
Bioactive Components in Oat and Barley Grain as a Promising Breeding Trend for Functional Food Production

 ,
,  ,
,
Abstract
1. Introduction
2. Bioactive Components in Oat and Barley Grain Used in Functional Nutrition
3. Regulatory and Structural Genes for the Biosynthesis of Anthocyanins in Barley and β-Glucans in Oats
3.1. Regulatory and Structural Genes for Anthocyanin Biosynthesis in Hordeum Vulgare
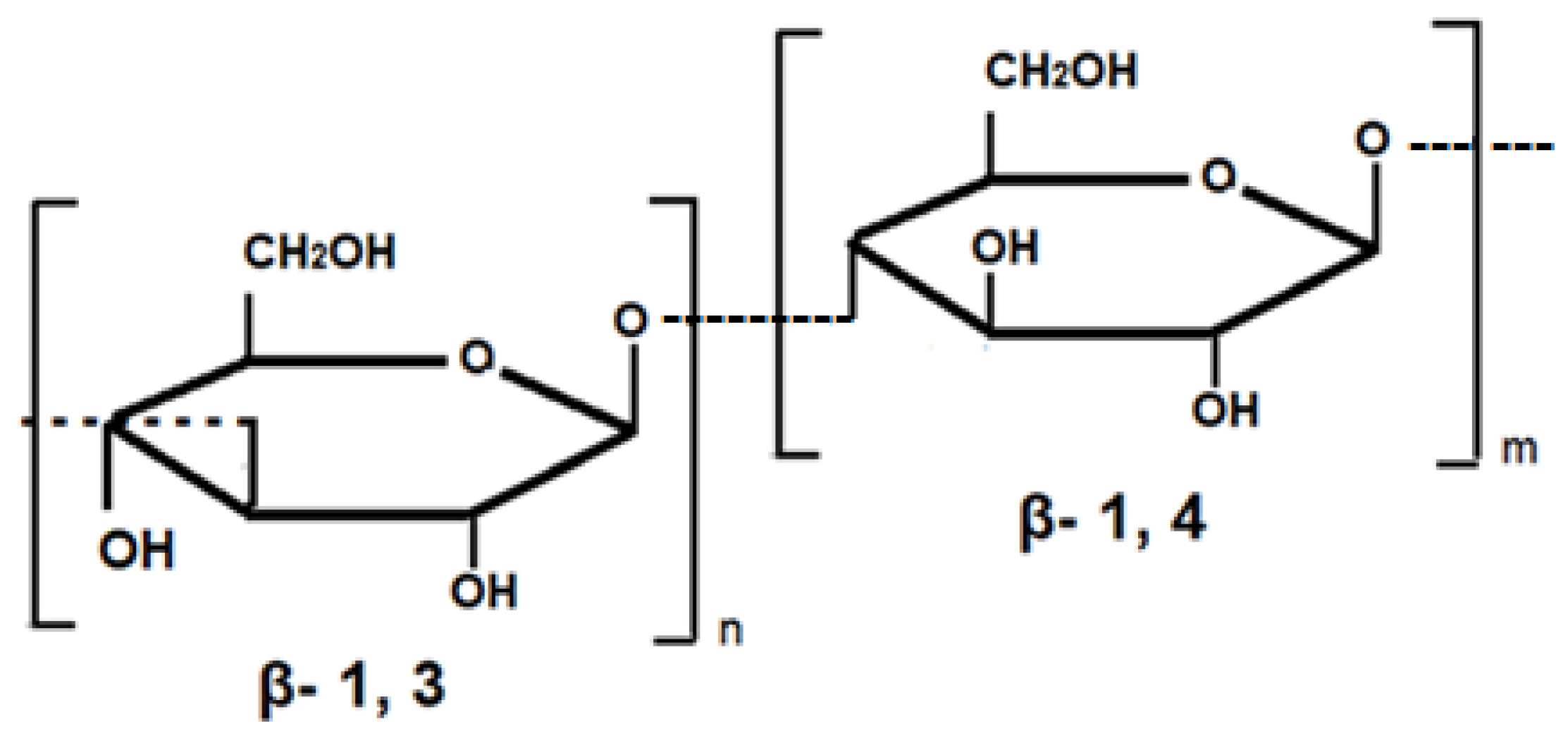
3.2. Genes for the Biosynthesis of β-Glucans in Barley and Oats
4. Study of Oat and Barley Accessions from the VIR Global Collection for the Content of Bioactive Components in Grain
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nikberg, I.I. Functional products in the structure of modern nutrition. Int. J. Endocrinol. 2011, 6, 64–69. (In Russian) [Google Scholar]
- Functional Foods Market Size, Share & Trends Analysis Report by Ingredient (Carotenoids, Prebiotics & Probiotics, Fatty Acids, Dietary Fibers), by Product, by Application, and Segment Forecasts, 2019–2025. Available online: https://www.researchandmarkets.com/reports/4764576/functional-foods-market-size-share-and-trends (accessed on 28 January 2021).
- Kornen, N.N.; Viktorova, E.P.; Evdokimova, O.V. Methodological approaches to the creation of healthy food. Nutr. Issues 2015, 84, 95–99. (In Russian) [Google Scholar]
- Cheryl, G.F.; Sachin, K.S.; Arya, S.S. Cereal based functional beverages: A review. Microbiol. Biotechnol. Food Sci. Fernandesetal 2018, 8, 914–919. [Google Scholar] [CrossRef]
- Aslam, H.; Green, J.; Jacka, F.N.; Collier, F.; Berk, M.; Pasco, J.; Samantha, L.D. Fermented foods, the gut and mental health: A mechanistic overview with implications for depression and anxiety. Nutr. Neurosci. 2020, 23, 659–671. [Google Scholar] [CrossRef]
- Popov, V.S.; Grigoriev, S.V.; Illarionova, K.V.; Shelenga, T.V. Fatty acid composition of hemp and cotton oils and the prospects of their use in the food industry and functional nutrition. Agrar. Russ. 2019, 8, 9–15. (In Russian) [Google Scholar] [CrossRef]
- Dydykin, A.S.; Ustinova, A.V.; Derevitskaya, O.K.; Aslanova, M.A.; Volovik, E.L. Functional meat and vegetable products using enriched vegetables. Food Ind. 2011, 8, 26–27. (In Russian) [Google Scholar]
- Popov, V.S.; Sergeeva, S.S.; Barsukova, N.V. Functional and technological properties of oat grain and a promising range of food products based on it. Bull. Technol. Univ. 2016, 16, 147–152. (In Russian) [Google Scholar]
- Koshevoy, O.S.; Fudina, E.V. Food security is the basis for ensuring economic security. News High. Educ. Inst. Volga Reg. 2015, 4, 188–196. (In Russian) [Google Scholar]
- Kayshev, V.G.; Seregin, S.N. Functional food products: The basis for disease prevention, health promotion and active longevity. Food Ind. 2017, 7, 8–14. (In Russian) [Google Scholar]
- Matveeva, T.V.; Koryachkina, S.Y. Flour Confectionery for Functional Purposes. Scientific Bases, Technologies, Recipes; FGOU VPO “State University—Educational, Research and Production Complex”: Orel, Russia, 2011; p. 358. ISBN 978-5-93932-312-3. (In Russian) [Google Scholar]
- Marklinder, I.; Johansson, L.; Haglund, A.; Nagel-Held, B.; Seibel, W. Effects of Flour from Different Barley Varieties on Barley Sour Dough Bread. Food Qual. Prefer. 1996, 7, 275–284. [Google Scholar] [CrossRef]
- Rico, D.; Peñas, E.; García, M.C.; Martínez-Villaluenga, C.; Rai, D.K.; Birsan, R.I.; Frias, J.; Martín-Diana, A.B. Sprouted barley flour as a nutritious and functional ingredient. Foods 2020, 9, 296. [Google Scholar] [CrossRef]
- Valevskaya, L.; Dzyuba, N.; Bunyak, E.; Evdokimova, G. The meaning of grain cultures in healthy food. Sci. Eur. 2017, 18, 71–73. [Google Scholar]
- Konuklar, G.; Inglett, G.E.; Warner, K.; Carriere, C.J. Use of a β-glucan hydrocolloidal suspension in the manufacture of low-fat Cheddar cheeses: Textural properties by instrumental methods and sensory panels. Food Hydrocoll. 2004, 18, 535–545. [Google Scholar] [CrossRef]
- Khomyakova, N.V. Oat bread—A concentrate of benefits and taste. Rus Bakery. 2007, 2, 36. [Google Scholar]
- Gormley, T.R.; Morrissey, A. A note on the evaluation of wheaten breads containing oat flour or oat flakes. Ir. J. Agric. Food Res. 1993, 32, 205–209. [Google Scholar]
- Saarela, M.; Virkajärvi, I.; Nohynek, L.; Vaari, A.; Mättö, J. Fibres as carriers for Lactobacillus rhamnosus during freeze-drying and storage in apple juice and chocolatecoated breakfast cereals. Int. J. Food Microbiol. 2006, 112, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Angelov, A.; Gotcheva, V.; Kuncheva, R.; Hristozova, T. Development of a new oat-based probiotic drink. Int. J. Food Microbiol. 2006, 112, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Selezneva, I.S. Influence of β-glucan from oats on the properties of low-fat yogurt. Sci. J. NRU ITMO Ser. Process. Appar. Food Prod. 2019, 4, 111–116. [Google Scholar]
- Sidhu, J.S.; Kabir, Y.; Huffman, F.G. Functional foods from cereal grains. Int. J. Food Prop. 2007, 10, 231–244. [Google Scholar] [CrossRef]
- Gorash, A.; Armoniene, R.; Fetch, J.M.; Liatukas, Z.; Danyte, V. Aspects in oat breeding: Nutrition quality, nakedness and disease resistance, challenges and perspectives: Review article. Ann. Appl. Biol. 2017, 171, 281–302. [Google Scholar] [CrossRef]
- Welch, R.W. Nutrient composition and nutritional quality of oats and comparisons with other cereals. In Oats: Chemistry and Technology, 2nd ed.; American Association of Cereal Chemists Inc.: St. Paul, MN, USA, 2011; pp. 95–107. ISBN 9780128104521. [Google Scholar]
- Shewry, P.R.; Piironen, V.; Lampi, A.M.; Nyström, L.; Li, L.; Rakszegi, M.; Fraś, A.; Boros, D.; Gebruers, K.; Courtin, C.M.; et al. Phytochemical and fiber components in oat varieties in the health grain diversity screen. J. Agric. Food Chem. 2008, 56, 9777–9784. [Google Scholar] [CrossRef] [PubMed]
- Loskutov, I.G.; Kovaleva, O.N.; Blinova, E.V. Genetic diversity barley and oats of N.I. Vavilov All-Russian Research Institute of Plant Industry collections for breeding. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2012, 66, 20–30. [Google Scholar] [CrossRef]
- Loskutov, I.G.; Rines, H.W. Avena L. In Wild Crop Relatives: Genomic and Breeding Resources, Cereals; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany; Institute of Nutraceutical Research, Clemson University: Clemson, SC, USA, 2011; pp. 109–183. ISBN 978-3-642-14228-4. [Google Scholar]
- Lapveteläinen, A.; Aro, T. Protein composition and functionality of high-protein oat four derived from integrated starch ethanol process. Cereal Chem. 1994, 71, 133–139. [Google Scholar]
- Bityutsky, N.P.; Loskutov, I.G.; Yakkonen, K.L.; Konarev, A.V.; Shelenga, T.V.; Khoreva, V.I.; Blinova, E.; Ryumin, A. Screening of Avena Sativa cultivars for iron, zinc, manganese, protein and oil content and fatty acid composition in whole grains. Cereal Res. Commun. 2020, 48, 87–94. [Google Scholar] [CrossRef]
- Zhoua, M.; Robards, K.; Glennie-Holmes, M.; Helliwell, S. Oat Lipids. J. Am. Oil Chem. Soc. 1999, 76, 159–169. [Google Scholar] [CrossRef]
- Loskutov, I.G. Oats (Avena L.)—Distribution, Taxonomy, Evolution and Breeding Value; VIR: Saint-Petersburg, Russia, 2007; p. 336. (In Russian) [Google Scholar]
- Batalova, G.A.; Krasilnikov, V.N.; Popov, V.S.; Safonova, E.E. Characteristics of the fatty acid composition of naked oats of Russian selection. In Proceedings of the IOP Conference Series: Earth and Environmental Science, St. Petersburg, Russia, 13–14 December 2018; IOP Publishing Ltd.: Bristol, UK, 2019; Volume 337, pp. 1–5. [Google Scholar]
- Krasilnikov, V.N.; Batalova, G.A.; Popov, V.S.; Sergeeva, S.S. Fatty acid composition of lipids in naked oat grain of domestic varieties. Russ. Agric. Sci. 2018, 44, 406–408. [Google Scholar] [CrossRef]
- Andreev, N.R.; Batalova, G.A.; Nosovskaya, L.P.; Adikaeva, L.V.; Goldstein, V.G.; Shevchenko, S.N. Evaluation of the technological properties of some cultivars of naked oats as a raw material for the production of starch. Legum. Cereal Crop. 2016, 1, 83–89. (In Russian) [Google Scholar]
- Leonova, S.; Shelenga, T.; Hamberg, M.; Konarev, A.V.; Loskutov, I.G.; Carlsson, A.S. Analysis of oil composition in cultivars and wild species of oat (Avena sp.). J. Agric. Food Chem. 2008, 56, 7983–7991. [Google Scholar] [CrossRef]
- Fincher, G.B.; Stone, B.A. Cell walls and their components in cereal grain technology. In Advances in Cereal Science and Technology; Pomeranz, Y., Ed.; American Association of Cereal Chemists Inc.: St. Paul, MN, USA, 1986; pp. 207–295. ISBN 978-0913250396. [Google Scholar]
- Krasilnikov, V.N.; Gavrilyuk, I.P.; Batalova, G.A.; Afonin, D.V.; Popov, V.S.; Sergeeva, S.S.; Loskutov, I.G.; Gubareva, N.K. Dietary fiber and avenins of grain of naked varieties of oats of new selection. Int. Sci. Res. J. 2017, 1, 111–116. (In Russian) [Google Scholar] [CrossRef]
- Wood, P.J.; Beer, M.U. Functional oat products. In Functional Foods. Biochemical & Processing Aspects, 1st ed.; Mazza, G., Ed.; Technomic Publishing Co.: Lancaster, PA, USA, 1998; Volume 1, pp. 1–37. ISBN 9780367400415. [Google Scholar]
- Wood, P.J.; Paton, D.; Siddiquil, R. Determination of beta-glucan in oats and barley. Cereal Chem. 1977, 54, 524–533. [Google Scholar]
- Khoury, D.E.; Cuda, C.; Luhovyy, B.L.; Anderson, G.H. Beta Glucan: Health Benefits in Obesity and Metabolic Syndrome. J. Nutr. Metab. 2012, 2012, 851362. [Google Scholar] [CrossRef]
- Popov, V.S.; Krasilnikov, V.N.; Barsukova, N.V. Oat β-glucans in functional and therapeutic nutrition. Probl. Econ. Manag. Trade Ind. 2014, 2, 78–83. (In Russian) [Google Scholar]
- Brownlee, I.A. The physiological roles of dietary fibre. Food Hydrocoll. 2011, 25, 238–250. [Google Scholar] [CrossRef]
- Chang, H.C.; Huang, C.N.; Yeh, D.M.; Wang, S.J.; Peng, C.H.; Wang, C.J. Oat prevents obesity and abdominal fat distribution, and improves liver function in humans. Plant Foods Hum. Nutr. 2013, 68, 18–23. [Google Scholar] [CrossRef]
- Harland, J. Authorised EU health claims for barley and oat beta-glucans. In Foods, Nutrients and Food Ingredients with Authorised EU Health Claims; Sadler, M.J., Ed.; Woodhead Publishing: Cambridge, UK, 2014; Volume 2, pp. 25–45. ISBN 978-0-85709-842-9. [Google Scholar]
- Wood, P.J. Oat and rye β-glucan: Properties and function. Cereal Chem. 2010, 87, 315–330. [Google Scholar] [CrossRef]
- Regand, A.; Chowdhury, Z.; Tosh, S.M.; Wolever, T.M.S.; Wood, P. The molecular weight, solubility and viscosity of oat beta-glucan affect human glycemic response by modifying starch digestibility. Food Chem. 2011, 129, 297–304. [Google Scholar] [CrossRef]
- Kumar, K.; Chauhan, D.; Kumar, S.; Sharma, M.; Kaur, R.; Vyas, P. Barley: A potential source of functional food ingredients. In Proceedings of the National Seminar on Technological Interventions in Food Processing and Preservation, Rajasthan, India, 17 November 2017; Amity University: Jaipur, India, 2017; pp. 85–86. [Google Scholar]
- Tiwari, P.K.; Sahu, R.K.; Sandey, K.K.; Tiwari, R.K. Importance of oats in human diet: A review. Bull. Environ. Pharmacol. Life Sci. 2017, 7, 125–130. [Google Scholar]
- Gao, C.; Gao, Z.; Greenway, F.L.; Burton, H.J.; Johnson, W.D.; Keenan, M.J.; Enright, F.M.; Martin, R.J.; Chu, Y.; Zheng, J. Oat consumption reduced intestinal fat deposition and improved health span in Caenorhabditis elegans model. Nutr. Res. 2015, 35, 834–843. [Google Scholar] [CrossRef]
- Thomas, M.; Kim, S.; Guo, W.; Collins, W.F.; Wise, M.L.; Meydani, M. High levels of avenanthramides in oat-based diet further suppress high fat diet-induced atherosclerosis in Ldlr−/− Mice. J. Agric. Food Chem. 2018, 66, 498–504. [Google Scholar] [CrossRef]
- Hurtado-Fernández, E.; Gómez-Romero, M.; Carrasco-Pancorbo, A.; Fernández-Gutiérrez, A. Application and potential of capillary electroseparation methods to determine antioxidant phenolic compounds from plant food material. J. Pharm. Biomed. Anal. 2010, 53, 1130–1160. [Google Scholar] [CrossRef]
- Zielinski, H.; Ciska, E.; Kozlowska, H. The cereal grains: Focus on vitamin E. Czech Food Sci. 2001, 19, 182–188. [Google Scholar] [CrossRef]
- Polonskiy, V.I.; Loskutov, I.G.; Sumina, A.V. Breeding for antioxidant content in grain as a promising trend in obtaining healthy food products. Russ. J. Genet. Appl. Res. 2018, 22, 343–352. [Google Scholar] [CrossRef]
- Makarenko, O.A.; Levitsky, A.P. Physiological functions of flavonoids in plants. Physiol. Biochem. Cultiv. Plants 2013, 45, 100–112. (In Russian) [Google Scholar]
- Adzhieva, V.F.; Babak, O.G.; Shoeva, O.J.; Kilchevsky, A.V.; Khlestkina, E.K. Molecular genetic mechanisms of the formation of the color of fruits and seeds of plants. Russ. J. Genet. Appl. Res. 2015, 19, 561–573. [Google Scholar] [CrossRef]
- Khlestkina, E.K. The adaptive role of flavonoids: Emphasis on cereals. Cereal Res. Commun. 2013, 41, 185–198. [Google Scholar] [CrossRef]
- Jende-Strid, B. Gene-enzyme relations in the pathway of flavonoid biosynthesis in barley. Theor. Appl. Genet. 1991, 81, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Meldgaard, M. Expression of chalcone synthase, dihydroflavonol reductase, and flavanone-3-hydroxylase in mutants of barley deficient in anthocyanin and proanthocyanidin biosynthesis. Theor. Appl. Genet. 1992, 83, 695–706. [Google Scholar] [CrossRef]
- Dubcovsky, J.; Luo, M.C.; Zhong, G.Y.; Bransteitter, R.; Desai, A.; Kilian, A.; Kleinhofs, A.; Dvorak, J. Genetic map of diploid wheat, Triticum monococcum L., and its comparison with maps of Hordeum vulgare L. Genetics 1996, 143, 983–999. [Google Scholar] [CrossRef]
- Strygina, K.V.; Börner, A.; Khlestkina, E.K. Identification and characterization of regulatory network components for anthocyanin synthesis in barley aleurone. BMC Plant Biol. 2017, 17, 184. [Google Scholar] [CrossRef]
- Shoeva, O.Y.; Strygina, K.V.; Khlestkina, E.K. Genes that control the synthesis of flavonoid and melanin pigments in barley. Russ. J. Genet. Appl. Res. 2018, 22, 333–342. [Google Scholar] [CrossRef]
- Vikhorev, A.V.; Strygina, K.V.; Khlestkina, E.K. Duplicated flavonoid 3′-hydroxylase and flavonoid 3′, 5′-hydroxylase genes in barley genome. PeerJ 2019, 7, e6266. [Google Scholar] [CrossRef]
- Khlestkina, E.K.; Shoeva, O.Y.; Gordeeva, E.I. Flavonoid biosynthesis genes in wheat. Russ. J. Genet. Appl. Res. 2014, 18, 784–796. [Google Scholar] [CrossRef]
- Shoeva, O.Y.; Khlestkina, E.K.; Berges, H.; Salina, E.A. The homeologous encoding chalcone-flavanone isomerase in Triticum aestivum L.: Structural characterization and expression in different parts of wheat plant. Gene 2014, 538, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Karre, S.; Kumar, A.; Yogendra, K.; Kage, U.; Kushalappa, A.; Charron, J.-B. HvWRKY23 regulates flavonoid glycoside and hydroxycinnamic acid amide biosynthetic genes in barley to combat Fusarium head blight. Plant Mol. Biol. 2019, 100, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Karre, S.; Kumar, A.; Dhokane, D.; Kushalappa, A.C. Metabolo-transcriptome profiling of barley reveals induction of chitin elicitor receptor kinase gene (HvCERK1) conferring resistance against Fusarium graminearum. Plant Mol. Biol. 2017, 93, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Druka, A.; Kudrna, D.; Rostoks, N.; Brueggeman, R.; Wettstein, D.; Kleinhofs, A. Chalcone isomerase gene from rice (Oryza sativa) and barley (Hordeum vulgare): Physical, genetic and mutation mapping. Gene 2003, 302, 171–178. [Google Scholar] [CrossRef]
- Kristiansen, K.N.; Rohde, W. Structure of the Hordeum vulgare gene encoding dihydroflavonol-4-reductase and molecular analysis of antl8 mutants blocked in flavonoid synthesis. Mol. Gen. Genet. 1991, 230, 49–59. [Google Scholar] [CrossRef]
- Christensen, A.B.; Gregersen, P.L.; Schröder, J.; Collinge, D.B. A chalcone synthase with an unusual substrate preference is expressed in barley leaves in response to UV light and pathogen attack. Plant Mol. Biol. 1998, 37, 849–857. [Google Scholar] [CrossRef]
- Pecchioni, N.; Vale, G.; Toubia-Rahme, H.; Faccioli, P.; Terzi, V.; Delogu, G.; Fischbeck, G. Barley-Pyrenophora graminea interaction: QTL analysis and gene mapping. Plant Breed. 1999, 118, 29–35. [Google Scholar] [CrossRef]
- Peukert, M.; Weise, S.; Röder, M.S.; Matthies, I.E. Development of SNP markers for genes of the phenylpropanoid pathway and their association to kernel and malting traits in barley. BMC Genet. 2013, 14, 97. [Google Scholar] [CrossRef]
- Khlestkina, E.K.; Salina, E.A.; Matthies, I.E.; Leonova, I.N.; Börner, A.; Röder, M.S. Comparative molecular marker-based genetic mapping of flavanone 3-hydroxylase genes in wheat, rye and barley. Euphytica 2011, 179, 333–341. [Google Scholar] [CrossRef]
- Cockram, J.; White, J.; Zuluaga, D.L.; Smith, D.; Comadran, J.; Macaulay, M.; Luo, Z.; Kearsey, M.J.; Werner, P.; Harrap, D.; et al. Genome wide association mapping to candidate polymorphism resolution in the unsequenced barley genome. Proc. Natl. Acad. Sci. USA 2010, 107, 21611–21616. [Google Scholar] [CrossRef]
- Himi, E.; Taketa, S. Isolation of candidate genes for the barley Ant1 and wheat Rc genes controlling anthocyanin pigmentation in different vegetative tissues. Mol. Genet. Genom. 2015, 290, 1287–1298. [Google Scholar] [CrossRef]
- Strygina, K.V.; Khlestkina, E.K. Structural and functional divergence of the Mpc1 genes in wheat and barley. BMC Evol. Biol. 2019, 19, 90–99. [Google Scholar] [CrossRef]
- Strygina, K.V.; Khlestkina, E.K. Structural and functional organization and evolution of the WD40 genes involved in the regulation of flavonoid biosynthesis in the Triticeae Tribe. Russ. J. Genet. 2019, 55, 1398–1405. [Google Scholar] [CrossRef]
- Doblin, M.S.; Pettolino, F.A.; Wilson, S.M.; Campbell, R.; Burton, R.A.; Fincher, G.B.; Newbigin, E.; Bacic, A. A barley cellulose synthase-like CSLH gene mediates (1,3;1,4)-β-d-glucan synthesis in transgenic Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 5996–6001. [Google Scholar] [CrossRef] [PubMed]
- Fincher, G.B. Exploring the evolution of (1,3;1,4)-β-d-glucans in plant cell walls: Comparative genomics can help! Curr. Opin. Plant Biol. 2009, 12, 140–147. [Google Scholar] [CrossRef]
- Buckeridge, M.S.; Rayon, C.; Urbanowicz, B.; Tiné, M.A.S.; Carpita, N.C. Mixed Linkage (1→3),(1→4)-β-d-Glucans of Grasses. Cereal Chem. J. 2004, 81, 115–127. [Google Scholar] [CrossRef]
- Delmer, D.P. Cellulose biosynthesis: Exciting times for a difficult field of study. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 245–276. [Google Scholar] [CrossRef] [PubMed]
- Delaney, B.; Nicolosi, R.J.; Wilson, T.A.; Carlson, T.; Frazer, S.; Zheng, G.H.; Hess, R.; Ostergren, K.; Haworth, J.; Knutson, N. B-Glucan fractions from barley and oats are similarly antiatherogenic in hypercholesterolemic Syrian Golden Hamsters. J. Nutr. 2003, 133, 468–475. [Google Scholar] [CrossRef]
- Li, J.; Burton, R.A.; Harvey, A.J.; Hrmova, M.; Wardak, A.Z.; Stone, B.A.; Fincher, G.B. Biochemical evidence linking a putative callose synthase gene with (1→3)-beta-d-glucan biosynthesis in barley. Plant Mol. Biol. 2003, 53, 213–225. [Google Scholar] [CrossRef]
- Richmond, T.A.; Somerville, C.R. The Cellulose Synthase Superfamily. Plant Physiol. 2000, 124, 495–498. [Google Scholar] [CrossRef]
- Farrokhi, N.; Burton, R.A.; Brownfield, L.; Hrmova, M.; Wilson, S.M.; Bacic, A.; Fincher, G.B. Plant cell wall biosynthesis: Genetic, biochemical and functional genomics approaches to the identification of key genes. Plant Biotechnol. J. 2006, 4, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Hazen, S.P.; Scott-Craig, J.S.; Walton, J.D. Cellulose synthase-like genes of rice. Plant Physiol. 2002, 128, 336–340. [Google Scholar] [CrossRef]
- Burton, R.A.; Collins, H.M.; Kibble, N.A.J.; Smith, J.A.; Shirley, N.J.; Jobling, S.A.; Henderson, M.; Singh, R.R.; Pettolino, F.; Wilson, S.M.; et al. Over-expression of specific HvCslF cellulose synthase-like genes in transgenic barley increases the levels of cell wall (1,3;1,4)-β-d-glucans and alters their fine structure. Plant Biotechnol. J. 2011, 9, 117–135. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, M.C.; Smith, S.M.; Sheridan, J.L.; Hu, G.; Islamovic, E.; Reid, R.; Jackson, E.W.; Maughan, P.J.; Ames, N.P.; Jellen, E.N.; et al. Identification of mixed linkage β-glucan quantitative trait loci and evaluation of AsCslF6 homoeologs in hexaploid oat. Crop Sci. 2020, 60, 914–933. [Google Scholar] [CrossRef]
- Burton, R.A.; Wilson, S.M.; Hrmova, M.; Harvey, A.J.; Shirley, N.J.; Medhurst, A.; Stone, B.A.; Newbigin, E.J.; Bacic, A.; Fincher, G.B. Cellulose synthase-like CslF genes mediate the synthesis of cell wall (1,3;1,4)-β-d-glucans. Science 2006, 311, 1940–1942. [Google Scholar] [CrossRef]
- Newell, M.A.; Asoro, F.G.; Scott, M.P.; White, P.J.; Beavis, W.D.; Jannink, J.-L. Genome-wide association study for oat (Avena sativa L.) beta-glucan concentration using germplasm of worldwide origin. Theor. Appl. Genet. 2012, 125, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Yanova, M.A.; Tsuglenok, G.I.; Ivanova, T.S. The use of naked forms of barley and oats in food production. Bull. Krasgau 2012, 4, 203–205. (In Russian) [Google Scholar]
- Nizova, G.K.; Yarosh, N.P. Influence of pre-sowing treatment of seeds with succinic acid on the quality of green mass and oat grain. Sci. Tech. Bull. N.I. Vavilov All-Russ. Res. Inst. Plant Ind. 1988, 184, 17. (In Russian) [Google Scholar]
- Ballabio, C.; Uberti, F.; Manferdelli, S.; Vacca, E.; Boggini, G.; Redaelli, R.; Catassie, C.; Lionetti, E.; Peñas, E.; Restani, P. Molecular characterisation of 36 oat varieties and in vitro assessment of their suitability for coeliacs’ diet. J. Cereal Sci. 2011, 54, 110–115. [Google Scholar] [CrossRef]
- Gavrilyuk, I.P.; Gubareva, N.K.; Perchuk, I.N.; Loskutov, I.G.; Konarev, A.V.; Oreshko, L.S. The avenins and celiac disease. In Proceedings of the 3rd International Symposium on Gluten-Free Cereal Products and Beverages, Vienna, Austria, 12–14 June 2013; p. 15. [Google Scholar]
- Konarev, A.V.; Shelenga, T.V.; Perchuk, I.N.; Blinova, E.V.; Loskutov, I.G. Characteristics of the diversity of oats (Avena L.) from the collection of VIR—The initial material for breeding for resistance to fusarium. Agrar. Russ. 2015, 5, 1–10. (In Russian) [Google Scholar] [CrossRef]
- Leonova, S.; Gnutikov, A.; Loskutov, I.; Blinova, E.; Gustafsson, K.E.; Olsson, O. Diversity of avenanthramide content in cultivated and wild oats. Proc. Appl. Bot. Genet. Breed. 2020, 181, 30–47. (In Russian) [Google Scholar] [CrossRef]
- Bityutskii, N.; Yakkonen, K.; Loskutov, I. Content of iron, zinc and manganese in grains of Triticum aestivum, Secale cereale, Hordeum vulgare and Avena sativa cultivars registered in Russia. Genet. Resour. Crop Evol. 2017, 64, 1955–1961. [Google Scholar] [CrossRef]
- Loskutov, I.G.; Shelenga, T.V.; Konarev, A.V.; Vargach, Y.I.; Porokhovinova, E.A.; Blinova, E.V.; Gnutikov, A.A.; Rodionov, A.V. Biochemical approach of structuring the variety diversity of the naked and covered forms of cultural oats (Avena Sativa L.). Ecol. Genet. 2020, 18, 27–41. (In Russian) [Google Scholar] [CrossRef]
- Loskutov, I.G.; Shelenga, T.V.; Konarev, A.V.; Shavarda, A.L.; Blinova, E.V.; Dzubenko, N.I. The metabolomic approach to the comparative analysis of wild and cultivated species of oats (Avena L.). Russ. J. Genet. Appl. Res. 2016, 20, 636–642. [Google Scholar] [CrossRef]
- Zhang, G.P.; Wang, J.M.; Chen, J.X. Analysis of β-glucan content in barley cultivars from different locations of China. Food Chem. 2002, 79, 251–254. [Google Scholar] [CrossRef]
- Fastnaught, C.E.; Berglund, P.T.; Holm, E.T.; Fox, G.J. Genetic and environmental variation in β-glucan content and quality parameters of barley for food. Crop Sci. 1996, 36, 941–946. [Google Scholar] [CrossRef]
- Yalcin, E.; Celik, S.; Akar, T.; Sayim, I.; Koksel, H. Effects of genotype and environment on β-glucan and dietary fibre contents of hull-less barleys grown in Turkey. Food Chem. 2007, 101, 171–176. [Google Scholar] [CrossRef]
- Lee, C.J.; Horsley, R.D.; Manthey, F.A.; Schwarz, P.B. Comparison of β-glucan content of barley and oat. Cereal Chem. 1997, 74, 571–575. [Google Scholar] [CrossRef]
- Hang, A.; Obert, D.; Gironella, A.I.N.; Burton, C.S. Barley amylase and β-glucan: Their relationships to protein, agronomic traits, and environmental factors. Crop Sci. 2007, 47, 1754–1760. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, J.; Wang, J.; Ding, S. Cultivar and environmental effects on (1-3, 1-4)-β-glucan and protein content in malting barley. J. Cereal Sci. 2001, 34, 295–301. [Google Scholar] [CrossRef]
- Rey, J.I.; Hayes, P.M.; Petrie, S.E.; Corey, A.; Flowers, M.; Ohm, J.B.; Ong, C.; Rhinhart, K.; Ross, A.S. Production of dryland barley for human food: Quality and agronomic performance. Crop Sci. 2009, 49, 347–355. [Google Scholar] [CrossRef]
- Aman, P.; Graham, H.; Tilley, A.-C. Content and solubility of mixed-linked (1-3;1-4)-β-glucan in barley and oats during kernel development and storage. J. Cereal Sci. 1989, 10, 45–50. [Google Scholar] [CrossRef]
- Griffey, C.; Brooks, W.; Kurantz, M.; Thomason, W.; Taylor, F.; Obert, D.; Morea, R.; Flores, R.; Sohn, M.; Hicks, K. Grain composition of Virginia winter barley and implications for use in feed, food, and biofuels production. J. Cereal Sci. 2010, 51, 41–49. [Google Scholar] [CrossRef]
- Knutsen, S.H.; Holtekjilen, A.K. Preparation and analysis of dietary fiber constituents in whole grain from hulled and hull-less barley. Food Chem. 2007, 102, 707–715. [Google Scholar] [CrossRef]
- Huth, M.; Dongowski, G.; Gebhart, E.; Flamme, W. Functional properties of dietary fibre enriched exudates from barley. J. Cereal Sci. 2002, 32, 115–117. [Google Scholar] [CrossRef]
- Konarev, A.V.; Loskutov, I.G.; Shelenga, T.V.; Khoreva, V.I.; Konarev, A.V. Plant genetic resources—An inexhaustible source of healthy food. Agrar. Russ. 2019, 2, 38–48. (In Russian) [Google Scholar] [CrossRef]
- Lukyanova, M.V.; Trofimovskaya, A.Y.; Gudkova, G.N.; Terentyeva, I.A.; Yarosh, N.P. Cultural Flora of the USSR: Barley; Agropromizdat: Leningrad, Russia, 1990; Volume 3, p. 421. (In Russian) [Google Scholar]
- Gagkaeva, T.Y.; Gavrilova, O.P.; Levitin, M.M.; Novozhilov, K.V. Fusariosis of grain crops. Suppl. J. Plant Prot. Q. 2011, 5, 69–120. (In Russian) [Google Scholar]
- Gagkaeva, T.Y.; Levitin, M.M.; Zuev, E.V.; Terentjeva, I.A. Evaluation of genetic resources of wheat and barley from Far East of Russia for resistance to Fusarium head blight. J. Appl. Genet. 2002, 43A, 229–236. [Google Scholar]
- Gagkaeva, T.Y.; Gavrilova, O.P. Fusarium head and grain of barley. Work. Appl. Bot. Breed. 2009, 165, 39–44. (In Russian) [Google Scholar]
- Mesterhasy, A. Types and components resistance to Fusarium head blight of wheat. Plant Breed. 1995, 114, 377–386. [Google Scholar] [CrossRef]
- Miller, J.D.; Young, J.C.; Sampson, R.D. Deoxynivalenol and Fusarium head blight resistance in spring cereals. J. Phytopathol. 1985, 113, 359–367. [Google Scholar] [CrossRef]
- Martin, C.; Schöneberg, T.; Vogelgsang, S.; Morisoli, R.; Bertossa, M.; Mauch-Mani, B.; Mascher, F. Resistance against Fusarium graminearum and the relationship to β-glucan content in barley grains. Eur. J. Plant Pathol. 2018, 152, 621–634. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Wild/ Cultivated | Oil Content | Pl | 1,2-DAG | 1,3-DAG | FFA | Unknown Lipids | TAG1 | TAG2 | TAG |
|---|---|---|---|---|---|---|---|---|---|
| Wild | 7.8 ± 0.2 | 15.4 ± 0.6 | 1.3 ± 0.1 | 1.9 ± 0.1 | 1.5 ± 0.2 | 1.0 ± 0.1 | 1.1 ± 0.1 | 0.9 ± 0.1 | 77.1 ± 0.9 |
| Cultivated | 5.9 ± 0.2 | 16.0 ± 0.7 | 1.1 ± 0.1 | 2.3 ± 0.1 | 2.5 ± 0.1 | 1.4 ± 0.1 | 1.4 ± 0.1 | 1.1 ± 0.0 | 74.2 ± 1.0 |
| F value | 47.5 *** | 0.4 NS | 1.3 NS | 10.6 ** | 10.2 ** | 4.2 * | 13.2 *** | 11.0 ** | 4.3 * |
| VIR Catalogue No. | Name of Cultivar | Origin | Content of Avenanthramides, mg/kg | |||
|---|---|---|---|---|---|---|
| I | II | III | Average | |||
| 14787 | Privet | RF, Moscow reg. | 23.46 | 36.92 | 30.39 | 30.26 |
| 15277 | Bulanyi | RF, Moscow reg. | 8.28 | 13.06 | 7.31 | 9.55 |
| 15187 | Eklips | RF, Kirov reg. | 92.60 | 121.67 | 144.32 | 119.53 |
| 14648 | Argamak | RF, Kirov reg. | 5.38 | 6.60 | 6.11 | 6.03 |
| 14857 | Krechet | RF, Kirov reg. | 20.27 | 19.91 | 29.90 | 23.36 |
| 15068 | Konkur | RF, Ul’yanovsk reg. | 54.32 | 53.76 | 50.49 | 52.86 |
| 14960 | Vyatskii * | RF, Kirov reg. | 214.10 | 169.50 | 261.20 | 214.93 |
| 15275 | Persheron * | RF, Kirov reg. | 62.35 | 68.82 | 54.41 | 61.86 |
| 15067 | Golets * | RF, Krasnoyask reg. | 77.62 | 82.14 | 79.24 | 79.67 |
| 15067 | Levsha * | RF, Kemerovo reg. | 59.81 | 72.77 | 67.65 | 66.74 |
| 15115 | Aldan * | RF, Kemerovo reg. | 60.54 | 83.74 | 56.44 | 66.91 |
| 15116 | Murom * | RF, Kemerovo reg. | 138.98 | 170.54 | 200.71 | 170.08 |
| 15117 | Pomor * | RF, Kemerovo reg. | 43.40 | 46.82 | 46.00 | 45.40 |
| 15183 | Taidon * | RF, Kemerovo reg. | 140.20 | 165.76 | 122.96 | 142.97 |
| 14851 | Numbat * | Australia | 358.87 | 460.00 | 403.78 | 407.55 |
| VIR Catalogue No. | Name of Cultivar | Origin | Content of Tocopherols, % | Content of Sterols, % |
|---|---|---|---|---|
| 5184 | Local | Spain | 283 | 1.18 |
| 11840 | Borrus | Germany | 184 | 1.00 |
| 14648 | Argamak | RF, Kirov reg. | 189 | 0.64 |
| 13780 | Skakun | RF, Moscow reg. | 180 | 0.64 |
| 13918 | Kirovets | RF, Kirov reg. | 227 | 0.72 |
| 13957 | Gunter | RF, Kirov reg. | 236 | 0.67 |
| 14373 | Fakir | RF, Kirov reg. | 235 | 0.81 |
| 14781 | Faust | RF, Kirov reg. | 195 | 0.77 |
| 14857 | Krechet | RF, Kirov reg. | 149 | 0.61 |
| 15177 | Derbi | RF, Ulaynovsk reg. | 169 | 0.62 |
| 15180 | Piruet | RF, Ulaynovsk reg. | 167 | 0.64 |
| 1931 | Local * | China | 223 | 0.74 |
| 2472 | Local * | Mongolia | 415 | 0.97 |
| 8317 | Local * | China | 106 | 0.85 |
| Name/Genomes | Wild Oats | Cultivated Oats | |||||
|---|---|---|---|---|---|---|---|
| C | A | AB | AC | CD | ACD | ACD | |
| Amino acids | 61.3 ± 0.03 | 67.5 ± 0.05 | 45.8 ± 0.02 | 19.3 ± 0.01 | 41.1 ± 0.02 | 30.4 ± 0.02 | 75.80 ± 0.04 |
| Fatty acids | 1058.8 ± 0.11 | 603.6 ± 0.08 | 1040.6 ± 0.10 | 412.0 ± 0.04 | 656.1 ± 0.07 | 981.5 ± 0.10 | 494.00 ± 0.10 |
| Sterols | 7.4 ± 0.00 | 13.6 ± 0.01 | 25.0 ± 0.01 | 13.5 ± 0.01 | 25.7 ± 0.01 | 26.4 ± 0.02 | 16.40 ± 0.01 |
| Organic acids | 99.6 ± 0.05 | 126.5 ± 0.08 | 167.4 ± 0.08 | 37.1 ± 0.02 | 115.7 ± 0.06 | 108.8 ± 0.07 | 49.90 ± 0.02 |
| Polyhydric alcohols | 370.3 ± 0.11 | 343.2 ± 0.10 | 342.4 ± 0.10 | 91.7 ± 0.03 | 171.3 ± 0.05 | 312.1 ± 0.09 | 189.90 ± 0.09 |
| Monosaccharides | 1194.6 ± 0.08 | 1159.1 ± 0.08 | 1429.5 ± 0.07 | 358.3 ± 0.02 | 329.8 ± 0.04 | 1217.4 ± 0.08 | 901.50 ± 0.09 |
| Disaccharides | 6943.7 ± 0.14 | 2243.1 ± 0.05 | 1588.0 ± 0.03 | 4979.1 ± 0.10 | 7424.2 ± 0.15 | 1448.4 ± 0.03 | 2361.40 ± 0.09 |
| Total sugars | 8138.3 ± 0.10 | 3402.2 ± 0.05 | 3017.5 ± 0.04 | 5337.4 ± 0.06 | 7754.1 ± 0.09 | 2665.8 ± 0.04 | 3262.90 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shvachko, N.A.; Loskutov, I.G.; Semilet, T.V.; Popov, V.S.; Kovaleva, O.N.; Konarev, A.V. Bioactive Components in Oat and Barley Grain as a Promising Breeding Trend for Functional Food Production. Molecules 2021, 26, 2260. https://doi.org/10.3390/molecules26082260
Shvachko NA, Loskutov IG, Semilet TV, Popov VS, Kovaleva ON, Konarev AV. Bioactive Components in Oat and Barley Grain as a Promising Breeding Trend for Functional Food Production. Molecules. 2021; 26(8):2260. https://doi.org/10.3390/molecules26082260
Chicago/Turabian StyleShvachko, Natalia A., Igor G. Loskutov, Tatyana V. Semilet, Vitaliy S. Popov, Olga N. Kovaleva, and Alexei V. Konarev. 2021. "Bioactive Components in Oat and Barley Grain as a Promising Breeding Trend for Functional Food Production" Molecules 26, no. 8: 2260. https://doi.org/10.3390/molecules26082260
APA StyleShvachko, N. A., Loskutov, I. G., Semilet, T. V., Popov, V. S., Kovaleva, O. N., & Konarev, A. V. (2021). Bioactive Components in Oat and Barley Grain as a Promising Breeding Trend for Functional Food Production. Molecules, 26(8), 2260. https://doi.org/10.3390/molecules26082260