
Towards a Synthetic Biology Toolset for Metallocluster Enzymes in Biosynthetic Pathways: What We Know and What We Need


1
INSERM U722, Faculté de Médecine, Université de Paris, Site Xavier Bichat, 75018 Paris, France
2
Department of Bionanoscience, Kavli Institute of Nanoscience, Delft University of Technology, 2629 HZ Delft, The Netherlands
*
Authors to whom correspondence should be addressed.
Molecules 2021, 26(22), 6930; https://doi.org/10.3390/molecules26226930
Submission received: 6 October 2021
/
Revised: 14 November 2021
/
Accepted: 15 November 2021
/
Published: 17 November 2021
(This article belongs to the Special Issue Molecular Metal Clusters: Fundamental and Applied Aspects)
Abstract
:Microbes are routinely engineered to synthesize high-value chemicals from renewable materials through synthetic biology and metabolic engineering. Microbial biosynthesis often relies on expression of heterologous biosynthetic pathways, i.e., enzymes transplanted from foreign organisms. Metallocluster enzymes are one of the most ubiquitous family of enzymes involved in natural product biosynthesis and are of great biotechnological importance. However, the functional expression of recombinant metallocluster enzymes in live cells is often challenging and represents a major bottleneck. The activity of metallocluster enzymes requires essential supporting pathways, involved in protein maturation, electron supply, and/or enzyme stability. Proper function of these supporting pathways involves specific protein–protein interactions that remain poorly characterized and are often overlooked by traditional synthetic biology approaches. Consequently, engineering approaches that focus on enzymatic expression and carbon flux alone often overlook the particular needs of metallocluster enzymes. This review highlights the biotechnological relevance of metallocluster enzymes and discusses novel synthetic biology strategies to advance their industrial application, with a particular focus on iron-sulfur cluster enzymes. Strategies to enable functional heterologous expression and enhance recombinant metallocluster enzyme activity in industrial hosts include: (1) optimizing specific maturation pathways; (2) improving catalytic stability; and (3) enhancing electron transfer. In addition, we suggest future directions for developing microbial cell factories that rely on metallocluster enzyme catalysis.
1. Introduction
In the two decades since its conception, the field of synthetic biology has moved genetic engineering considerably closer to an actual engineering discipline. The combination of synthetic biology tools with the principles of metabolic engineering have tremendously advanced our ability to engineer microorganisms to produce valuable products [1]. This progress is evident in the gradual shift of production of select commodities and fine chemicals to favor bio-based processes [2,3]. Central to this approach is the use of heterologous hosts as biosynthetic platforms, such as Escherichia coli [4], Streptomycetes [5] or yeast [6]. A classic synthetic biology approach to improve microbial biosynthesis commonly involves cloning codon-optimized heterologous genes into the platform host, followed by engineering-controlled expression/balancing of pathway enzymes, and optimizing supplies of pathway precursors and cofactors [7]. However, while synthetic biology has had some success in making genes universally compatible with heterologous hosts, far fewer tools have been developed that can adapt enzymes to foreign hosts. If enzymes lose catalytic activity when expressed in heterologous hosts, the gene-focused tools of synthetic biology cannot engineer microbial production.
Metallocluster enzymes, one of the most ubiquitous and versatile families of metalloproteins, employ metal-containing cofactors to catalyze some of the most remarkable reactions in nature [8,9]. These enzymes are of great biotechnological relevance as they are involved in the production of valuable natural products—such as pharmaceuticals or biofuels—or in metabolic pathways that utilize alternative carbon sources. However, their application in heterologous biosynthesis is limited, as metallocluster-containing enzymes often display little-to-no activity when expressed in heterologous organisms, leading to low product yields. Consequently, they are a significant bottleneck in the microbial production of industrially important chemicals. This is notably the case for numerous enzymes that contain iron-sulfur (FeS) clusters, which are required for the biosynthesis of a wide variety of useful chemicals. Unfortunately, these enzymes often display limited activity in foreign hosts, hence precluding the development of many relevant microbial cell factories.
The poor activity of metallocluster-containing enzymes in heterologous organisms is often considered to stem from their dependence on poorly characterized essential supporting pathways for their maturation and catalytic cycle. Therefore, traditional metabolic engineering approaches may be blind to the challenges in reconstituting and optimizing metabolic pathways that rely on these enzymes. In this review, we highlight the biotechnological relevance of metallocluster-containing enzymes and the challenges to their application in microbial biosynthesis. Finally, we detail successful engineering strategies that enable the functional expression of metallocluster enzymes to improve recombinant pathway yields for microbial cell factories, with an emphasis on FeS cluster enzymes.
2. Biotechnological Relevance of Metallocluster Enzymes: Potential and Challenges
2.1. Many Industrially Important Pathways Rely on Metallocluster Enzyme Activity
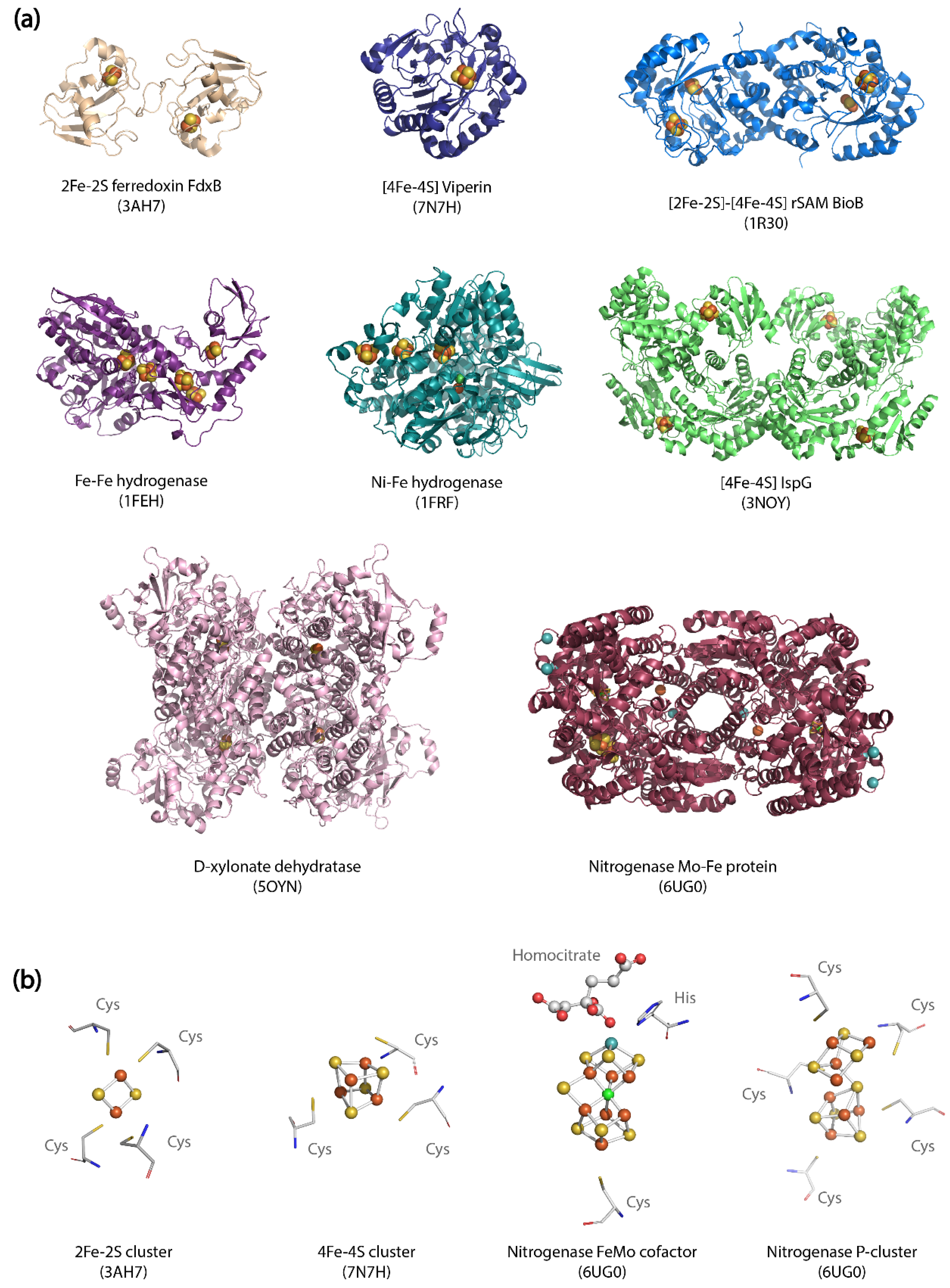
Enzymes that contain metallocofactors composed of homo- or hetero-nuclear clusters (i.e., [Fe-Fe], [Fe-Ni], [Fe-S] and [Fe-Mo] clusters) [8] are involved in a wide array of metabolic pathways. Metalloclusters participate in reactions either directly by providing catalytic sites, or by acting as electron transport centers. The absence or incomplete maturation of metalloclusters completely eliminates enzymatic activity. The natural ability of metalloclusters to bind a wide range of protein structures also accounts for their biosynthetic versatility (Figure 1). Because metallocluster-containing enzymes facilitate a huge variety of reactions that include oxidative transformations, epimerization, hydroxylation, epoxidation, methyl transfers, alkylation or oxygenation, they hold a tremendous potential for microbial biosynthesis [9,10]. Table 1 highlights metabolic pathways of prominent biotechnological relevance that rely upon reactions catalyzed by metallocluster enzymes.
2.2. Challenges Related to Recombinant Pathways That Contain Metallocluster Enzymes
The functional expression of recombinant metallocluster enzymes is often challenging. The abundance of functional metalloenzyme relies on the efficient synthesis and incorporation of metalloclusters into inactive apoenzymes. This maturation process usually involves complex post-translational modifications coordinated by specialized proteins. The proper maturation and incorporation of every metallocluster is essential for catalysis. Furthermore, the catalytic activity of many metalloenzymes requires specific electron transfer proteins.
The complexity of the essential supporting systems that drive metallocluster enzymes is often overlooked by traditional synthetic biology and metabolic engineering strategies. Therefore, the functional expression of metallocluster enzymes must overcome several challenges, including: (i) lack of activity in heterologous hosts; (ii) absent/ineffective maturation pathways; (iii) poorly characterized cofactor requirements or absent/ineffective electron transfer pathways; and (iv) oxygen sensitivity. Traditional strain engineers are often unfamiliar with these complications, which frustrates the development of microbial cell factories that rely on metalloenzyme catalysis.
2.2.1. The Functional Expression of Metallocluster Enzymes Requires Specific Maturation Pathways
Specific post-translational modifications mediated by specialized proteins [27] are essential for the maturation of metallocluster enzymes. For instance, the functional expression of most FeS cluster enzymes [28], nitrogenases [29], or [NiFe]- and [FeFe]- hydrogenases [25] involve several helper proteins that assemble and deliver the clusters to the apoenzyme via sequential ligand exchange reactions. Heterologous hosts that lack these essential pathways will be unable to functionally express such foreign metallocluster enzymes [30]. Moreover, even when compatible maturation pathways are present in the host, they may not be optimal for a given heterologous enzyme. At least three different FeS cluster maturation systems (called ISC, SUF and NIF) are found in prokaryotes. The function, protein composition and distribution of each system can differ across bacterial species [31]. Within each system, the individual steps required to assemble and deliver FeS clusters are executed by different enzymes. In the E. coli ISC pathway, reduced elemental sulfur is generated from cysteine by the cysteine desulfurase IscS and delivered to a cluster scaffold protein, IscU. In the SUF pathway, SufS generates elemental sulfur, while the SufBCD complex serves as the site of FeS cluster assembly [32]. The expression of FeS maturation pathways may also vary and be adapted to growth and environmental conditions. For instance, in E. coli, the protein ErpA delivers FeS clusters to the 4Fe-4S enzyme IspG in the presence of oxygen, while IscA mediates this delivery under anaerobic conditions [33].
Furthermore, the maturation of enzymes that contain multiple metalloclusters and/or protein components is far more complex, as multiple maturation steps must be coordinated. For instance, the Mo nitrogenase of Azotobacter vinelandii is a two-component enzyme comprised of (i) an Fe protein reductase (NifH) containing a [4Fe4S] cluster; (ii) a catalytic MoFe protein (the α2β2-tetramer NifDK) containing four complex metalloclusters: two M-clusters (or FeMo cofactor), and two P-clusters (Figure 1b). Therefore, multiple helper proteins are essential for its maturation and catalytic function. The maturation of the reductase NifH is ensured by the maturase NifM, and proteins NifUS for FeS cluster biosynthesis [29,34]. The in-situ assembly and maturation of the P-clusters in NifDK is a stepwise process performed by the proteins NifHZ. P-cluster assembly in NifDK is essential for the subsequent insertion of M-clusters. The biosynthesis and insertion of M-clusters is a highly complex process performed by the coordinated action of at least seven helper proteins [35]. The optimal expression and proper functioning of all these components are required for Mo nitrogenase catalysis in heterologous hosts.
2.2.2. Metalloenzyme Catalysis Often Requires Specific Electron Transfer Proteins
Many metallocluster enzymes are electronically coupled to small molecule reduced cofactors (i.e., NADH, NADPH, FAD) to sustain catalytic turnover. In some cases, these cofactors can directly deliver electrons to the enzyme, but the large majority of metalloclusters receive electrons via specific electron transfer proteins (ETPs), such as ferredoxins, flavodoxins or reductases [25,37]. The efficiency of electron transfer between protein partners is highly dependent on specific protein–protein contacts, redox potentials and protein concentrations [37]. Because the activity of many metallocluster enzymes is directly coupled to the abundance and efficiency of compatible ETPs, insufficient electron supply constitutes a major limitation to pathway optimization. For instance, the methylerythritol phosphate pathway (MEP) pathway for isoprenoid biosynthesis is known to be limited by the activity of two 4Fe-4S enzymes, IspG and IspH [13]. IspG/H both require an effective reducing partner that shuttles electrons from NADPH via a specific ETP [38]. Diverse studies have demonstrated that electron supply to these enzymes is a limiting factor in increasing the flux through the MEP pathway [38,39]. Insufficient electron transfer to FeS enzymes IspG and IspH could explain why the mevalonate pathway is still favored by biotechnologists for engineered isoprenoid synthesis, despite the higher theoretical carbon yield of the MEP pathway [40].
Moreover, the requirement for specific ETPs can prevent the functional expression of heterologous metallocluster enzymes. For instance, it has been demonstrated that the activity of IspG orthologs in E. coli is limited by the need for taxa-specific ETPs [30]. Similarly, different ETPs and cofactors are involved in nitrogenase activity depending on their genomic and metabolic backgrounds [41]. However, the identification of ETPs that support the activity of a heterologous metallocluster enzyme can be a challenging task, as most genes that encode them are not adjacent to metalloenzyme genes [30]. Identifying compatible ETPs is particularly crucial for ensuring enzyme activity in heterologous hosts, as strain engineers need to ensure that compatible ETPs are present in the host of choice. As the functional compatibilities between ETPs and metallocluster enzymes remain poorly characterized, their transferability across different organisms is often unpredictable. Therefore, difficulties in identifying efficient ETPs to sustain the activity of relevant metalloenzymes remain a major barrier to their full exploitation in microbial biosynthesis.
3. Strategies to the Promote Abundance of Functional Metallocluster Enzymes in Production Hosts
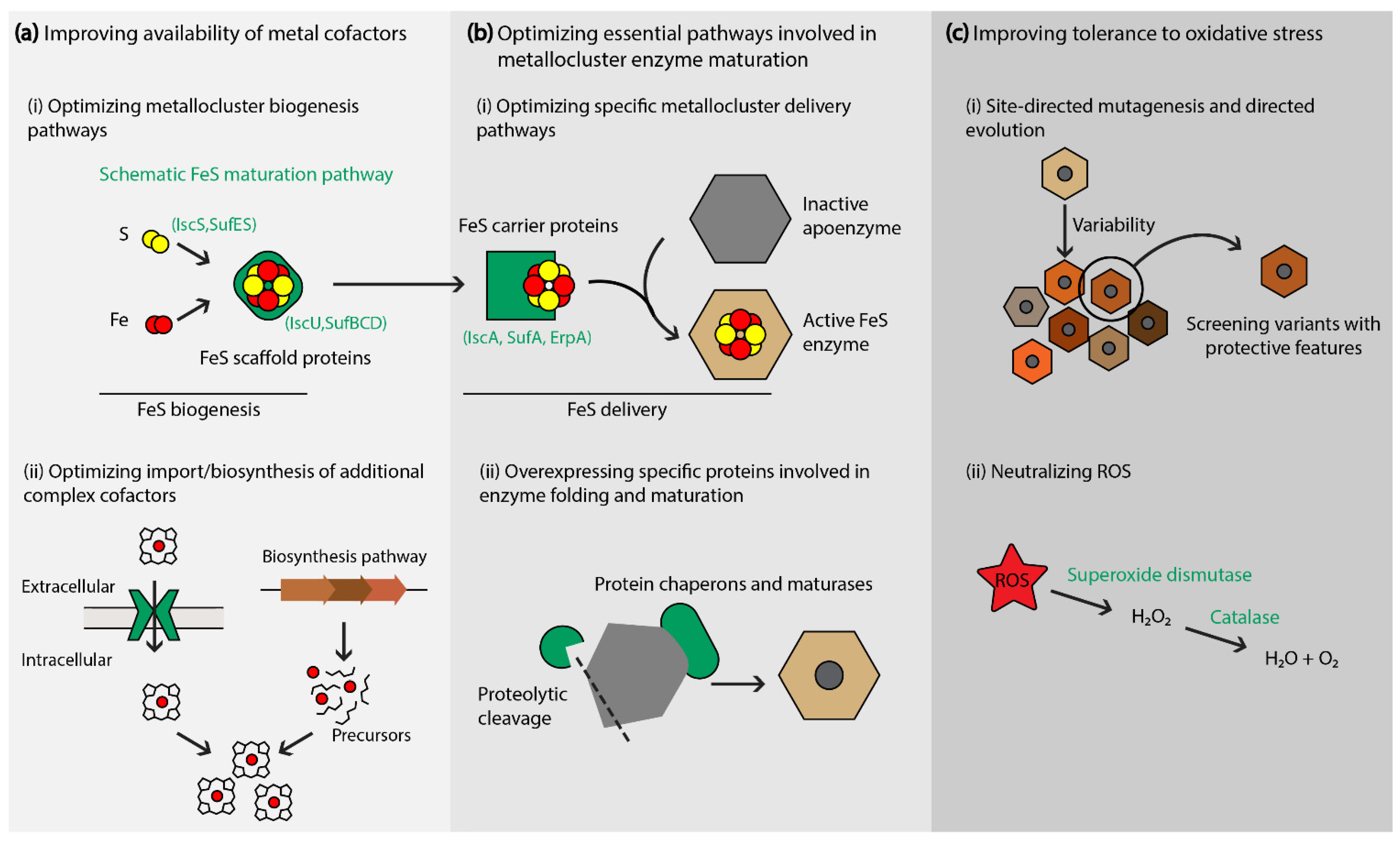
Strategies that enhance the peripheral molecular systems involved in metalloenzyme activity—maturation and electron transfer pathways—have been proven effective (and often necessary) in overcoming the challenges related to metabolic pathways that rely on recombinant metallocluster enzymes. The production of active metallocluster enzymes relies on the efficient synthesis and insertion of the metallic cofactor(s) into apoenzyme proteins [27]. The dedicated processes that assemble, deliver, and insert metallic clusters are coordinated by diverse helper proteins. Hence, the production of functional metalloenzymes requires the co-expression of helper proteins that ensure their maturation, activation and catalytic stability (Figure 2).
3.1. Optimizing Specific Maturation Pathways
Metalloenzyme maturation pathways are multi-protein systems that execute sequential steps coordinated through specific protein–protein interactions [42]. Such protein systems may be highly specific to the particular metallocluster enzyme and are thus unlikely to be found in heterologous hosts. In such cases, the co-expression of the required maturation pathways in the chassis of choice is the most direct way to ensure the maturation of a given enzyme. For instance, [Fe-Fe]- and [NiFe]-hydrogenases rely on a maturation pathway composed of a dozen proteins [25] that mediate subunit assembly, cofactor incorporation, and in some cases, proteolytic processing [43]. Regardless of this complexity, functional expression of heterologous hydrogenases from diverse organisms is possible when co-expressed with their respective maturation proteins in E. coli [44,45,46,47]. Similarly, the functional expression of a heterologous nitrogenase in E. coli was first achieved by transferring the full nif cluster from Klebsiella pneumoniae, which comprises all genes involved in nitrogenase maturation [48]. Recently, another study identified the minimal protein requirements for heterologous metallocluster biosynthesis and maturation of an engineered FeFe nitrogenase system in E. coli, comprised of only eight genes [49]. Both heterologous H2 biosynthesis and nitrogen fixation illustrate the success of co-expressing maturation pathways in achieving foreign metalloenzyme activity. However, the identification and expression of the required proteins may not be straightforward, hence still limiting the transfer of many interesting metallocluster enzymes [50].
On the contrary, some maturation pathways are ubiquitous and highly conserved across bacterial phyla. This is the case for the maturation pathways of FeS enzymes, which are essential to most cellular organisms (Figure 2). In a given prokaryote, these pathways can be encoded by up to three operons called SUF, ISC, or NIF. While FeS enzymes have an absolute requirement for FeS maturation pathways, compatible systems may be provided by the heterologous host. Although the different FeS cluster assembly and delivery systems rely on specific protein–protein interactions, their components show some promiscuity as they ensure the maturation of different FeS enzymes within the same species (up to 150) [28], and in some cases heterologous FeS enzymes [30,51]. The versatility of FeS maturation pathways notably enables the import FeS orthologs with improved kinetic properties, as demonstrated by the improved isoprenoid production resulting from the functional expression of IspG/H from T. elongatus in E. coli [39]. Therefore, strategies to boost the host’s FeS maturation pathways have been proven successful in optimizing pathways that rely on FeS enzymes. The enhanced expression of the isc operon, either by direct overexpression of the native isc genes [52] or by deletion of the negative regulator IscR [39], has been used for instance to improve the activities of IspG/H from the MEP pathway in E. coli. The production of antiviral molecules in E. coli by diverse prokaryotic Viperins also required the overexpression of the native isc operon by deletion of iscR [21].
However, many FeS enzymes lose their activity when heterologously expressed, indicating that the promiscuity of FeS maturation pathways does not necessarily extend beyond all species boundaries. Therefore, the construction of chassis strains that express heterologous FeS maturation pathways can be a strategy to restore the function of foreign FeS enzymes. For instance, the over-expression of the ISC operon from Azetobacter vinelandii is a common approach used by biochemistry groups to ensure functional expression of foreign FeS enzymes [16,47,49]. Yet, using the A. vinelandii pathway is not sufficient to deliver FeS clusters to all FeS. Little is known about functional versatility of FeS maturation pathways across different organisms, as the protein components and mechanisms required for FeS assembly and incorporation can differ across species [28]. However, a recent study demonstrated that the co-expression of heterologous FeS maturation pathways effectively increases the phylogenetic range of foreign FeS enzymes that can be functionally expressed. Indeed, the expression of the SUF operon from B. subtilis not only restored the function of B. subtilis NadA in E. coli, but also activated eight NadA orthologs from diverse phylogenetic origins [30]. Consequently, the expression of FeS maturation pathways may often broaden host compatibility with certain heterologous FeS enzymes. The cross-species compatibility of FeS maturation pathways also remains to be systematically explored. Furthermore, as all maturation pathway components should be expressed at an appropriate stoichiometry relative to other components, optimizing the abundance of each maturation protein may present an additional challenge for expressing heterologous maturation pathways. Further studies are required to determine the functionality of specific maturation pathways in heterologous hosts, especially pathways that require refactoring due to codon optimization.
3.2. Increasing the Synthesis of Additional Essential Metal Cofactors
In addition to metal clusters, additional complex metal cofactors—such as hemes or vitamins—may be required for catalytic activity. Such cofactors are not available in many popular chassis strains or are produced at insufficient levels for high pathway performance. Engineering and optimizing cofactor import/biosynthesis in host strains is therefore an efficient approach to enable/improve the activity of metallocluster enzymes that rely on them. For instance, the heme supply in E. coli was improved by optimizing the endogenous production of its precursor 5-aminolevulinic acid (ALA), which subsequently enhanced the activity of heterologous heme-containing enzymes [53]. Another example is the need of cobalamin (vitamin B12) for the activity of diverse radical SAM FeS methyltransferases. The activity these enzymes can be enhanced by co-expressing cobalamin import and trafficking pathways [54], such as the btu operon from E. coli, which enabled the functional expression of S. laurentii TsrM in E. coli [30]. Alternatively, engineering hosts for de novo biosynthesis of cobalamin could be an approach to optimize pathways that rely on B12-dependent FeS enzymes [55].
3.3. Improving the Tolerance of Metallic Clusters to Oxidative Stress
The stability of some metallocluster enzymes is greatly hindered by oxidative stress caused by molecular oxygen and/or reactive oxygen species (ROS). The catalytic inactivation of hydrogenases by O2, is considered as the major limitation in their biotechnological use for hydrogen production. Similarly, oxidative stress was identified as one of the barriers to the functionality of the bacterial FeS enzymes IspG and IspH in yeast [56], implying that cluster stability after initial FeS cluster assembly is also a limiting factor in metalloenzyme transferability.
Nature has evolved specific amino-acid features and structural arrangements that improve the tolerance of metallic clusters to oxidative damage and that are essential to their stability in aerobic conditions [57]. For a metallocluster enzyme of interest, selecting analogs that have naturally evolved protective features against oxidative stress can be a strategy to stabilize activity in foreign organisms and aerobic conditions. For instance, the O2-tolerant membrane-bound [NiFe] hydrogenase (MBH) from Ralstonia eutropha holds a special [4Fe-3S] cluster that enables the enzyme to reduce O2 into harmless water [58]. Heterologous H2 production has been achieved in aerobic conditions by overexpressing MBH homologs in E. coli. Cultures expressing functional MBH from Hydrogenovibrio marinus increased hydrogen production by two-fold compared to cultures overexpressing the oxygen sensitive hydrogenase Hyd-1 from E. coli [59]. Additionally, site-directed mutagenesis and directed evolution can be a reliable method to evolve such protective features. The identification of key residues involved in oxygen sensitivity of the [NiFe] hydrogenase from Desulfovibrio fructosovorans enabled the design of a mutated variant with improved O2 tolerance and faster reactivation times [60]. In combination with structural and mechanistic studies, these techniques could lead the rational design of enzymes with improved oxygen tolerance without compromising protein function.
Moreover, some metallocluster proteins are protected from oxygen damage by specific helper proteins found in native organisms, which can be co-expressed in heterologous hosts to improve metallocluster stability. For example, the Shethna Protein II [61] and Anf3 oxidase [62] from A. vinelandii protect nitrogenase activity from inactivation by oxygen. Additional strategies could rely on the co-expression of enzymes that neutralize ROS, such as superoxide dismutases or catalases, which are known to protect FeS clusters in aerobic organisms [63]. The optimization of natural FeS cluster repair systems in chassis strains could be investigated to improve the performance of FeS enzymes. Expression of metalloenzymes in mitochondria may also protect oxygen-sensitive enzymes from damage, as discussed further below.
4. Optimizing Electron Transfer Pathways to Boost Redox Active Metallocluster Enzymes
Because the activity of redox active metallocluster enzymes relies on efficient electron supply, optimizing electron transfer pathways linked to biosynthetic enzymes can be an effective strategy to improve pathway performance (Figure 3). This approach is especially relevant when cluster-containing recombinant enzymes display no/low activity in the production host or when direct manipulations of a metabolic pathway (e.g., balancing multi-enzymatic expression) are proven insufficient to further increase product synthesis. To this purpose, the identification, production, and optimization of essential redox partners is crucial.
4.1. Identification of Redox Partners Required for Metallocluster Enzyme Activity
To sustain the activity of a heterologous redox active metalloenzyme, it is necessary to ensure that compatible ETPs and reduced cofactors are present in the host. Diverse in vitro assays have been successfully employed to identify the small-molecule cofactors and ETPs that are naturally coupled to metallocluster enzymes. For certain enzymes found in secondary metabolic pathways, the genes that encode partner ETPs can often be within the same biosynthetic gene cluster. If the specific partner ETPs are unknown or are not contained within the same biosynthetic gene cluster, a bioinformatics analysis could help identify potential redox partners in the genomes of native or heterologous organisms. For instance, putative electron delivery systems needed for nitrogenase activity have been identified through genome mining [41]. However, while prokaryotic genomes usually encode multiple ETPs, no universal bioinformatics method to identify partner ETPs for a given enzyme currently exists. Yet, since ETPs display some functional promiscuity and can mediate electron transfer to a diversity of interacting proteins, cellular assays have been successful in identifying compatible ETPs for target enzymes [30,37]. These studies demonstrate that both binding specificity and reduction potentials determine the favorable interactions between ETPs and heterologous metallocluster enzymes. For instance, the expression of ferredoxins from diverse species can support the activity of heterologous IspG homologs from different organisms in E. coli [30]. This functional versatility can be an advantage in optimizing electron supply to relevant biosynthetic metallocluster enzymes.
4.2. Increasing the Amount of Redox Partners Enhances Electron Supply and Catalytic Activity
4.2.1. Improving Availability of Small-Molecule Cofactors
A straightforward approach to increase electron supply to an enzyme of interest is to optimize the regeneration and availability of its small-molecule redox partner(s) [64]. For instance, strategies that improve NADPH pools can result in increased performance of pathways limited by NADPH-dependent FeS cluster enzymes [38]. The deletion of an enzyme that consumes NADPH in E. coli, coupled with expression of a NADH kinase from S. cerevisiae, boosted the metabolic flux towards IspG/H enzymes of the MEP pathway.
4.2.2. Overexpression of Partner ETPs to Direct Electron Flow to Metallocluster Enzymes
More targeted approaches to increase the electron flow through given biosynthetic metalloenzymes involve the manipulation of specific redox chains composed of partner ETPs. Because ETPs are genetically encoded, synthetic biology tools can be employed to control their expression, improve interaction affinities with protein partners, or control their allosteric conformations [37].
Approaches that boost the expression of partner ETPs enhance electron supply to specific metallocluster enzymes and improve pathway performance. For instance, overexpression of FldA and Fpr, the partner ETPs of the FeS enzymes IspG/H from the MEP pathway in E. coli, improved titers of an isoprenoid product by three-fold [38]. In cyanobacteria, the activity of IspG/H is modulated by a different set of ETPs: the ferredoxin PetF and its associated ferredoxin-NADP+ reductase PetH. Isoprene production was increased by 1.5-fold in Synechocystis sp. PCC 6803 resulting from the overexpression of PetF/H [65]. Improved metabolic flux through the MEP pathway in E. coli was achieved by the heterologous co-expression of IspG/H from Thermosynechococcus elongatus combined with PetF/H from the same organism (2.7-fold improvement) [39].
The ability of ETPs to mediate electron transfer between different partner proteins, and their functionality across a wide range of hosts, make them useful tools for engineering synthetic redox chains that drive heterologous metalloenzyme activity [66,67]. For instance, a recent study explored the cross-species compatibility of diverse IspG orthologs with heterologous electron transfer proteins obtained from different species in E. coli [30]. The results show that IspG orthologs exhibit activity with multiple ETPs, and vice-versa. This study also identified ETPs from Streptomyces cattleya that sustain the activity of the rSAM FeS enzyme TsrM from Streptomyces laurentii in E. coli, highlighting the importance of expressing surrogate ETPs. Another study identified housekeeping ETP orthologs of NifJ and NifF to partially support an artificial nitrogenase system in E. coli, a surrogate electron transport pathway that could be further optimized [49].
Finally, the overexpression of ETPs can lead to excessive redox activities and to the accumulation of reactive oxygen species, causing global imbalances that hinder cell viability and production [37]. Therefore, it is important to tightly regulate the expression of reducing partners (relative to their target enzymes) to optimize electron flux without compromising cell viability.
4.3. Insulation Strategies for Electron Transfer Pathways
4.3.1. Removing Competing Reactions
Increasing electron flow to specific metalloenzymes can achieved by preventing electron transfer to other biochemical processes in the cell. An approach to redirect electron flow into a pathway of interest is to remove competing electron acceptors from the host. Agapakis et al. increased hydrogen production in E. coli by 40% by deleting a gene encoding a protein that was suspected to interact with the synthetic electron transfer pathway of [Fe-Fe] hydrogenases [46]. Moreover, as protein–protein binding affinities control electron flow between redox partners, targeted mutations in ETPs can reduce or suppress their interaction with competing binding partners, hence redirecting electron flow to biosynthetic metalloenzymes and improving product yields [43,61]. For instance, targeted mutations of a hydrogenase at the interaction surface to enhance the charge-complementarity with a partner ferredoxin improved hydrogen production up to two-fold, compared to the wild-type hydrogenase [46].
4.3.2. Covalent Fusions with Redox Partner
Other methods to enhance electron transfer between desired protein partners are based on increasing their relative local concentration within the cell. Indeed, incrementing the chance of contact between enzymes and ETPs is an efficient strategy to direct the flux into biosynthetic pathways of interest. Improved enzymatic activity and pathway performance can be achieved by directly fusing metalloenzymes to their reducing partner using genetic engineering tools. Such chimeric protein complexes can be synthetically constructed by fusing the desired partners through protein linkers. For instance, the direct fusion of the hydrogenase from C. acetobutylicum with a ferredoxin resulted in a 4.4-fold increase in hydrogen production [46]. However, it is difficult to predict how these covalent fusions will impact enzymatic activity [46].
The physical linkage of metalloenzymes with electron carriers can also be employed to engineer alternative electron transfer pathways with improved efficiency. For instance, alternative electron donors have been engineered in E. coli to uncouple the activity of the [NiFe]-hydrogenase-3 (Hyd-3) from its natural electron donor formate. An engineered Hyd-3 covalently attached to a ferredoxin from Thermotoga maritima accepts electron from pyruvate instead, and sustains in vivo hydrogen production when co-expressed with a pyruvate-ferredoxin oxidoreductase (PFOR) [68].
4.3.3. Spatial Organization of Enzymes and Partner ETPs
Since the physical interaction between partner proteins is critical for electron transfer, their spatial organization can be optimized to enhance electron flux through desired metabolic pathways. Optimized spatial organization of electron donor and acceptor proteins increases their chance of physical contact, and reduces cross-talk with other cellular pathways [69]. Molecular scaffolds can be designed to attach desired redox partner proteins and increase pathway yields in vivo. This approach has been applied to optimize the spatial organization of hydrogenases and ferredoxins to improve hydrogen production in E. coli. Attachment of a hydrogenase and a ferredoxin on a synthetic protein scaffold improved hydrogen production by three-fold [46]. The use of rationally designed RNA structures with protein-docking sites to optimize spatial organization and electron transfer boosted hydrogen yields by 50-fold [70]. Other strategies based on subcellular compartmentalization to encapsulate metabolic pathways can be employed to enhance electron transfer and pathway performance [71].
5. Towards Transferring Metallocluster Enzymes from Bacteria to Eukaryotes
Eukaryotic organisms, such as yeast or microalgae, are essential hosts in the development of microbial cell factories. As many industrial bioprocesses use eukaryotic microbial hosts, implementing biotechnologically relevant pathways in these organisms is crucial to the large-scale production of chemicals. Unfortunately, attempts of transferring pathways that rely on the activity of bacterial metallocluster enzymes into eukaryotes have encountered limited success. For instance, many bacterial FeS enzymes involved in industrially relevant pathways display little-to-no activity when expressed in yeast, and common synthetic biology interventions have failed to improve their activity [63]. Attempts to engineer the MEP pathway [72], the Entner–Doudoroff pathway [73], or the Weimberg pathway [74] in yeast have met limited success due to the poor activity of bacterial FeS cluster enzymes. The specific challenges encountered when expressing bacterial metalloenzymes within eukaryotes are more fully explored in reference [63], but are briefly discussed here.
The barriers to the functional expression of recombinant metallocluster enzymes in eukaryotic organisms are analogous to those that exist between bacterial species: (i) inefficient maturation and/or electron supply due to incompatibilities between phyla-specific systems; and (ii) oxidative stress. The co-expression of bacterial FeS maturation and reducing pathways in yeast is unfortunately insufficient to enhance enzymatic activity, as demonstrated for the enzymes IspG and IspH [75]. Other strategies focused on disturbing the host’s iron regulation to increase FeS cluster availability have been successfully employed to improve IlvD activity and isobutanol production [76]. Finally, the compartmentalization of bacterial FeS enzymes in yeast organelles (i.e., mitochondria or chloroplasts) remains another promising strategy to sustain their activity in eukaryotes. Indeed, these organelles contain FeS maturation pathways that resemble bacterial systems, as well as compatible ETPs to support catalysis [77]. Moreover, compartmentalization significantly reduces oxidative stress of metallocluster enzymes, hence increasing their catalytic stability [11]. While these strategies are promising, they remain insufficient to the development of viable yeast bioprocesses and additional research is required.
Engineering nitrogen fixation in eukaryotes by expressing bacterial nitrogenases has been a long-sought goal due to the cost of synthetic nitrogen fertilizers; however, nitrogen fixation in plants for self-fertilization has not been achieved due to difficulties in expressing functional nitrogenase systems [78]. Nevertheless, nitrogenase expression in eukaryotes serves as a case study that demonstrates the progress made through the use of many approaches described in this review. The oxygen sensitivity of the nitrogenase maturation pathway was circumvented by targeting expression to the mitochondria [79]. Highlighting the importance of identifying specific electron transfer proteins, a recent study used an E. coli cell chassis to determine that electron transfer pathways from chloroplasts, but not mitochondria, are capable of supporting bacterial nitrogenase systems [80]. More recently, 32 orthologs of an iron reductase (NifH) were expressed in tobacco mitochondria, together with protein cofactors involved with FeS transfer [81]. This work specifically identified Fe-S cluster assembly as a significant barrier to nitrogenase function, underscoring the need for further research.
6. Conclusions and Perspectives
The aim of making biology easier to engineer is the central goal of synthetic biology. To this purpose, tools and standardized principles for manipulating DNA have been developed over the past two decades. Genetic engineering technologies have enabled the engineering of a wide range of organisms with novel biological functions, including biotechnologically relevant microbial cell factories. However standard synthetic biology tools focused on genetic manipulation have faced challenges in engineering pathways that rely on many catalytically powerful metalloenzymes. Here, we presented an outline of the obstacles to the functional expression and optimization of heterologous metallocluster enzymes in biosynthetic pathways. We also described novel synthetic biology strategies and investigations that could enable harnessing their full biotechnological potential. Such efforts are focused on overcoming obstacles specific to the functional requirements of catalytic metallocluster enzymes: (i) engineering essential supporting pathways involved in enzyme stability and maturation; and (ii) engineering pathways for electron supply that sustain redox active catalysis. The combination and further development of these strategies could be used to optimize novel microbial cell factories with economically viable yields.
However, major challenges to the full exploitation of these engineering approaches remain, as our fundamental knowledge on the biochemistry and barriers to the heterologous expression of metallocluster enzymes is still very limited. Indeed, synthetic biologists cannot easily predict the transferability of a given metallocluster enzyme, nor anticipate what will be required for their activation in heterologous hosts. More studies of phylogenetic compatibility and functional requirements will help develop intuitive rules for harnessing heterologous metallocluster enzymes. To this purpose, original research at the intersection between biochemistry, metabolomics, functional genomics, and synthetic biology is necessary. For instance, large-scale cellular assays could be developed to screen the in vivo functionality of relevant metallocluster enzymes in foreign hosts, and to address the barriers to their transferability and catalytic activity [30]. Furthermore, such studies could also help developing strategies to transfer bacterial metalloenzymes into industrial eukaryotic hosts.
Moreover, recent advances in synthetic biology have enabled de novo engineering of artificial metallocluster enzymes by incorporating catalytic metallocofactors (including abiotic cofactors) into designed protein scaffolds. These artificial enzymes catalyze a remarkable range of natural and synthetic reactions [82], and their biosynthetic potential is tremendous. Yet, the same challenges encountered with natural metalloenzymes often limit their activity, including electron transfer efficiency [83] and catalytic stability [82]. Strategies to enhance the maturation, stability and biochemistry of natural metallocluster enzymes will likely be of significant relevance for improving artificial or bioinspired catalysts [84]. Moreover, designing custom FeS proteins that mediate tunable electron transfer could be of considerable interest for engineering novel redox chains to support metallocluster enzyme activity [85,86]. For instance, synthetic chemical-dependent ETPs have been recently designed to support electron transfer in vivo, in the presence of the chemical inducer rapamycin [85]. Such protein switches could represent a new class of ETPs for the precise regulation of electron transfer across a wide range of redox potentials with direct applications in metabolic engineering.
Besides their potential for microbial biosynthesis, metallocluster enzymes participate in a wide range of key biological processes across kingdoms, including some of technological importance, such as antiviral [21] or antibiotic resistance [87]. Therefore, the relevance of understanding their biochemistry and addressing the barriers to their activity goes beyond applications to microbial cell factories.
Author Contributions
Conceptualization, H.S.; writing—original draft preparation, H.S.; writing—review and editing, G.B.; supervision, G.B. All authors have read and agreed to the published version of the manuscript.
Funding
Work has been supported with funding from the European Union’s Horizon 2020 research and innovation program under grant 722361 (project IRONPLUGNPLAY).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Frédéric Barras, Beatrice Py, Sandrine Ollagnier de Choudens, and members of the Bokinsky Lab for valuable discussions.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
Table A1.
Crystal structures of metalloenzymes.
| Enzyme | Origin | Uniprot ID | PDB ID |
|---|---|---|---|
| Fe-Fe hydrogenase | Clostridium pasteurianum | P29166 | 1FEH |
| Ni-Fe hydrogenase | Desulfovibrio fructosovorans | P18187 | 1FRF |
| [2Fe-2S] ferredoxin FdxB | Pseudomonas putida | Q76CS9 | 3AH7 |
| D-xylonate dehydratase | Caulobacter vibrioides CB15 | Q9A9Z2 | 5OYN |
| [2Fe-2S] and [4Fe-4S] rSAM enzyme BioB | Escherichia coli | P12996 | 1R30 |
| Nitrogenase molybdenum-iron protein | Azotobacter vinelandii | P07328 | 6UG0 |
| [4Fe-4S] Viperin | Nematostella vectensis | A7RNF3 | 7N7H |
| [4Fe-4S] enzyme IspG | Aquifex aeolicus | O67496 | 3NOY |
References
- Nielsen, J.; Keasling, J.D. Engineering Cellular Metabolism. Cell 2016, 164, 1185–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jullesson, D.; David, F.; Pfleger, B.; Nielsen, J. Impact of synthetic biology and metabolic engineering on industrial production of fine chemicals. Biotechnol. Adv. 2015, 33, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, M.; Lee, S.Y. Prospects of microbial cell factories developed through systems metabolic engineering. Microb. Biotechnol. 2016, 9, 610–617. [Google Scholar] [CrossRef]
- Becker, J.; Wittmann, C. Systems metabolic engineering of Escherichia coli for the heterologous production of high value molecules—A veteran at new shores. Curr. Opin. Biotechnol. 2016, 42, 178–188. [Google Scholar] [CrossRef]
- Robertsen, H.L.; Weber, T.; Kim, H.U.; Lee, S.Y. Toward Systems Metabolic Engineering of Streptomycetes for Secondary Metabolites Production. Biotechnol. J. 2018, 13, 1700465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsui, R.; Yamada, R. Saccharomyces cerevisiae as a microbial cell factory. In Microbial Cell Factories Engineering for Production of Biomolecules; Academic Press: Cambridge, MA, USA, 2021; pp. 319–333. [Google Scholar] [CrossRef]
- Stephanopoulos, G. Synthetic Biology and Metabolic Engineering. ACS Synth. Biol. 2012, 1, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.C. Great Metalloclusters in Enzymology. Annu. Rev. Biochem. 2002, 71, 221–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R. A comparison of types of catalyst: The quality of metallo-enzymes. J. Inorg. Biochem. 2008, 102, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.S.; Drennan, C.L. Metalloenzymes in natural product biosynthetic pathways. Nat. Prod. Rep. 2018, 35, 612–614. [Google Scholar] [CrossRef] [PubMed]
- Bloch, S.E.; Ryu, M.-H.; Ozaydin, B.; Broglie, R. Harnessing atmospheric nitrogen for cereal crop production. Curr. Opin. Biotechnol. 2020, 62, 181–188. [Google Scholar] [CrossRef]
- Shen, L.; Kohlhaas, M.; Enoki, J.; Meier, R.; Schönenberger, B.; Wohlgemuth, R.; Kourist, R.; Niemeyer, F.; Van Niekerk, D.; Bräsen, C.; et al. A combined experimental and modelling approach for the Weimberg pathway optimisation. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Chang, W.-C.; Xiao, Y.; Liu, H.-W.; Liu, P. Methylerythritol Phosphate Pathway of Isoprenoid Biosynthesis. Annu. Rev. Biochem. 2013, 82, 497–530. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Zhou, J.; Zhang, Z.; Kim, C.H.; Jiang, B.; Shi, J.; Hao, J. Isobutanol and 2-ketoisovalerate production by Klebsiella pneumoniae via a native pathway. Metab. Eng. 2017, 43, 71–84. [Google Scholar] [CrossRef]
- Streit, W.R.; Entcheva, P. Biotin in microbes, the genes involved in its biosynthesis, its biochemical role and perspectives for biotechnological production. Appl. Microbiol. Biotechnol. 2003, 61, 21–31. [Google Scholar] [CrossRef]
- Marous, D.R.; Lloyd, E.P.; Buller, A.R.; Moshos, K.A.; Grove, T.L.; Blaszczyk, A.J.; Booker, S.J.; Townsend, C.A. Consecutive radical S-adenosylmethionine methylations form the ethyl side chain in thienamycin biosynthesis. Proc. Natl. Acad. Sci. USA 2015, 112, 10354–10358. [Google Scholar] [CrossRef] [Green Version]
- Freeman, M.F.; Vagstad, A.L.; Piel, J. Polytheonamide biosynthesis showcasing the metabolic potential of sponge-associated uncultivated ‘Entotheonella’ bacteria. Curr. Opin. Chem. Biol. 2016, 31, 8–14. [Google Scholar] [CrossRef]
- Sato, S.; Kudo, F.; Kim, S.-Y.; Kuzuyama, T.; Eguchi, T. Methylcobalamin-Dependent Radical SAM C-Methyltransferase Fom3 Recognizes Cytidylyl-2-hydroxyethylphosphonate and Catalyzes the Nonstereoselective C-Methylation in Fosfomycin Biosynthesis. Biochemistry 2017, 56, 3519–3522. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Guo, J.; Reva, A.; Huang, F.; Xiong, B.; Liu, Y.; Deng, Z.; Leadlay, P.F.; Sun, Y. Methyltransferases of gentamicin biosynthesis. Proc. Natl. Acad. Sci. USA 2018, 115, 1340–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Xu, H.; Li, Y.; Zhang, F.; Chen, X.-Y.; He, Q.-L.; Igarashi, Y.; Tang, G.-L. Characterization of Yatakemycin Gene Cluster Revealing a Radical S-Adenosylmethionine Dependent Methyltransferase and Highlighting Spirocyclopropane Biosynthesis. J. Am. Chem. Soc. 2012, 134, 8831–8840. [Google Scholar] [CrossRef] [PubMed]
- Bernheim, A.; Millman, A.; Ofir, G.; Meitav, G.; Avraham, C.; Shomar, H.; Rosenberg, M.M.; Tal, N.; Melamed, S.; Amitai, G.; et al. Prokaryotic viperins produce diverse antiviral molecules. Nat. Cell Biol. 2021, 589, 120–124. [Google Scholar] [CrossRef]
- Fang, H.; Kang, J.; Zhang, D. Microbial production of vitamin B12: A review and future perspectives. Microb. Cell Factories 2017, 16, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, J.C.; Khusnutdinova, A.N.; Flick, R.; Kim, T.; Bornscheuer, U.T.; Yakunin, A.F.; Mahadevan, R. Alkene hydrogenation activity of enoate reductases for an environmentally benign biosynthesis of adipic acid. Chem. Sci. 2017, 8, 1406–1413. [Google Scholar] [CrossRef] [Green Version]
- Micallef, M.L.; Sharma, D.; Bunn, B.M.; Gerwick, L.; Viswanathan, R.; Moffitt, M.C. Comparative analysis of hapalindole, ambiguine and welwitindolinone gene clusters and reconstitution of indole-isonitrile biosynthesis from cyanobacteria. BMC Microbiol. 2014, 14, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, J.W.; Schut, G.J.; Boyd, E.S.; Mulder, D.W.; Shepard, E.M.; Broderick, J.B.; King, P.W.; Adams, M.W.W. [FeFe]- and [NiFe]-hydrogenase diversity, mechanism, and maturation. Biochim. Biophys. Acta—Mol. Cell Res. 2015, 1853, 1350–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekhar, K.; Lee, Y.-J.; Lee, D.-W. Biohydrogen Production: Strategies to Improve Process Efficiency through Microbial Routes. Int. J. Mol. Sci. 2015, 16, 8266–8293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldron, K.; Robinson, N. How do bacterial cells ensure that metalloproteins get the correct metal? Nat. Rev. Microbiol. 2009, 7, 25–35. [Google Scholar] [CrossRef]
- Roche, B.; Aussel, L.; Ezraty, B.; Mandin, P.; Py, B.; Barras, F. Iron/sulfur proteins biogenesis in prokaryotes: Formation, regulation and diversity. Biochim. Biophys. Acta—Bioenerg. 2013, 1827, 923–937. [Google Scholar] [CrossRef] [Green Version]
- Rubio, L.M.; Ludden, P.W. Maturation of Nitrogenase: A Biochemical Puzzle. J. Bacteriol. 2005, 187, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Shomar, H.; Garcia, P.S.; Fernández-Fueyo, E.; D’angelo, F.; Pelosse, M.; Manuel, R.R.; Büke, F.; Liu, S.; van den Broek, N.; Duraffourg, N.; et al. Resolving phylogenetic and biochemical barriers to functional expression of heterologous iron-sulphur cluster enzymes. bioRxiv 2021. bioRxiv:2021.02.02.429153. [Google Scholar]
- Johnson, D.C.; Dean, D.R.; Smith, A.D.; Johnson, M.K. Structure, function, and formation of biological iron-sulfur clusters. Annu. Rev. Biochem. 2005, 74, 247–281. [Google Scholar] [CrossRef]
- Yokoyama, N.; Nonaka, C.; Ohashi, Y.; Shioda, M.; Terahata, T.; Chen, W.; Sakamoto, K.; Maruyama, C.; Saito, T.; Yuda, E.; et al. Distinct roles for U-type proteins in iron–sulfur cluster biosynthesis revealed by genetic analysis of the Bacillus subtilis sufCDSUB operon. Mol. Microbiol. 2018, 107, 688–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinella, D.; Brochier-Armanet, C.; Loiseau, L.; Talla, E.; Barras, F. Iron-Sulfur (Fe/S) Protein Biogenesis: Phylogenomic and Genetic Studies of A-Type Carriers. PLoS Genet. 2009, 5, e1000497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, P.C.; Johnson, D.C.; Ragle, B.E.; Unciuleac, M.C.; Dean, D.R. Controlled expression of nif and isc iron-sulfur protein maturation components reveals target specificity and limited functional replacement between the two systems. J. Bacteriol. 2007, 189, 2854–2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Ribbe, M.W. Nitrogenase assembly. Biochim. Biophys. Acta—Bioenerg. 2013, 1827, 1112–1122. [Google Scholar] [CrossRef] [Green Version]
- Biphenyls, C.P. Structure and Function of Four Classes of the 4Fe-4S Protein, IspH. Biochemistry 2016, 55, 4119–4129. [Google Scholar]
- Atkinson, J.; Campbell, I.; Bennett, G.N.; Silberg, J.J. Cellular Assays for Ferredoxins: A Strategy for Understanding Electron Flow through Protein Carriers That Link Metabolic Pathways. Biochemistry 2016, 55, 7047–7064. [Google Scholar] [CrossRef]
- Zhou, J.; Yang, L.; Wang, C.; Choi, E.-S.; Kim, S.-W. Enhanced performance of the methylerythritol phosphate pathway by manipulation of redox reactions relevant to IspC, IspG, and IspH. J. Biotechnol. 2017, 248, 1–8. [Google Scholar] [CrossRef]
- Chotani, G.K.; Mcauliffe, J.C.; Miller, M.C.; Muir, R.E.; Vavlline, D.V.; Weyler, W. Isoprene Production Using the DXP and MVA Pathway. U.S. Patent 8,507,235 B2, 20 January 2011. [Google Scholar]
- Vickers, C.E.; Sabri, S. Isoprene. In Biotechnology of Isoprenoids; Schrader, J., Bohlmann, J., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 289–317. [Google Scholar]
- Poudel, S.; Colman, D.R.; Fixen, K.R.; Ledbetter, R.N.; Zheng, Y.; Pence, N.; Seefeldt, L.C.; Peters, J.W.; Harwood, C.S.; Boyd, E.S. Electron Transfer to Nitrogenase in Different Genomic and Metabolic Backgrounds. J. Bacteriol. 2018, 200. [Google Scholar] [CrossRef] [Green Version]
- Kuchar, J.; Hausinger, R.P. Biosynthesis of Metal Sites. Chem. Rev. 2004, 104, 509–526. [Google Scholar] [CrossRef]
- Senger, M.; Stripp, S.T.; Soboh, B. Proteolytic cleavage orchestrates cofactor insertion and protein assembly in [NiFe]-hydrogenase biosynthesis. J. Biol. Chem. 2017, 292, 11670–11681. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Hopkins, R.C.; Jenney, F.E.; McTernan, P.M.; Adams, M.W.W. Heterologous expression and maturation of an NADP-Dependent [NiFe]-Hydrogenase: A key enzyme in biofuel production. PLoS ONE 2010, 5, e10526. [Google Scholar] [CrossRef] [Green Version]
- Weyman, P.D.; Vargas, W.A.; Chuang, R.-Y.; Chang, Y.; Smith, H.O.; Xu, Q. Heterologous expression of Alteromonas macleodii and Thiocapsa roseopersicina [NiFe] hydrogenases in Escherichia coli. Microbiology 2011, 157, 1363–1374. [Google Scholar] [CrossRef] [Green Version]
- Agapakis, C.M.; Ducat, D.C.; Boyle, P.M.; Wintermute, E.H.; Way, J.C.; Silver, P.A. Insulation of a synthetic hydrogen metabolism circuit in bacteria. J. Biol. Eng. 2010, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Rosales-Colunga, L.M.; Rodríguez, A.D.L. Escherichia coli and its application to biohydrogen production. Rev. Environ. Sci. Bio/Technol. 2015, 14, 123–135. [Google Scholar] [CrossRef]
- Dixon, R.A.; Postgate, J.R. Transfer of Nitrogen-fixation Genes by Conjugation in Klebsiella pneumoniae. Nature 1971, 234, 47–48. [Google Scholar] [CrossRef]
- Yang, J.; Xie, X.; Wang, X.; Dixon, R.; Wang, Y. Reconstruction and minimal gene requirements for the alternative iron-only nitrogenase in Escherichia coli. Proc. Natl. Acad. Sci. USA 2014, 111, E3718–E3725. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Li, M.; Li, J.; Wang, G.; Li, F.; Xiong, M. Chaperone-assisted maturation of the recombinant Fe-type nitrile hydratase is insufficient for fully active expression in Escherichia coli. Process. Biochem. 2017, 56, 37–44. [Google Scholar] [CrossRef]
- Zheng, L.; Cash, V.L.; Flint, D.H.; Dean, D.R. Assembly of Iron-Sulfur Clusters—Identification of an iscSUA-hscBA-fdx gene cluster from Azotobacter vinelandii. J. Biol. Chem. 1998, 273, 13264–13272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zepeck, F.; Gräwert, T.; Kaiser, J.; Schramek, N.; Eisenreich, W.; Bacher, A.A.; Rohdich, F. Biosynthesis of Isoprenoids. Purification and Properties of IspG Protein from Escherichia coli. J. Org. Chem. 2005, 70, 9168–9174. [Google Scholar] [CrossRef] [PubMed]
- Ramzi, A.B.; Hyeon, J.E.; Han, S.O. Improved catalytic activities of a dye-decolorizing peroxidase (DyP) by overexpression of ALA and heme biosynthesis genes in Escherichia coli. Process. Biochem. 2015, 50, 1272–1276. [Google Scholar] [CrossRef]
- Lanz, N.D.; Blaszczyk, A.J.; McCarthy, E.L.; Wang, B.; Wang, R.X.; Jones, B.S.; Booker, S.J. Enhanced Solubilization of Class B Radical S-Adenosylmethionine Methylases by Improved Cobalamin Uptake in Escherichia coli. Biochemistry 2018, 57, 1475–1490. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Li, D.; Kang, J.; Jiang, P.; Sun, J.; Zhang, D. Metabolic engineering of Escherichia coli for de novo biosynthesis of vitamin B12. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kirby, J.; Dietzel, K.L.; Wichmann, G.; Chan, R.; Antipov, E.; Moss, N.; Baidoo, E.E.; Jackson, P.; Gaucher, S.P.; Gottlieb, S.; et al. Engineering a functional 1-deoxy-D-xylulose 5-phosphate (DXP) pathway in Saccharomyces cerevisiae. Metab. Eng. 2016, 38, 494–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imlay, J.A.; Sethu, R.; Rohaun, S.K. Evolutionary adaptations that enable enzymes to tolerate oxidative stress. Free Radic. Biol. Med. 2019, 140, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Kalms, J.; Schmidt, A.; Frielingsdorf, S.; Utesch, T.; Gotthard, G.; von Stetten, D.; van der Linden, P.; Royant, A.; Mroginski, M.A.; Carpentier, P.; et al. Tracking the route of molecular oxygen in O2-tolerant membrane-bound [NiFe] hydrogenase. Proc. Natl. Acad. Sci. USA 2018, 115, E2229–E2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.Y.H.; Jo, B.H.; Cha, H.J. Production of biohydrogen by heterologous expression of oxygen-tolerant Hydrogenovibrio marinus [NiFe]-hydrogenase in Escherichia coli. J. Biotechnol. 2011, 155, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Liebgott, P.-P.; de Lacey, A.L.; Burlat, B.; Cournac, L.; Richaud, P.; Brugna, M.; Fernandez, V.M.; Guigliarelli, B.; Rousset, M.; Léger, C.; et al. Original Design of an Oxygen-Tolerant [NiFe] Hydrogenase: Major Effect of a Valine-to-Cysteine Mutation near the Active Site. J. Am. Chem. Soc. 2010, 133, 986–997. [Google Scholar] [CrossRef]
- Schlesier, J.; Rohde, M.; Gerhardt, S.; Einsle, O. A Conformational Switch Triggers Nitrogenase Protection from Oxygen Damage by Shethna Protein II (FeSII). J. Am. Chem. Soc. 2016, 138, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Varghese, F.; Kabasakal, B.V.; Cotton, C.A.R.; Schumacher, J.; Rutherford, A.W.; Fantuzzi, A.; Murray, J.W. A low-potential terminal oxidase associated with the iron-only nitrogenase from the nitrogen-fixing bacterium Azotobacter vinelandii. J. Biol. Chem. 2019, 294, 9367–9376. [Google Scholar] [CrossRef] [Green Version]
- Biz, A.; Mahadevan, R. Overcoming Challenges in Expressing Iron–Sulfur Enzymes in Yeast. Trends Biotechnol. 2021, 39, 665–677. [Google Scholar] [CrossRef]
- Wang, Y.; San, K.-Y.; Bennett, G.N. Cofactor engineering for advancing chemical biotechnology. Curr. Opin. Biotechnol. 2013, 24, 994–999. [Google Scholar] [CrossRef] [Green Version]
- Englund, E.; Shabestary, K.; Hudson, E.; Lindberg, P. Systematic overexpression study to find target enzymes enhancing production of terpenes in Synechocystis PCC 6803, using isoprene as a model compound. Metab. Eng. 2018, 49, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Agapakis, C.M.; Silver, P.A. Modular electron transfer circuits for synthetic biology. Bioeng. Bugs 2010, 1, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Rumpel, S.; Siebel, J.F.; Farès, C.; Duan, J.; Reijerse, E.; Happe, T.; Lubitz, W.; Winkler, M. Enhancing hydrogen production of microalgae by redirecting electrons from photosystem I to hydrogenase. Energy Environ. Sci. 2014, 7, 3296–3301. [Google Scholar] [CrossRef]
- Lamont, C.M.; Kelly, C.; Pinske, C.; Buchanan, G.; Palmer, T.; Sargent, F. Expanding the substrates for a bacterial hydrogenlyase reaction. Microbiology 2017, 163, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Agapakis, C.M.; Boyle, P.M.; Silver, P.A. Natural strategies for the spatial optimization of metabolism in synthetic biology. Nat. Chem. Biol. 2012, 8, 527–535. [Google Scholar] [CrossRef]
- Delebecque, C.J.; Lindner, A.B.; Silver, P.A.; Aldaye, F.A. Organization of Intracellular Reactions with Rationally Designed RNA Assemblies. Science 2011, 333, 470–474. [Google Scholar] [CrossRef]
- Lee, H.; DeLoache, W.C.; Dueber, J.E. Spatial organization of enzymes for metabolic engineering. Metab. Eng. 2012, 14, 242–251. [Google Scholar] [CrossRef]
- Carlsen, S.; Ajikumar, P.K.; Formenti, L.R.; Zhou, K.; Phon, T.H.; Nielsen, M.L.; Lantz, A.E.; Kielland-Brandt, M.C.; Stephanopoulos, G. Heterologous expression and characterization of bacterial 2-C-methyl-d-erythritol-4-phosphate pathway in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2013, 97, 5753–5769. [Google Scholar] [CrossRef] [Green Version]
- Benisch, F.; Boles, E. The bacterial Entner–Doudoroff pathway does not replace glycolysis in Saccharomyces cerevisiae due to the lack of activity of iron–sulfur cluster enzyme 6-phosphogluconate dehydratase. J. Biotechnol. 2014, 171, 45–55. [Google Scholar] [CrossRef]
- Wasserstrom, L.; Portugal-Nunes, D.; Almqvist, H.; Sandström, A.G.; Lidén, G.; Gorwa-Grauslund, M.F. Exploring d-xylose oxidation in Saccharomyces cerevisiae through the Weimberg pathway. AMB Express 2018, 8, 1–16. [Google Scholar] [CrossRef]
- Partow, S.; Siewers, V.; Daviet, L.; Schalk, M.; Nielsen, J. Reconstruction and Evaluation of the Synthetic Bacterial MEP Pathway in Saccharomyces cerevisiae. PLoS ONE 2012, 7, e52498. [Google Scholar] [CrossRef] [Green Version]
- Anthony, L.C.; Maggio-Hall, L.A.; Rothman, S.C.; Tomb, J.-F. Increased Heterologous Fe-S Enzyme Activity in Yeast. U.S. Patent US20,160,024,534A1, 28 January 2016. [Google Scholar]
- Salusjärvi, L.; Toivari, M.; Vehkomäki, M.-L.; Koivistoinen, O.; Mojzita, D.; Niemelä, K.; Penttilä, M.; Ruohonen, L. Production of ethylene glycol or glycolic acid from D-xylose in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 8151–8163. [Google Scholar] [CrossRef] [PubMed]
- Vicente, E.J.; Dean, D.R. Keeping the nitrogen-fixation dream alive. Proc. Natl. Acad. Sci. USA 2017, 114, 3009–3011. [Google Scholar] [CrossRef] [Green Version]
- López-Torrejón, G.; Jiménez-Vicente, E.; Buesa, J.M.; Hernandez, J.A.; Verma, H.K.; Rubio, L.M. Expression of a functional oxygen-labile nitrogenase component in the mitochondrial matrix of aerobically grown yeast. Nat. Commun. 2016, 7, 11426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Xie, X.; Yang, M.; Dixon, R.; Wang, Y. Modular electron-transport chains from eukaryotic organelles function to support nitrogenase activity. Proc. Natl. Acad. Sci. USA 2017, 114, E2460–E2465. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Payá-Tormo, L.; Coroian, D.; García-Rubio, I.; Castellanos-Rueda, R.; Eseverri, Á.; López-Torrejón, G.; Burén, S.; Rubio, L.M. Exploiting genetic diversity and gene synthesis to identify superior nitrogenase NifH protein variants to engineer N2-fixation in plants. Commun. Biol. 2021, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Schwizer, F.; Okamoto, Y.; Heinisch, T.; Gu, Y.; Pellizzoni, M.M.; Lebrun, V.; Reuter, R.; Köhler, V.; Lewis, J.C.; Ward, T.R. Artificial Metalloenzymes: Reaction Scope and Optimization Strategies. Chem. Rev. 2018, 118, 142–231. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Yu, Y.; Wang, J. Improving artificial metalloenzymes’ activity by optimizing electron transfer. Chem. Commun. 2017, 53, 4173–4186. [Google Scholar] [CrossRef]
- Artero, V.; Berggren, G.; Atta, M.; Caserta, G.; Roy, S.; Pecqueur, L.; Fontecave, M. From Enzyme Maturation to Synthetic Chemistry: The Case of Hydrogenases. Acc. Chem. Res. 2015, 48, 2380–2387. [Google Scholar] [CrossRef]
- Atkinson, J.T.; Campbell, I.; Thomas, E.E.; Bonitatibus, S.C.; Elliott, S.J.; Bennett, G.N.; Silberg, J.J. Metalloprotein switches that display chemical-dependent electron transfer in cells. Nat. Chem. Biol. 2019, 15, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Mutter, A.C.; Tyryshkin, A.M.; Campbell, I.; Poudel, S.; Bennett, G.N.; Silberg, J.J.; Nanda, V.; Falkowski, P.G. De novo design of symmetric ferredoxins that shuttle electrons in vivo. Proc. Natl. Acad. Sci. USA 2019, 116, 14557–14562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, R.J.; Margolis, P.S.; Trias, J.; Yuan, Z. Targeting metalloenzymes: A strategy that works. Curr. Opin. Pharmacol. 2003, 3, 502–507. [Google Scholar] [CrossRef]
Figure 1.
Structural diversity of metallocluster enzymes. (a) Crystal structures of diverse metallocluster enzymes bound to their metal cofactors. PDB entries and origin of each protein structures are listed in Appendix A Table A1. (b) Structure and environment of different metalloclusters from biotechnologically relevant enzymes.
Figure 1.
Structural diversity of metallocluster enzymes. (a) Crystal structures of diverse metallocluster enzymes bound to their metal cofactors. PDB entries and origin of each protein structures are listed in Appendix A Table A1. (b) Structure and environment of different metalloclusters from biotechnologically relevant enzymes.

Figure 2.
Strategies to enhance metallocluster enzyme maturation and stability. (a,b) A schematic representation of prokaryotic FeS maturation is presented as an example of the different steps and specific proteins involved in metallocluster enzyme maturation. The availability of metal cofactors can be improved by optimizing their biosynthesis or cellular import (a), or by optimizing essential pathways involved in post-translational modifications and protein maturation (b). (c) Reducing effects of oxidative stress.
Figure 2.
Strategies to enhance metallocluster enzyme maturation and stability. (a,b) A schematic representation of prokaryotic FeS maturation is presented as an example of the different steps and specific proteins involved in metallocluster enzyme maturation. The availability of metal cofactors can be improved by optimizing their biosynthesis or cellular import (a), or by optimizing essential pathways involved in post-translational modifications and protein maturation (b). (c) Reducing effects of oxidative stress.
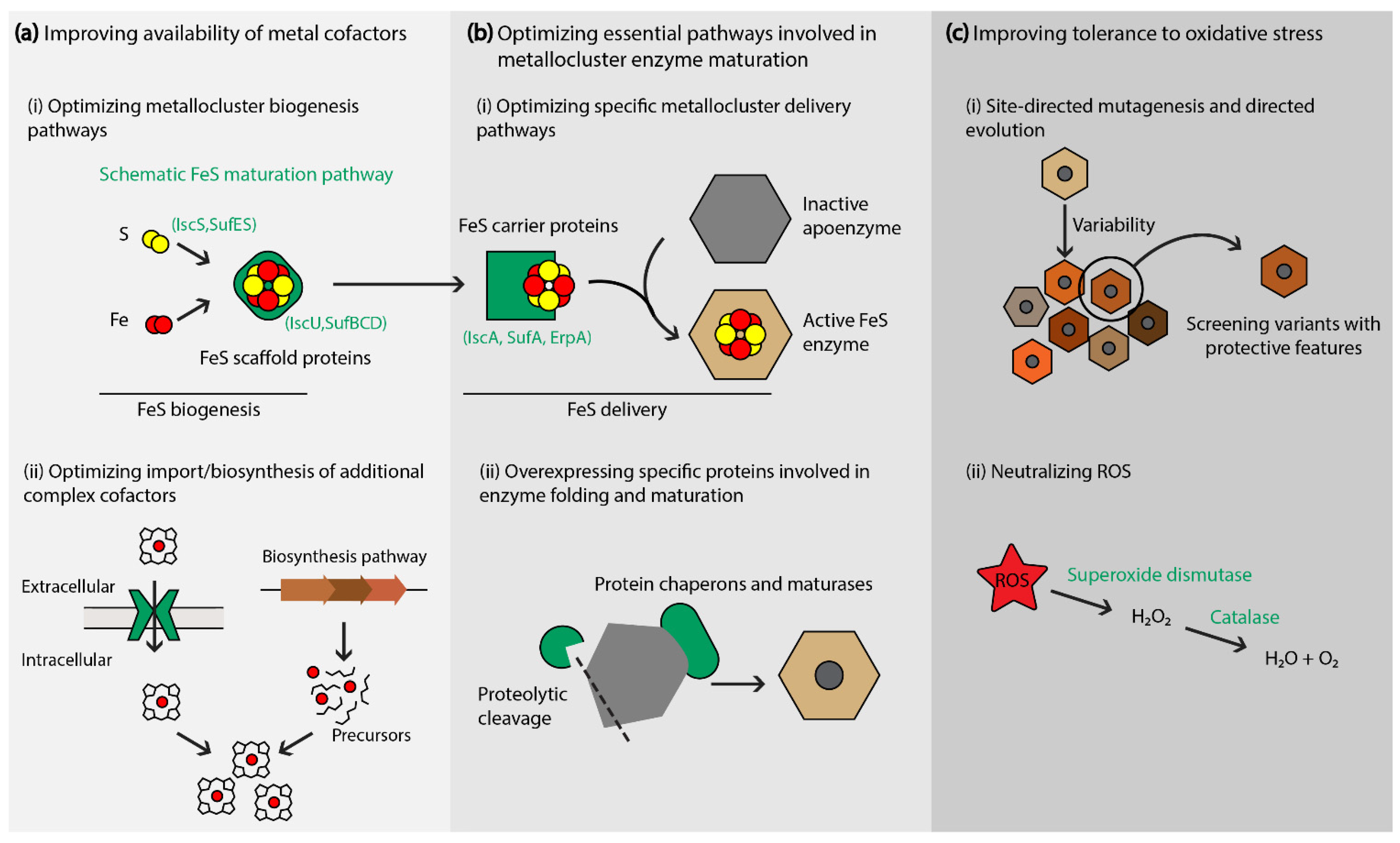
Figure 3.
Strategies to enhance electron supply to redox active metallocluster enzymes. (a) Schematic representation of the diversity of electron transfer pathways coupled to the activity of different metallocluster enzymes. Most enzymes are coupled to one or multiple ETPs that shuttle electrons from cellular cofactors or photosynthetic chains (such as PSI). Examples of natural redox partners involved in each pathway are given in parenthesis. (b) Increasing levels of natural or alternative reducing partners is a successful strategy to enhance electron supply to a given metallocluster enzyme. (c) Strategies to insulate electron transfer pathways can be employed to optimize and direct electron flow.
Figure 3.
Strategies to enhance electron supply to redox active metallocluster enzymes. (a) Schematic representation of the diversity of electron transfer pathways coupled to the activity of different metallocluster enzymes. Most enzymes are coupled to one or multiple ETPs that shuttle electrons from cellular cofactors or photosynthetic chains (such as PSI). Examples of natural redox partners involved in each pathway are given in parenthesis. (b) Increasing levels of natural or alternative reducing partners is a successful strategy to enhance electron supply to a given metallocluster enzyme. (c) Strategies to insulate electron transfer pathways can be employed to optimize and direct electron flow.
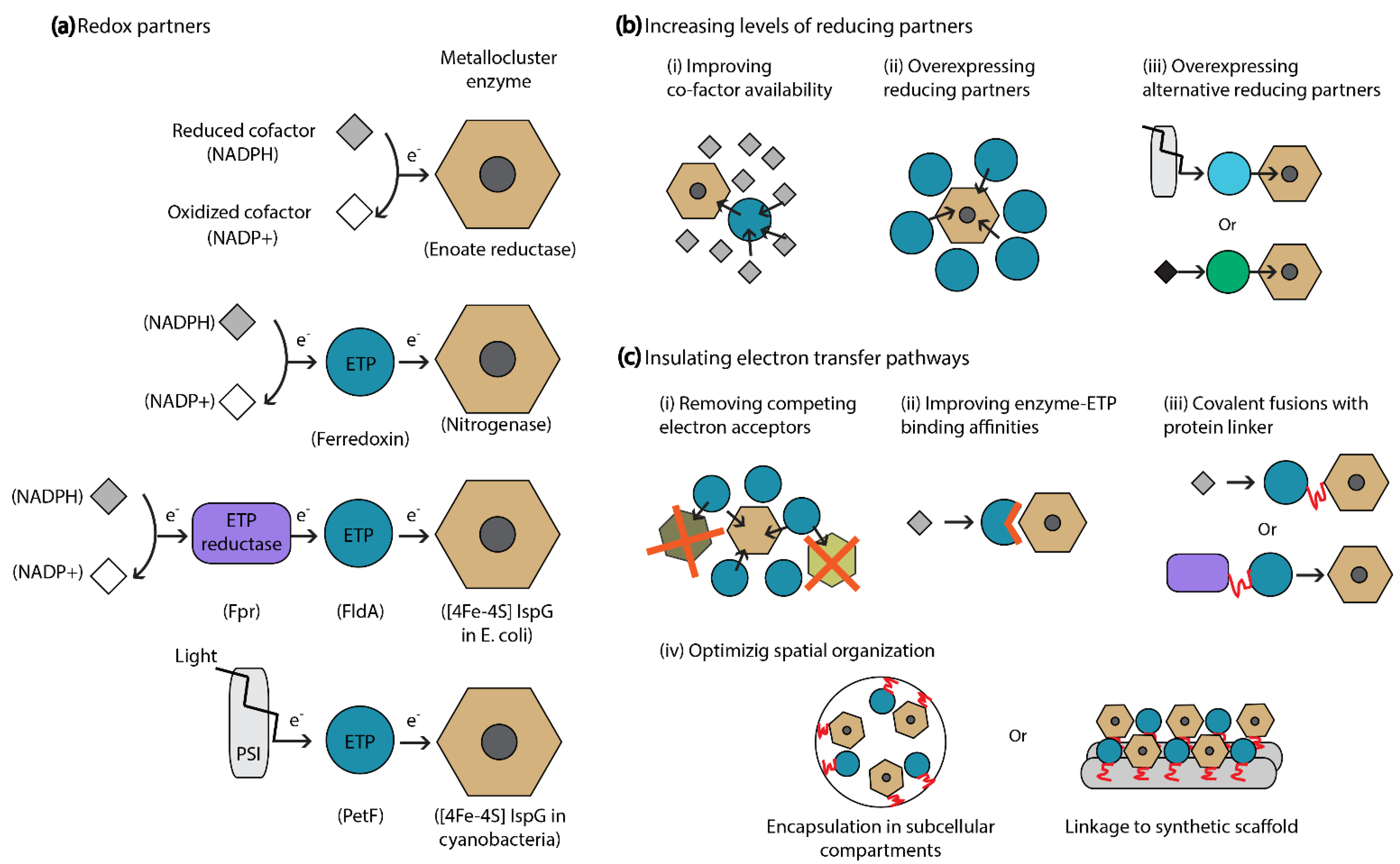
Table 1.
Biotechnological relevance of metalloenzymes.
| Name/Type | Metal Cluster Composition | Compound or Compound Family | Native Pathway | Biotechnological Relevance | Ref. |
|---|---|---|---|---|---|
| Nitrogenase | Complex FeS cluster [X–7Fe–C-9S] where “X” is molybdenum, vanadium, or iron | Reduction of N2 to bioavailable NH3 | Nitrogen fixation | Agriculture; plant-associated nitrogen-fixing bacteria | [11] |
| D-xylonate dehydratase | [2Fe-2S] cluster | Xylose catabolism, 1,2,4 butanetriol production | Weimberg pathway | Renewable carbon sources | [12] |
| IspG, IspH | [4Fe-4S] cluster | Isoprenoid precursors | MEP pathway | Pharmaceuticals, fragances, flavors, biofuels, polymers | [13] |
| IlvD | [4Fe-4S] cluster | Isobutyraldehyde isobutanol | Isoleucine and valine pathway | Biofuels | [14] |
| BioB | B12-Radical SAM [2Fe-2S] and [4Fe-4S] clusters | Biotin | Biotin biosynthesis | Food supplements, pharmaceuticals, cosmetics, animal feed | [15] |
| ThnK, ThnL, ThnP | B12-Radical SAM [4Fe-4S] clusters | Thienamycin and carbapenem derivatives | Thienamycin pathway | Antibiotics | [16] |
| PoyB, PoyC, PoyD | Radical SAM [4Fe-4S] clusters | Polytheonamide cytotoxins | Polytheonamide biosynthesis | Antibiotics | [17] |
| Fom3 | B12-Radical SAM [4Fe-4S] cluster | Fosfomycin | Fosfomycin biosynthesis | Antibiotics | [18] |
| GenK, GenD1 | B12-Radical SAM [4Fe-4S] cluster | Gentamicin | Gentamicin biosynthesis | Antibiotics | [19] |
| YtkT (radical SAM/FeS) | Radical SAM [4Fe-4S] cluster | Yatakemycin | Yatakemycin biosynthesis | Antitumor | [20] |
| Viperin | Radical SAM [4Fe-4S] cluster | Antiviral ribonucleotides | Antiviral defense | Antivirals | [21] |
| Cob enzymes | Contain [4Fe-4S] clusters | Vitamin B12 | Cobalamin biosynthesis | Medical and food industries | [22] |
| Enoate reductases | [4Fe-4S] cluster | Adipic acid, precursor for Nylon-6,6 polymer | Phenylalanine pathway | Commodity chemicals | [23] |
| [2Fe-2S] Rieske-type oxygenases | Iron-sulfur Rieske domain and non-heme Fe(II)-binding motif | Hapalindole-type products | Hapalindole biosynthesis | Antimycotic insecticidal | [24] |
| Hydrogenases | [FeFe]- or [NiFe] active site. Contain multiple Fe-S subclusters | Hydrogen (H2) gas | Hydrogen production | Biofuels | [25,26] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shomar, H.; Bokinsky, G. Towards a Synthetic Biology Toolset for Metallocluster Enzymes in Biosynthetic Pathways: What We Know and What We Need. Molecules 2021, 26, 6930. https://doi.org/10.3390/molecules26226930
AMA Style
Shomar H, Bokinsky G. Towards a Synthetic Biology Toolset for Metallocluster Enzymes in Biosynthetic Pathways: What We Know and What We Need. Molecules. 2021; 26(22):6930. https://doi.org/10.3390/molecules26226930
Chicago/Turabian StyleShomar, Helena, and Gregory Bokinsky. 2021. "Towards a Synthetic Biology Toolset for Metallocluster Enzymes in Biosynthetic Pathways: What We Know and What We Need" Molecules 26, no. 22: 6930. https://doi.org/10.3390/molecules26226930