TGF-β2, EGF, and FGF21 Growth Factors Present in Breast Milk Promote Mesenteric Lymph Node Lymphocytes Maturation in Suckling Rats

 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Dietary Supplementation
2.3. Measurement of Growth and Development
2.4. Sample Collection and Processing
2.5. Quantification of Intestinal IgA and IgM by ELISA
2.6. Lymphocyte Isolation from Mesenteric Lymph Nodes
2.7. Mesenteric Lymph Node Cells Stimulation and Proliferation Assay
2.8. Quantification of Cytokine Secretion by Mesenteric Lymph Node Lymphocytes
2.9. Immunofluorescence Staining and Flow Cytometry Analysis
2.10. Statistical Analysis
3. Results
3.1. Animal Growth
3.2. IgA and IgM Concentration in Gut Wash
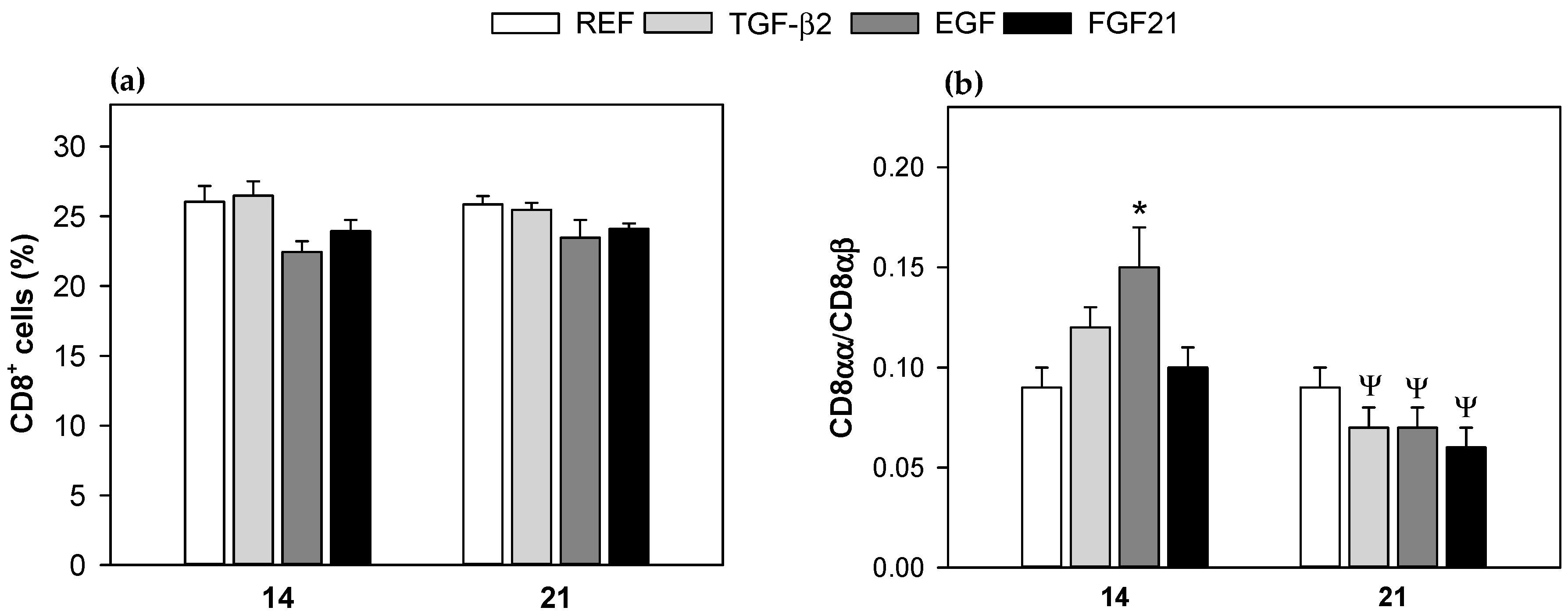
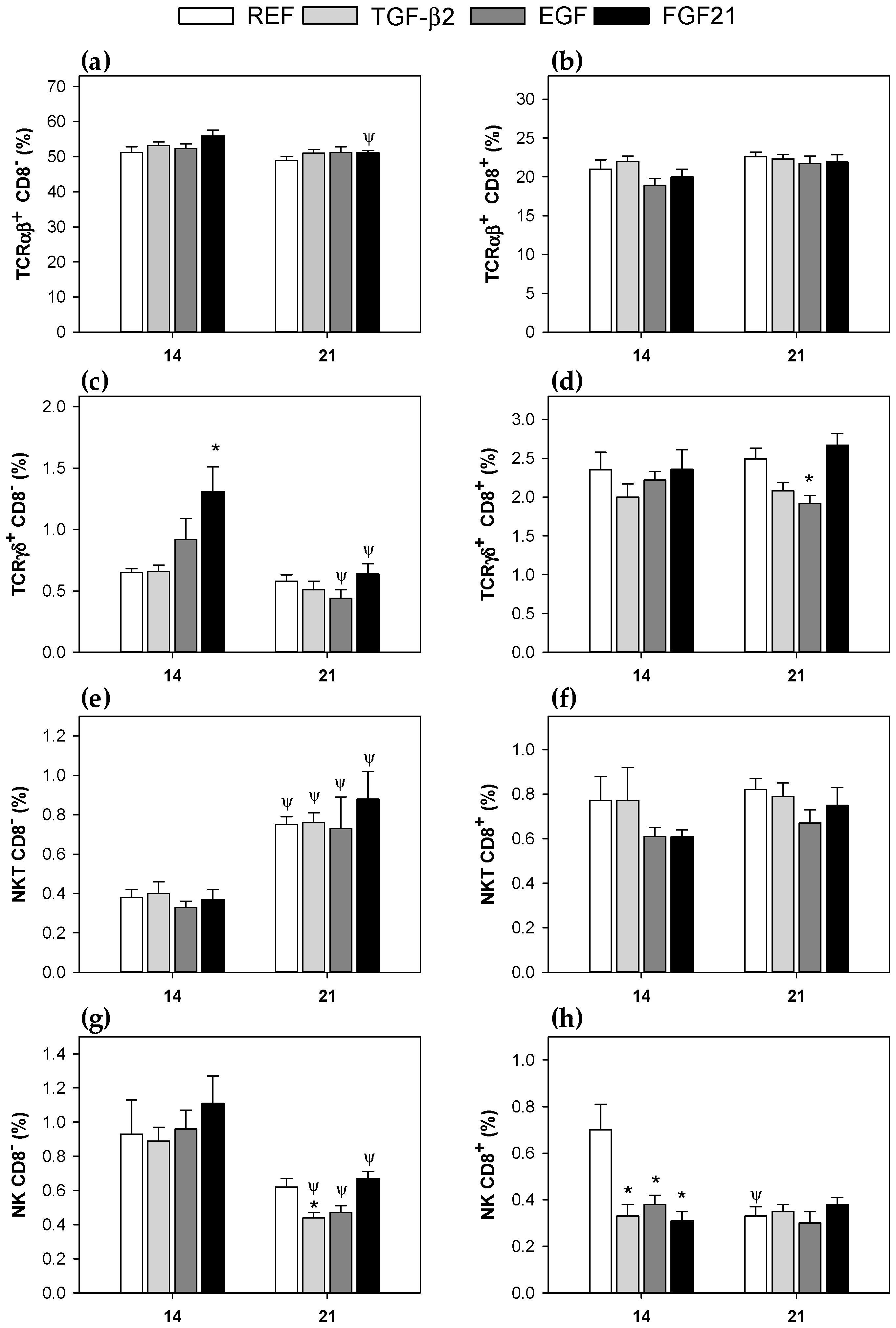
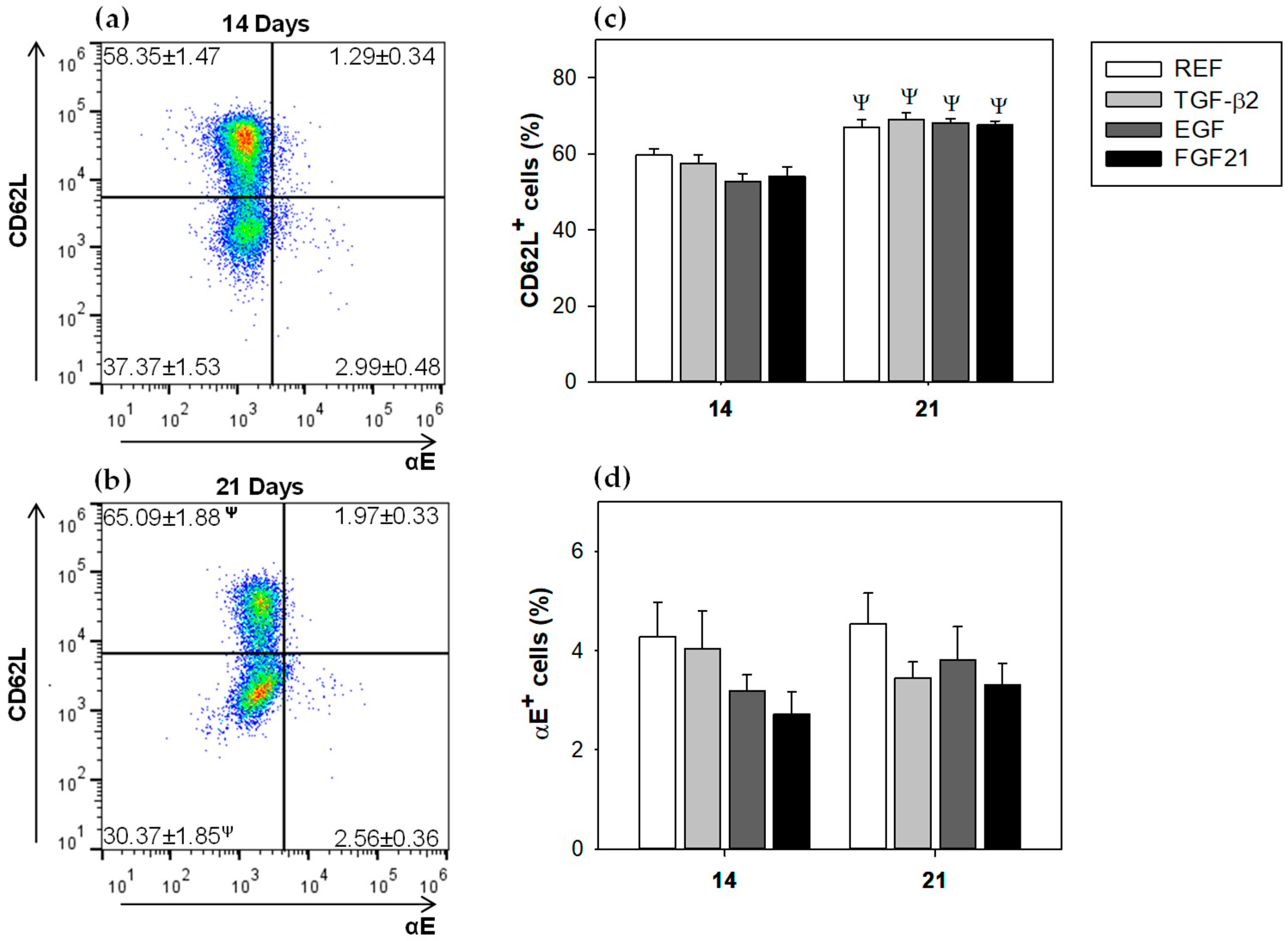
3.3. Mesenteric Lymph Node Lymphocytes Composition
3.4. Proliferation and Cytokine Production by Mesenteric Lymph Nodes Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Turfkruyer, M.; Verhasselt, V. Breast milk and its impact on maturation of the neonatal immune system. Curr. Opin. Infect. Dis. 2015, 28, 199–206. [Google Scholar] [CrossRef] [PubMed]
- García, C.; Duan, R.D.; Brévaut-Malaty, V.; Gire, C.; Millet, V.; Simeoni, U.; Bernard, M.; Armand, M. Bioactive compounds in human milk and intestinal health and maturity in preterm newborn: An overview. Cell. Mol. Biol. 2013, 59, 108–131. [Google Scholar] [CrossRef] [PubMed]
- Penttila, I.A.; van Spriel, A.B.; Zhang, M.F.; Xian, C.J.; Steeb, C.B.; Cummins, A.G.; Zola, H.; Read, L.C. Transforming growth factor-β levels in maternal milk and expression in postnatal rat duodenum and ileum. Pediatr. Res. 1998, 44, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Buddington, R.K.; Sangild, P.T. Companion animals symposium: Development of the mammalian gastrointestinal tract, the resident microbiota, and the role of diet in early life. J. Anim. Sci. 2011, 89, 1506–1519. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, S.K.; Odle, J. Nutritional factors influencing intestinal health of the neonate. Adv. Nutr. 2012, 3, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Castellote, C.; Casillas, R.; Ramírez-Santana, C.; Pérez-Cano, F.J.; Castell, M.; Moretones, M.G.; López-Sabater, M.C.; Franch, À. Premature delivery influences the immunological composition of colostrum and transitional and mature human milk. J. Nutr. 2011, 141, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Field, C.J. The immunological components of human milk and their effect on immune development in infants. J. Nutr. 2005, 135, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Andreas, N.J.; Kampmann, B.; Mehring Le-Doare, K. Human breast milk: A review on its composition and bioactivity. Early Hum. Dev. 2015, 91, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Penttila, I.A.; Flesch, I.E.; McCue, A.L.; Powell, B.C.; Zhou, F.H.; Read, L.C.; Zola, H. Maternal milk regulation of cell infiltration and interleukin 18 in the intestine of suckling rat pups. Gut 2003, 52, 1579–1586. [Google Scholar] [CrossRef] [PubMed]
- Sanjabi, S.; Oh, S.A.; Li, M.O. Regulation of the immune response by TGF-β: From conception to autoimmunity and infection. Cold Spring Harb. Perspect. Biol. 2017, 9, a022236. [Google Scholar] [CrossRef] [PubMed]
- Penttila, I.A. Milk-derived transforming growth factor-beta and the infant immune response. J. Pediatr. 2010, 156, S21–S25. [Google Scholar] [CrossRef] [PubMed]
- Penttila, I. Effects of transforming growth factor-beta and formula feeding on systemic immune responses to dietary β-lactoglobulin in allergy-prone rats. Pediatr. Res. 2006, 59, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Liu, H.; Yang, S.; Li, Z.; Zhong, J.; Fang, R. Epidermal growth factor and intestinal barrier function. Mediat. Inflamm. 2016, 2016, 1927348. [Google Scholar] [CrossRef] [PubMed]
- Berseth, C.L. Enhancement of intestinal growth in neonatal rats by epidermal growth factor in milk. Am. J. Physiol. 1987, 253, G662–G665. [Google Scholar] [CrossRef] [PubMed]
- Gavaldà-Navarro, A.; Hondares, E.; Giralt, M.; Mampel, T.; Iglesias, R.; Villarroya, F. Fibroblast growth factor 21 in breast milk controls neonatal intestine function. Sci. Rep. 2015, 5, 13717. [Google Scholar] [CrossRef] [PubMed]
- Coursodon, C.F.; Dvorak, B. Epidermal growth factor and necrotizing enterocolitis. Curr. Opin. Pediatr. 2012, 24, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Kharitonenkov, A.; DiMarchi, R. Fibroblast growth factor 21 night watch: Advances and uncertainties in the field. J. Intern. Med. 2017, 281, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Grases-Pintó, B.; Abril-Gil, M.; Rodríguez-Lagunas, M.J.; Castell, M.; Pérez-Cano, F.J.; Franch, À. Leptin and adiponectin supplementation modifies mesenteric lymph node lymphocyte composition and functionality in suckling rats. Br. J. Nutr. 2018, 119, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Rigo-Adrover, M.D.M.; Van Limpt, K.; Knipping, K.; Garssen, J.; Knol, J.; Costabile, A.; Franch, À.; Castell, M.; Pérez-Cano, F.J. Preventive effect of a synbiotic combination of galacto- and fructooligosaccharides mixture with Bifidobacterium breve M-16V in a model of multiple rotavirus infections. Front. Immunol. 2018, 9, 1318. [Google Scholar] [CrossRef] [PubMed]
- Maynard, A.A.; Dvorak, K.; Khailova, L.; Dobrenen, H.; Arganbright, K.M.; Halpern, M.D.; Kurundkar, A.R.; Maheshwari, A.; Dvorak, B. Epidermal growth factor reduces autophagy in intestinal epithelium and in the rat model of necrotizing enterocolitis. Am. J. Physiol. Liver Physiol. 2010, 299, G614–G622. [Google Scholar] [CrossRef] [PubMed]
- Camps-Bossacoma, M.; Pérez-Cano, F.J.; Franch, À.; Untersmayr, E.; Castell, M. Effect of a cocoa diet on the small intestine and gut-associated lymphoid tissue composition in an oral sensitization model in rats. J. Nutr. Biochem. 2017, 42, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Basha, S.; Surendran, N.; Pichichero, M. Immune responses in neonates. Expert Rev. Clin. Immunol. 2014, 10, 1171–1184. [Google Scholar] [CrossRef] [PubMed]
- Hormi, K.; Lehy, T. Transforming growth factor-alpha in vivo stimulates epithelial cell proliferation in digestive tissues of suckling rats. Gut 1996, 39, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Pollack, P.F.; Goda, T.; Colony, P.C.; Edmond, J.; Thornburg, W.; Korc, M.; Koldovský, O. Effects of enterally fed epidermal growth factor on the small and large intestine of the suckling rat. Regul. Pept. 1987, 17, 121–132. [Google Scholar] [CrossRef]
- Camporez, J.P.; Asrih, M.; Zhang, D.; Kahn, M.; Samuel, V.T.; Jurczak, M.J.; Jornayvaz, F.R. Hepatic insulin resistance and increased hepatic glucose production in mice lacking FGF21. J. Endocrinol. 2015, 226, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cano, F.J.; Franch, À.; Castellote, C.; Castell, M. The suckling rat as a model for immunonutrition studies in early life. Clin. Dev. Immunol. 2012, 2012, 537310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.F. The Role of Milk Transforming Growth Factor-β (TGF-β) in the Development of the Infant Gut and Gut Mucosal Immune System. Ph.D. Thesis, University of Adelaide, Adelaide, Australia, 2000. [Google Scholar]
- Wills-Karp, M.; Luyimbazi, J.; Xu, X.; Schofield, B.; Neben, T.Y.; Karp, C.L.; Donaldson, D.D. Interleukin-13: Central mediator of allergic asthma. Science 1998, 282, 2258–2261. [Google Scholar] [CrossRef] [PubMed]
- Kubo, T.; Morita, H.; Sugita, K.; Akdis, C.A. Introduction to mechanisms of allergic diseases. In Middleton’s Allergy Essentials; Holgate, S., Sheikh, A., Eds.; Elsevier: New York, NY, USA, 2017; pp. 1–27. ISBN 978-0-323-37579-5. [Google Scholar]
- Bao, K.; Reinhardt, R.L. The differential expression of IL-4 and IL-13 and its impact on type-2 immunity. Cytokine 2015, 75, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Munblit, D.; Treneva, M.; Peroni, D.G.; Colicino, S.; Chow, L.Y.; Dissanayeke, S.; Pampura, A.; Boner, A.L.; Geddes, D.T.; Boyle, R.J.; et al. Immune components in human milk are associated with early infant immunological health outcomes: A prospective three-country analysis. Nutrients 2017, 9, 532. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, K.; Koyanagi, A.; Kamachi, F.; Harauma, A.; Chiba, A.; Hisata, K.; Moriguchi, T.; Shimizu, T.; Miyake, S. Breast-feeding regulates immune system development via transforming growth factor-β in mice pups. Pediatr. Int. 2018, 60, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, B.; Halpern, M.D.; Holubec, H.; Williams, C.S.; McWilliam, D.L.; Dominguez, J.A.; Stepankova, R.; Payne, C.M.; McCuskey, R.S. Epidermal growth factor reduces the development of necrotizing enterocolitis in a neonatal rat model. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G156–G164. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.T.; Walker, W.A. Epidermal growth factor and the developing human gut. Gastroenterology 1988, 94, 845–847. [Google Scholar] [CrossRef]
- Wong, W.M.; Wright, N.A. Epidermal growth factor, epidermal growth factor receptors, intestinal growth, and adaptation. JPEN J. Parenter. Enteral Nutr. 1999, 23, S83–S88. [Google Scholar] [CrossRef] [PubMed]
- Halpern, M.D.; Dominguez, J.A.; Dvorakova, K.; Holubec, H.; Williams, C.S.; Meza, Y.G.; Ruth, M.C.; Dvorak, B. Ileal cytokine dysregulation in experimental necrotizing enterocolitis is reduced by epidermal growth factor. J. Pediatr. Gastroenterol. Nutr. 2003, 36, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Hanson, L.Å.; Korotkova, M. The role of breastfeeding in prevention of neonatal infection. Semin. Neonatol. 2002, 7, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Takeyama, K.; Dabbagh, K.; Lee, H.M.; Agustí, C.; Lausier, J.A.; Ueki, I.F.; Grattan, K.M.; Nadel, J.A. Epidermal growth factor system regulates mucin production in airways. Proc. Natl. Acad. Sci. USA 1999, 96, 3081–3086. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, J.; Huang, H.; Huang, Y.; Mi, T.; Zhang, C.; Zhang, L. Fibroblast growth factor 21 delayed endothelial replicative senescence and protected cells from H2O2-induced premature senescence through SIRT1. Am. J. Transl. Res. 2017, 9, 4492–4501. [Google Scholar] [PubMed]
- Li, S.M.; Yu, Y.H.; Li, L.; Wang, W.F.; Li, D.S. Treatment of CIA mice with FGF21 down-regulates TH17-IL-17 axis. Inflammation 2016, 39, 309–319. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| BMI | Lee Index | SI Relative Weight | SI Relative Length | ||
|---|---|---|---|---|---|
| (g/cm2) | ((∛g/cm) × 1000) | (%) | (%) | ||
| Day 14 | REF | 0.365 ± 0.009 | 332.9 ± 3.473 | 3.51 ± 0.05 | 118.29 ± 5.49 |
| TGF-β2 | 0.373 ± 0.010 | 336.9 ± 3.844 | 3.59 ± 0.14 | 114.34 ± 4.28 | |
| EGF | 0.340 ± 0.007 | 327.0 ± 2.845 | 3.48 ± 0.06 | 122.64 ± 2.91 | |
| FGF21 | 0.371 ± 0.009 | 337.4 ± 4.044 | 3.18 ± 0.06 | 105.59 ± 5.31 | |
| Day 21 | REF | 0.404 ± 0.010 Ψ | 319.5 ± 2.551 Ψ | 4.15 ± 0.24 Ψ | 89.57 ± 3.12 Ψ |
| TGF-β2 | 0.404 ± 0.009 Ψ | 320.8 ± 3.225 Ψ | 4.25 ± 0.09 Ψ | 85.65 ± 2.20 Ψ | |
| EGF | 0.404 ± 0.007 Ψ | 322.9 ± 2.574 | 4.38 ± 0.07 Ψ | 88.34 ± 2.20 Ψ | |
| FGF21 | 0.420 ± 0.011 Ψ | 329.0 ± 4.053 | 4.78 ± 0.17 Ψ | 89.97 ± 4.01 Ψ |
| Reference | TGF-β2 | EGF | FGF21 | ||
|---|---|---|---|---|---|
| Day 14 | T cells (%) | 75.90 ± 1.80 | 77.90 ± 1.39 | 74.36 ± 1.88 | 79.60 ± 1.50 |
| T TCRαβ+ (%) | 72.90 ± 1.98 | 75.24 ± 1.29 | 71.22 ± 1.94 | 75.90 ± 1.20 | |
| T TCRγδ+ (%) | 3.00 ± 0.20 | 2.66 ± 0.20 | 3.14 ± 0.23 | 3.70 ± 0.40 | |
| NK (%) | 1.63 ± 0.30 | 1.22 ± 0.08 | 1.35 ± 0.12 | 1.42 ± 0.17 | |
| NKT (%) | 1.15 ± 0.14 | 1.18 ± 0.20 | 0.94 ± 0.05 | 0.98 ± 0.06 | |
| B cells (%) | 17.65 ± 1.65 | 16.19 ± 1.40 | 19.62 ± 1.23 | 15.86 ± 1.07 | |
| Day 21 | T cells (%) | 74.60 ± 1.36 | 75.86 ± 1.10 | 75.29 ± 1.20 | 76.41 ± 0.69 |
| T TCRαβ+ (%) | 71.57 ± 1.44 | 73.28 ± 1.14 | 72.93 ± 1.26 | 73.10 ± 0.77 | |
| T TCRγδ+ (%) | 3.07 ± 0.15 | 2.58 ± 0.15 | 2.36 ± 0.13 *,Ψ | 3.31 ± 0.13 | |
| NK (%) | 0.95 ± 0.07 Ψ | 0.79 ± 0.06 Ψ | 0.77 ± 0.08 Ψ | 1.05 ± 0.07 | |
| NKT (%) | 1.57 ± 0.07 Ψ | 1.55 ± 0.11 | 1.39 ± 0.21 | 1.63 ± 0.18 Ψ | |
| B cells (%) | 14.67 ± 0.71 | 14.30 ± 0.64 | 12.95 ± 1.19 Ψ | 13.23 ± 0.62 |
| 14 Days | |||||
| Reference | TGF-β2 | EGF | FGF21 | ||
| %αE | CD8+ | 2.75 ± 0.72 | 2.92 ± 0.49 | 2.94 ± 0.52 | 2.88 ± 0.44 |
| CD4+ | 2.81 ± 0.39 | 3.07 ± 0.45 | 4.46 ± 0.34 | 3.80 ± 0.26 | |
| B cells | 15.98 ± 1.61 | 18.33 ± 1.39 | 18.92 ± 2.50 | 20.32 ± 3.12 | |
| %CD62L | CD8+ | 65.27 ± 1.56 | 62.38 ± 3.47 | 59.25 ± 2.52 | 59.21 ± 3.55 |
| CD4+ | 59.55 ± 1.76 | 55.41 ± 2.36 | 53.61 ± 1.37 | 53.65 ± 2.40 | |
| B cells | 48.43 ± 2.77 | 46.6 ± 2.05 | 38.77 ± 2.08 | 37.29 ± 4.78 | |
| 21 Days | |||||
| Reference | TGF-β2 | EGF | FGF21 | ||
| %αE | CD8+ | 3.84 ± 0.48 | 2.98 ± 1.04 | 3.70 ± 0.48 | 3.97 ± 0.66 |
| CD4+ | 2.83 ± 0.75 | 2.61 ± 0.52 | 1.94 ± 0.18 Ψ | 2.52 ± 0.30 Ψ | |
| B cells | 18.43 ± 1.83 Ψ | 13.03 ± 1.55 Ψ | 14.95 ± 1.69 Ψ | 15.75 ± 1.16 Ψ | |
| %CD62L | CD8+ | 71.49 ± 1.38 Ψ | 72.10 ± 1.93 Ψ | 70.78 ± 2.19 Ψ | 72.88 ± 2.37 Ψ |
| CD4+ | 66.02 ± 1.74 Ψ | 68.24 ± 1.64 Ψ | 66.66 ± 0.42 Ψ | 66.80 ± 1.22 Ψ | |
| B cells | 56.79 ± 2.25 | 60.92 ± 2.17 Ψ | 60.82 ± 3.92 | 57.61 ± 2.37 | |
| 14 Days | ||||
| (pg/mL) | Reference | TGF-β2 | EGF | FGF21 |
| IL-2 | 42.71 ± 5.96 | 39.24 ± 5.66 | 50.45 ± 6.66 | 56.72 ± 11.04 |
| IL-4 | 89.82 ± 13.00 | 57.14 ± 7.81 | 82.65 ± 12.47 | 74.97 ± 12.18 |
| IL-10 | 879.82 ± 239.40 | 688.01 ± 145.14 | 1450.87 ± 296.29 | 1326.99 ± 217.06 |
| IL-13 | 25.38 ± 5.64 | 17.53 ± 3.03 | 31.91 ± 4.28 | 26.18 ± 4.03 |
| IL-17A | 71.98 ± 26.92 | 70.22 ± 20.16 | 60.12 ± 13.61 | 50.82 ± 9.53 |
| IFN-γ | 10323.26 ± 2391.46 | 8302.75 ± 1964.09 | 9329.17 ± 1662.87 | 7882.51 ± 2810.34 |
| TNF-α | 8.76 ± 0.98 | 9.48 ± 0.68 | 9.19 ± 1.16 | 7.40 ± 0.25 |
| IL-10/TNF-α | 115.00 ± 33.11 | 79.71 ± 21.80 | 170.87 ± 43.10 | 178.05 ± 29.25 |
| IFN-γ/IL-4 | 110.35 ± 20.65 | 195.32 ± 61.39 | 124.85 ± 37.55 | 103.80 ± 26.93 |
| 21 Days | ||||
| (pg/mL) | Reference | TGF-β2 | EGF | FGF21 |
| IL-2 | 390.57 ± 141.49 Ψ | 383.80 ± 89.21 Ψ | 244.79 ± 80.37 Ψ | 270.16 ± 114.54 |
| IL-4 | 70.52 ± 10.51 | 52.21 ± 7.69 | 48.52 ± 1.91 Ψ | 66.60 ± 8.80 |
| IL-10 | 705.45 ± 63.95 | 514.70 ± 55.30 | 508.26 ± 61.43 Ψ | 542.36 ± 78.82 Ψ |
| IL-13 | 19.51 ± 2.44 | 12.92 ± 0.74 * | 9.91 ± 1.24 *,Ψ | 9.83 ± 1.65 *,Ψ |
| IL-17A | 59.97 ± 7.14 | 46.91 ± 4.03 | 52.30 ± 10.7022 | 41.24 ± 11.77 |
| IFN-γ | 4231.26 ± 365.41 Ψ | 4488.50 ± 416.14 | 6900.74 ± 1129.78 | 4595.11 ± 1058.87 |
| TNF-α | 6.63 ± 0.11 Ψ | 6.55 ± 0.20 Ψ | 10.02 ± 0.40 * | 5.91 ± 0.38 Ψ |
| IL-10/TNF-α | 106.47 ± 9.73 | 80.13 ± 9.55 | 50.48 ± 5.72 *,Ψ | 95.64 ± 17.03 Ψ |
| IFN-γ/IL-4 | 64.25 ± 9.61 | 111.99 ± 25.23 | 147.25 ± 27.9166 | 81.50 ± 25.37 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Castro, P.; Abril-Gil, M.; Rodríguez-Lagunas, M.J.; Castell, M.; Pérez-Cano, F.J.; Franch, À. TGF-β2, EGF, and FGF21 Growth Factors Present in Breast Milk Promote Mesenteric Lymph Node Lymphocytes Maturation in Suckling Rats. Nutrients 2018, 10, 1171. https://doi.org/10.3390/nu10091171
Torres-Castro P, Abril-Gil M, Rodríguez-Lagunas MJ, Castell M, Pérez-Cano FJ, Franch À. TGF-β2, EGF, and FGF21 Growth Factors Present in Breast Milk Promote Mesenteric Lymph Node Lymphocytes Maturation in Suckling Rats. Nutrients. 2018; 10(9):1171. https://doi.org/10.3390/nu10091171
Chicago/Turabian StyleTorres-Castro, Paulina, Mar Abril-Gil, María J. Rodríguez-Lagunas, Margarida Castell, Francisco J. Pérez-Cano, and Àngels Franch. 2018. "TGF-β2, EGF, and FGF21 Growth Factors Present in Breast Milk Promote Mesenteric Lymph Node Lymphocytes Maturation in Suckling Rats" Nutrients 10, no. 9: 1171. https://doi.org/10.3390/nu10091171
APA StyleTorres-Castro, P., Abril-Gil, M., Rodríguez-Lagunas, M. J., Castell, M., Pérez-Cano, F. J., & Franch, À. (2018). TGF-β2, EGF, and FGF21 Growth Factors Present in Breast Milk Promote Mesenteric Lymph Node Lymphocytes Maturation in Suckling Rats. Nutrients, 10(9), 1171. https://doi.org/10.3390/nu10091171