann. Nutr. Metab. 2003, 47, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.S.; Whiting, S.J.; Barton, C.N. Vitamin D fortification in the United States and Canada: Current status and data needs. Am. J. Clin. Nutr. 2004, 80, 1710S–1716S. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D: A millenium perspective. J. Cell. Biochem. 2003, 88, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.W. The history of the discovery of vitamin D and its daughter steroid hormone. Ann. Nutr. Metab. 2012, 61, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Harvey, N.C.; Holroyd, C.; Ntani, G.; Javaid, K.; Cooper, P.; Moon, R.; Cole, Z.; Tinati, T.; Godfrey, K.; Dennison, E.; et al. Vitamin D supplementation in pregnancy: A systematic review. Health Technol. Assess. 2014, 18, 1–190. [Google Scholar] [CrossRef] [PubMed]
- De-Regil, L.M.; Palacios, C.; Lombardo, L.K.; Peña-Rosas, J.P. Vitamin D supplementation for women during pregnancy. Sao Paulo Med. J. 2016, 134, 274–275. [Google Scholar] [CrossRef] [PubMed]
- Pérez-López, F.R.; Pasupuleti, V.; Mezones-Holguin, E.; Benites-Zapata, V.A.; Thota, P.; Deshpande, A.; Hernandez, A.V. Effect of vitamin D supplementation during pregnancy on maternal and neonatal outcomes: A systematic review and meta-analysis of randomized controlled trials. Fertil. Steril. 2015, 103, 1278–1288.e4. [Google Scholar] [CrossRef] [PubMed]
- Thorne-Lyman, A.; Fawzi, W.W. Vitamin D during pregnancy and maternal, neonatal and infant health outcomes: A systematic review and meta-analysis. Paediatr. Perinat. Epidemiol. 2012, 26 (Suppl. 1), 75–90. [Google Scholar] [CrossRef] [PubMed]
- Strugnell, S.A.; Deluca, H.F. The vitamin d receptor-structure and transcriptional activation. Proc. Soc. Exp. Biol. Med. 1997, 215, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, J.; DeLuca, H.F. Where is the vitamin D receptor? Arch. Biochem. Biophys. 2012, 523, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Freedman, L.P.; Arce, V.; Perez Fernandez, R. DNA sequences that act as high affinity targets for the vitamin D3 receptor in the absence of the retinoid x receptor. Mol. Endocrinol. 1994, 8, 265–273. [Google Scholar] [PubMed]
- Karras, S.N.; Wagner, C.L.; Castracane, V.D. Understanding vitamin d metabolism in pregnancy: From physiology to pathophysiology and clinical outcomes. Metabolism 2018, 86, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Javorski, N.; Lima, C.A.D.; Silva, L.V.C.; Crovella, S.; de Azêvedo Silva, J. Vitamin D receptor (VDR) polymorphisms are associated to spontaneous preterm birth and maternal aspects. Gene 2018, 642, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, T.; Salem, H.; Altarescu, G.; Grisaru-Granovsky, S.; Tevet, A.; Birk, R. Maternal-fetal vitamin D receptor polymorphisms significantly associated with preterm birth. Arch. Gynecol. Obstet. 2017, 296, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Workalemahu, T.; Badon, S.E.; Dishi-Galitzky, M.; Qiu, C.; Williams, M.A.; Sorensen, T.; Enquobahrie, D.A. Placental genetic variations in vitamin D metabolism and birthweight. Placenta 2017, 50, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Manzon, L.; Altarescu, G.; Tevet, A.; Schimmel, M.S.; Elstein, D.; Samueloff, A.; Grisaru-Granovsky, S. Vitamin D receptor polymorphism foki is associated with spontaneous idiopathic preterm birth in an israeli population. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 177, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Swamy, G.K.; Garrett, M.E.; Miranda, M.L.; Ashley-Koch, A.E. Maternal vitamin D receptor genetic variation contributes to infant birthweight among black mothers. Am. J. Med. Genet. A 2011, 155A, 1264–1271. [Google Scholar] [CrossRef] [PubMed]
- Lorentzon, M.; Lorentzon, R.; Nordström, P. Vitamin D receptor gene polymorphism is associated with birth height, growth to adolescence, and adult stature in healthy Caucasian men: A cross-sectional and longitudinal study. J. Clin. Endocrinol. Metab. 2000, 85, 1666–1670. [Google Scholar] [PubMed]
- Tao, C.; Yu, T.; Garnett, S.; Briody, J.; Knight, J.; Woodhead, H.; Cowell, C.T. Vitamin D receptor alleles predict growth and bone density in girls. Arch. Dis. Child. 1998, 79, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Suarez, F.; Zeghoud, F.; Rossignol, C.; Walrant, O.; Garabédian, M. Association between vitamin d receptor gene polymorphism and sex-dependent growth during the first two years of life. J. Clin. Endocrinol. Metab. 1997, 82, 2966–2970. [Google Scholar] [CrossRef] [PubMed]
- Silvano, L.; Miras, M.; Pérez, A.; Picotto, G.; Díaz de Barboza, G.; Muñoz, L.; Martin, S.; Sobrero, G.; Armelini, P.; Mericq, V.; et al. Comparative analysis of clinical, biochemical and genetic aspects associated with bone mineral density in small for gestational age children. J. Pediatr. Endocrinol. Metab. 2011, 24, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Suarez, F.; Rossignol, C.; Garabédian, M. Interactive effect of estradiol and vitamin D receptor gene polymorphisms as a possible determinant of growth in male and female infants. J. Clin. Endocrinol. Metab. 1998, 83, 3563–3568. [Google Scholar] [CrossRef] [PubMed]
- Knabl, J.; Vattai, A.; Ye, Y.; Jueckstock, J.; Hutter, S.; Kainer, F.; Mahner, S.; Jeschke, U. Role of placental VDR expression and function in common late pregnancy disorders. Int. J. Mol. Sci. 2017, 18, E2340. [Google Scholar] [CrossRef] [PubMed]
- Agodi, A.; Barchitta, M.; Quattrocchi, A.; Maugeri, A.; Vinciguerra, M. Dapk1 promoter methylation and cervical cancer risk: A systematic review and a meta-analysis. PLoS ONE 2015, 10, e0135078. [Google Scholar] [CrossRef] [PubMed]
- Agodi, A.; Barchitta, M.; Quattrocchi, A.; Maugeri, A.; Canto, C.; Marchese, A.E.; Vinciguerra, M. Low fruit consumption and folate deficiency are associated with line-1 hypomethylation in women of a cancer-free population. Genes Nutr. 2015, 10, 480. [Google Scholar] [CrossRef] [PubMed]
- Barchitta, M.; Quattrocchi, A.; Maugeri, A.; Vinciguerra, M.; Agodi, A. Line-1 hypomethylation in blood and tissue samples as an epigenetic marker for cancer risk: A systematic review and meta-analysis. PLoS ONE 2014, 9, e109478. [Google Scholar] [CrossRef] [PubMed]
- Barchitta, M.; Quattrocchi, A.; Maugeri, A.; Canto, C.; La Rosa, N.; Cantarella, M.A.; Spampinato, G.; Scalisi, A.; Agodi, A. Line-1 hypermethylation in white blood cell dna is associated with high-grade cervical intraepithelial neoplasia. BMC Cancer 2017, 17, 601. [Google Scholar] [CrossRef] [PubMed]
- Barchitta, M.; Maugeri, A.; Quattrocchi, A.; Agrifoglio, O.; Agodi, A. The role of mirnas as biomarkers for pregnancy outcomes: A comprehensive review. Int. J. Genomics 2017, 2017, 8067972. [Google Scholar] [CrossRef] [PubMed]
- Physical status: The use and interpretation of anthropometry: Report of a WHO expert committee. World Health Organ. Tech. Rep. Ser. 1995, 854, 1–452.
- Moore Simas, T.A.; Waring, M.E.; Sullivan, G.M.; Liao, X.; Rosal, M.C.; Hardy, J.R.; Berry, R.E. Institute of medicine 2009 gestational weight gain guideline knowledge: Survey of obstetrics/gynecology and family medicine residents of the united states. Birth 2013, 40, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Bertino, E.; Spada, E.; Occhi, L.; Coscia, A.; Giuliani, F.; Gagliardi, L.; Gilli, G.; Bona, G.; Fabris, C.; De Curtis, M.; et al. Neonatal anthropometric charts: The Italian neonatal study compared with other European studies. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Agodi, A.; Barchitta, M.; Valenti, G.; Marzagalli, R.; Frontini, V.; Marchese, A.E. Increase in the prevalence of the mthfr 677 tt polymorphism in women born since 1959: Potential implications for folate requirements. Eur. J. Clin. Nutr. 2011, 65, 1302–1308. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.; Green, S. Cochrane Handbook for Systematic Reviews of Interventions, Version 5.1.0 ed. 2008. Available online: https://handbook-5-1.cochrane.org/ (accessed on 1 March 2018).
- Begg, C.B.; Mazumdar, M. Operating characteristics of a rank correlation test for publication bias. Biometrics 1994, 50, 1088–1101. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Davey Smith, G.; Schneider, M.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.S.; Choi, M.Y.; Longtine, M.S.; Nelson, D.M. Vitamin D effects on pregnancy and the placenta. Placenta 2010, 31, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.N.; Bulmer, J.N.; Kilby, M.D.; Hewison, M. Vitamin D and placental-decidual function. J. Soc. Gynecol. Investig. 2004, 11, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Stephanou, A.; Ross, R.; Handwerger, S. Regulation of human placental lactogen expression by 1,25-dihydroxyvitamin d3. Endocrinology 1994, 135, 2651–2656. [Google Scholar] [CrossRef] [PubMed]
- Avila, E.; Díaz, L.; Barrera, D.; Halhali, A.; Méndez, I.; González, L.; Zuegel, U.; Steinmeyer, A.; Larrea, F. Regulation of vitamin D hydroxylases gene expression by 1,25-dihydroxyvitamin d3 and cyclic amp in cultured human syncytiotrophoblasts. J. Steroid. Biochem. Mol. Biol. 2007, 103, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.N.; Nguyen, L.; Chan, J.; Innes, B.A.; Bulmer, J.N.; Kilby, M.D.; Hewison, M. Effects of 25-hydroxyvitamin d3 and 1,25-dihydroxyvitamin d3 on cytokine production by human decidual cells. Biol. Reprod. 2006, 75, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Murthi, P.; Yong, H.E.; Ngyuen, T.P.; Ellery, S.; Singh, H.; Rahman, R.; Dickinson, H.; Walker, D.W.; Davies-Tuck, M.; Wallace, E.M.; et al. Role of the placental vitamin d receptor in modulating feto-placental growth in fetal growth restriction and preeclampsia-affected pregnancies. Front. Physiol. 2016, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Jelliffe-Pawlowski, L.L.; Rand, L.; Bedell, B.; Baer, R.J.; Oltman, S.P.; Norton, M.E.; Shaw, G.M.; Stevenson, D.K.; Murray, J.C.; Ryckman, K.K. Correction: Prediction of preterm birth with and without preeclampsia using mid-pregnancy immune and growth-related molecular factors and maternal characteristics. J. Perinatol. 2018, 38, 946. [Google Scholar] [CrossRef] [PubMed]
- Karras, S.; Paschou, S.A.; Kandaraki, E.; Anagnostis, P.; Annweiler, C.; Tarlatzis, B.C.; Hollis, B.W.; Grant, W.B.; Goulis, D.G. Hypovitaminosis D in pregnancy in the Mediterranean region: A systematic review. Eur. J. Clin. Nutr. 2016, 70, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Zhang, F.; Richards, J.B.; Kestenbaum, B.; van Meurs, J.B.; Berry, D.; Kiel, D.P.; Streeten, E.A.; Ohlsson, C.; Koller, D.L.; et al. Common genetic determinants of vitamin d insufficiency: A genome-wide association study. Lancet 2010, 376, 180–188. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Study Population (n = 187) | PTB (n = 17) | Full-Term (n = 170) | p-Value |
|---|---|---|---|---|
| Age, years a | 37.0 (4) | 37.0 (5) | 38.0 (4) | 0.648 |
| Gestational age at enrolment, weeks | 16.0 (4) | 16.0 (4) | 16.0 (4) | 0.691 |
| Educational level (% low-medium) | 13.9% | 5.9% | 14.7% | 0.316 |
| Employment status (% employed) | 55.6% | 47.1% | 56.5% | 0.456 |
| Smoking (% current smokers) | 18.3% | 5.8% | 19.5% | 0.165 |
| Pre-gestational nutritional status | ||||
| Underweight | 8.1% | 11.8% | 7.7% | 0.522 |
| Normal weight | 65.6% | 58.8% | 66.3% | |
| Overweight | 17.2% | 11.8% | 17.8% | |
| Obese | 9.1% | 17.6% | 8.3% | |
| GWG, kg a | 12.0 (7) | 11.0 (10) | 12.0 (6.9) | 0.630 |
| GWG classification | ||||
| Reduced | 30.9% | 35.3% | 30.5% | 0.903 |
| Adequate | 42% | 41.2% | 42.1% | |
| Excessive | 27.1% | 23.5% | 27.4% | |
| Vitamin D intake, µg/day a | 3.7 (3.5) | 3.7 (4.3) | 3.1 (3.6) | 0.808 |
| Vitamin D supplements (% users) | 10.7% | 5.9% | 11.2% | 0.501 |
| Gestational duration, weeks | 39.0 (2) | 35.5 (2) | 39.0 (2) | <0.001 |
| Sex (% male) | 50.3% | 47.1% | 50.6% | 0.781 |
| Birth weight, kg a | 3.2 (0.6) | 2.44 0.5) | 3.2 (0.6) | <0.001 |
| Birth length, cm a | 50.0 (2) | 47.5 (4) | 50.0 (2) | <0.001 |
| Type of delivery | ||||
| Natural | 55.1% | 47.1% | 55.9% | 0.486 |
| Caesarean section | 44.9% | 52.9% | 44.1% | |
| Underweight (%) | 8% | 58.8% | 2.9% | <0.001 |
| Macrosomia (%) | 7.5% | 0% | 8.2% | 0.219 |
| Weight for gestational age | ||||
| SGA | 8% | 5.9% | 8.2% | 0.283 |
| AGA | 80.2% | 70.6% | 81.2% | |
| LGA | 11.8% | 23.5% | 10.6% |
| Authors | Country | Study Design | Ethnicity | Sample Size | Sample | SNPs | Genotyping Method |
|---|---|---|---|---|---|---|---|
| Baczyńska-Strzecha et al., 2016 [9] | Poland | Case-control | Caucasian | 199 | Maternal blood | ApaI BsmI TaqI | TaqMan Assay |
| Javorski et al., 2018 [33] | Brazil | Case-control | Mixed | 189 | Maternal blood | FokI | TaqMan Assay |
| Rosenfeld et al., 2017 [34] | Israel | Case-control | Mixed | 375 | Maternal and fetal blood | ApaI BsmI FokI TaqI | RFLP |
| Mamma & Bambino Cohort, 2018 | Italy | Prospective cohort | Caucasian | 187 | Maternal blood | ApaI BsmI FokI TaqI | TaqMan Assay |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barchitta, M.; Maugeri, A.; La Rosa, M.C.; Magnano San Lio, R.; Favara, G.; Panella, M.; Cianci, A.; Agodi, A. Single Nucleotide Polymorphisms in Vitamin D Receptor Gene Affect Birth Weight and the Risk of Preterm Birth: Results From the “Mamma & Bambino” Cohort and A Meta-Analysis. Nutrients 2018, 10, 1172. https://doi.org/10.3390/nu10091172
Barchitta M, Maugeri A, La Rosa MC, Magnano San Lio R, Favara G, Panella M, Cianci A, Agodi A. Single Nucleotide Polymorphisms in Vitamin D Receptor Gene Affect Birth Weight and the Risk of Preterm Birth: Results From the “Mamma & Bambino” Cohort and A Meta-Analysis. Nutrients. 2018; 10(9):1172. https://doi.org/10.3390/nu10091172
Chicago/Turabian StyleBarchitta, Martina, Andrea Maugeri, Maria Clara La Rosa, Roberta Magnano San Lio, Giuliana Favara, Marco Panella, Antonio Cianci, and Antonella Agodi. 2018. "Single Nucleotide Polymorphisms in Vitamin D Receptor Gene Affect Birth Weight and the Risk of Preterm Birth: Results From the “Mamma & Bambino” Cohort and A Meta-Analysis" Nutrients 10, no. 9: 1172. https://doi.org/10.3390/nu10091172
APA StyleBarchitta, M., Maugeri, A., La Rosa, M. C., Magnano San Lio, R., Favara, G., Panella, M., Cianci, A., & Agodi, A. (2018). Single Nucleotide Polymorphisms in Vitamin D Receptor Gene Affect Birth Weight and the Risk of Preterm Birth: Results From the “Mamma & Bambino” Cohort and A Meta-Analysis. Nutrients, 10(9), 1172. https://doi.org/10.3390/nu10091172