
Structures, Mechanisms, and Physiological Functions of Zinc Transporters in Different Biological Kingdoms


Abstract
1. Introduction
2. Zn2+ Transport Systems in Prokaryotes and Eukaryotes
3. ZnTs
3.1. Mammalian ZnTs
3.2. Plant ZnTs
3.3. Yeast ZnTs
3.4. Bacterial ZnTs
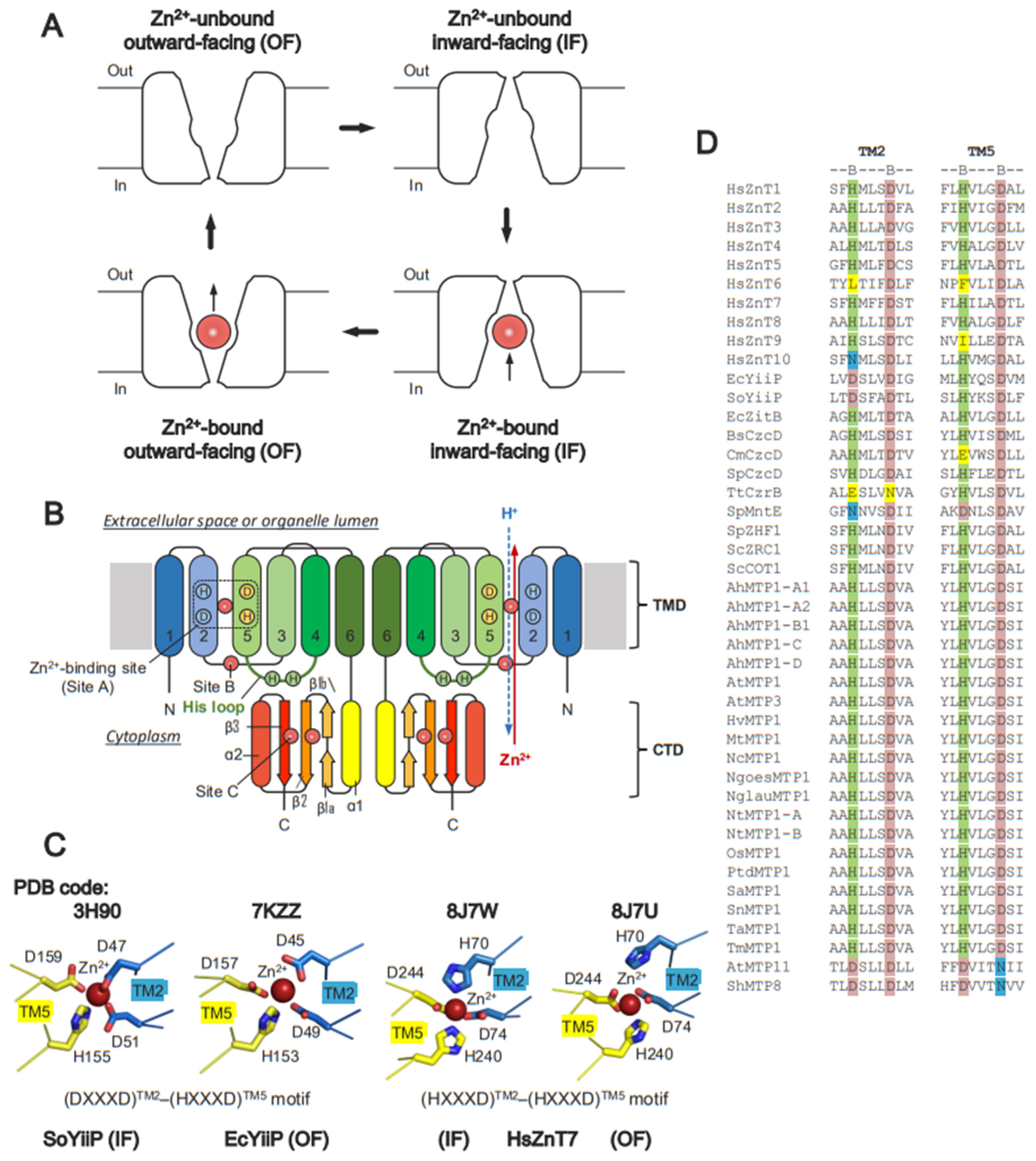
4. Structural Basis of ZnTs
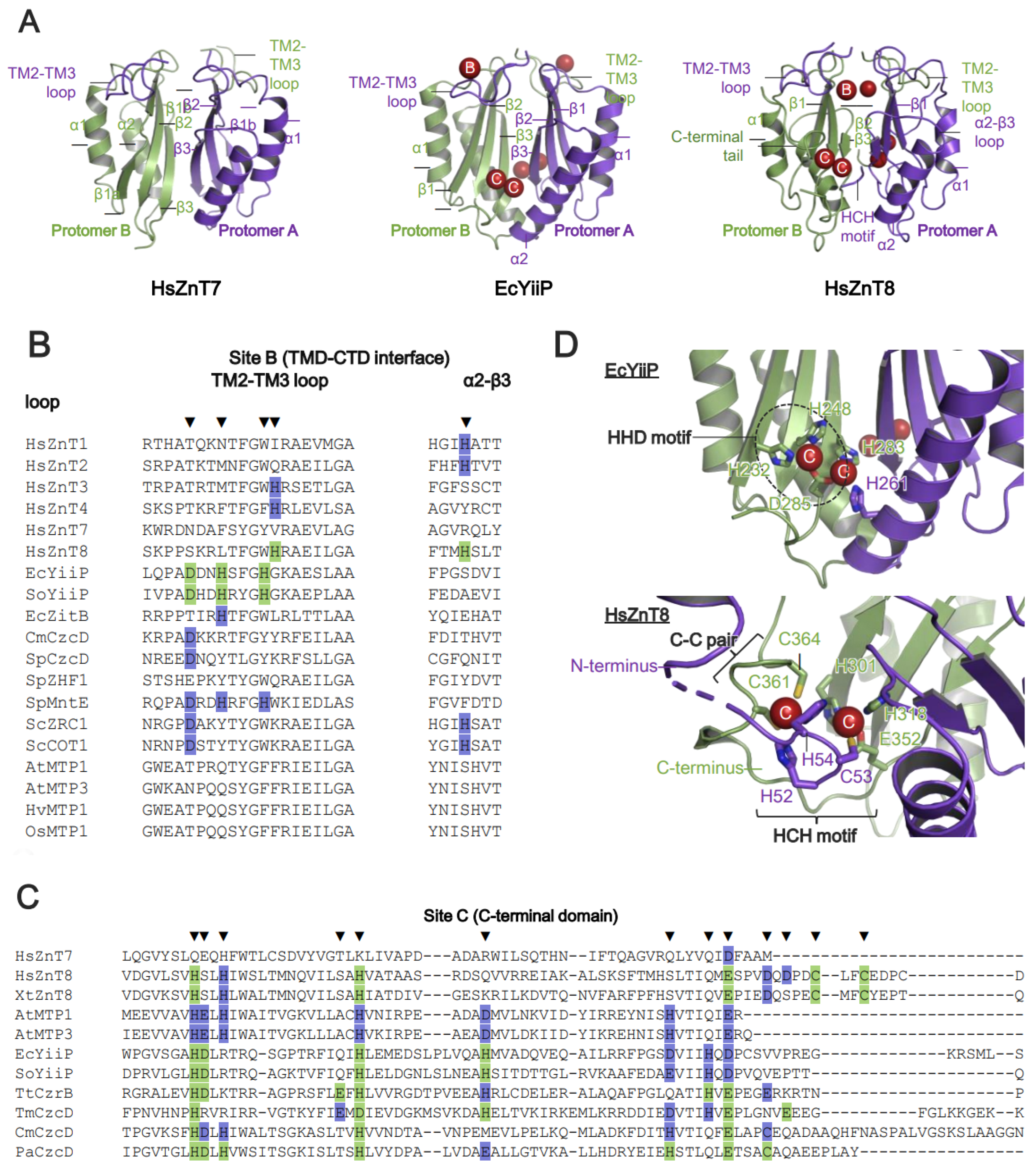
5. Zn2+-Binding Sites on ZnTs
Zn2+-Binding Sites and Metal Ion Selectivity
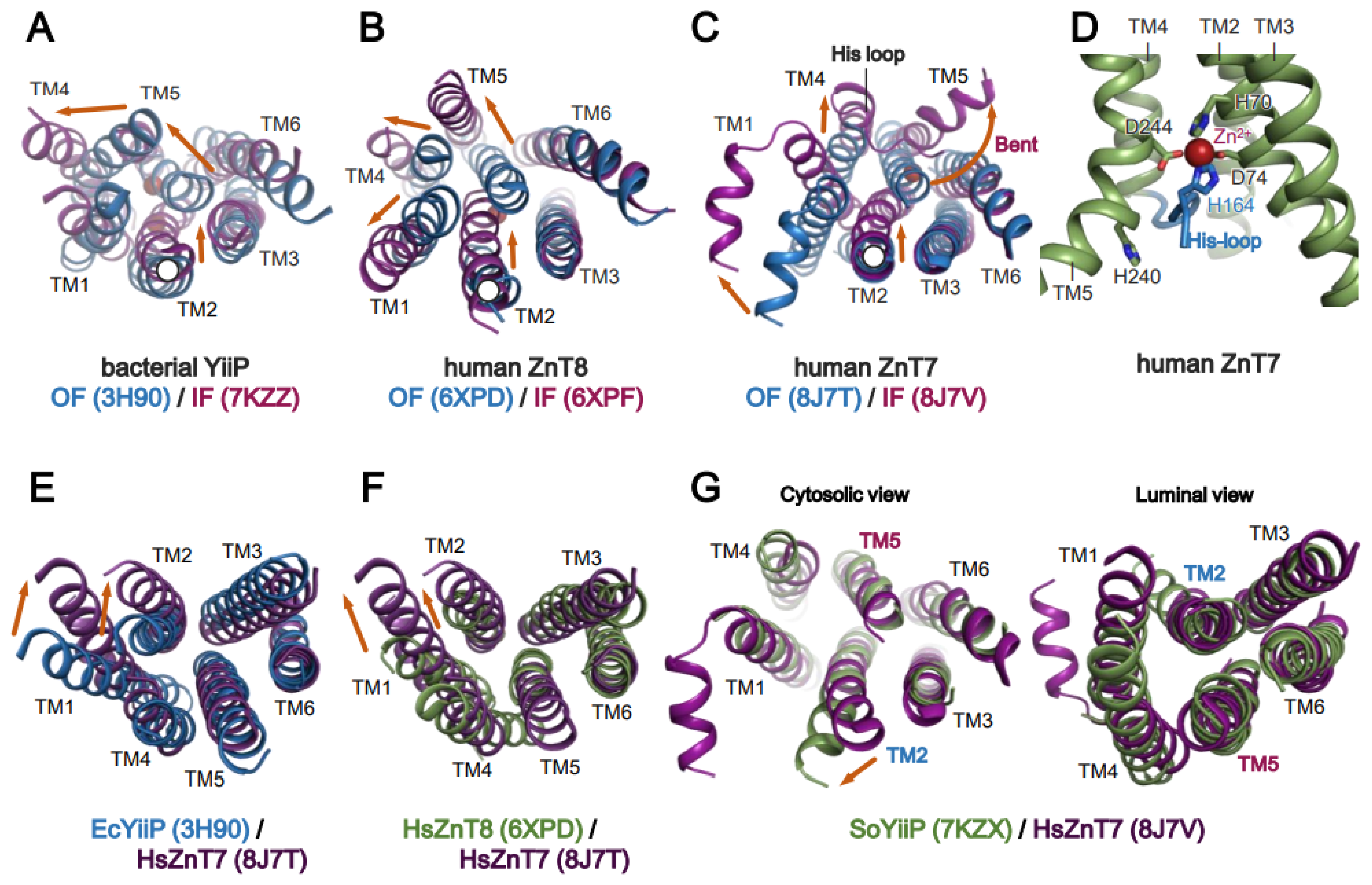
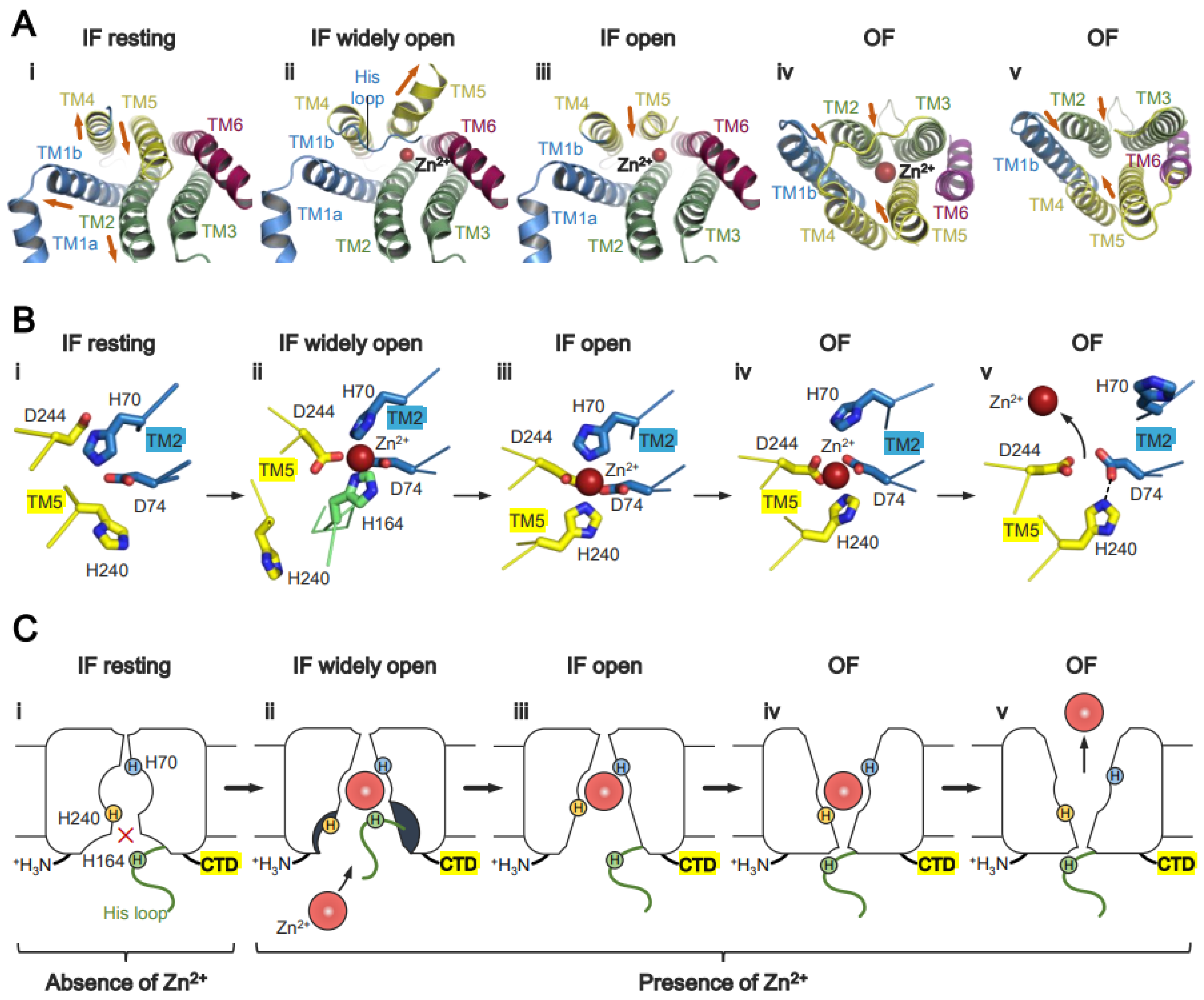
6. Mechanism of Zn2+ Transport by Human ZnT7 and Bacterial YiiP
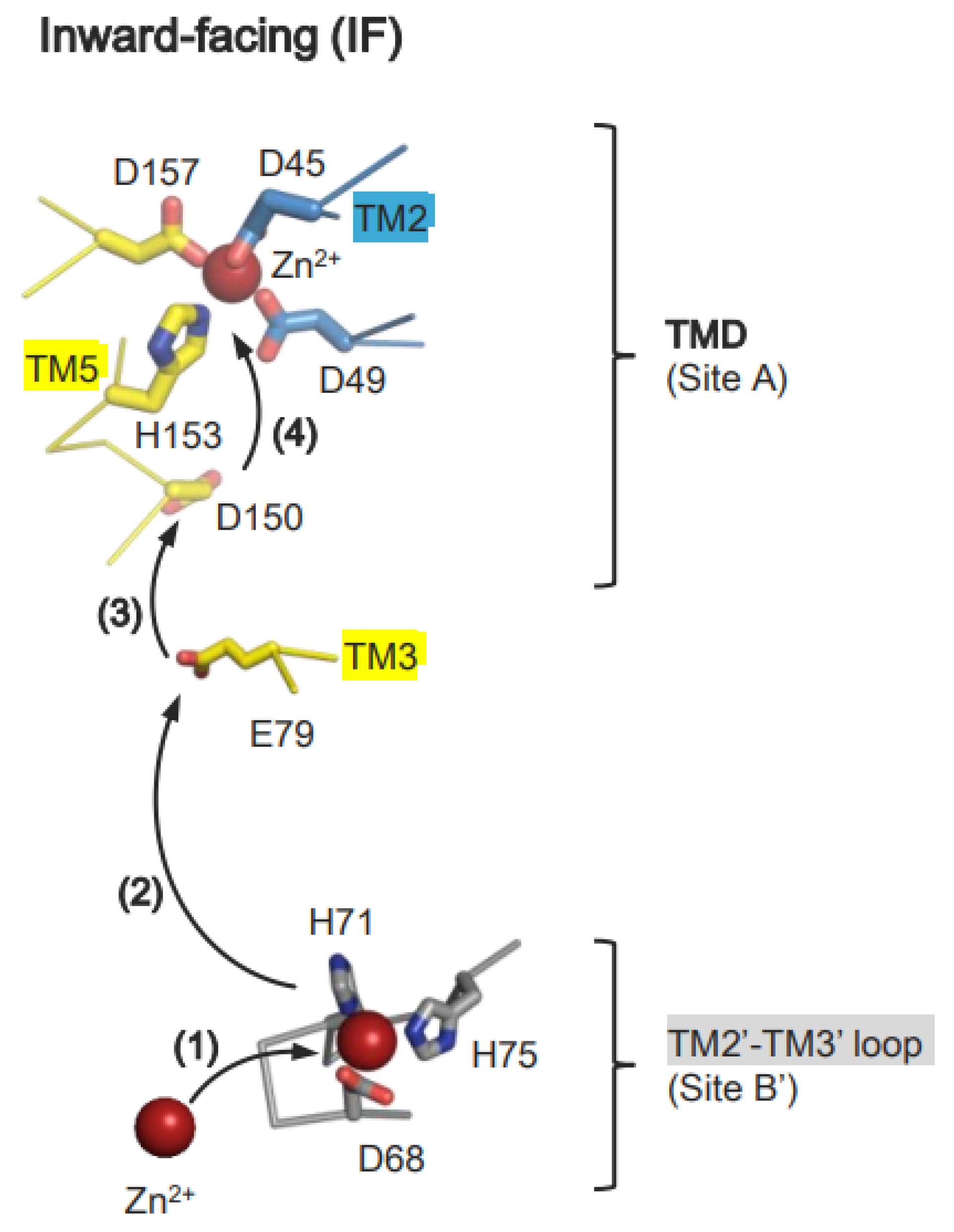
6.1. Zn2+ Transport by HsZnT7
6.2. Zn2+ Transport by Bacterial YiiP
6.3. Role of the Proton Gradient in ZnT-Mediated Zn2+ Transport
7. Structural and Functional Roles of the Histidine-Rich Loop (His-Loop)
8. Zn2+ Transport by Other Zn2+ Transport Systems
8.1. ZIPs
8.2. ZntB
8.3. P-Type ATPases with Zn2+ Transport Activity
8.4. Zn2+ Transport by ATP-Binding Cassette Transporters
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Skalny, A.V.; Aschner, M.; Tinkov, A.A. Zinc. Adv. Food Nutr. Res. 2021, 96, 251–310. [Google Scholar] [CrossRef]
- Zaher, D.M.; El-Gamal, M.I.; Omar, H.A.; Aljareh, S.N.; Al-Shamma, S.A.; Ali, A.J.; Zaib, S.; Iqbal, J. Recent advances with alkaline phosphatase isoenzymes and their inhibitors. Arch. Pharm. 2020, 353, e2000011. [Google Scholar] [CrossRef]
- Xiong, L.; Peng, M.; Zhao, M.; Liang, Z. Truncated expression of a Carboxypeptidase A from bovine improves its enzymatic properties and detoxification efficiency of Ochratoxin A. Toxins 2020, 12, 680. [Google Scholar] [CrossRef]
- Luan, R.; Ding, D.; Xue, Q.; Li, H.; Wang, Y.; Yang, J. Protective role of zinc in the pathogenesis of respiratory diseases. Eur. J. Clin. Nutr. 2023, 77, 427–435. [Google Scholar] [CrossRef]
- Begum, F.; Me, H.M.; Christov, M. The role of zinc in cardiovascular disease. Cardiol. Rev. 2022, 30, 100–108. [Google Scholar] [CrossRef]
- Gembillo, G.; Visconti, L.; Giuffrida, A.E.; Labbozzetta, V.; Peritore, L.; Lipari, A.; Calabrese, V.; Piccoli, G.B.; Torreggiani, M.; Siligato, R.; et al. Role of zinc in diabetic kidney disease. Nutrients 2022, 14, 1353. [Google Scholar] [CrossRef]
- Li, J.; Cao, D.; Huang, Y.; Chen, B.; Chen, Z.; Wang, R.; Dong, Q.; Wei, Q.; Liu, L. Zinc intakes and health outcomes: An umbrella review. Front. Nutr. 2022, 9, 798078. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Y.; Wei, R.; Yong, V.W.; Xue, M. The important role of zinc in neurological diseases. Biomolecules 2022, 13, 28. [Google Scholar] [CrossRef]
- Garner, T.B.; Hester, J.M.; Carothers, A.; Diaz, F.J. Role of zinc in female reproduction. Biol. Reprod. 2021, 104, 976–994. [Google Scholar] [CrossRef] [PubMed]
- Banupriya, N.; Bhat, B.V.; Sridhar, M.G. Role of zinc in neonatal sepsis. Indian J. Pediatr. 2021, 88, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Tamura, Y. The role of zinc homeostasis in the prevention of diabetes mellitus and cardiovascular diseases. J. Atheroscler. Thromb. 2021, 28, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Allouche-Fitoussi, D.; Breitbart, H. The role of zinc in male fertility. Int. J. Mol. Sci. 2020, 21, 7796. [Google Scholar] [CrossRef]
- Michalczyk, K.; Cymbaluk-Płoska, A. The role of zinc and copper in gynecological malignancies. Nutrients 2020, 12, 3732. [Google Scholar] [CrossRef] [PubMed]
- Granero, R.; Pardo-Garrido, A.; Carpio-Toro, I.L.; Ramírez-Coronel, A.A.; Martínez-Suárez, P.C.; Reivan-Ortiz, G.G. The role of iron and zinc in the treatment of ADHD among children and adolescents: A systematic review of randomized clinical trials. Nutrients 2021, 13, 4059. [Google Scholar] [CrossRef]
- Moshtagh, M.; Amiri, R. Role of zinc supplementation in the improvement of acute respiratory infections among Iranian children: A systematic review. Tanaffos 2020, 19, 1–9. [Google Scholar]
- Wang, M.X.; Win, S.S.; Pang, J. Zinc supplementation reduces common cold duration among healthy adults: A systematic review of randomized controlled trials with micronutrients supplementation. Am. J. Trop. Med. Hyg. 2020, 103, 86–99. [Google Scholar] [CrossRef]
- Korkmaz-Icöz, S.; Atmanli, A.; Radovits, T.; Li, S.; Hegedüs, P.; Ruppert, M.; Brlecic, P.; Yoshikawa, Y.; Yasui, H.; Karck, M.; et al. Administration of zinc complex of acetylsalicylic acid after the onset of myocardial injury protects the heart by upregulation of antioxidant enzymes. J. Physiol. Sci. 2016, 66, 113–125. [Google Scholar] [CrossRef]
- Barnett, J.B.; Dao, M.C.; Hamer, D.H.; Kandel, R.; Brandeis, G.; Wu, D.; Dallal, G.E.; Jacques, P.F.; Schreiber, R.; Kong, E.; et al. Effect of zinc supplementation on serum zinc concentration and T cell proliferation in nursing home elderly: A randomized, double-blind, placebo-controlled trial. Am. J. Clin. Nutr. 2016, 103, 942–951. [Google Scholar] [CrossRef]
- Cope, E.C.; Levenson, C.W. Role of zinc in the development and treatment of mood disorders. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 685–689. [Google Scholar] [CrossRef]
- Plum, L.M.; Rink, L.; Haase, H. The essential toxin: Impact of zinc on human health. Int. J. Environ. Res. Public Health 2010, 7, 1342–1365. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.R.S. Zinc Excess. In Zinc in Human Biology. ILSI Human Nutrition Reviews; Mills, C.F., Ed.; Springer: London, UK, 1989; pp. 365–370. [Google Scholar] [CrossRef]
- Brown, M.A.; Thom, J.V.; Orth, G.L.; Cova, P.; Juarez, J. Food poisoning involving zinc contamination. Arch. Environ. Health 1964, 8, 657–660. [Google Scholar] [CrossRef]
- Skalny, A.V.; Aschner, M.; Lei, X.G.; Gritsenko, V.A.; Santamaria, A.; Alekseenko, S.I.; Prakash, N.T.; Chang, J.S.; Sizova, E.A.; Chao, J.C.J.; et al. Gut Microbiota as a Mediator of Essential and Toxic Effects of Zinc in the Intestines and Other Tissues. Int. J. Mol. Sci. 2021, 22, 13074. [Google Scholar] [CrossRef] [PubMed]
- Hamzah-Saleem, M.; Usman, K.; Rizwan, M.; Al-Jabri, H.; Alsafran, M. Functions and strategies for enhancing zinc availability in plants for sustainable agriculture. Front. Plant Sci. 2022, 13, 1033092. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, R.; Srivastava, S. Zinc resistance mechanisms in bacteria. Curr. Sci. 2001, 81, 768–775. [Google Scholar]
- Pandey, N.; Pathak, G.C.; Sharma, C.P. Zinc is critically required for pollen function and fertilisation in lentil. J. Trace Elem. Med. Biol. 2006, 20, 89–96. [Google Scholar] [CrossRef]
- Wissuwa, M.; Ismail, A.M.; Yanagihara, S. Effects of zinc deficiency on rice growth and genetic factors contributing to tolerance. Plant Physiol. 2006, 142, 731–741. [Google Scholar] [CrossRef]
- Corbett, D.; Wang, J.; Schuler, S.; Lopez-Castejon, G.; Glenn, S.; Brough, D.; Andrew, P.W.; Cavet, J.S.; Roberts, I.S. Two zinc uptake systems contribute to the full virulence of Listeria monocytogenes during growth in vitro and in vivo. Infect. Immun. 2012, 80, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Yoshigai, E.; Ohashi, T.; Fukada, T. Zinc transporters as potential therapeutic targets: An updated review. J. Pharmacol. Sci. 2022, 148, 221–228. [Google Scholar] [CrossRef]
- Zhu, B.; Huo, R.; Zhi, Q.; Zhan, M.; Chen, X.; Hua, Z.C. Increased expression of zinc transporter ZIP4, ZIP11, ZnT1, and ZnT6 predicts poor prognosis in pancreatic cancer. J. Trace Elem. Med. Biol. 2021, 65, 126734. [Google Scholar] [CrossRef]
- Lei, P.; Ayton, S.; Bush, A.I. The essential elements of Alzheimer’s disease. J. Biol. Chem. 2021, 296, 100105. [Google Scholar] [CrossRef]
- Xu, Y.; Xiao, G.; Liu, L.; Lang, M. Zinc transporters in Alzheimer’s disease. Mol. Brain 2019, 12, 106. [Google Scholar] [CrossRef] [PubMed]
- Sikora, J.; Ouagazzal, A.M. Synaptic zinc: An emerging player in Parkinson’s disease. Int. J. Mol. Sci. 2021, 22, 4724. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.N.; Strong, M.D.; Chambers, E.; Hart, M.D.; Bettaieb, A.; Clarke, S.L.; Smith, B.J.; Stoecker, B.J.; Lucas, E.A.; Lin, D.; et al. A role for zinc transporter gene SLC39A12 in the nervous system and beyond. Gene 2021, 799, 145824. [Google Scholar] [CrossRef] [PubMed]
- Chowanadisai, W.; Lönnerdal, B.; Kelleher, S.L. Identification of a mutation in SLC30A2 (ZnT-2) in women with low milk zinc concentration that results in transient neonatal zinc deficiency. J. Biol. Chem. 2006, 281, 39699–39707. [Google Scholar] [CrossRef] [PubMed]
- Lieberwirth, J.K.; Joset, P.; Heinze, A.; Hentschel, J.; Stein, A.; Iannaccone, A.; Steindl, K.; Kuechler, A.; Abou-Jamra, R. Bi-allelic loss of function variants in SLC30A5 as cause of perinatal lethal cardiomyopathy. Eur. J. Hum. Genet. 2021, 29, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, M.S.; Phillips, A.M.; Mullen, S.A.; Adlard, P.A.; Hardies, K.; Damiano, J.A.; Wimmer, V.; Bellows, S.T.; McMahon, J.M.; Burgess, R.; et al. Loss of synaptic Zn2+ transporter function increases risk of febrile seizures. Sci. Rep. 2015, 5, 17816. [Google Scholar] [CrossRef] [PubMed]
- Pérez, Y.; Shorer, Z.; Liani-Leibson, K.; Chabosseau, P.; Kadir, R.; Volodarsky, M.; Halperin, D.; Barber-Zucker, S.; Shalev, H.; Schreiber, R.; et al. SLC30A9 mutation affecting intracellular zinc homeostasis causes a novel cerebro-renal syndrome. Brain 2017, 140, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Tavasoli, A.; Arjmandi-Rafsanjani, K.; Hemmati, S.; Mojbafan, M.; Zarei, E.; Hosseini, S. A case of dystonia with polycythemia and hypermanganesemia caused by SLC30A10 mutation: A treatable inborn error of manganese metabolism. BMC Pediatr. 2019, 19, 229. [Google Scholar] [CrossRef]
- Quadri, M.; Federico, A.; Zhao, T.; Breedveld, G.J.; Battisti, C.; Delnooz, C.; Severijnen, L.A.; Di Toro Mammarella, L.; Mignarri, A.; Monti, L.; et al. Mutations in SLC30A10 cause parkinsonism and dystonia with hypermanganesemia, polycythemia, and chronic liver disease. Am. J. Hum. Genet. 2012, 90, 467–477. [Google Scholar] [CrossRef]
- Lu, M.; Fu, D. Structure of the zinc transporter YiiP. Science 2007, 317, 1746–1748. [Google Scholar] [CrossRef]
- Lu, M.; Chai, J.; Fu, D. Structural basis for autoregulation of the zinc transporter YiiP. Nat. Struct. Mol. Biol. 2009, 16, 1063–1067. [Google Scholar] [CrossRef]
- Hussein, A.; Fan, S.; Lopez-Redondo, M.L.; Kenney, I.; Zhang, X.; Beckstein, O.; Stokes, D.L. Energy coupling and stoichiometry of Zn2+/H+ antiport by the cation diffusion facilitator YiiP. eLife 2023, 12, RP87167. [Google Scholar] [CrossRef]
- Lopez-Redondo, M.L.; Fan, S.; Koide, A.; Koide, S.; Beckstein, O.; Stokes, D.L. Zinc binding alters the conformational dynamics and drives the transport cycle of the cation diffusion facilitator YiiP. J. Gen. Physiol. 2021, 153, e202112873. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Redondo, M.L.; Coudray, N.; Zhang, Z.; Alexopoulos, J.; Stokes, D.L. Structural basis for the alternating access mechanism of the cation diffusion facilitator YiiP. Proc. Natl. Acad. Sci. USA 2018, 115, 3042–3047. [Google Scholar] [CrossRef] [PubMed]
- Coudray, N.; Valvo, S.; Hu, M.; Lasala, R.; Kim, C.; Vink, M.; Zhou, M.; Provasi, D.; Filizola, M.; Tao, J.; et al. Inward-facing conformation of the zinc transporter YiiP revealed by cryoelectron microscopy. Proc. Natl. Acad. Sci. USA 2013, 110, 2140–2145. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.; Fu, D. Kinetic study of the antiport mechanism of an Escherichia coli zinc transporter, ZitB. J. Biol. Chem. 2004, 279, 12043–12050. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.; Fu, D. Thermodynamic studies of the mechanism of metal binding to the Escherichia coli zinc transporter YiiP. J. Biol. Chem. 2004, 279, 17173–17180. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Merz, K.M. Mechanism of zinc transport through the zinc transporter YiiP. J. Chem. Theory Comput. 2022, 18, 2556–2568. [Google Scholar] [CrossRef] [PubMed]
- Bui, H.B.; Watanabe, S.; Nomura, N.; Liu, K.; Uemura, T.; Inoue, M.; Tsutsumi, A.; Fujita, H.; Kinoshita, K.; Kato, Y.; et al. Cryo-EM structures of human zinc transporter ZnT7 reveal the mechanism of Zn2+ uptake into the Golgi apparatus. Nat. Commun. 2023, 14, 4770. [Google Scholar] [CrossRef]
- Xue, J.; Xie, T.; Zeng, W.; Jiang, Y.; Bai, X.C. Cryo-EM structures of human ZnT8 in both outward- and inward-facing conformations. eLife 2020, 9, e58823. [Google Scholar] [CrossRef]
- Zhang, S.; Fu, C.; Luo, Y.; Xie, Q.; Xu, T.; Sun, Z.; Su, Z.; Zhou, X. Cryo-EM structure of a eukaryotic zinc transporter at a low pH suggests its Zn2+-releasing mechanism. J. Struct. Biol. 2023, 215, 107926. [Google Scholar] [CrossRef] [PubMed]
- Gati, C.; Stetsenko, A.; Slotboom, D.J.; Scheres, S.H.W.; Guskov, A. The structural basis of proton driven zinc transport by ZntB. Nat. Commun. 2017, 8, 1313. [Google Scholar] [CrossRef] [PubMed]
- Golan, Y.; Alhadeff, R.; Warshel, A.; Assaraf, Y.G. ZnT2 is an electroneutral proton-coupled vesicular antiporter displaying an apparent stoichiometry of two protons per zinc ion. PLoS Comput. Biol. 2019, 15, e1006882. [Google Scholar] [CrossRef] [PubMed]
- Cotrim, C.A.; Jarrott, R.J.; Martin, J.L.; Drew, D. A structural overview of the zinc transporters in the cation diffusion facilitator family. Acta Crystallogr. D Struct. Biol. 2019, 75, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Pang, C.; Chai, J.; Zhu, P.; Shanklin, J.; Liu, Q. Structural mechanism of intracellular autoregulation of zinc uptake in ZIP transporters. Nat. Commun. 2023, 14, 3404. [Google Scholar] [CrossRef]
- Montanini, B.; Blaudez, D.; Jeandroz, S.; Sanders, D.; Chalot, M. Phylogenetic and functional analysis of the Cation Diffusion Facilitator (CDF) family: Improved signature and prediction of substrate specificity. BMC Genom. 2007, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Gaither, L.A.; Eide, D.J. Eukaryotic zinc transporters and their regulation. Biometals 2001, 14, 251–270. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T.; Taylor, K.M.; Fu, D. Zinc transporters and their functional integration in mammalian cells. J. Biol. Chem. 2021, 296, 100320. [Google Scholar] [CrossRef]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The physiological, biochemical, and molecular roles of zinc transporters in zinc homeostasis and metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef]
- Hara, T.; Takeda, T.A.; Takagishi, T.; Fukue, K.; Kambe, T.; Fukada, T. Physiological roles of zinc transporters: Molecular and genetic importance in zinc homeostasis. J. Physiol. Sci. 2017, 67, 283–301. [Google Scholar] [CrossRef]
- Fukunaka, A.; Suzuki, T.; Kurokawa, Y.; Yamazaki, T.; Fujiwara, N.; Ishihara, K.; Migaki, H.; Okumura, K.; Masuda, S.; Yamaguchi-Iwai, Y.; et al. Demonstration and characterization of the heterodimerization of ZnT5 and ZnT6 in the early secretory pathway. J. Biol. Chem. 2009, 284, 30798–30806. [Google Scholar] [CrossRef]
- Huang, L.; Tepaamorndech, S. The SLC30 family of zinc transporters: A review of current understanding of their biological and pathophysiological roles. Mol. Asp. Med. 2013, 34, 548–560. [Google Scholar] [CrossRef]
- Amagai, Y.; Yamada, M.; Kowada, T.; Watanabe, T.; Du, Y.; Liu, R.; Naramoto, S.; Watanabe, S.; Kyozuka, J.; Anelli, T.; et al. Zinc homeostasis governed by Golgi-resident ZnT family members regulates ERp44-mediated proteostasis at the ER-Golgi interface. Nat. Commun. 2023, 14, 2683. [Google Scholar] [CrossRef]
- Kambe, T.; Wagatsuma, T. Metalation and activation of Zn2+ enzymes via early secretory pathway-resident ZNT proteins. Biophys. Rev. 2023, 4, 041302. [Google Scholar] [CrossRef]
- Ricachenevsky, F.K.; Menguer, P.K.; Sperotto, R.A.; Williams, L.E.; Fett, J.P. Roles of plant metal tolerance proteins (MTP) in metal storage and potential use in biofortification strategies. Front. Plant Sci. 2013, 4, 144. [Google Scholar] [CrossRef]
- Gustin, J.L.; Zanis, M.J.; Salt, D.E. Structure and evolution of the plant cation diffusion facilitator family of ion transporters. BMC Evol. Biol. 2011, 11, 76. [Google Scholar] [CrossRef]
- Gao, Y.; Yang, F.; Liu, J.; Xie, W.; Zhang, L.; Chen, Z.; Peng, Z.; Ou, Y.; Yao, Y. Genome-wide identification of metal tolerance protein genes in Populus trichocarpa and their roles in response to various heavy metal stresses. Int. J. Mol. Sci. 2020, 21, 1680. [Google Scholar] [CrossRef]
- Arrivault, S.; Senger, T.; Krämer, U. The Arabidopsis metal tolerance protein AtMTP3 maintains metal homeostasis by mediating Zn exclusion from the shoot under Fe deficiency and Zn oversupply. Plant J. 2006, 46, 861–879. [Google Scholar] [CrossRef] [PubMed]
- Desbrosses-Fonrouge, A.G.; Voigt, K.; Schröder, A.; Arrivault, S.; Thomine, S.; Krämer, U. Arabidopsis thaliana MTP1 is a Zn transporter in the vacuolar membrane which mediates Zn detoxification and drives leaf Zn accumulation. FEBS Lett. 2005, 579, 4165–4174. [Google Scholar] [CrossRef] [PubMed]
- Kobae, Y.; Uemura, T.; Sato, M.H.; Ohnishi, M.; Mimura, T.; Nakagawa, T.; Maeshima, M. Zinc transporter of Arabidopsis thaliana AtMTP1 is localized to vacuolar membranes and implicated in zinc homeostasis. Plant Cell Physiol. 2004, 45, 1749–1758. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, S.A.; Krämer, U. The zinc homeostasis network of land plants. Biochim. Biophys. Acta 2012, 1823, 1553–1567. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Kawachi, M.; Sato, Y.; Mori, H.; Kutsuna, N.; Hasezawa, S.; Maeshima, M. A high molecular mass zinc transporter MTP12 forms a functional heteromeric complex with MTP5 in the Golgi in Arabidopsis thaliana. FEBS J. 2015, 282, 1965–1979. [Google Scholar] [CrossRef]
- Miyabe, S.; Izawa, S.; Inoue, Y. The Zrc1 is involved in zinc transport system between vacuole and cytosol in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2001, 282, 79–83. [Google Scholar] [CrossRef]
- MacDiarmid, C.W.; Milanick, M.A.; Eide, D.J. Biochemical properties of vacuolar zinc transport systems of Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 39187–39194. [Google Scholar] [CrossRef]
- MacDiarmid, C.W.; Milanick, M.A.; Eide, D.J. Induction of the ZRC1 metal tolerance gene in zinc-limited yeast confers resistance to zinc shock. J. Biol. Chem. 2003, 278, 15065–15072. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Kumánovics, A.; Nelson, J.M.; Warner, D.E.; Ward, D.M.; Kaplan, J. A single amino acid change in the yeast vacuolar metal transporters ZRC1 and COT1 alters their substrate specificity. J. Biol. Chem. 2008, 283, 33865–33873. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Kaplan, J. The yeast gene MSC2, a member of the cation diffusion facilitator family, affects the cellular distribution of zinc. J. Biol. Chem. 2001, 276, 5036–5043. [Google Scholar] [CrossRef]
- Wu, Y.H.; Taggart, J.; Song, P.X.; MacDiarmid, C.; Eide, D.J. An MSC2 promoter-lacZ fusion gene reveals zinc-responsive changes in sites of transcription initiation that occur across the yeast genome. PLoS ONE 2016, 11, e0163256. [Google Scholar] [CrossRef]
- Ellis, C.D.; Macdiarmid, C.W.; Eide, D.J. Heteromeric protein complexes mediate zinc transport into the secretory pathway of eukaryotic cells. J. Biol. Chem. 2005, 280, 28811–28818. [Google Scholar] [CrossRef]
- Ellis, C.D.; Wang, F.; MacDiarmid, C.W.; Clark, S.; Lyons, T.; Eide, D.J. Zinc and the Msc2 zinc transporter protein are required for endoplasmic reticulum function. J. Cell Biol. 2004, 166, 325–335. [Google Scholar] [CrossRef]
- Clemens, S.; Bloss, T.; Vess, C.; Neumann, D.; Nies, D.H.; Zur Nieden, U. A transporter in the endoplasmic reticulum of Schizosaccharomyces pombe cells mediates zinc storage and differentially affects transition metal tolerance. J. Biol. Chem. 2002, 277, 18215–18221. [Google Scholar] [CrossRef] [PubMed]
- Grass, G.; Otto, M.; Fricke, B.; Haney, C.J.; Rensing, C.; Nies, D.H.; Munkelt, D. FieF (YiiP) from Escherichia coli mediates decreased cellular accumulation of iron and relieves iron stress. Arch. Microbiol. 2005, 183, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Fu, D. Selective metal binding to a membrane-embedded aspartate in the Escherichia coli metal transporter YiiP (FieF). J. Biol. Chem. 2005, 280, 33716–33724. [Google Scholar] [CrossRef] [PubMed]
- Grass, G.; Fan, B.; Rosen, B.P.; Franke, S.; Nies, D.H.; Rensing, C. ZitB (YbgR), a member of the cation diffusion facilitator family, is an additional zinc transporter in Escherichia coli. J. Bacteriol. 2001, 183, 4664–4667. [Google Scholar] [CrossRef] [PubMed]
- Guffanti, A.A.; Wei, Y.; Rood, S.V.; Krulwich, T.A. An antiport mechanism for a member of the cation diffusion facilitator family: Divalent cations efflux in exchange for K+ and H+. Mol. Microbiol. 2002, 45, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Garaeva, A.A.; Slotboom, D.J. Elevator-type mechanisms of membrane transport. Biochem. Soc. Trans. 2020, 48, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Nishito, Y.; Tsuji, N.; Fujishiro, H.; Takeda, T.A.; Yamazaki, T.; Teranishi, F.; Okazaki, F.; Matsunaga, A.; Tuschl, K.; Rao, R.; et al. Direct comparison of manganese detoxification/efflux proteins and molecular characterization of ZnT10 protein as a manganese transporter. J. Biol. Chem. 2016, 291, 14773–14787. [Google Scholar] [CrossRef]
- Zogzas, C.E.; Aschner, M.; Mukhopadhyay, S. Structural elements in the transmembrane and cytoplasmic domains of the metal transporter SLC30A10 are required for its manganese efflux activity. J. Biol. Chem. 2016, 291, 15940–15957. [Google Scholar] [CrossRef]
- Leyva-Illades, D.; Chen, P.; Zogzas, C.E.; Hutchens, S.; Mercado, J.M.; Swaim, C.D.; Morrisett, R.A.; Bowman, A.B.; Aschner, M.; Mukhopadhyay, S. SLC30A10 is a cell surface-localized manganese efflux transporter, and parkinsonism-causing mutations block its intracellular trafficking and efflux activity. J. Neurosci. 2014, 34, 14079–14095. [Google Scholar] [CrossRef]
- Kawachi, M.; Kobae, Y.; Kogawa, S.; Mimura, T.; Krämer, U.; Maeshima, M. Amino acid screening based on structural modeling identifies critical residues for the function, ion selectivity and structure of Arabidopsis MTP1. FEBS J. 2012, 279, 2339–2356. [Google Scholar] [CrossRef]
- Sala, D.; Giachetti, A.; Rosato, A. An atomistic view of the YiiP structural changes upon zinc(II) binding. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 1560–1567. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Chai, J.; Cheng, J.; D’Mello, R.; Chance, M.R.; Fu, D. Visualizing the kinetic power stroke that drives proton-coupled zinc(II) transport. Nature 2014, 512, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Kaback, H.R. A chemiosmotic mechanism of symport. Proc. Natl. Acad. Sci. USA 2015, 112, 1259–1264. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Kowada, T.; Du, Y.; Amagai, Y.; Matsui, T.; Inaba, K.; Mizukami, S. Organelle-level labile Zn2+ mapping based on targetable fluorescent sensors. ACS Sens. 2022, 7, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Kowada, T.; Watanabe, T.; Amagai, Y.; Liu, R.; Yamada, M.; Takahashi, H.; Matsui, T.; Inaba, K.; Mizukami, S. Quantitative imaging of labile Zn2+ in the Golgi apparatus using a localizable small-molecule fluorescent probe. Cell Chem. Biol. 2020, 27, 1521–1531.e8. [Google Scholar] [CrossRef]
- Maeda, Y.; Ide, T.; Koike, M.; Uchiyama, Y.; Kinoshita, T. GPHR is a novel anion channel critical for acidification and functions of the Golgi apparatus. Nat. Cell Biol. 2008, 10, 1135–1145. [Google Scholar] [CrossRef]
- Kawachi, M.; Kobae, Y.; Mimura, T.; Maeshima, M. Deletion of a histidine-rich loop of AtMTP1, a vacuolar Zn2+/H+ antiporter of Arabidopsis thaliana, stimulates the transport activity. J. Biol. Chem. 2008, 283, 8374–8383. [Google Scholar] [CrossRef]
- Podar, D.; Scherer, J.; Noordally, Z.; Herzyk, P.; Nies, D.; Sanders, D. Metal selectivity determinants in a family of transition metal transporters. J. Biol. Chem. 2012, 287, 3185–3196. [Google Scholar] [CrossRef]
- Tanaka, N.; Fujiwara, T.; Tomioka, R.; Krämer, U.; Kawachi, M.; Maeshima, M. Characterization of the histidine-rich loop of Arabidopsis vacuolar membrane zinc transporter AtMTP1 as a sensor of zinc level in the cytosol. Plant Cell Physiol. 2015, 56, 510–519. [Google Scholar] [CrossRef]
- Tanaka, N.; Kawachi, M.; Fujiwara, T.; Maeshima, M. Zinc-binding and structural properties of the histidine-rich loop of Arabidopsis thaliana vacuolar membrane zinc transporter MTP1. FEBS Open Bio 2013, 3, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Eide, D.J. The SLC39 family of zinc transporters. Mol. Asp. Med. 2013, 34, 612–619. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, Y.; Gao, K.; Sui, D.; Yu, P.; Su, M.; Wei, G.W.; Hu, J. Structural insights into the elevator-type transport mechanism of a bacterial ZIP metal transporter. Nat. Commun. 2023, 14, 385. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, J.; Fellner, M.; Zhang, C.; Sui, D.; Hu, J. Crystal structures of a ZIP zinc transporter reveal a binuclear metal center in the transport pathway. Sci. Adv. 2017, 3, e1700344. [Google Scholar] [CrossRef]
- Hoch, E.; Levy, M.; Hershfinkel, M.; Sekler, I. Elucidating the H+ coupled Zn2+ transport mechanism of ZIP4: Implications in acrodermatitis enteropathica. Int. J. Mol. Sci. 2020, 21, 734. [Google Scholar] [CrossRef] [PubMed]
- Franz, M.C.; Pujol-Giménez, J.; Montalbetti, N.; Fernandez-Tenorio, M.; DeGrado, T.R.; Niggli, E.; Romero, M.F.; Hediger, M.A. Reassessment of the transport mechanism of the human zinc transporter SLC39A2. Biochemistry 2018, 57, 3976–3986. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Chai, J.; Love, J.; Fu, D. Selective electrodiffusion of zinc ions in a Zrt-, Irt-like protein, ZIPB. J. Biol. Chem. 2010, 285, 39013–39020. [Google Scholar] [CrossRef]
- Zhang, T.; Sui, D.; Zhang, C.; Cole, L.; Hu, J. Asymmetric functions of a binuclear metal center within the transport pathway of a human zinc transporter ZIP4. FASEB J. 2020, 34, 237–247. [Google Scholar] [CrossRef]
- Mao, X.; Kim, B.E.; Wang, F.; Eide, D.J.; Petris, M.J. A histidine-rich cluster mediates the ubiquitination and degradation of the human zinc transporter, hZIP4, and protects against zinc cytotoxicity. J. Biol. Chem. 2007, 282, 6992–7000. [Google Scholar] [CrossRef] [PubMed]
- Ajeesh-Krishna, T.P.; Maharajan, T.; Victor-Roch, G.; Ignacimuthu, S.; Antony Ceasar, S. Structure, function, regulation and phylogenetic relationship of ZIP family transporters of plants. Front. Plant Sci. 2020, 11, 662. [Google Scholar] [CrossRef]
- Tan, K.; Sather, A.; Robertson, J.L.; Moy, S.; Roux, B.; Joachimiak, A. Structure and electrostatic property of cytoplasmic domain of ZntB transporter. Protein Sci. 2009, 18, 2043–2052. [Google Scholar] [CrossRef]
- Worlock, A.J.; Smith, R.L. ZntB is a novel Zn2+ transporter in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2002, 184, 4369–4373. [Google Scholar] [CrossRef]
- Stetsenko, A.; Stehantsev, P.; Dranenko, N.O.; Gelfand, M.S.; Guskov, A. Structural and biochemical characterization of a novel ZntB (CmaX) transporter protein from Pseudomonas aeruginosa. Int. J. Biol. Macromol. 2021, 184, 760–767. [Google Scholar] [CrossRef]
- Pfoh, R.; Li, A.; Chakrabarti, N.; Payandeh, J.; Pomès, R.; Pai, E.F. Structural asymmetry in the magnesium channel CorA points to sequential allosteric regulation. Proc. Natl. Acad. Sci. USA 2012, 109, 18809–18814. [Google Scholar] [CrossRef]
- Wang, D.; Hurst, T.K.; Thompson, R.B.; Fierke, C.A. Genetically encoded ratiometric biosensors to measure intracellular exchangeable zinc in Escherichia coli. J. Biomed. Opt. 2011, 16, 087011. [Google Scholar] [CrossRef]
- Mills, R.F.; Francini, A.; Ferreira da Rocha, P.S.; Baccarini, P.J.; Aylett, M.; Krijger, G.C.; Williams, L.E. The plant P1B-type ATPase AtHMA4 transports Zn and Cd and plays a role in detoxification of transition metals supplied at elevated levels. FEBS Lett. 2005, 579, 783–791. [Google Scholar] [CrossRef]
- Wang, K.; Sitsel, O.; Meloni, G.; Autzen, H.E.; Andersson, M.; Klymchuk, T.; Nielsen, A.M.; Rees, D.C.; Nissen, P.; Gourdon, P. Structure and mechanism of Zn2+-transporting P-type ATPases. Nature 2014, 514, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Dyla, M.; Kjærgaard, M.; Poulsen, H.; Nissen, P. Structure and mechanism of P-type ATPase ion pumps. Annu. Rev. Biochem. 2020, 89, 583–603. [Google Scholar] [CrossRef]
- Raimunda, D.; Subramanian, P.; Stemmler, T.; Argüello, J.M. A tetrahedral coordination of zinc during transmembrane transport by P-type Zn2+-ATPases. Biochim. Biophys. Acta 2012, 1818, 1374–1377. [Google Scholar] [CrossRef]
- Kühlbrandt, W. Biology, structure and mechanism of P-type ATPases. Nat. Rev. Mol. Cell Biol. 2004, 5, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Albers, R.W. Biochemical aspects of active transport. Annu. Rev. Biochem. 1967, 36, 727–756. [Google Scholar] [CrossRef] [PubMed]
- Argüello, J.M.; Eren, E.; González-Guerrero, M. The structure and function of heavy metal transport P1B-ATPases. Biometals 2007, 20, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef]
- Yatsunyk, L.A.; Easton, J.A.; Kim, L.R.; Sugarbaker, S.A.; Bennett, B.; Breece, R.M.; Vorontsov, I.I.; Tierney, D.L.; Crowder, M.W.; Rosenzweig, A.C. Structure and metal binding properties of ZnuA, a periplasmic zinc transporter from Escherichia coli. J. Biol. Inorg. Chem. 2008, 13, 271–288. [Google Scholar] [CrossRef]
- Patzer, S.I.; Hantke, K. The ZnuABC high-affinity zinc uptake system and its regulator Zur in Escherichia coli. Mol. Microbiol. 1998, 28, 1199–1210. [Google Scholar] [CrossRef] [PubMed]
- Gabbianelli, R.; Scotti, R.; Ammendola, S.; Petrarca, P.; Nicolini, L.; Battistoni, A. Role of ZnuABC and ZinT in Escherichia coli O157:H7 zinc acquisition and interaction with epithelial cells. BMC Microbiol. 2001, 11, 36. [Google Scholar] [CrossRef]
- Li, H.; Jogl, G. Crystal structure of the zinc-binding transport protein ZnuA from Escherichia coli reveals an unexpected variation in metal coordination. J. Mol. Biol. 2007, 368, 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Chandra, B.R.; Yogavel, M.; Sharma, A. Structural analysis of ABC-family periplasmic zinc binding protein provides new insights into mechanism of ligand uptake and release. J. Mol. Biol. 2007, 367, 970–982. [Google Scholar] [CrossRef]
- Ilari, A.; Alaleona, F.; Petrarca, P.; Battistoni, A.; Chiancone, E. The X-ray structure of the zinc transporter ZnuA from Salmonella enterica discloses a unique triad of zinc-coordinating histidines. J. Mol. Biol. 2011, 409, 630–641. [Google Scholar] [CrossRef]
- Penon-Portmann, M.; Eldomery, M.K.; Potocki, L.; Marafi, D.; Posey, J.E.; Coban-Akdemir, Z.; Harel, T.; Grochowski, C.M.; Loucks, H.; Devine, W.P.; et al. De novo heterozygous variants in SLC30A7 are a candidate cause for Joubert syndrome. Am. J. Med. Genet. A 2022, 188, 2360–2366. [Google Scholar] [CrossRef]
- Watanabe, S.; Amagai, Y.; Sannino, S.; Tempio, T.; Anelli, T.; Harayama, M.; Masui, S.; Sorrentino, I.; Yamada, M.; Sitia, R.; et al. Zinc regulates ERp44-dependent protein quality control in the early secretory pathway. Nat. Commun. 2009, 10, 603. [Google Scholar] [CrossRef]
- Fan, Y.G.; Wu, T.Y.; Zhao, L.X.; Jia, R.J.; Ren, H.; Hou, W.J.; Wang, Z.Y. From zinc homeostasis to disease progression: Unveiling the neurodegenerative puzzle. Pharmacol. Res. 2023, 18, 107039. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Chen, H. Aberrance of zinc metalloenzymes-induced human diseases and its potential mechanisms. Nutrients 2021, 13, 4456. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T.; Matsunaga, M.; Takeda, T.A. Understanding the contribution of zinc transporters in the function of the early secretory pathway. Int. J. Mol. Sci. 2017, 18, 2179. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Hirano, T. Intracellular zinc homeostasis and zinc signaling. Cancer Sci. 2008, 99, 1515–1522. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | Main Functions | Organisms | States | Conformations (PDB Code) | Ligands | Methods | References |
|---|---|---|---|---|---|---|---|
| YiiP | Transport Zn2+ out of the cytoplasm and into the periplasm | Escherichia coli | Homodimer | Outward-facing (2QFI, 3H90) | Zn2+ | X-ray diffraction | [41,42] |
| Shewanella oneidensis | Homodimer | Inward-facing (3J1Z, 5VRF, 7KZZ (1)) | Zn2+ | Electron microscopy | [44,45,46] | ||
| Homodimer | Inward-facing occluded (7KZX) | Zn2+ | [43] | ||||
| ZnT7 | Transport Zn2+ out of the cytoplasm and into the Golgi lumen | Homo sapiens | Homodimer | Outward-facing (8J7T) | Apo | Electron microscopy | [50] |
| Homodimer | Outward-facing (8J7U) | Zn2+ | |||||
| Heterodimer | Inward-facing and outward-facing (8J7V (2)) | Apo | |||||
| Heterodimer | Inward-facing with Zn2+ and outward-facing (8J80 (3)) | Zn2+, Apo | |||||
| Heterodimer | Inward-facing with Zn2+ and outward-facing with Zn2+ (8J7W) (4) | Zn2+ | |||||
| ZnT8 | Transport Zn2+ out of the cytoplasm and into the insulin secretory granule | H. sapiens | Homodimer | Outward-facing (6XPE) | Zn2+ | Electron microscopy | [51] |
| Heterodimer | Outward-facing and inward-facing (6XPF) | Apo | |||||
| Xenopus tropicalis | Homodimer | Outward-facing (7Y5G) | Zn2+ | [52] | |||
| Homodimer | Outward-facing (7Y5H (5)) | Apo |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bui, H.B.; Inaba, K. Structures, Mechanisms, and Physiological Functions of Zinc Transporters in Different Biological Kingdoms. Int. J. Mol. Sci. 2024, 25, 3045. https://doi.org/10.3390/ijms25053045
Bui HB, Inaba K. Structures, Mechanisms, and Physiological Functions of Zinc Transporters in Different Biological Kingdoms. International Journal of Molecular Sciences. 2024; 25(5):3045. https://doi.org/10.3390/ijms25053045
Chicago/Turabian StyleBui, Han Ba, and Kenji Inaba. 2024. "Structures, Mechanisms, and Physiological Functions of Zinc Transporters in Different Biological Kingdoms" International Journal of Molecular Sciences 25, no. 5: 3045. https://doi.org/10.3390/ijms25053045
APA StyleBui, H. B., & Inaba, K. (2024). Structures, Mechanisms, and Physiological Functions of Zinc Transporters in Different Biological Kingdoms. International Journal of Molecular Sciences, 25(5), 3045. https://doi.org/10.3390/ijms25053045