Cell-Free Demineralized Bone Matrix for Mesenchymal Stem Cells Survival and Colonization

 ,
,  ,
, 

Abstract
1. Introduction
2. Results
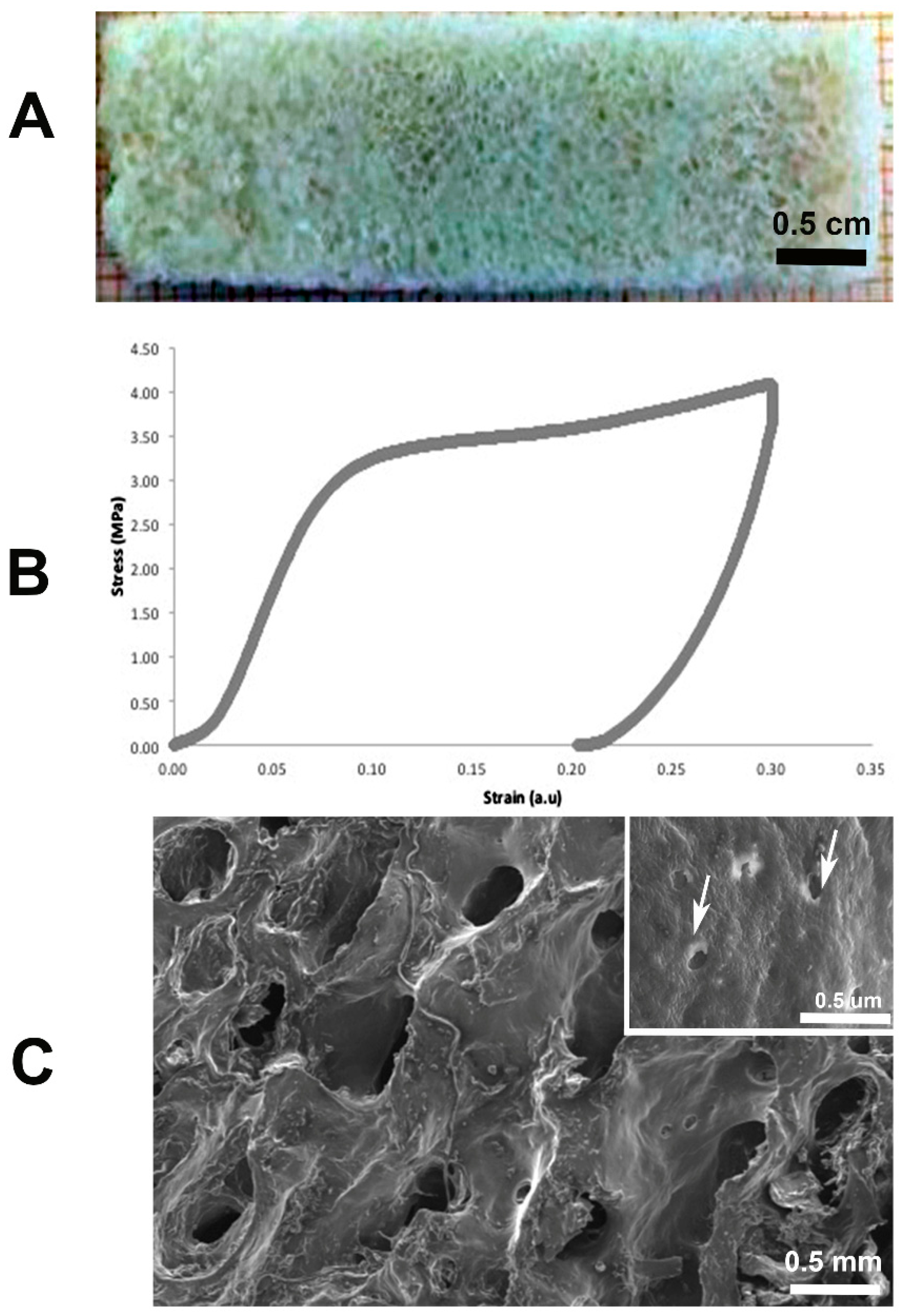
2.1. Bone Demineralization and Mechanical Tests
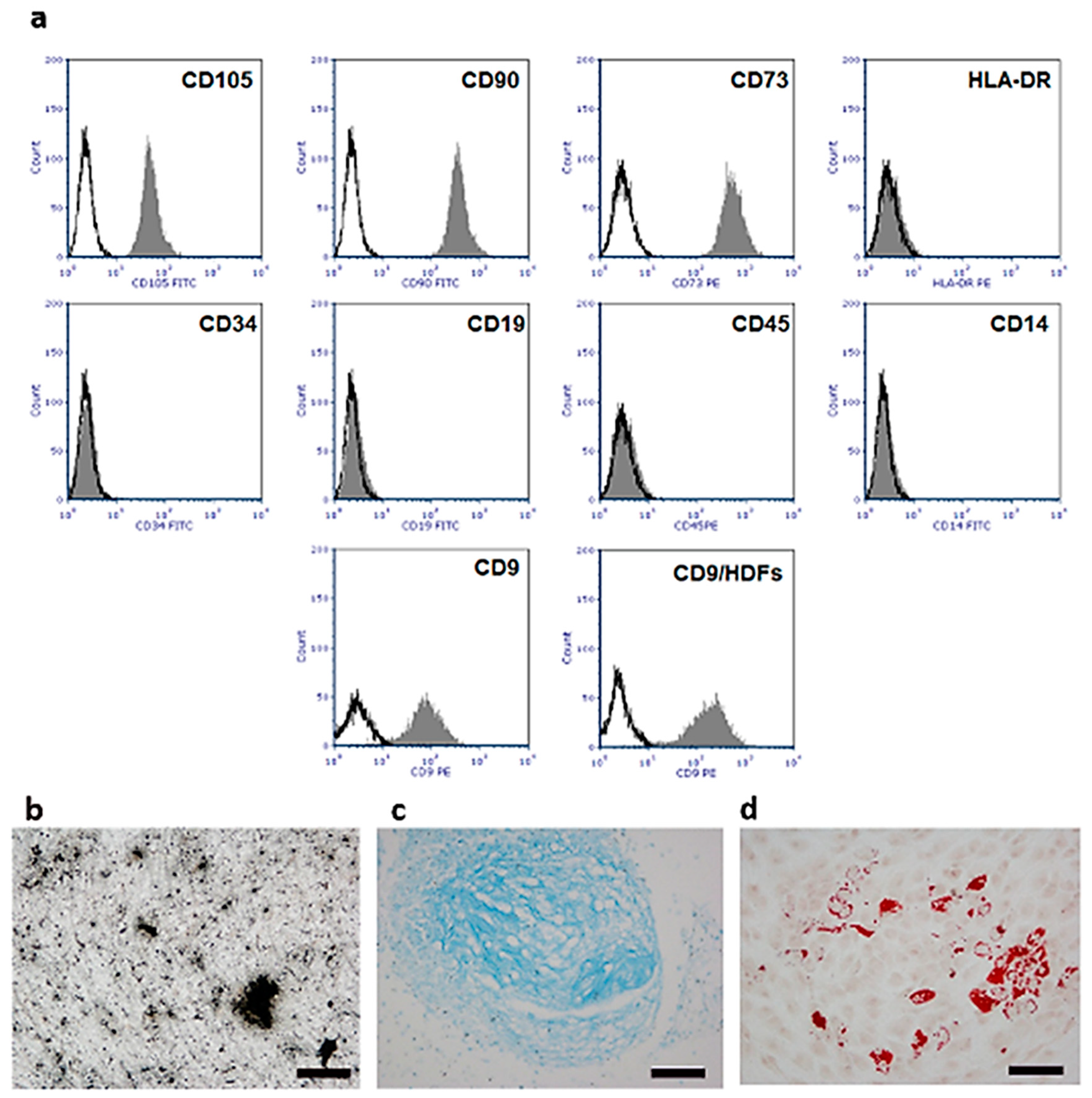
2.2. Isolation and Characterization of Human Umbilical Cord Stem Cells (hUC-SCs)
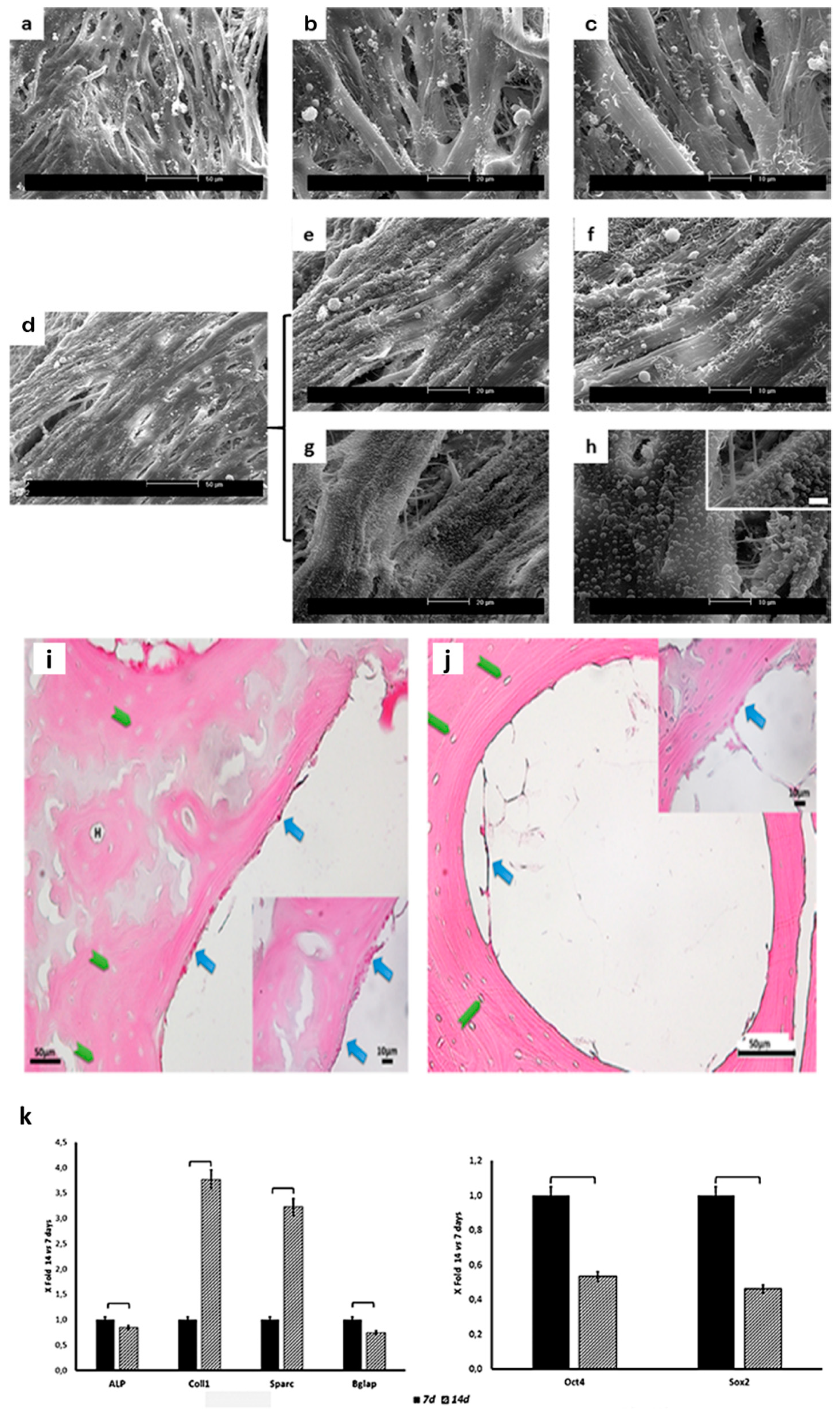
2.3. Scaffold Colonization
2.4. Gene Expression
3. Discussion
4. Materials and Methods
4.1. Bone Demineralization and Mechanical Tests
4.2. Isolation and Culture of Human Umbilical Cord Mesenchymal Stem Cells (hUC-SCs)
4.3. hUC-SC In Vitro Differentiation
4.4. Scaffold Seeding and Culture
4.5. Scanning Electron Microscopy (SEM)
4.6. Light Microscopy (LM)
4.7. Real-Time Quantitative Reverse Transcription PCR (qRT-PCR)
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murphy, W.L.; McDevitt, T.C.; Engler, A.J. Materials as stem cell regulators. Nat. Mater. 2014, 13, 547–557. [Google Scholar] [CrossRef]
- Mattioli-Belmonte, M.; Vozzi, G.; Kyriakidou, K.; Pulieri, E.; Lucarini, G.; Vinci, B.; Pugnaloni, A.; Biagini, G.; Ahluwalia, A. Rapid-prototyped and salt-leached PLGA scaffolds condition cell morpho-functional behavior. J. Biomed. Mater. Res. A 2008, 85, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Santoro, M.; Perale, G. Polymeric scaffolds as stem cell carriers in bone repair. J. Tissue Eng. Regen. Med. 2015, 9, 1093–1119. [Google Scholar] [PubMed]
- Ciardelli, G.; Gentile, P.; Chiono, V.; Mattioli-Belmonte, M.; Vozzi, G.; Barbani, N.; Giusti, P. Enzymatically crosslinked porous composite matrices for bone tissue regeneration. J. Biomed. Mater. Res. A 2010, 92, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Jelen, C.; Mattei, G.; Montemurro, F.; De Maria, C.; Mattioli-Belmonte, M.; Vozzi, G. Bone scaffolds with homogeneous and discrete gradient mechanical properties. Mat. Sci. Eng. C. 2013, 3, 28–36. [Google Scholar] [CrossRef]
- Rajangam, T.; An, S.S. Fibrinogen and fibrin-based micro and nano scaffolds incorporated with drugs, proteins, cells and genes for therapeutic biomedical applications. Int. J. Nanomed. 2013, 8, 3641–3662. [Google Scholar]
- Kruger, T.E.; Miller, A.H.; Wang, J. Collagen scaffolds in bone sialoprotein-mediated bone regeneration. Sci. World J. 2013. [Google Scholar] [CrossRef]
- Vindigni, V.; Cortivo, R.; Iacobellis, L.; Abatangelo, G.; Zavan, B. Hyaluronan benzyl ester as a scaffold for tissue engineering. Int. J. Mol. Sci. 2009, 10, 2972–2985. [Google Scholar] [CrossRef]
- Gloria, A.; De Santis, R.; Ambrosio, L. Polymer-based composite scaffolds for tissue engineering. J. Appl. Biomater. Biomech. 2010, 8, 57–67. [Google Scholar] [PubMed]
- Boccaccini, A.R.; Blaker, J.J. Bioactive composite materials for tissue engineering scaffolds. Expert. Rev. Med. Devices 2005, 2, 303–317. [Google Scholar] [CrossRef]
- Shi, X.; Sitharaman, B.; Pham, Q.P.; Liang, F.; Wu, K.; Edward Billups, W.; Wilson, L.J.; Mikos, A.G. Fabrication of porous ultra-short single-walled carbon nanotube nanocomposite scaffolds for bone tissue engineering. Biomaterials 2007, 28, 4078–4090. [Google Scholar] [CrossRef] [PubMed]
- Veetil, J.V.; Ye, K. Tailored carbon nanotubes for tissue engineering applications. Biotechnol. Prog. 2009, 25, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Mattioli-Belmonte, M.; Vozzi, G.; Whulanza, Y.; Seggiani, M.; Fantauzzi, V.; Orsini, G.; Ahluwalia, A. Tuning polycaprolactone–carbon nanotube composites for bone tissue engineering scaffolds. Mat. Sci. Eng. C 2012, 32, 152–159. [Google Scholar] [CrossRef]
- Whulanza, Y.; Battini, E.; Vannozzi, L.; Vomero, M.; Ahluwalia, A.; Vozzi, G. Electrical and Mechanical Characterisation of single wall carbon nanotubes-based composites for tissue engineering applications. J. Nanosci. Nanotechnol. 2013, 13, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Mattioli-Belmonte, M.; De Maria, C.; Vitale-Brovarone, C.; Baino, F.; Dicarlo, M.; Vozzi, G. Pressure-activated microsyringe (PAM) fabrication of bioactive glass-poly (lactic-co-glycolic acid) composite scaffolds for bone tissue regeneration. J. Tissue Eng. Regen. Med. 2017, 11, 1986–1997. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.A., Jr.; Hoehn, J.G. Use of commercial porcine skin for wound dressings. Plast. Reconstr. Surg. 1973, 52, 401–405. [Google Scholar] [CrossRef]
- Edgar, L.; McNamara, K.; Wong, T.; Tamburrini, R.; Katari, R.; Orlando, G. Heterogeneity of Scaffold Biomaterials in Tissue Engineering. Materials 2016, 9, 332. [Google Scholar] [CrossRef]
- Bissell, M.J.; Hall, H.G.; Parry, G. How does the extracellular matrix direct gene expression? J. Theor. Biol. 1982, 99, 31–68. [Google Scholar] [CrossRef]
- Lin, X.; Fang, X.; Wang, Q.; Hu, Z.; Chen, K.; Shan, Z.; Chen, S.; Wang, J.; Mo, J.; Ma, J.; et al. Decellularized allogeneic intervertebral disc: Natural biomaterials for regenerating disc degeneration. Oncotarget 2016, 7, 12121–12136. [Google Scholar] [CrossRef][Green Version]
- Saldin, L.T.; Cramer, M.C.; Velankar, S.S.; White, L.J.; Badylak, S.F. Extracellular matrix hydrogels from decellularized tissues: Structure and function. Acta Biomater. 2017, 49, 1–15. [Google Scholar] [CrossRef]
- Cheng, C.W.; Solorio, L.D.; Alsberg, E. Decellularized tissue and cell-derived extracellular matrices as scaffolds for orthopaedic tissue engineering. Biotechn. Adv. 2014, 32, 462–484. [Google Scholar] [CrossRef]
- Roberts, T.T.; Rosenbaum, A.J. Bone grafts, bone substitutes and orthobiologics. The bridge between basic science and clinical advancements in fracture healing. Organogenesis 2012, 8, 114–124. [Google Scholar] [CrossRef]
- Mattei, G.; Ferretti, C.; Tirella, A.; Ahluwalia, A.; Mattioli-Belmonte, M. Decoupling the role of stiffness from other hydroxyapatite signalling cues in periosteal derived stem cell differentiation. Sci. Rep. 2015, 5, 10778. [Google Scholar] [CrossRef]
- Even-Ram, S.; Artym, V.; Yamada, K.M. Matrix control of stem cell fate. Cell 2006, 126, 645–647. [Google Scholar] [CrossRef]
- Eagle, M.J.; Rooney, P.; Kearney, J.N. Optimized demineralization of human cancellous bone by application of a vacuum. J. Biomed. Mater. Res. B 2015, 103, 1023–1029. [Google Scholar] [CrossRef]
- Chen, P.Y.; McKittric, J. Compressive mechanical properties of demineralized and deproteinized cancellous bone. J. Mech. Behav. Biomed. Mater. 2011, 4, 961–973. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for defining multipotent mesenchymal stem cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Halfon, S.; Abramov, N.; Grinblat, B.; Ginis, I. Markers distinguishing mesenchymal stem cells from fibroblasts are downregulated with passaging. Stem Cells Dev. 2011, 20, 53–66. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Keane, T.J.; Swinehart, I.T.; Badylak, S.F. Methods of tissue decellularization used for preparation of biologic scaffolds and in vivo relevance. Methods 2015, 84, 25–34. [Google Scholar] [CrossRef]
- Xing, Q.; Yates, K.; Tahtinen, M.; Shearier, E.; Qian, Z.; Zhao, F. Decellularization of fibroblast cell sheets for natural extracellular matrix scaffold preparation. Tissue Eng. Part C Methods. 2014, 21, 77–87. [Google Scholar] [CrossRef]
- Voytik-Harbin, S.L.; Brightman, A.O. Small intestinal submucosa: A tissue derived extracellular matrix that promotes tissue-specific growth and differentiation of cells in vitro. Tissue Eng. 1998, 4, 157–174. [Google Scholar] [CrossRef]
- Badylak, S.F.; Freytes, D.O.; Gilbert, T.W. Extracellular matrix as a biological scaffold material: Structure and function. Acta Biomater. 2009, 5, 1–13. [Google Scholar] [CrossRef]
- Porzionato, A.; Stocco, E.; Barbon, S.; Grandi, F.; Macchi, V.; De Caro, R. Tissue-Engineered Grafts from Human Decellularized Extracellular Matrices: A Systematic Review and Future Perspectives. Int. J. Mol. Sci. 2018, 19, 4117. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, W.; Zhou, G.; Cao, Y.; Liu, W.; Zhang, Z.Y. Demineralized bone matrix-based microcarrier scaffold favors vascularized large bone regeneration in vivo in a rat model. J. Biomater. Appl. 2018, 33, 182–195. [Google Scholar] [CrossRef]
- Li, T.; Xia, M.; Gao, Y.; Chen, Y.; Xu, Y. Human umbilical cord mesenchymal stem cells: An overview of their potential in cell-based therapy. Expert Opin. Biol. Ther. 2015, 15, 1293–1306. [Google Scholar] [CrossRef]
- Wang, W.T.; Gu, P.; Qiu, F.C.; Zhang, L.N.; Zhang, Z.X.; Xie, B.C.; Dong, C.; Han, R.; Liu, H.M.; Yan, B.Y. Intravenous transplantation of Allograft hUC-MSC was more effective than subarachnoid transplantation of BM-MSCs in patients with Parkinson’s Syndrome and Secondary Parkinson’s Syndrome. J. Biomater. Tiss. Eng. 2016, 6, 158–164. [Google Scholar] [CrossRef]
- Moroncini, G.; Paolini, C.; Orlando, F.; Capelli, C.; Grieco, A.; Tonnini, C.; Agarbati, S.; Mondini, E.; Saccomanno, S.; Goteri, G.; et al. Mesenchymal stromal cells from human umbilical cord prevent the development of lung fibrosis in immunocompetent mice. PLoS ONE 2018, 13, e0196048. [Google Scholar] [CrossRef]
- Merlo, B.; Teti, G.; Mazzotti, E.; Ingrà, L.; Salvatore, V.; Buzzi, M.; Cerqueni, G.; Dicarlo, M.; Lanci, A.; Castagnetti, C.; et al. Wharton’s Jelly Derived Mesenchymal Stem Cells: Comparing Human and Horse. Stem Cell Rev. 2018, 14, 574–584. [Google Scholar] [CrossRef]
- Xu, J.; Sun, M.; Tan, Y.; Wang, H.; Wang, H.; Li, P.; Xu, Z.; Xia, Y.; Li, L.; Li, Y. Effect of matrix stiffness on the proliferation and differentiation of umbilical cord mesenchymal stem cells. Differentiation 2017, 96, 30–39. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef]
- Witkowska-Zimny, M.; Walenko, K.; Walkiewicz, A.E.; Pojda, Z.; Przybylski, J.; Lewandowska-Szumiel, M. Effect of substrate stiffness on differentiation of umbilical cord stem cells. Acta Biochim. Pol. 2016, 59, 261–264. [Google Scholar] [CrossRef]
- Duchamp de Lageneste, O.; Julien, A.; Abou-Khalil, R.; Frangi, G.; Carvalho, C.; Cagnard, N.; Cordier, C.; Conway, S.J.; Colnot, C. Periosteum contains skeletal stem cells with high bone regenerative potential controlled by Periostin. Nat. Commun. 2018, 9, 773. [Google Scholar] [CrossRef]
- Manilay, Z.; Novitskaya, E.; Sadovnikov, E.; McKittrick, J. A comparative study of young and mature bovine cortical bone. Acta Biomater. 2013, 9, 5280–5288. [Google Scholar] [CrossRef]
- Ramakrishnan, S.; Sulochana, KN. Manual of Medical Laboratory Techniques; JP Medical Ltd.: London, UK, 2012; p. 388. [Google Scholar]
- Ragni, E.; Viganò, M.; Rebulla, P.; Giordano, R.; Lazzari, L. What is beyond a qRT-PCR study on mesenchymal stem cell differentiation properties: How to choose the most reliable housekeeping genes. J. Cell. Mol. Med. 2013, 17, 168–180. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genes | 7 Days | 14 Days | |||
|---|---|---|---|---|---|
| Mean | DS | Mean | DS | p | |
| ALP | 2.93 × 10−4 | 1.47 × 10−5 | 2.46 × 10−4 | 1.23 × 10−5 | 0.039 |
| COL1 | 1.33 × 10−1 | 6.64 × 10−3 | 4.99 × 10−1 | 2.50 × 10−2 | 0.0096 |
| SPARC | 1.52 × 10−1 | 7.59 × 10−3 | 4.89 × 10−1 | 2.44 × 10−2 | 0.013 |
| BGLAP | 4.96 × 10−6 | 2.48 × 10−7 | 3.64 × 10−6 | 1.82 × 10−7 | 0.047 |
| Oct4 | 9.79 × 10−4 | 4.89 × 10−5 | 5.21 × 10−4 | 2.60 × 10−5 | 0.034 |
| Sox2 | 3.93 × 10−5 | 1.97 × 10−6 | 1.77 × 10−5 | 8.85 × 10−7 | 0.042 |
| Genes | Detected Transcript | Primer Forward (5′→3′) | Primer Reverse (3′→5′) | Amplicon Length (bp) |
|---|---|---|---|---|
| ALP | NM_007431 | GGCCAGCTACACCACAACA | CTGAGCGTTGGTGTTATATGTCTT | 96 |
| COL1 | NM_000088.3 | CCAACCCTTCCACCTTTGGAAGT | CCGGAGGTCCACAAAGCTGAA | 132 |
| SPARC | NM_003118.3 | CCTGAGGCTGTAACTGAGAGAAAG | GTGGGAGGGGAAACAAGAAGATAA | 142 |
| BGLAP | NM_199173 | GACTGTGACGAGTTGGCTGA | GCCCACAGATTCCTCTTCTG | 119 |
| Sox2 | NM_003106.3 | ACACCAATCCCATCCACACT | GCAAACTTCCTGCAAAGCTC | 198 |
| Oct4 | NM_203289.4 | AGCGAACCAGTATCGAGAAC | GCCTCAAAATCCTCTCGTTG | 199 |
| GUSB * | NM_000181.2 | AAACGATTGCAGGGTTTCAC | TCTCGTCGGTGACTGTTCA | 81 |
| GAPDH * | NM_002046.3 | AGCCACATCGCTCAGACAC | GCCCAATACGACCAAATCC | 200 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattioli-Belmonte, M.; Montemurro, F.; Licini, C.; Iezzi, I.; Dicarlo, M.; Cerqueni, G.; Coro, F.; Vozzi, G. Cell-Free Demineralized Bone Matrix for Mesenchymal Stem Cells Survival and Colonization. Materials 2019, 12, 1360. https://doi.org/10.3390/ma12091360
Mattioli-Belmonte M, Montemurro F, Licini C, Iezzi I, Dicarlo M, Cerqueni G, Coro F, Vozzi G. Cell-Free Demineralized Bone Matrix for Mesenchymal Stem Cells Survival and Colonization. Materials. 2019; 12(9):1360. https://doi.org/10.3390/ma12091360
Chicago/Turabian StyleMattioli-Belmonte, Monica, Francesca Montemurro, Caterina Licini, Iolanda Iezzi, Manuela Dicarlo, Giorgia Cerqueni, Florinda Coro, and Giovanni Vozzi. 2019. "Cell-Free Demineralized Bone Matrix for Mesenchymal Stem Cells Survival and Colonization" Materials 12, no. 9: 1360. https://doi.org/10.3390/ma12091360
APA StyleMattioli-Belmonte, M., Montemurro, F., Licini, C., Iezzi, I., Dicarlo, M., Cerqueni, G., Coro, F., & Vozzi, G. (2019). Cell-Free Demineralized Bone Matrix for Mesenchymal Stem Cells Survival and Colonization. Materials, 12(9), 1360. https://doi.org/10.3390/ma12091360