
The Effects of Prolonged Storage on ARPE-19 Cells Stored at Three Different Storage Temperatures

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Effect of Three-Week Storage on the Metabolism of Cultured ARPE-19 Cells
2.2. Effect of Three-Week Storage on the Morphology of Cultured ARPE-19 Cells
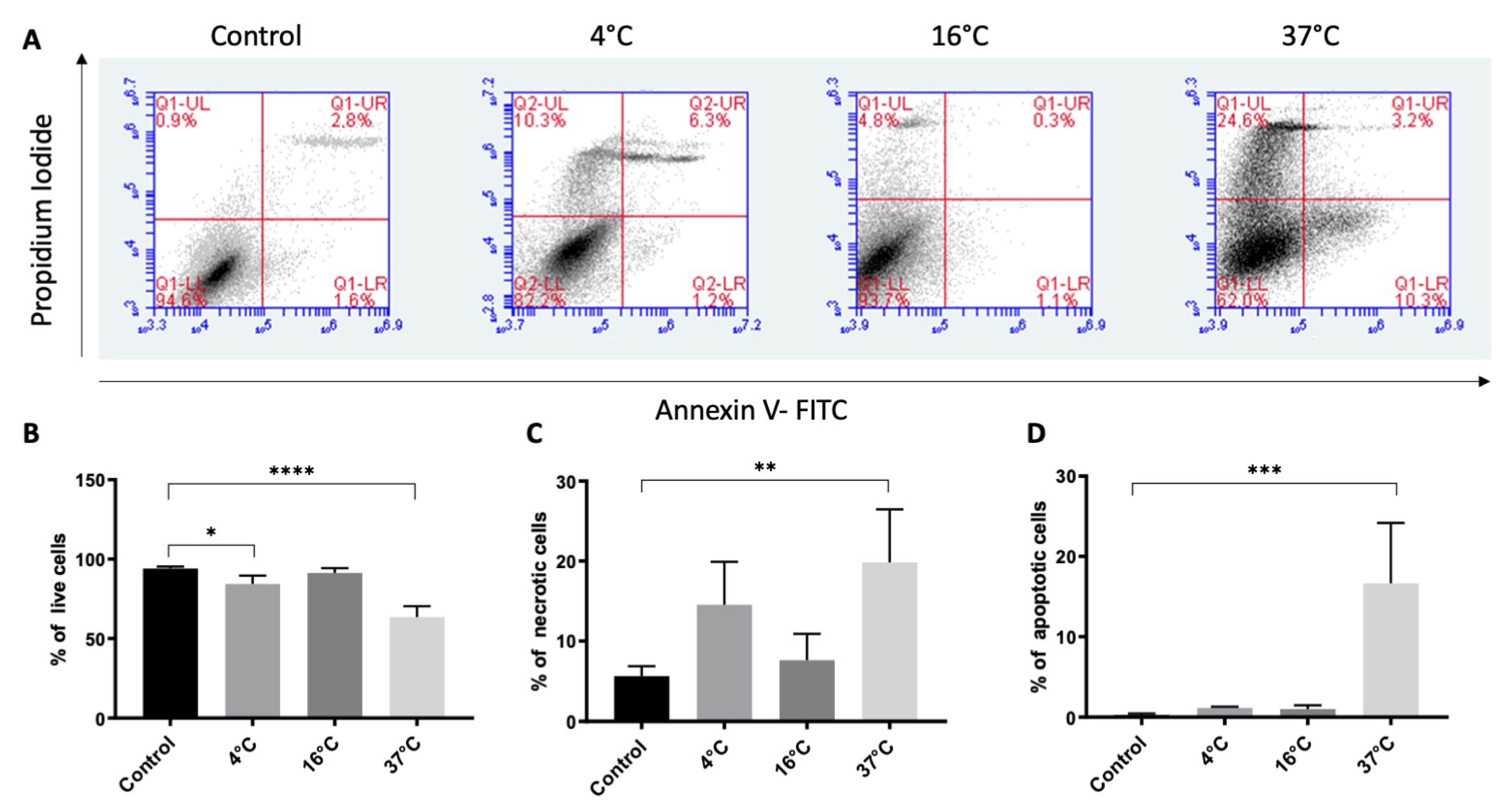
2.3. Effect of Three-Week Storage on the Viability of Cultured ARPE-19 Cells
2.4. Effect of Three-Week Storage on the Phenotype of Cultured ARPE-19 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture Media and Reagents
4.2. Culture of ARPE-19 Cells
4.3. Storage of ARPE-19 Cells
4.4. Metabolic Analysis
4.5. Morphology Analysis
4.6. Viability Analysis
4.7. Phenotype Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Marmorstein, A.D. The Polarity of the Retinal Pigment Epithelium. Traffic 2001, 2, 867–872. [Google Scholar] [CrossRef]
- De Jong, P.T.V.M. Age-related macular degeneration. N. Engl. J. Med. 2006, 355, 1474–1485. [Google Scholar] [CrossRef]
- Hicks, D.; Hamel, C.P. The Retinal Pigment Epithelium in Health and Disease. Curr. Mol. Med. 2010, 10, 802–823. [Google Scholar] [CrossRef]
- Vaajasaari, H.; Ilmarinen, T.; Juuti-Uusitalo, K.; Rajala, K.; Onnela, N.; Narkilahti, S.; Suuronen, R.; Hyttinen, J.; Uusitalo, H.; Skottman, H. Toward the defined and xeno-free differentiation of functional human pluripotent stem cell–derived retinal pigment epithelial cells. Mol. Vis. 2011, 17, 558–575. [Google Scholar]
- Alexander, P.; Thomson, H.A.J.; Luff, A.J.; Lotery, A.J. Retinal pigment epithelium transplantation: Concepts, challenges, and future prospects. Eye 2015, 29, 992–1002. [Google Scholar] [CrossRef]
- Nazari, H.; Zhang, L.; Zhu, D.; Chader, G.J.; Falabella, P.; Stefanini, F.R.; Rowland, T.J.; Clegg, D.O.; Kashani, A.H.; Hinton, D.R.; et al. Stem cell based therapies for age-related macular degeneration: The promises and the challenges. Prog. Retin. Eye Res. 2015, 48, 39. [Google Scholar] [CrossRef]
- Mandai, M.; Watanabe, A.; Kurimoto, Y.; Hirami, Y.; Morinaga, C.; Daimon, T.; Fujihara, M.; Akimaru, H.; Sakai, N.; Shibata, Y.; et al. Autologous Induced Stem-Cell–Derived Retinal Cells for Macular Degeneration. N. Engl. J. Med. 2017, 376, 1038–1046. [Google Scholar] [CrossRef]
- Crafoord, S.; Algvere, P.V.; Seregard, S.; Kopp, E.D. Long-term outcome of RPE allografts to the subretinal space of rabbits. Acta Ophthalmol. Scand. 1999, 77, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Lund, R.D.; Adamson, P.; Sauvé, Y.; Keegan, D.J.; Girman, S.V.; Wang, S.; Winton, H.; Kanuga, N.; Kwan, A.S.L.; Beauchène, L.; et al. Subretinal transplantation of genetically modified human cell lines attenuates loss of visual function in dystrophic rats. Proc. Natl. Acad. Sci. USA 2001, 98, 9942–9947. [Google Scholar] [CrossRef]
- Wang, H.; Leonard, D.S.; Castellarin, A.; Tsukahara, I.; Ninomiya, Y.; Yagi, F.; Cheewatrakoolpong, N.; Sugino, I.K.; Zarbin, M. Short-term study of allogeneic retinal pigment epithelium transplants onto debrided Bruch’s membrane. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2990–2999. [Google Scholar]
- Coffey, P.J.; Girman, S.; Wang, S.M.; Hetherington, L.; Keegan, D.J.; Adamson, P.; Greenwood, J.; Lund, R.D. Long-term preservation of cortically dependent visual function in RCS rats by transplantation. Nat. Neurosci. 2001, 5, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Girman, S.V.; Wang, S.; Lund, R.D. Cortical visual functions can be preserved by subretinal RPE cell grafting in RCS rats. Vis. Res. 2003, 43, 1817–1827. [Google Scholar] [CrossRef]
- Haruta, M.; Sasai, Y.; Kawasaki, H.; Amemiya, K.; Ooto, S.; Kitada, M.; Suemori, H.; Nakatsuji, N.; Ide, C.; Honda, Y.; et al. In vitro and in vivo characterization of pigment epithelial cells differentiated from primate embryonic stem cells. Investig. Opthalmol. Vis. Sci. 2004, 45, 1020–1025. [Google Scholar] [CrossRef] [PubMed]
- McGill, T.; Lund, R.; Douglas, R.; Wang, S.; Lu, B.; Prusky, G. Preservation of vision following cell-based therapies in a model of retinal degenerative disease. Vis. Res. 2004, 44, 2559–2566. [Google Scholar] [CrossRef][Green Version]
- Del Priore, L.V.; Tezel, T.H.; Kaplan, H.J. Survival of allogeneic porcine retinal pigment epithelial sheets after subretinal transplantation. Investig. Opthalmol. Vis. Sci. 2004, 45, 985–992. [Google Scholar] [CrossRef]
- Wang, S.; Lu, B.; Wood, P.; Lund, R.D. Grafting of ARPE-19 and Schwann Cells to the Subretinal Space in RCS Rats. Investig. Opthalmol. Vis. Sci. 2005, 46, 2552–2560. [Google Scholar] [CrossRef]
- Lund, R.D.; Wang, S.; Klimanskaya, I.; Holmes, T.; Ramos-Kelsey, R.; Lu, B.; Girman, S.; Bischoff, N.; Sauvé, Y.; Lanza, R. Human Embryonic Stem Cell–Derived Cells Rescue Visual Function in Dystrophic RCS Rats. Cloning Stem Cells 2006, 8, 189–199. [Google Scholar] [CrossRef]
- Yaji, N.; Yamato, M.; Yang, J.; Okano, T.; Hori, S. Transplantation of tissue-engineered retinal pigment epithelial cell sheets in a rabbit model. Biomaterials 2009, 30, 797–803. [Google Scholar] [CrossRef]
- Carr, A.-J.F.; Vugler, A.A.; Hikita, S.T.; Lawrence, J.M.; Gias, C.; Chen, L.L.; Buchholz, D.E.; Ahmado, A.; Semo, M.; Smart, M.J.K.; et al. Protective Effects of Human iPS-Derived Retinal Pigment Epithelium Cell Transplantation in the Retinal Dystrophic Rat. PLoS ONE 2009, 4, e8152. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, L.; Lu, B.; Zhu, D.; Ribeiro, R.; Diniz, B.; Thomas, P.B.; Ahuja, A.K.; Hinton, D.R.; Tai, Y.-C.; et al. A Novel Approach for Subretinal Implantation of Ultrathin Substrates Containing Stem Cell-Derived Retinal Pigment Epithelium Monolayer. Ophthalmic Res. 2012, 48, 186–191. [Google Scholar] [CrossRef]
- Li, Y.; Tsai, Y.-T.; Hsu, C.-W.; Erol, D.; Yang, J.; Wu, W.-H.; Davis, R.J.; Egli, D.; Tsang, S.H. Long-term Safety and Efficacy of Human-Induced Pluripotent Stem Cell (iPS) Grafts in a Preclinical Model of Retinitis Pigmentosa. Mol. Med. 2012, 18, 1312–1319. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Mandai, M.; Kamao, H.; Hashiguchi, T.; Shikamura, M.; Kawamata, S.; Sugita, S.; Takahashi, M. Protective Effects of Human iPS-Derived Retinal Pigmented Epithelial Cells in Comparison with Human Mesenchymal Stromal Cells and Human Neural Stem Cells on the Degenerating Retina inrd1mice. Stem Cells 2015, 33, 1543–1553. [Google Scholar] [CrossRef]
- Schwartz, S.D.; Hubschman, J.-P.; Heilwell, G.; Franco-Cardenas, V.; Pan, C.K.; Ostrick, R.M.; Mickunas, E.; Gay, R.; Klimanskaya, I.; Lanza, R. Embryonic stem cell trials for macular degeneration: A preliminary report. Lancet 2012, 379, 713–720. [Google Scholar] [CrossRef]
- Schwartz, S.D.; Regillo, C.D.; Lam, B.L.; Eliott, D.; Rosenfeld, P.J.; Gregori, N.Z.; Hubschman, J.-P.; Davis, J.L.; Heilwell, G.; Spirn, M.; et al. Human embryonic stem cell-derived retinal pigment epithelium in patients with age-related macular degeneration and Stargardt’s macular dystrophy: Follow-up of two open-label phase 1/2 studies. Lancet 2015, 385, 509–516. [Google Scholar] [CrossRef]
- Schwartz, S.D.; Tan, G.; Hosseini, H.; Nagiel, A. Subretinal Transplantation of Embryonic Stem Cell–Derived Retinal Pigment Epithelium for the Treatment of Macular Degeneration: An Assessment at 4 Years. Investig. Opthalmol. Vis. Sci. 2016, 57, ORSFc1–ORSFc9. [Google Scholar] [CrossRef]
- Da Cruz, L.; Fynes, K.; Georgiadis, O.; Kerby, J.; Luo, Y.H.; Ahmado, A.; Vernon, A.; Daniels, J.T.; Nommiste, B.; Hasan, S.M.; et al. Phase 1 clinical study of an embryonic stem cell–derived retinal pigment epithelium patch in age-related macular degeneration. Nat. Biotechnol. 2018, 36, 328–337. [Google Scholar] [CrossRef]
- Bracha, P.; A Moore, N.; A Ciulla, T. Induced pluripotent stem cell-based therapy for age-related macular degeneration. Expert Opin. Biol. Ther. 2017, 17, 1113–1126. [Google Scholar] [CrossRef]
- Ramsden, C.M.; Powner, M.B.; Carr, A.-J.F.; Smart, M.J.K.; Da Cruz, L.; Coffey, P.J. Stem cells in retinal regeneration: Past, present and future. Deversity 2013, 140, 2576–2585. [Google Scholar] [CrossRef]
- Ramsden, C.M.; Da Cruz, L.; Coffey, P. Stemming the Tide of Age-Related Macular Degeneration: New Therapies for Old Retinas. Investig. Opthalmol. Vis. Sci. 2016, 57. [Google Scholar] [CrossRef]
- Daniels, J.T.; Secker, G.A.; Shortt, A.J.; Tuft, S.J.; Seetharaman, S. Stem cell therapy delivery: Treading the regulatory tightrope. Regen. Med. 2006, 1, 715–719. [Google Scholar] [CrossRef]
- Oie, Y.; Nozaki, T.; Takayanagi, H.; Hara, S.; Hayashi, R.; Takeda, S.; Mori, K.; Moriya, N.; Soma, T.; Tsujikawa, M.; et al. Development of a Cell Sheet Transportation Technique for Regenerative Medicine. Tissue Eng. Part C Methods 2014, 20, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Pasovic, L.; Utheim, T.P.; Maria, R.; Lyberg, T.; Messelt, E.B.; Aabel, P.; Chen, D.F.; Chen, X.; Eidet, J.R. Optimization of Storage Temperature for Cultured ARPE-19 Cells. J. Ophthalmol. 2013, 2013, 11. [Google Scholar] [CrossRef] [PubMed]
- Pasovic, L.; Eidet, J.R.; Brusletto, B.S.; Lyberg, T.; Utheim, T.P. Effect of Storage Temperature on Key Functions of Cultured Retinal Pigment Epithelial Cells. J. Ophthalmol. 2015, 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Pasovic, L.; Eidet, J.R.; Olstad, O.K.; Chen, D.F.; Lyberg, T.; Utheim, T.P. Impact of Storage Temperature on the Expression of Cell Survival Genes in Cultured ARPE-19 Cells. Curr. Eye Res. 2016, 42, 134–144. [Google Scholar] [CrossRef]
- Khan, A.Z.; Utheim, T.P.; Reppe, S.; Sandvik, L.; Lyberg, T.; Roald, B.B.-H.; Ibrahim, I.B.; Eidet, J.R. Cultured Human Retinal Pigment Epithelial (hRPE) Sheets: A Search for Suitable Storage Conditions. Microsc. Microanal. 2018, 24, 147–155. [Google Scholar] [CrossRef]
- Kitahata, S.; Tanaka, Y.; Hori, K.; Kime, C.; Sugita, S.; Ueda, H.; Takahashi, M. Critical Functionality Effects from Storage Temperature on Human Induced Pluripotent Stem Cell-Derived Retinal Pigment Epithelium Cell Suspensions. Sci. Rep. 2019, 9, 2891. [Google Scholar] [CrossRef]
- Wang, A.W.; Zhang, H.; Ikemoto, I.; Anderson, D.J.; Loughlin, K.R. Reactive oxygen species generation by seminal cells during cryopreservation. Urology 1997, 49, 921–925. [Google Scholar] [CrossRef]
- Honda, S.; Weigel, A.; Hjelmeland, L.M.; Handa, J.T. Induction of Telomere Shortening and Replicative Senescence by Cryopreservation. Biochem. Biophys. Res. Commun. 2001, 282, 493–498. [Google Scholar] [CrossRef]
- Pegg, D.E. The History and Principles of Cryopreservation. Semin. Reprod. Med. 2002, 20, 005–014. [Google Scholar] [CrossRef]
- Ahmado, A.; Carr, A.-J.; Vugler, A.A.; Semo, M.; Gias, C.; Lawrence, J.M.; Chen, L.L.; Chen, F.K.; Turowski, P.; Da Cruz, L.; et al. Induction of Differentiation by Pyruvate and DMEM in the Human Retinal Pigment Epithelium Cell Line ARPE-19. Investig. Opthalmol. Vis. Sci. 2011, 52, 7148–7159. [Google Scholar] [CrossRef]
- Jackson, C.J.; Aabel, P.; Eidet, J.R.; Messelt, E.B.; Lyberg, T.; Von Unge, M.; Utheim, T.P. Effect of Storage Temperature on Cultured Epidermal Cell Sheets Stored in Xenobiotic-Free Medium. PLoS ONE 2014, 9, e105808. [Google Scholar] [CrossRef]
- Eidet, J.R.; Utheim, Ø.A.; Islam, R.; Lyberg, T.; Messelt, E.B.; Dartt, D.A.; Utheim, T.P. The Impact of Storage Temperature on the Morphology, Viability, Cell Number and Metabolism of Cultured Human Conjunctival Epithelium. Curr. Eye Res. 2015, 40, 30–39. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Islam, R.; Jackson, C.J.; Eidet, J.R.; Messelt, E.B.; Corraya, R.M.; Lyberg, T.; Griffith, M.; Dartt, D.A.; Utheim, T.P. Effect of Storage Temperature on Structure and Function of Cultured Human Oral Keratinocytes. PLoS ONE 2015, 10, e0128306. [Google Scholar] [CrossRef] [PubMed]
- Slikker, W.; Desai, V.G.; Duhart, H.; Feuers, R.; Imam, S.Z. Hypothermia enhances bcl-2 expression and protects against oxidative stress-induced cell death in chinese hamster ovary cells. Free. Radic. Biol. Med. 2001, 31, 405–411. [Google Scholar] [CrossRef]
- Park, H.J.; Lyons, J.C.; Ohtsubo, T.; Song, C.W. Acidic environment causes apoptosis by increasing caspase activity. Br. J. Cancer 1999, 80, 1892–1897. [Google Scholar] [CrossRef] [PubMed]
- Marmor, M.F. Control of subretinal fluid: Experimental and clinical studies. Eye 1990, 4, 340–344. [Google Scholar] [CrossRef]
- Dunn, K.; Aotaki-Keen, A.; Putkey, F.; Hjelmeland, L. ARPE-19, A Human Retinal Pigment Epithelial Cell Line with Differentiated Properties. Exp. Eye Res. 1996, 62, 155–170. [Google Scholar] [CrossRef]
- Dunn, K.C.; Marmorstein, A.D.; Bonilha, V.L.; Rodriguez-Boulan, E.; Giordano, F.; Hjelmeland, L.M. Use of the ARPE-19 cell line as a model of RPE polarity: Basolateral secretion of FGF5. Investig. Ophthalmol. Vis. Sci. 1998, 39, 6. [Google Scholar]
- Tian, J.; Ishibashi, K.; Honda, S.; A Boylan, S.; Hjelmeland, L.M.; Handa, J.T. The expression of native and cultured human retinal pigment epithelial cells grown in different culture conditions. Br. J. Ophthalmol. 2005, 89, 1510–1517. [Google Scholar] [CrossRef]
- Luo, Y.; Zhuo, Y.; Fukuhara, M.; Rizzolo, L.J. Effects of Culture Conditions on Heterogeneity and the Apical Junctional Complex of the ARPE-19 Cell Line. Investig. Opthalmol. Vis. Sci. 2006, 47, 3644–3655. [Google Scholar] [CrossRef][Green Version]
- Geisen, P.; McColm, J.R.; King, B.M.; Hartnett, M.E. Characterization of Barrier Properties and Inducible VEGF Expression of Several Types of Retinal Pigment Epithelium in Medium-Term Culture. Curr. Eye Res. 2006, 31, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Samuel, W.; Jaworski, C.; Postnikova, O.A.; Kutty, R.K.; Duncan, T.; Tan, L.X.; Poliakov, E.; Lakkaraju, A.; Redmond, T.M. Appropriately differentiated ARPE-19 cells regain phenotype and gene expression profiles similar to those of native RPE cells. Mol. Vis. 2017, 23, 60–89. [Google Scholar] [PubMed]
- Tian, J.; Ishibashi, K.; Handa, J.T. The expression of native and cultured RPE grown on different matrices. Physiol. Genom. 2004, 17, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Utheim, T.P.; Raeder, S.; Utheim, Ø.A.; Cai, Y.; Roald, B.; Drolsum, L.; Lyberg, T.; Nicolaissen, B. A novel method for preserving cultured limbal epithelial cells. Br. J. Ophthalmol. 2006, 91, 797–800. [Google Scholar] [CrossRef] [PubMed]
- Utheim, T.P.; Islam, R.; Fostad, I.G.; Eidet, J.R.; Sehic, A.; Olstad, O.K.; Dartt, D.A.; Messelt, E.B.; Griffith, M.; Pasovic, L. Storage Temperature Alters the Expression of Differentiation-Related Genes in Cultured Oral Keratinocytes. PLoS ONE 2016, 11, e0152526. [Google Scholar] [CrossRef]
- Jackson, C.; Eidet, J.R.; Reppe, S.; Aass, H.C.D.; Tønseth, K.A.; Roald, B.; Lyberg, T.; Utheim, T.P. Effect of Storage Temperature on the Phenotype of Cultured Epidermal Cells Stored in Xenobiotic-Free Medium. Curr. Eye Res. 2015, 41, 757–768. [Google Scholar] [CrossRef]
- Pellegrin, S.; Mellor, H. Actin stress fibres. J. Cell Sci. 2007, 120, 3491–3499. [Google Scholar] [CrossRef]
- Tojkander, S.; Gateva, G.; Lappalainen, P. Actin stress fibers—assembly, dynamics and biological roles. J. Cell Sci. 2012, 125, 1855–1864. [Google Scholar] [CrossRef]
- Narimatsu, T.; Ozawa, Y.; Miyake, S.; Kubota, S.; Hirasawa, M.; Nagai, N.; Shimmura, S.; Tsubota, K. Disruption of Cell-Cell Junctions and Induction of Pathological Cytokines in the Retinal Pigment Epithelium of Light-Exposed Mice. Investig. Opthalmol. Vis. Sci. 2013, 54, 4555–4562. [Google Scholar] [CrossRef]
- Eidet, J.R.; Pasovic, L.; Maria, R.; Jackson, C.J.; Utheim, T.P. Objective assessment of changes in nuclear morphology and cell distribution following induction of apoptosis. Diagn. Pathol. 2014, 9, 92. [Google Scholar] [CrossRef]
- Vermes, I.; Haanen, C.; Steffens-Nakken, H.; Reutellingsperger, C. A novel assay for apoptosis Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled Annexin V. J. Immunol. Methods 1995, 184, 39–51. [Google Scholar] [CrossRef]
- Khan, A.Z.; Utheim, T.P.; Jackson, C.J.; Reppe, S.; Lyberg, T.; Eidet, J.R. Nucleus Morphometry in Cultured Epithelial Cells Correlates with Phenotype. Microsc. Microanal. 2016, 22, 612–620. [Google Scholar] [CrossRef] [PubMed]




Sample Availability: Samples of the compounds are not available from the authors. Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, R.; Corraya, R.M.; Pasovic, L.; Khan, A.Z.; Aass, H.C.D.; Eidet, J.R.; Utheim, T.P. The Effects of Prolonged Storage on ARPE-19 Cells Stored at Three Different Storage Temperatures. Molecules 2020, 25, 5809. https://doi.org/10.3390/molecules25245809
Islam R, Corraya RM, Pasovic L, Khan AZ, Aass HCD, Eidet JR, Utheim TP. The Effects of Prolonged Storage on ARPE-19 Cells Stored at Three Different Storage Temperatures. Molecules. 2020; 25(24):5809. https://doi.org/10.3390/molecules25245809
Chicago/Turabian StyleIslam, Rakibul, Rima Maria Corraya, Lara Pasovic, Ayyad Zartasht Khan, Hans Christian D. Aass, Jon Roger Eidet, and Tor Paaske Utheim. 2020. "The Effects of Prolonged Storage on ARPE-19 Cells Stored at Three Different Storage Temperatures" Molecules 25, no. 24: 5809. https://doi.org/10.3390/molecules25245809
APA StyleIslam, R., Corraya, R. M., Pasovic, L., Khan, A. Z., Aass, H. C. D., Eidet, J. R., & Utheim, T. P. (2020). The Effects of Prolonged Storage on ARPE-19 Cells Stored at Three Different Storage Temperatures. Molecules, 25(24), 5809. https://doi.org/10.3390/molecules25245809