
Nitric Oxide Regulates Seed Germination by Integrating Multiple Signalling Pathways
by
 ,
,
Yue Zhang
1,
Ruirui Wang
1,
Xiaodong Wang
1,
Caihong Zhao
1,
Hailong Shen
1,2,* and
Ling Yang
1,* 1
State Key Laboratory of Tree Genetics and Breeding, School of Forestry, Northeast Forestry University, Harbin 150040, China
2
Research Center of Korean Pine Engineering and Technology, National Forestry and Grassland Administration, Harbin 150040, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(10), 9052; https://doi.org/10.3390/ijms24109052
Submission received: 16 April 2023
/
Revised: 16 May 2023
/
Accepted: 17 May 2023
/
Published: 21 May 2023
(This article belongs to the Section Molecular Plant Sciences)
Abstract
:Seed germination is of great significance for plant development and crop yield. Recently, nitric oxide (NO) has been shown to not only serve as an important nitrogen source during seed development but also to participate in a variety of stress responses in plants to high salt, drought, and high temperature. In addition, NO can affect the process of seed germination by integrating multiple signaling pathways. However, due to the instability of NO gas activity, the network mechanism for its fine regulation of seed germination remains unclear. Therefore, this review aims to summarize the complex anabolic processes of NO in plants, to analyze the interaction mechanisms between NO-triggered signaling pathways and different plant hormones such as abscisic acid (ABA) and gibberellic acid (GA), ethylene (ET) and reactive oxygen species (ROS) signaling molecules, and to discuss the physiological responses and molecular mechanisms of seeds during the involvement of NO in abiotic stress, so as to provide a reference for solving the problems of seed dormancy release and improving plant stress tolerance.
1. Introduction
Plants reproduce by producing seeds, and the seed dormancy-germination process, which initiates the plant life cycle, is crucial for the establishment of seedlings and their ability to survive adverse external environments [1,2]. Seed dormancy refers to the physiological process of primary dormancy, which prevents the germination of viable seeds under favorable conditions [3,4,5]. The main types of dormancy include physiological dormancy, morphological dormancy, physical dormancy, and combinational dormancy [6,7]. Physiological dormancy occurs mainly in the seeds of gymnosperms and angiosperms and has been extensively studied in model plants such as Arabidopsis thaliana [8] and Nicotiana tabacum [9]; temperature stratification can break this type of dormancy. Morphologically dormant seeds are very clearly differentiated morphologically, but the internal embryo is not fully developed, which leads to delayed germination. Physical dormancy is mainly caused by mechanical hindrance from the external seed coat, and damage to the seed coat can induce seed germination under unfavorable conditions and break seed dormancy. Most seeds have a variable integrated dormancy state, which may have both physiological and morphological dormancy or both physiological and physical dormancy [6,7]. However, the combined dormant state of seeds can change depending on environmental conditions, and seeds can initiate germination to break dormancy when the external environment becomes favorable for plant growth [10]. There are three main stages in the process of breaking seed dormancy and germination: Initially, dry seeds absorb water (imbibition). Then, the embryo begins to expand and enzymes become activated. Finally, the protrusion of the embryo radicle and cotyl elongation complete the process of germination [4,11]. Thus, the transition from seed dormancy to germination is a complex process. In addition, this process is affected by internal genetic and external environmental factors as well as available nitrogen sources [12]. Light, temperature, nitrogenous compounds as signal inputs for seed dormancy and germination, activate changes in the levels of internal hormones such as ABA and GA. Seed dormancy is mainly induced by ABA. However, Seed germination is mainly induced by GA. The dynamic balance between ABA and GA is an important factor for breaking dormancy and promoting germination [12].
Nitrogen is an essential nutrient for plant growth and development. Reactive nitrogen species (RNS) have important regulatory functions in plant physiological activities. Recent studies have reported that the main forms of RNS in plants include nitric oxide (NO), nitrogen dioxide (NO2), peroxynitrite (ONOO−), nitrate (NO3−), and nitrite (HNO2) [13,14,15,16]. Among these forms, NO acts as a key gaseous molecule for regulating seed germination and improving crop productivity. It can promote seed germination by regulating ABA metabolism and GA synthesis pathways [11].
Evidence is accumulating that NO donors, such as sodium nitroprusside (SNP), S-nitroso-N-acetyl-D-penicillamine (SNAP), and S-nitrosoglutathione (GSNO), can break seed dormancy. In contrast, the NO scavenger (2-(4-carboxyphenyl)-4, 4, 5, 5-tetramethylimidazoline-1-oxyl-3 oxide; cPTIO) inhibited seed germination [17]. For example, Indian mustard (Brassica juncea L.) was treated with 0–250 μM SNP, and it was found that 100 μM SNP was the best promoter of seed germination [18]. A low SNP concentration of 25 μM reduced dormancy of Arabidopsis seeds, while an SNP concentration of more than 250 μM inhibited germination [19]. Fumigation of apple (Malus domestica Borkh.) embryos with 3 mM SNAP promoted germination, while 0.3 mM cPTIO treatment significantly inhibited germination [20]. SNAP treatment of 500 µM significantly accelerated the germination rate of chickpea seeds [21]. The germination index of rice seed treated with 100 μM GSNO significantly increased [22].
In addition, during seed germination of different types of plants, NO signals can affect physiological responses to a variety of factors, including light, temperature, drought, and salinity [12,23,24]. Moreover, a large number of studies have emphasized that the interaction between NO and phytohormones is crucial in seed biology [25]. Therefore, this review aims to summarize the interactions of NO with a variety of pathways to enhance our understanding of the physiological and potential molecular mechanisms regulating seed germination.
2. Main Text
2.1. Synthesis and Decomposition of Nitric Oxide in Plants
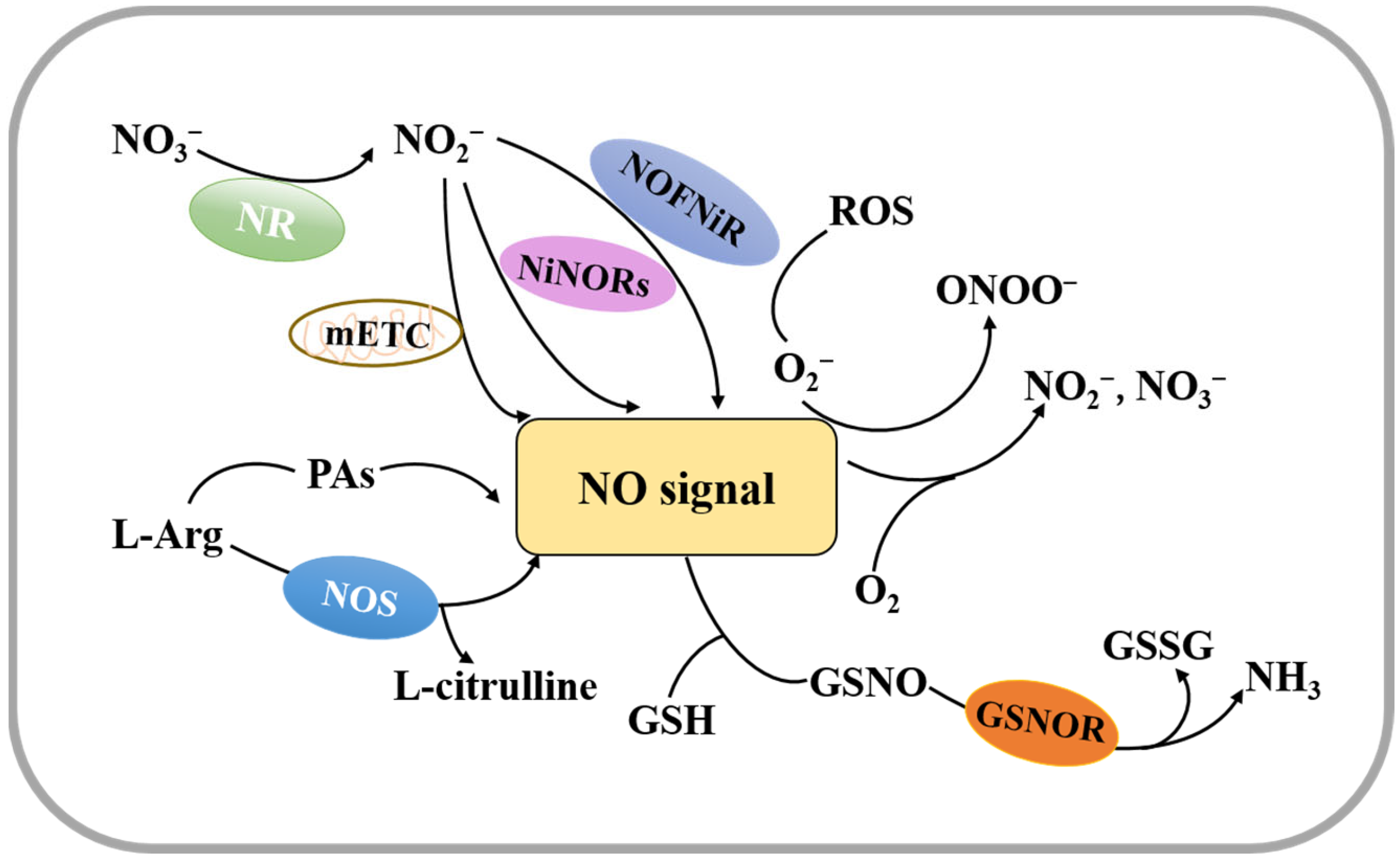
Nitrogen displays a range of oxidation states between a strongly oxidized form and a fully reduced form and can produce NO through oxidation or reduction mechanisms [26]. Current studies have shown that there are two pathways for plant NO synthesis (Figure 1). One is through a nitrate/nitrite-dependent reduction pathway, and the other is through an NO synthase (NOS)-mediated oxidation pathway [26,27,28,29].
The reduction pathway can be catalyzed by a variety of reductases, including nitrate reductase (NR), NO-forming nitrite reductase (NOFNiR), or a non-enzymatic pathway that occurs in the mitochondrial electron transport chain (mETC). Among these pathways, the mETC-dependent reductions of nitrite to NO are mainly through electron transfer between complex III and complex IV [27,28,30,31]. A recent study reported that there are two genes encoding NR in the Arabidopsis genome, NIA1/NR1 and NIA2/NR2 [31]. NR-mediated reduction of nitrite to NO is the major source of NO in higher plants, and this reaction occurs under aerobic or acidic conditions when concentrations of nitrite and nitrate are high [27,32]. With NR, nitrate is first reduced to nitrite, and then nitrite is reduced to NO [27].
The oxidative pathway is linked to the oxidation of L-arginine (Arg). Similar to NO synthesis in mammals, plants catalyze the oxidation of Arg to citrulline and NO using NOS [33,34]. Additionally, there is a polyamine synthesis pathway with Arg as a precursor, and studies have reported that increased levels of polyamines (PAs), such as spermine (Spm) and spermidine (Spd), lead to NO release in a variety of plants [35].
The non-enzymatic pathway to produce NO is utilized under anaerobic conditions with nitrite as the electron acceptor, which maintains plant respiration [27]. Under hypoxic conditions, plant cells produce NO mainly through the mETC [36], suggesting that plants may induce NO production through a variety of pathways (Figure 1). Therefore, future research on the complex mechanism of NO production is needed.
NO is unstable, and when it reacts with oxygen, NO3− or NO2− is formed [26]. NO can be scavenged both by reacting with ROS and by interacting with superoxide anions (O2−) to produce ONOO− [37,38]. It can also react with glutathione (GSH) to form GSNO, which is further reduced to glutathione disulfide (GSSG) and ammonia (NH3) by GSNO reductase (GSNOR) [39,40]. In addition, NO removal can be achieved by oxidation of NO to nitrate by hemoglobin (Figure 1). Maintaining the homeostasis of NO in plant cells by controlling its concentration is a significant factor for the seed germination process [41].
2.2. Nitric Oxide and Phytochrome Signaling Pathways Jointly Regulate Seed Germination under Light
Light is an important signal that affects plant survival; through photosynthesis, plants obtain energy to maintain growth. Current studies have reported that plants activate multiple photoreceptors, which include blue light receptors, red light/far-red light receptors and other photoreceptors that respond to light signals at different wavelengths [42,43]. There are five phytochrome (PHY) proteins in Arabidopsis; PHYA and PHYB are the photoreceptors primarily responsible for light-induced seed germination. PHYA responds to far-red light, and PHYB responds to red light [44,45].
Beligni explored the effect of NO on plant photomorphogenesis for the first time, revealing that NO donor SNP can induce lettuce seed germination depending on light [46]. Recent studies have shown that empress trees (Paulownia elongata) have physiological dormancy characteristics, and the seed germination rate after SNP treatment and continuous illumination for 12 h is significantly higher than that of unilluminated seeds [47]. NO stimulation also led to greening of etiolated seedlings of wheat (Triticum aestivum L. cv. Buck Patacon), which contained 30–40% more chlorophyll than controls when treated with 100 μM SNP and grown in the dark [46]. Another photoresponse to NO stimulation is the inhibition of hypocotyl and internode elongation. The germination rate of lettuce (Lactuca sativa L. cv. Grand Rapids) seeds treated with 100 μM SNP or 100 μM SNAP (both NO donors) and given red light (20 μmol s−1m−2) pulses was greater than 90%. For Arabidopsis (Arabidopsis thaliana (L.) Heynh.) treated with NO donors under dark conditions or potato (Solanum tuberosum (L.) cv. Pampeana) tubers under low light intensity, hypocotyl elongation was significantly inhibited, and the internode length of the plants was shortened [46]. These results indicate that NO-dependent photoreceptors affect seed germination, hypocotyl elongation, and plant greening.
In addition, the mechanism of action in Arabidopsis in which NO interacts with PHY to collectively affect seed germination was further elucidated (Figure 2). Exogenous application of the nitrogenous compound potassium nitrate (KNO3) to Arabidopsis seeds, followed by exposure to far-red light/red light for a short period of time, stimulates the activity of PHYA and PHYB, which in turn will promote rapid seed germination [48]. Within this process, PHYB is particularly crucial during its early stages and regulates seed germination by controlling the stability of the protein PHYTOCHROME-INTERACTION FACTOR 1 (PIF1) under red/far-red light [44]. An antagonistic relationship between PHYB and PIF1 has been reported (Figure 2). In the presence of red light, PHYB moves from the cytoplasm to the nucleus and promotes seed germination promoting expression of the 26S proteasome pathway leading to PIF1 degradation [49]. Significant hypocotyl elongation and increased anthocyanin levels were found in the NO biosynthesis-deficient Arabidopsis nia1, 2noa1-2 mutant after exposure to red light pulses. Moreover, PHYB protein content was reduced and PIF1, PIF3, and PIF4 expression was enhanced in the red light-induced mutants, and these results further support that NO and PHYB interact during seed germination [50]. In addition, NO in light can increase the accumulation of PHYB, which mediates the degradation of PIF3, leading to the inhibition of initial root elongation in Arabidopsis [51]. SOMNUS (SOM), as a direct target gene downstream of PIF1, can affect the expression levels of related genes in the abscisic acid (ABA) and gibberellic acid (GA) signaling pathways in photochrome-dependent photomorphogenesis (Figure 2). In the ABA signaling pathway, SOM promotes ABA biosynthesis by activating three ABA anabolic genes, ABA-DEFICIENT1 (ABA1), NINE-CIS-EPOXYCAROTENOID DEOXYGENASE6 (NCED6), and NCED9, and inhibits the expression of an ABA catabolic gene (CYP707A2), leading to inhibition of seed germination. In the GA signaling pathway, SOM suppresses GA biosynthesis by inhibiting GA synthesis metabolism genes (GA3ox1 and GA3ox2) and activating a catabolic gene (GA2ox2), leading to lower GA and inhibition of seed germination (Figure 2) [52]. In addition, the DELLA protein plays an important regulatory role in the GA signaling pathway by affecting the activity of PIF transcription factors under red light irradiation [50]. An antagonistic relationship between NO and GA was found during seed germination under red light irradiation. With increased concentration of the nitric oxide donor SNP, GA accumulation was inhibited, and GA-regulated DELLA protein accumulation was induced, which reduced the expression of PIFs, resulting in the inhibition of hypocotyl elongation (Figure 2) [50].
In addition to the regulation of seed germination under light through PHYB and its interacting protein PIFs, a member of the basic helix-loop-helix (bHLH) transcription factor family, LONG HYPOCOTYL IN FAR-RED (HFR1), was found to form dimers with PIF1, reducing the transcriptional activity of PIF1 (Figure 2). HFR1 accumulation increased under light stimulation, and the interaction of HFR1-PIF1 produced more precise regulation of light-initiated seed germination [53]. Recently, NO accumulation under red light conditions was found to not only inhibit PIF1 transcription but also enhance the HFR1-PIF1 interaction, ultimately leading to a weakened inhibitory effect of PIF1 on seed germination (Figure 2) [54].
NO regulation of seed germination is also associated with blue photoreceptors and functions in cooperation with regulatory factors such as methyl jasmonate (MJ) and phospholipase D (PLD). Blue light at 20 μmol m−2 s−1 inhibited germination of dormant wheat; however, when NO and MJ were applied simultaneously, the inhibitory effect of blue light was attenuated. Inhibition of synthesis of the ABA coding gene TaNCED1 and increased expression of the metabolic gene TaABA8′OH-1 decreased the ABA content, thereby reducing grain dormancy [55]. NO was also involved in the specific response of tomato seeds to blue light under osmotic stress. Exogenous addition of 0.2 mM S-nitrosoglutathione (SNG), an NO donor, promoted seed germination under 10 μmol m−2 s−1 blue light, while exogenous addition of 0.1 mM PTIO, an NO scavenger, inhibited seed germination [56]. PLD, a member of an important family of phospholipases in plants (Figure 2), hydrolyzes phospholipids to produce phosphatidic acid (PA) [57]. Activation of PLD and the PA produced by its hydrolysis during seed germination can regulate cytoskeletal organization and mediate the transport of Ca2+ signals in the cytoplasm [57,58]. During seed germination, NO content and PLD activity are increased by light. Indeed, in lettuce seeds, light induces NO production, stimulates PLD activity, and ultimately leads to the production of more PA, which in turn promotes seed germination [59]. However, further studies are still needed on the specific functions of light-induced NO production and the type of PA produced by PLD during seed germination.
2.3. The Crosstalk between Nitric Oxide and Plant Hormone Signaling Pathways in Seed Dormancy and Germination
2.3.1. Nitric Oxide and Abscisic Acid Signaling Pathways Jointly Regulate Seed Germination
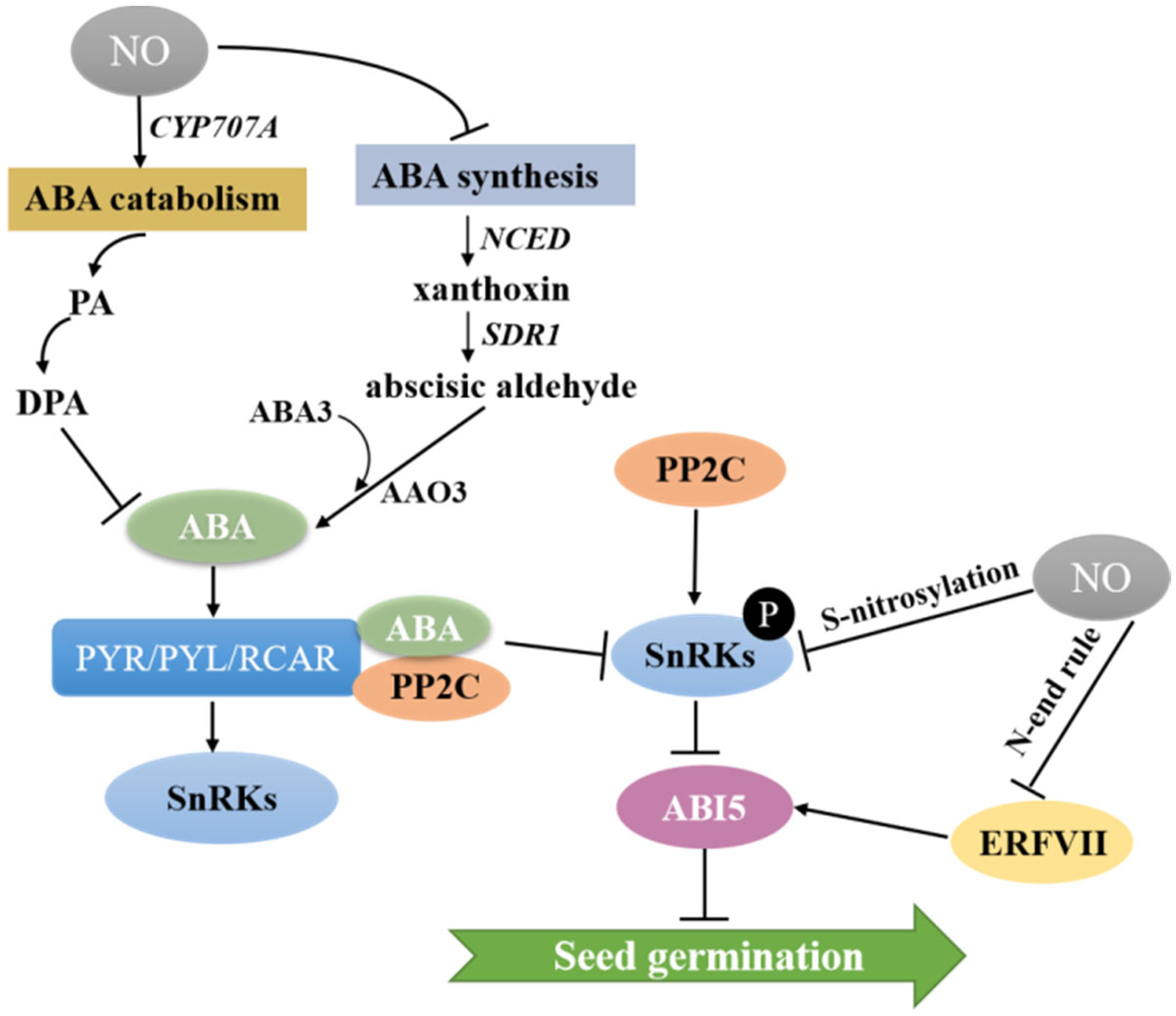
It is well known that the phytohormone ABA often plays a key role in the induction of seed dormancy [6,60]. In contrast, the presence of NO has been reported in many studies to reduce the sensitivity of seeds to ABA, and the interaction of NO and ABA in the regulation of seed germination has been demonstrated in plants such as Arabidopsis, switchgrass and warm-season C4 grasses [61,62,63,64]. The seed coat of Arabidopsis is composed of a dead coat and a layer of living aleurone cells. As the only endosperm tissue, the living, single-cell thick, aleurone layer stores lipids and proteins during seed maturation [65,66]. Therefore, the aleurone layer is the single most important factor determining seed dormancy. Arabidopsis and barley maintain the growth of embryos by storing lipids and proteins in the aleurone layer and increasing the expression of the NO oxide synthase gene (AtNOS1), while NO acts upstream of ABA and GA to increase biosynthesis of GA and inhibit biosynthesis of ABA, which conserves nutrients and promotes the normal germination of seeds [67]. The enzyme NCED catalyzes ABA biosynthesis to produce xanthoxin (Figure 3). Then, xanthoxin is converted to abscisic aldehyde by short-chain dehydrogenase reductase (SDR1), followed by oxidation of abscisic aldehyde oxidase (AAO3) to ABA catalyzed by ABA3 (Figure 3). Further, 8′-hydroxylation by the enzyme CYP707A, which can hydrolyze ABA into phaseic acid (PA) and dihydrophaseic acid (DPA), is thought to be the main pathway of ABA catabolism [68]. In the model plant Arabidopsis, NO induced rapid accumulation of ABA in the endosperm layer during early germination, leading to reduced ABA concentration in the embryo and thus promoting seed germination by upregulating the expression of ABA 8′-hydroxylase encoded by CYP707A2 and inducing catabolism of ABA [61,69]. In model plants, it is well established that NO regulates seed germination by affecting the expression of genes associated with ABA metabolism. A recent study reported that NO produced by NOS or NR in potato tubers significantly induced sprouting of the tubers by promoting the expression of the ABA catabolic gene StCYP707A1 while inhibiting the expression of the ABA biosynthesis-related gene StNCED1, thereby reducing ABA content and altering the ABA-GA balance [70]. Additionally, in the embryonic axes isolated from dormant apple embryos, NO reduced ABA concentration by promoting the expression of ABA catabolism gene CYP707A2 and inhibiting the expression of ABA synthesis genes (NCED3 and NCED9) [71].
NO also affects ABA signaling in guard cells during seed dormancy [72]. The mechanism is shown in Figure 3: ABA binds to its receptor PYR/PYL/RCAR, resulting in the inactivation of type 2C protein phosphatases (PP2C), which stimulates the action of SNF1-related protein kinase 2 (SnRK2). The transcription of ABSCISIC ACID INSENSITIVE5 (ABI5), an important repressor of seed germination, is promoted by phosphorylation, which leads to seed dormancy. When exogenous SNP is added or endogenous NO accumulates, NO-dependent protein modifications repress ABA signaling through S-nitrosylation of two important kinases of the SnRK2 family, SnRK2.2 and SnRK2.3, which in turn break seed dormancy [73,74,75]. In addition to mediating S-nitrosylation of SnRK2s, crosstalk between NO and ABA signaling also plays an important role in seed germination by indirectly regulating the expression of ABI5 in the N-end rule pathway (Figure 3). Group VII of the ethylene response factor (ERF/AP2) family (ERFVIIs) has a conserved the N-terminal domain. In the presence of nitric oxide, ERFVII is destabilized and exposed N-terminal Cysteine (Cys) residues are susceptible to oxidation, followed by arginylation by Arg-tRNA protein transferase and finally ubiquitination by E3 ligases, leading to repression of the downstream transcription factor ABI5, which in turn promotes seed germination [76,77].
2.3.2. Nitric Oxide and Auxin Signaling Pathways Jointly Regulate Seed Germination
During seed germination in higher plants, NO-mediated release of embryonic dormancy is positively correlated with ethylene synthesis [68]. Ethylene synthesis begins with the activation of methionine (Met) by S-adenosyl-L-methionine synthetase (SAM synthetase) to produce S-adenosyl-methionine (S-AdoMet) (Figure 4). The transformation to 1-aminocyclopropane-1-carboxylic acid (ACC) and the by-product 5′-methylthioadenosine (MTA) is catalyzed by ACC synthase, S-adenosyl-L-methionine methylthioadenosine-lyase (ACS), and then ACC is oxidized using ACC oxidase (ACO) to produce ethylene, CO2, and hydrogen cyanide (HCN) [78]. MTA regenerates Met through the methionine cycle [78]. To date, five ethylene receptors have been identified in the model plant Arabidopsis. Among them, ethylene resistant 1 (ETR1) plays an important role in NO-mediated ethylene signal transduction [79,80]. Ethylene signaling is regulated by constitutive triple response 1 (CTR1), and when the ethylene receptor ETR1 on the endoplasmic reticulum receives the ethylene signal, it leads to the inactivation of the receptor and CTR1. CTR1 is a negative regulator of ETHYLENE INSENSITIVE2 (EIN2), while EIN2 acts as a positive regulator of ethylene signaling by increasing the activity of the nuclear transcription factor EIN3, which in turn activates the transcription of downstream ethylene response elements such as ethylene-responsive factor (ERF) and other genes that promote seed germination [81]. Recent studies have also shown that EIN2, a key factor in the ethylene signaling pathway, is involved in jasmonic acid (JA)-induced primary root growth through effects on NO accumulation [82]. The involvement of ETR1, EIN2, and EIN3 in NIA1/2-mediated NO production is associated with salicylic acid (SA)-induced stomatal closure. All these findings confirmed the important connections between NO and the ethylene signaling pathway [83].
NO-ethylene interactions promote seed germination in a variety of plants. For instance, exogenous NO donors SNP or SNAP can increase the germination of apple embryos, while the ethylene synthesis inhibitor aminooxyacetate acid (AOA) represses the germination of embryos by NO donors [84]. Moreover, the effect of NO on ethylene production was associated with the accumulation of ROS. Pretreatment of apple embryos with HCN or SNP leads to short-term accumulation of ROS (H2O2 and O2−), which breaks the deep dormancy of apple embryos. However, the exogenous addition of ethylene glycol or the ethylene precursor ACC induced normal growth of embryonic roots and greening of cotyledons after embryo germination [85]. Gniazdowska et al. investigated the relationship between ethylene and the role of NO and HCN in mediating seed germination and demonstrated that early in the process of seed germination, NO and HCN pretreatment could directly change the activities of ACS and ACO, key enzymes of the ethylene synthesis process. Moreover, ACC can be oxidized to ethylene by ROS free radicals in the early stage of seed germination, leading to increased ethylene content [86]. In addition, a recent study found that increased SNP during germination of Sorbus pohuashanensis is related to ethylene synthesis and ROS accumulation. NO promoted hypocotyl and radicle growth by inducing ethylene biosynthesis and ROS accumulation, enhancing the capacity of antioxidant defense systems, and reducing ABA content [87]. Exogenous NO donors increase the concentrations of endogenous ethylene and ROS, promoting seed germination. Studies have shown that exogenous ethylene treatments can increase the production of ROS in the embryo axis and break sunflower seed dormancy. Moreover, exogenous application of 0.1 mM methyl viologen (MV), an ROS-inducing herbicide, can alleviate the inhibitory effect of ABA on sunflower seed germination and reduce the ABA content in the embryo axis, reducing the inhibitory effect on seed germination [88].
Karrikin-1 (KAR1) is an active compound derived from smoke that affects seed germination in different types of plants [89]. Sami et al. showed that Brassica oleracea L. (Chinese cabbage) seeds treated with NO and KAR1 had reduced ABA content and increased GA and ROS content, but the activities of antioxidant system enzymes, including catalase (CAT) and glutathione reductase (GR), increased [90]. The expression levels of ethylene biosynthesis genes (BOACS7, BOACS9, and BOACS11), ethylene receptors (BOETR1 and BOETR2), and an ACC oxidase ACO gene (BOACO1) were significantly increased with induction by NO and KAR1 [90]. Furthermore, KAR1 and HCN treatments reduced the ABA content and increased the GA content in B. oleracea seeds with secondary dormancy and induced the expression of ethylene synthesis genes and ethylene receptor genes, releasing seed dormancy [91]. However, the mechanism of the interaction of NO with KAR1 and HCN to release seed dormancy by mediating ethylene synthesis still requires further exploration. Further, ethylene, HCN, and NO gases applied directly to dry seeds can break seed dormancy [92]. Therefore, in addition to the application of solutions, the direct application of gas can induce release from seed dormancy; future research can reveal new insights on the process of seed dormancy release in different types of plants.
2.3.3. Nitric Oxide and Gibberellin Signaling Pathways Jointly Regulate Seed Germination
NO-GA interactions often are antagonistic to the regulatory role of ABA during seed germination [25]. Exogenous SNP and GA3 can improve seed viability and seed germination in grapevines [93]. In Arabidopsis, NO increases GA biosynthesis by inducing downstream transcript levels of gibberellin 3-oxidase 1 (GA3ox1) and GA3ox2, which promotes seed germination [67]. NO not only regulates the GA signal but also interacts with ethylene to regulate seed germination. For example, the exogenous hormone ethephon or ethylene precursor ACC can release ethylene and GA3 to break seed dormancy and induce seed germination [94,95]. Additionally, seeds are sensitive to dry environments, and the physiological response of seeds to GA3 and ethylene is enhanced with dry storage and stratified environments [95]. Kepczynski et al. further investigated the interactions between NO, GA3, and ethylene during seed germination, and confirmed that the stimulation of seed germination by NO and GA3 was positively correlated with ethylene production by measuring the ethylene content of the radicle before it protruded from the seed coat. The exogenous addition of the NO inhibitor cPTIO or the ethylene receptor inhibitor 2,5-norbornadiene (NBD) inhibited the induction of seed germination by NO and GA3, whereas the repression was alleviated when ethylene was added in combination with NBD [96]. These results suggest that the release of seed dormancy by GA3 and NO is dependent on the function of ethylene.
2.3.4. Nitric Oxide Regulates Seed Germination Together with Polyamines and Jasmonic Acid Signaling Pathways
In addition to inducing the synthesis of GAs to promote seed germination, NO can stimulate seed dormancy release by regulating the synthesis of JA in wheat grains and apple embryos [55,71]. It was found that NO could induce the expression of the genes AOS1, JMT, and JAR1 encoding JA and JA derivatives JA methylester (MeJA) and JA-isoleucine (JA-Ile), respectively, promoting seed germination [71].
Polyamines (PAs) are an important plant growth regulator [97]. Studies have shown that PA can rapidly induce the release of NO in embryogenic suspension cultures of Araucaria angustifolia [98]. In plant cells, the synthesis of PA is related to the ethylene synthesis pathway (Figure 4). SAM is central to ethylene synthesis, then decarboxylated SAM (dcSAM) is produced under the action of SAM decarboxylase (SAMDC), and finally, PA is produced by PA synthase. Moreover, PA oxidase (PAO) catalyzes the oxidative deamination of PAs, producing H2O2 [99,100,101]. However, increased levels of PAs, including (Spm) and Spermidine (Spd) also resulted in a significant increase in NO released by the Arg-dependent pathway. [101]. These results suggest a close regulatory network between NO and PAs, but the mechanism of their interaction during the transition from seed dormancy to germination is not well understood. Recent studies have reported that at the transcriptional level, NO can increase the expression of PA biosynthetic genes MdPAO, MdSAMDC1, and MdSPDS2a to release apple embryonic dormancy and induce seed germination [101,102].
2.4. Regulation of Nitric Oxide on Seed Germination under Abiotic Stress
2.4.1. Mechanism of Nitric Oxide Action on Seed Germination under Salt Stress
The NO synthetic protein is AtNOA1 in Arabidopsis genome and OsNOA1 in Oryza sativa L. genome. The molecular mechanisms of the two proteins are similar in regulating the salt tolerance of plants during seed germination. OsNOA1 can complement the atnoa1 mutant phenotype and the loss of chlorophyll synthesis by promoting NO release, and improve the salt tolerance of plants during seed germination by decreasing the Na+/K+ ratio in the mutant (Table 1) [103,104]. Seed germination can also be promoted by magnetic fields. For example, NO promoted α-amylase activity and the physiological response of seed germination in maize (Zea mays) under magnetic initiation [105]. In soybean (Glycine max (L.) Merrill) seeds, GmNOS2 and GmNR1 may be ideal candidate genes involved in NO production (Table 1). Magnetopriming-mediated NO production decreased the Na+/K+ ratio and maintained the balance of ABA, GA, and ondole-3-acetic acid (IAA) hormones, thereby improving the salt tolerance index of soybean [106]. NO not only affects ion homeostasis under salt stress but also alleviates oxidative damage in plants by upregulating the antioxidant defense system and maintaining ROS homeostasis under stress conditions [107,108,109,110]. In tomatoes, overexpression of tomato glutathione reductase (SlGR) led to the accumulation of more antioxidant substances under 100 mM NaCl treatment (Table 1) [111]. Additionally, 100 μM SNP, by releasing cyanide, reduced lipid peroxidation and increased the activity of antioxidant enzymes, which in turn increased the speed and rate of germination [112]. Recently, Hajihashemi et al. found that RNS and ROS, together with Ca2+, regulate the physiological response of Chenopodium quinoa seeds under salt stress. Treatment with a combination of 5 mM CaCl2, 5 mM H2O2, and 0.2 mM SNP leading to starch hydrolysis and increased water-soluble sugar content, effectively alleviating the adverse effects of salt stress on seed germination [113].
In addition to promoting seed germination under salt stress by providing NO through SNP, KNO3, the major form of nitrate, is useful for breaking seed dormancy in plants such as Arabidopsis [114] and maize [115]. However, the specific mechanism of nitrate-mediated responses of plants to salt remains unclear. Studies have reported that the main regulators of nitrate signaling include Chlorate-resistant1 (NRT1.1, CHL1, NPF6.3), Arabidopsis nitrate regulated 1(ANR1), Teosinte branched1/cycloidea/proliferating cell factor1-20 (TCP20), and NIN-like protein (NLP) [116,117,118,119]. NRT1.1 is a dual-affinity nitrate transporter protein in Arabidopsis that acts as a plasma membrane nitrate receptor to sense external nitrate signals [120]. It has been reported that NRT1.1 relies on NO3− for Na+ transport (Table 1). In the presence of nitrate, Na+ accumulation in the nrt1.1 mutant was significantly lower than that of wild type [121]. However, when NH4+ was the only nitrogen source, the nrt1.1 mutant significantly reduced the absorption of Cl− and eliminated the salt-sensitive reaction caused by NH4+ after stress treatment with 25 mM NaCl [122]. ANR1 also responds to abiotic stress, and overexpression of ANR1 produces a salt-sensitive phenotype (Table 1) [123]. The NLP gene family encodes core transcription factors that regulate nitrate signaling in plants [124]. In Arabidopsis, nine members of this family have conserved phosphorylation sites in the N terminus that are critical for responding to nitrate signaling [125,126]. NLP8, a nitrate-activated transcription factor, was found to activate the expression of the ABA catabolic gene CYP707A2 in Arabidopsis and is essential for nitrate-induced seed germination (Table 1) [127]. The NR pathway is the main pathway for NO production [128]. Nitrate release of NO under NR can promote seed germination. In NaCl-stressed Arabidopsis seeds when exogenously treated with KNO3, nitrate serves as a nitrogen source and counteracts NaCl inhibition of seed germination by upregulating the expression of the GA20ox1 gene encoding GA synthase. The SPATULA (SPT) gene encodes a bHLH transcription factor, and the CYP707A2 gene encodes an ABA catabolic enzyme [114]. During the germination of cucumber seeds treated with 50 mM NaCl, NO-dependent NR activity also significantly improved salt tolerance [129]. Rice with defective OsNR gene function exhibits a phenotype that is sensitive to salt stress and shows delayed germination (Table 1). During rice seed germination, the nitrate-responsive transcription factor NLP2 promotes the expression of NO-activated ABA catabolic genes ABA8ox1 and ABAox2 through the NR pathway, thereby improving plant salt tolerance [130]. NLP7 and TCP20 are major regulators of nitrate response genes in Arabidopsis roots and regulate the primary nitrate response [117,131], and both NLP6 and NLP7 strongly interact with TCP20 to influence cell proliferation factors that regulate the cell cycle [132]. Recently, deletion of the NLP7 function has been shown to lead to a significant reduction in the transcript level of the ABA biosynthesis gene NCED3, which in turn improves salt tolerance in plants (Table 1) [133]. However, the interaction of other NLP family transcription factors with nitrate signaling response genes during salt stress-inhibited seed germination still needs to be further explored. The interaction of NO and ethylene also reduced the inhibitory effect of salt stress on seed germination [134]. It was found that the NO-dependent ethylene signaling factor EIN3 can resist high salt stress to promote seed germination (Table 1) [135]. Additionally, recent studies have found that RNS can promote embryo germination of S. pohuashanensis under NaHCO3 stress mediated by exogenous ethylene [136].
2.4.2. Mechanism of Nitric Oxide Action on Seed Germination under Drought Stress
NO can significantly increase the tolerance of plants to drought stress and improve seed viability and crop yield [137]. For example, the exogenous NO donors SNP and GSNO induced the transcription of Oryza sativa plasma membrane channel protein (OsPIP1;3), which in turn induced seed germination under water stress conditions (Table 1) [22]. Transformation of the unicellular marine alga Ostreococcus tauri NOS gene (OtNOS) into Arabidopsis significantly elevated NO accumulation and enhanced seed tolerance to salt stress [138].
ABA, an important signaling molecule for water stress, regulates the sensitivity of plants to ABA under osmotic stress by interacting with GAP1, the gene encoding the NAC family transcription factor ANAC089 (Table 1) [139]. Further, inhibition of ABA synthesis under abiotic stress by ANAC089 through regulation of NO homeostasis during seed development has been suggested [139]. Moreover, ABA-induced stomatal closure leads to a reduction in transpiration rate; NO mediation of stomatal development under drought stress also affects seed germination. Recent studies found that exogenous NO treatment mediated ABA synthesis in guard cells or induced quickly activating Anion Channel 1/Aluminum-activated Malate Transporter 12 (QUAC1/ALMT12) anion channels leading to stomatal closure and enhanced drought tolerance in plants [140]. In addition to exogenous NO affecting ABA synthesis, the response to the ABA signaling pathway via the NR pathway or NOA1-dependent NO production is important [141]. NO synthesis gene mutant nia1nia2 results in lower than 1% bioactivity of NR and reduced accumulation of NO in guard cells [142]. Further, although NIA1 and NIA2 are functionally redundant, only NIA1 is required for NO production in response to ABA [143]. It has been reported that Atnoa1 mutants reduced NO response to ABA in root tips and guard cells [103]. Furthermore, the reduced NO content in Atnoa1 mutants was associated with protein synthesis in chloroplasts [144]. In addition, nia1nia2noa1-2 plants had reduced seed germination potential and were overly sensitive to ABA [137]. Hence, the interaction between NR/NIA and AtNOA1-mediated NO-ABA in guard cell signaling deserves considerable attention in future investigations into the molecular mechanisms of seed germination.
NO-dependent post-translational modifications also play an important role in the regulation of seed germination during abiotic stress [77]. NO inhibits the activity of SNF1-associated protein kinase (SnRK2.6/OST1) through GSNO, thereby inhibiting ABA signaling in guard cells (Table 1) [75]. Furthermore, dried seeds induced the expression of ABA-induced growth inhibitor ABI5; however, NO and ABA had antagonistic effects on ABI5 expression. Large amounts of NO are produced at the early stage of seed imbibition, which induces S-nitrosylation of ABI5 at Cys153 and promotes seed germination through proteasomal degradation [77]. NO not only affects seed germination through S-nitrosylation but also regulates the degradation of ethylene response factor ERFVII, a substrate of E3 ubiquitin ligase PROTEOLYSIS6 (PRT6), which suppresses ABI5 expression through ubiquitination modifications. Arabidopsis prt6-1nlp7-1 seeds were found to be hypersensitive to ABA, suggesting that NLP7 and PRT6 interactions enhanced seed tolerance to ABA (Table 1) [76,145,146].
2.4.3. Involvement of Nitric Oxide in the Regulation of Seed Germination by Ambient Temperature
Seed germination is often related to temperature, and low temperature stratification plays a positive role in the breaking dormancy of most seeds. Many studies have shown that RNS, ROS, and HCN can induce the breaking of deep dormancy during seed development. For example, SNP and cold stratification together significantly increased the germination rate of Empress Tree seeds. NO scavenger cPTIO significantly inhibited the rapid development of apple embryos pretreated with NO and HCN into seedlings [47,85]. Subsequently, it was found that NO or HCN induced transient accumulation of ROS during the enhancement of germination in apple embryos and sunflower (Helianthus annuus) embryos, which was associated with the AsA-GSH system [147,148,149]. The GSH system of seeds stratified at low temperatures for 40 d showed significantly higher glutathione peroxidase (GPX) and glutathione reductase (GR) activities, which in turn promoted seed germination [150]. GSNO is an important source of stable release of NO [151,152]. GSNOR and GR enzymes can break seed dormancy by regulating NO levels during low temperature stratification-mediated germination of apple embryos. Reduction of GSNO by GSNOR leads to the formation of oxidized forms of GSH and NH2OH, which in turn eliminates GSNO to maintain cellular RNS levels and prevent excessive accumulation of NO. GSSG is further reduced to GSH by the action of GR, which promotes seed germination [151,153,154].
However, ambient temperatures that are too low or too high often cause adverse effects on plants, resulting in suppressed seed germination and inhibited plant growth. A recent study found that irrigation of 89.4 mg/L SNP on two rapeseed (Brassica napus L.) varieties (ZY15 and HY49) significantly improved the resistance of rapeseeds to low temperature and drought stress, thus increasing the seed germination rate [155]. Tomato growth is inhibited at temperatures below 25 °C and growth nearly ceases below 6 °C. In tomato seeds, SNP treatment at 10 °C significantly increased amylase activity and soluble sugar content and improved low temperature tolerance [156]. In addition, KNO3 has an important effect on seed germination of Sorbus pohuashanensis. For seeds in long-term cold storage, incubation at variable temperatures (5 °C followed by 25 °C followed by 5 °C) after pretreatment with KNO3 significantly increased the germination rate compared to seeds incubated at 5 °C or 25 °C followed by 5 °C [157]. However, recent studies have found that methylglyoxal (MG) can promote maize germination by reducing endogenous NO levels. This finding suggests that, in addition to the degradation of ABA, NO may induce other signaling pathways to enhance maize germination at low temperatures (13 °C) [158].
High temperatures also reduced seed germination, while 1 mM SNP significantly increased the pod number and seed numbers in lentils (Lens culinaris Medik.) exposed to high temperature stress of 32 °C [159]. High temperature stress also led to a decrease in GSNO reductase (GSNOR1) activity preventing the degradation of the NO donor GSNO, which resulted in the accumulation of more RNS and reduced seed germination rate. Genetic analysis showed that overexpression of GSNOR1 increased the nitrosation modification level of ABI5 and induced the degradation of downstream ABI5 protein increasing seed germination (Table 1) [160]. These studies suggest directions for future research on different types of gene modification to improve plant heat tolerance.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of NO-regulated target genes on seed germination under abiotic stress.
| Stress Types | Gene | Species | Function Description | References |
|---|---|---|---|---|
| Salt Stress | OsNOA1 | Oryza sativa | Overexpression of OsNOA1 gene improved plant salt tolerance by reducing Na+/K+ ratio in the mutant. | [104] |
| GmNOS2 GmNR1 | Glycine max | The Na+/K+ ratio was decreased and the balance of ABA, GA and IAA hormones was maintained, improving the salt tolerance index of soybean. | [106] | |
| SlGR | Solanum lycopersicum | Overexpression of SlGR significantly increased the activity of antioxidant enzymes and reduced the oxidative damage of tobacco seeds. | [111] | |
| NRT1.1 | Arabidopsis thaliana | Na+ accumulation was promoted after NO3- treatment, and Cl- accumulation was promoted after NH4+ treatment. | [121,122] | |
| ANR1 | Arabidopsis thaliana | Overexpression of ANR1 produces a salt-sensitive phenotype and inhibited seed germination | [123] | |
| NLP8 | Arabidopsis thaliana | Activate ABA catabolic gene CYP707A2 to promote germination. | [127] | |
| NLP2 | Arabidopsis thaliana | The NLP2-NR pathway activates the expression of ABA catabolic genes ABA8ox1 and ABAox2. | [130] | |
| NLP7 | Arabidopsis thaliana | Increasing the transcription level of ABA biosynthesis gene NCED3 to inhibit seed germination. | [133] | |
| NIA1,2 | Arabidopsis thaliana | It can resist salt stress by promoting EIN3 expression and transcription of downstream ethylene response genes. | [135] | |
| Drought stress | OsPIP1;3 | Oryza sativa | Overexpression of OsPIP1;3 induced germination under water stress conditions. | [22] |
| Drought stress | OtNOS | Arabidopsis thaliana | Overexpression of OtNOS elevated NO accumulation and enhanced seed tolerance to salt stress. | [138] |
| GAP1 | Arabidopsis thaliana | Functionally deficient mutants showed insensitivity to ABA. | [139] | |
| SnRK2.6 | Arabidopsis thaliana | NO inhibits the activity of SNF1-associated protein kinase (SnRK2.6) through GSNO, thereby inhibiting ABA signaling in guard cells. | [75] | |
| PRT6-1 NLP7-1 | Arabidopsis thaliana | The interaction between NLP7 and PRT6 enhanced seed tolerance to ABA. | [146] | |
| Heat stress | GSNOR1 | Arabidopsis thaliana | Overexpression of GSNOR1 increased the nitrosation modification level of ABI5 and induced the degradation of downstream ABI5 protein increasing seed germination | [160] |
3. Conclusions and Future Perspectives
Over the past 10 years, analysis of different types of RNS revealed that the seed germination rate could be significantly increased by gaseous NO or NO donor (SNP, SNAP, and GSNO) pretreatment. However, the NO synthesis and catabolism pathway remains a controversial topic. Although RNS and ROS can collectively affect seed germination, it has been reported that RNS and ROS were unstable and both induced and hindered seed germination. Studies showed that different concentrations of NO had different effects on seed germination. Low concentration of NO promoted seed germination, while high concentration of NO inhibited seed germination [18,20]. Moreover, exogenous NO interacted with phytohormone signaling pathways, such as ethylene synthesis, ABA catabolism, and GA synthesis, during early seed germination and increased seed germination by affecting the accumulation of ROS and the synthesis of enzymes related to the antioxidant system. However, excessive accumulation of H2O2 can also have a negative impact on plants. Although previously there have been extensive studies on the effect of NO on the physiological processes of seeds, whether the mechanisms of action on different phases of germination during the breaking of seed dormancy in different plants are similar or different remains an open question. In addition, the complex network of interactions between NO and different phytohormone signals deserves further exploration.
Under abiotic stress conditions, NO synthesis via the nitrate reduction (NR) pathway or NO synthase (NOS) has major effects on seed dormancy and germination. In the model plant Arabidopsis, the expression of downstream genes in the ABA and ethylene signaling pathways is regulated by the expression of NLP transcription factors in the NO3− signaling pathway, which has an important function in improving plant tolerance to abiotic stress. This mechanism can also serve as a reference for the investigation of transcription factors during seed germination in non-model plants. Additionally, NO-dependent post-translational modifications (PTM), such as S-nitrosylation or ubiquitination, can regulate protein expression in response to stress. However, the effects of different types of PTM on NO-mediated seed germination require further investigation. With the continuous development of genome sequencing, the use of multi-omics to explore the key genes and gene interactions in the regulation by NO of seed dormancy and germination will provide a reference for the elucidation of the molecular mechanism of seed germination.
Author Contributions
Y.Z. contributed to the writing of the manuscript and figure preparation. R.W. checked the English expression form. Supervision: C.Z. and X.W. L.Y. and H.S. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was provided by the National Natural Science Foundation of China (No. 32071757) and the National Natural Science Foundation of Heilongjiang Province of China (C201407).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Reed, R.C.; Bradford, K.J.; Khanday, I. Seed germination and vigor: Ensuring crop sustainability in a changing climate. Heredity 2022, 128, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed]
- Chahtane, H.; Kim, W.; Lopez-Molina, L. Primary seed dormancy: A temporally multilayered riddle waiting to be unlocked. J. Exp. Bot. 2017, 68, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J. Seed germination and dormancy. Plant Cell 1997, 9, 1055. [Google Scholar] [CrossRef]
- Hilhorst, H.W. A critical update on seed dormancy. I. Primary dormancy1. Seed Sci. Res. 1995, 5, 61–73. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New. Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Deng, G.; Sun, H.; Hu, Y.; Yang, Y.; Li, P.; Chen, Y.; Zhu, Y.; Zhou, Y.; Huang, J.; Neill, S.; et al. A Transcription Factor WRKY36 Interacts with AFP2 to Break Primary Seed Dormancy by Progressively Silencing DOG1 in Arabidopsis. New. Phytol. 2023, 238, 688–704. [Google Scholar] [CrossRef]
- Grainge, G.; Nakabayashi, K.; Iza, F.; Leubner-Metzger, G.; Steinbrecher, T. Gas-Plasma-Activated Water Impact on Photo-Dependent Dormancy Mechanisms in Nicotiana tabacum Seeds. Int. J. Mol. Sci. 2022, 23, 6709. [Google Scholar] [CrossRef]
- Née, G.; Xiang, Y.; Soppe, W.J. The release of dormancy, a wake-up call for seeds to germinate. Curr. Opin. Plant Biol. 2017, 35, 8–14. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Chen, Z. The control of seed dormancy and germination by temperature, light and nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar] [CrossRef]
- Del Río, L.A. ROS and RNS in plant physiology: An overview. J. Exp. Bot. 2015, 66, 2827–2837. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef]
- Augusto, O.; Bonini, M.G.; Amanso, A.M.; Linares, E.; Santos, C.C.; De Menezes, S.L. Nitrogen dioxide and carbonate radical anion: Two emerging radicals in biology. Free Radic. Biol. Med. 2002, 32, 841–859. [Google Scholar] [CrossRef]
- Domingos, P.; Prado, A.M.; Wong, A.; Gehring, C.; Feijo, J.A. Nitric oxide: A multitasked signaling gas in plants. Mol. Plant 2015, 8, 506–520. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Malvankar, M.R.; Karle, S.B.; Kumar, K. Reactive nitrogen species: Paradigms of cellular signaling and regulation of salt stress in plants. Environ. Exp. Bot. 2019, 161, 86–97. [Google Scholar] [CrossRef]
- Rather, B.A.; Mir, I.R.; Masood, A.; Anjum, N.A.; Khan, N.A. Nitric Oxide Pre-Treatment Advances Seed Germination and Alleviates Copper-Induced Photosynthetic Inhibition in Indian Mustard. Plants 2020, 9, 776. [Google Scholar] [CrossRef]
- Bethke, P.C.; Libourel, I.G.L.; Jones, R.L. Nitric oxide reduces seed dormancy in Arabidopsis. J. Exp. Bot. 2005, 57, 517–526. [Google Scholar] [CrossRef]
- Krasuska, U.; Ciacka, K.; Orzechowski, S.; Fettke, J.; Bogatek, R.; Gniazdowska, A. Modification of the endogenous NO level influences apple embryos dormancy by alterations of nitrated and biotinylated protein patterns. Planta 2016, 244, 877–891. [Google Scholar] [CrossRef]
- Pandey, S.; Kumari, A.; Shree, M.; Kumar, V.; Singh, P.; Bharadwaj, C.; Loake, G.J.; Parida, S.K.; Masakapalli, S.K.; Gupta, K.J. Nitric oxide accelerates germination via the regulation of respiration in chickpea. J. Exp. Bot. 2019, 70, 4539–4555. [Google Scholar] [CrossRef]
- Liu, H.Y.; Yu, X.; Cui, D.Y.; Sun, M.H.; Sun, W.N.; Tang, Z.C.; Kwak, S.S.; Su, W.A. The role of water channel proteins and nitric oxide signaling in rice seed germination. Cell Res. 2007, 17, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.X.; Li, X.; Li, C.; Zhao, L. The role of nitric oxide in plant responses to salt stress. Int. J. Mol. Sci. 2022, 23, 6167. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Oliveira, P.J.; Oliveira, H.C.; Kolbert, Z.; Freschi, L. The light and dark sides of nitric oxide: Multifaceted roles of nitric oxide in plant responses to light. J. Exp. Bot. 2021, 72, 885–903. [Google Scholar] [CrossRef] [PubMed]
- Sanz, L.; Albertos, P.; Mateos, I.; Sánchez-Vicente, I.; Lechón, T.; Fernández-Marcos, M.; Lorenzo, O. Nitric oxide (NO) and phytohormones crosstalk during early plant development. J. Exp. Bot. 2015, 66, 2857–2868. [Google Scholar] [CrossRef] [PubMed]
- Astier, J.; Gross, I.; Durner, J. Nitric oxide production in plants: An update. J. Exp. Bot. 2018, 69, 3401–3411. [Google Scholar] [CrossRef]
- León, J.; Costa-Broseta, Á. Present knowledge and controversies, deficiencies, and misconceptions on nitric oxide synthesis, sensing, and signaling in plants. Plant Cell Environ. 2020, 43, 1–15. [Google Scholar] [CrossRef]
- Yu, M.; Lamattina, L.; Spoel, S.H.; Loake, G.J. Nitric oxide function in plant biology: A redox cue in deconvolution. New. Phytol. 2014, 202, 1142–1156. [Google Scholar] [CrossRef]
- Besson-Bard, A.; Pugin, A.; Wendehenne, D. New insights into nitric oxide signaling in plants. Annu. Rev. Plant Biol. 2008, 59, 21–39. [Google Scholar] [CrossRef]
- Rockel, P.; Strube, F.; Rockel, A.; Wildt, J.; Kaiser, W.M. Regulation of nitric oxide (NO) production by plant nitrate reductase in vivo and in vitro. J. Exp. Bot. 2002, 53, 103–110. [Google Scholar] [CrossRef]
- Yamasaki, H. Nitrite-dependent nitric oxide production pathway: Implications for involvement of active nitrogen species in photoinhibition in vivo. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhang, Y.; Liu, L.; Liu, X.; Li, B.; Jin, C.; Lin, X. Molecular functions of nitric oxide and its potential applications in horticultural crops. Hort. Res. 2021, 8, 71. [Google Scholar] [CrossRef] [PubMed]
- Zemojtel, T.; Fröhlich, A.; Palmieri, M.C.; Kolanczyk, M.; Mikula, I.; Wyrwicz, L.S.; Wanker, E.E.; Mundlos, S.; Vingron, M.; Martasek, P.; et al. Plant nitric oxide synthase: A never-ending story? Trends Plant Sci. 2006, 11, 524–525. [Google Scholar] [CrossRef] [PubMed]
- Barroso, J.B.; Corpas, F.J.; Carreras, A.; Sandalio, L.M.; Valderrama, R.; Palma, J.; Lupiánez, J.A.; del Rıo, L.A. Localization of nitric-oxide synthase in plant peroxisomes. J. Biol. Chem. 1999, 274, 36729–36733. [Google Scholar] [CrossRef]
- Tun, N.N.; Santa-Catarina, C.; Begum, T.; Silveira, V.; Handro, W.; Floh, E.I.; Scherer, G.F. Polyamines induce rapid biosynthesis of nitric oxide (NO) in Arabidopsis thaliana seedlings. Plant Cell Physiol. 2006, 47, 346–354. [Google Scholar] [CrossRef]
- Gupta, K.J.; Stoimenova, M.; Kaiser, W.M. In higher plants, only root mitochondria, but not leaf mitochondria reduce nitrite to NO, in vitro and in situ. J. Exp. Bot. 2005, 56, 2601–2609. [Google Scholar] [CrossRef]
- Begara-Morales, J.C.; Sánchez-Calvo, B.; Chaki, M.; Valderrama, R.; Mata-Pérez, C.; López-Jaramillo, J.; Padilla, M.N.; Carreras, A.; Corpas, F.J.; Barroso, J.B. Dual regulation of cytosolic ascorbate peroxidase (APX) by tyrosine nitration and S-nitrosylation. J. Exp. Bot. 2014, 65, 527–538. [Google Scholar] [CrossRef]
- Gaupels, F.; Spiazzi-Vandelle, E.; Yang, D.; Delledonne, M. Detection of peroxynitrite accumulation in Arabidopsis thaliana during the hypersensitive defense response. Nitric Oxide 2011, 25, 222–228. [Google Scholar] [CrossRef]
- Kwon, E.; Feechan, A.; Yun, B.W.; Hwang, B.H.; Pallas, J.A.; Kang, J.G.; Loake, G.J. AtGSNOR1 function is required for multiple developmental programs in Arabidopsis. Planta 2012, 236, 887–900. [Google Scholar] [CrossRef]
- Leterrier, M.; Chaki, M.; Airaki, M.; Valderrama, R.; Palma, J.M.; Barroso, J.B.; Corpas, F.J. Function of S-nitrosoglutathione reductase (GSNOR) in plant development and under biotic/abiotic stress. Plant Signal. Behav. 2011, 6, 789–793. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate Reductase Regulates Plant Nitric Oxide Homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Yang, Z.; Gomez, A.; Liu, B.; Lin, C.; Oka, Y. Signaling mechanisms of plant cryptochromes in Arabidopsis thaliana. J. Plant Res. 2016, 129, 137–148. [Google Scholar] [CrossRef]
- Tripathi, S.; Hoang, Q.T.; Han, Y.J.; Kim, J.I. Regulation of photomorphogenic development by plant phytochromes. Int. J. Mol. Sci. 2019, 20, 6165. [Google Scholar] [CrossRef] [PubMed]
- Quail, P.H. Photosensory perception and signalling in plant cells: New paradigms? Curr. Opin. Cell Biol. 2002, 14, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Nambara, E.; Choi, G.; Yamaguchi, S. Interaction of light and hormone signals in germinating seeds. Plant Mol. Biol. 2009, 69, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Beligni, M.V.; Lamattina, L. Nitric oxide stimulates seed germination and de-etiolation, and inhibits hypocotyl elongation, three light-inducible responses in plants. Planta 2000, 210, 215–221. [Google Scholar] [CrossRef]
- Liu, J.; Xue, T.; Shen, Y.J.H. Effect of Nitric Oxide on Seed Germination and Dormancy in Empress Trees. Hort Technol. 2019, 29, 271–275. [Google Scholar] [CrossRef]
- Batak, I.; Dević, M.; Gibal, Z.; Grubišić, D.; Poff, K.L.; Konjević, R. The effects of potassium nitrate and NO-donors on phytochrome A-and phytochrome B-specific induced germination of Arabidopsis thaliana seeds. Seed Sci. Res. 2002, 12, 253–259. [Google Scholar] [CrossRef]
- Shen, H.; Moon, J.; Huq, E. PIF1 is regulated by light-mediated degradation through the ubiquitin-26S proteasome pathway to optimize photomorphogenesis of seedlings in Arabidopsis. Plant J. 2005, 44, 1023–1035. [Google Scholar] [CrossRef]
- Lozano-Juste, J.; León, J. Nitric oxide regulates DELLA content and PIF expression to promote photomorphogenesis in Arabidopsis. Plant Physiol. 2011, 156, 1410–1423. [Google Scholar] [CrossRef]
- Bai, S.; Yao, T.; Li, M.; Guo, X.; Zhang, Y.; Zhu, S.; He, Y. PIF3 is involved in the primary root growth inhibition of Arabidopsis induced by nitric oxide in the light. Mol. Plant 2014, 7, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Yamaguchi, S.; Lim, S.; Oh, E.; Park, J.; Hanada, A.; Kamiya, Y.; Choi, G. Corrigendum to: SOMNUS, a CCCH-Type Zinc Finger Protein in Arabidopsis, Negatively Regulates Light-Dependent Seed Germination Downstream of PIL5. Plant Cell 2021, 33, 2093–2095. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Zhong, S.; Mo, X.; Liu, N.; Nezames, C.D.; Deng, X.W. HFR1 sequesters PIF1 to govern the transcriptional network underlying light-initiated seed germination in Arabidopsis. Plant Cell 2013, 25, 3770–3784. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jia, Y.; Yu, L.; Yang, W.; Chen, Z.; Chen, H.; Hu, X. Nitric oxide promotes light-initiated seed germination by repressing PIF1 expression and stabilizing HFR1. Plant Physiol. Biochem. 2018, 123, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, J.V.; Barrero, J.M.; Hughes, T.; Julkowska, M.; Taylor, J.M.; Xu, Q.; Gubler, F. Roles for blue light, jasmonate and nitric oxide in the regulation of dormancy and germination in wheat grain (Triticum aestivum L.). Planta 2013, 238, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Piterková, J.; Luhová, L.; Hofman, J.; Turecková, V.; Novák, O.; Petrivalsky, M.; Fellner, M. Nitric oxide is involved in light-specific responses of tomato during germination under normal and osmotic stress conditions. Ann. Bot. 2012, 110, 767–776. [Google Scholar] [CrossRef]
- Wang, X. Regulatory functions of phospholipase D and phosphatidic acid in plant growth, development, and stress responses. Plant Physiol. 2005, 139, 566–573. [Google Scholar] [CrossRef]
- Berridge, M.J. Inositol trisphosphate and calcium signalling. Nature 1993, 361, 315–325. [Google Scholar] [CrossRef]
- An, Z.; Zhou, C.J. Light induces lettuce seed germination through promoting nitric oxide production and phospholipase D-derived phosphatidic acid formation. S. Afr. J. Bot. 2017, 108, 416–422. [Google Scholar] [CrossRef]
- Freschi, L. Nitric oxide and phytohormone interactions: Current status and perspectives. Front. Plant Sci. 2013, 4, 398. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, L.; Ye, N.; Liu, R.; Jia, W.; Zhang, J. Nitric oxide-induced rapid decrease of abscisic acid concentration is required in breaking seed dormancy in Arabidopsis. New Phytol. 2009, 183, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.G.; Chen, L.; Zhang, L.L.; Zhang, W.H. Nitric reductase-dependent nitric oxide production is involved in cold acclimation and freezing tolerance in Arabidopsis. Plant Physiol. 2009, 151, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Sarath, G.; Hou, G.; Baird, L.M.; Mitchell, R.B. ABA, ROS and NO are Key Players During Switchgrass Seed Germination. Plant Signal. Behav. 2007, 2, 492–493. [Google Scholar] [CrossRef]
- Sarath, G.; Hou, G.; Baird, L.M.; Mitchell, R.B. Reactive oxygen species, ABA and nitric oxide interactions on the germination of warm-season C4-grasses. Planta 2007, 226, 697–708. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, A.; Vandewiele, M.; de Rycke, R.; van Damme, J.; van Montagu, M.; Krebbers, E.; Vandekerckhove, J. Expression and Processing of an Arabidopsis 2S Albumin in Transgenic Tobacco. Plant Physiol. 1990, 92, 899–907. [Google Scholar] [CrossRef]
- Penfield, S.; Rylott, E.L.; Gilday, A.D.; Graham, S.; Larson, T.R.; Graham, I.A. Reserve mobilization in the Arabidopsis endosperm fuels hypocotyl elongation in the dark, is independent of abscisic acid, and requires PHOSPHOENOLPYRUVATE CARBOXYKINASE1. Plant Cell 2004, 16, 2705–2718. [Google Scholar] [CrossRef]
- Bethke, P.C.; Libourel, I.G.; Aoyama, N.; Chung, Y.Y.; Still, D.W.; Jones, R.L. The Arabidopsis aleurone layer responds to nitric oxide, gibberellin, and abscisic acid and is sufficient and necessary for seed dormancy. Plant Physiol. 2007, 143, 1173–1188. [Google Scholar] [CrossRef]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Front. Plant Sci. 2013, 4, 63. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, J. Rapid accumulation of NO regulates ABA catabolism and seed dormancy during imbibition in Arabidopsis. Plant Signal. Behav. 2009, 4, 905–907. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, R.; Zhao, M.; Wang, F.; Zhang, N.; Si, H. NO and ABA Interaction Regulates Tuber Dormancy and Sprouting in Potato. Front. Plant Sci. 2020, 11, 311. [Google Scholar] [CrossRef]
- Andryka-Dudek, P.; Ciacka, K.; Wiśniewska, A.; Bogatek, R.; Gniazdowska, A. Nitric Oxide-Induced Dormancy Removal of Apple Embryos Is Linked to Alterations in Expression of Genes Encoding ABA and JA Biosynthetic or Transduction Pathways and RNA Nitration. Int. J. Mol. Sci. 2019, 20, 1007. [Google Scholar] [CrossRef]
- Sun, L.R.; Yue, C.M.; Hao, F.S. Update on roles of nitric oxide in regulating stomatal closure. Plant Signal. Behav. 2019, 14, e1649569. [Google Scholar] [CrossRef] [PubMed]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The Role and Regulation of ABI5 (ABA-Insensitive 5) in Plant Development, Abiotic Stress Responses and Phytohormone Crosstalk. Front. Plant Sci. 2016, 7, 1884. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.; Considine, M.J. Nitric Oxide Enables Germination by a Four-Pronged Attack on ABA-Induced Seed Dormancy. Front. Plant Sci. 2018, 9, 296. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhu, J.K.; Lang, Z. Nitric oxide suppresses the inhibitory effect of abscisic acid on seed germination by S-nitrosylation of SnRK2 proteins. Plant Signal. Behav. 2015, 10, e1031939. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.J.; Md Isa, N.; Movahedi, M.; Lozano-Juste, J.; Mendiondo, G.M.; Berckhan, S.; Marín-de la Rosa, N.; Vicente Conde, J.; Sousa Correia, C.; Pearce, S.P.; et al. Nitric oxide sensing in plants is mediated by proteolytic control of group VII ERF transcription factors. Mol. Cell 2014, 53, 369–379. [Google Scholar] [CrossRef]
- Albertos, P.; Romero-Puertas, M.C.; Tatematsu, K.; Mateos, I.; Sánchez-Vicente, I.; Nambara, E.; Lorenzo, O. S-nitrosylation triggers ABI5 degradation to promote seed germination and seedling growth. Nat. Commun. 2015, 6, 8669. [Google Scholar] [CrossRef]
- Yang, S.F.; Hoffman, N.E. Ethylene biosynthesis and its regulation in higher plants. Annu. Rev. Plant Biol. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- Ju, C.; Chang, C. Mechanistic insights in ethylene perception and signal transduction. Plant Physiol. 2015, 169, 85–95. [Google Scholar] [CrossRef]
- Azhar, B.J.; Zulfiqar, A.; Shakeel, S.N.; Schaller, G.E. Amplification and adaptation in the ethylene signaling pathway. Small Methods 2020, 4, 1900452. [Google Scholar] [CrossRef]
- Corbineau, F.; Xia, Q.; Bailly, C.; El-Maarouf-Bouteau, H. Ethylene, a key factor in the regulation of seed dormancy. Front. Plant Sci. 2014, 5, 539. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Ortiz, S.; Garnica-Vergara, A.; Esparza-Reynoso, S.; García-Cárdenas, E.; Raya-González, J.; Francisco Ruiz-Herrera, L.; López-Bucio, J. Jasmonic acid-ethylene crosstalk via ETHYLENE INSENSITIVE 2 reprograms Arabidopsis root system architecture through nitric oxide accumulation. J. Plant Growth Regul. 2018, 37, 438–451. [Google Scholar] [CrossRef]
- Wang, H.Q.; Sun, L.P.; Wang, L.X.; Fang, X.W.; Li, Z.Q.; Zhang, F.F.; Hu, X.; Qi, C.; He, J.M. Ethylene mediates salicylic-acid-induced stomatal closure by controlling reactive oxygen species and nitric oxide production in Arabidopsis. Plant Sci. 2020, 294, 110464. [Google Scholar] [CrossRef] [PubMed]
- Gniazdowska, A.; Dobrzyńska, U.; Babańczyk, T.; Bogatek, R. Breaking the apple embryo dormancy by nitric oxide involves the stimulation of ethylene production. Planta 2007, 225, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Gniazdowska, A.; Krasuska, U.; Czajkowska, K.; Bogatek, R. Nitric oxide, hydrogen cyanide and ethylene are required in the control of germination and undisturbed development of young apple seedlings. Plant Growth Regul. 2010, 61, 75–84. [Google Scholar] [CrossRef]
- Gniazdowska, A.; Krasuska, U.; Bogatek, R. Dormancy removal in apple embryos by nitric oxide or cyanide involves modifications in ethylene biosynthetic pathway. Planta 2010, 232, 1397–1407. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tang, S.; Wang, J.; Shen, H.; Yang, L. Interaction between reactive oxygen species and hormones during the breaking of embryo dormancy in Sorbus pohuashanensis by exogenous nitric oxide. J. For. Res. 2022, 33, 435–444. [Google Scholar] [CrossRef]
- El-Maarouf-Bouteau, H.; Sajjad, Y.; Bazin, J.; Langlade, N.; Cristescu, S.M.; Balzergue, S.; Baudouin, E.; Bailly, C. Reactive oxygen species, abscisic acid and ethylene interact to regulate sunflower seed germination. Plant Cell Environ. 2015, 38, 364–374. [Google Scholar] [CrossRef]
- Dixon, K.; Merritt, D.; Flematti, G.; Ghisalberti, E.L. Karrikinolide—A phytoreactive compound derived from smoke with applications in horticulture, ecological restoration and agriculture. Acta Hortic. 2009, 813, 155–170. [Google Scholar] [CrossRef]
- Sami, A.; Riaz, M.W.; Zhou, X.; Zhu, Z.; Zhou, K. Alleviating dormancy in Brassica oleracea seeds using NO and KAR1 with ethylene biosynthetic pathway, ROS and antioxidant enzymes modifications. BMC Plant Biol. 2019, 19, 577. [Google Scholar] [CrossRef]
- Sami, A.; Rehman, S.; Tanvir, M.A.; Zhou, X.Y.; Zhu, Z.H.; Zhou, K. Assessment of the germination potential of Brassica oleracea seeds treated with karrikin 1 and cyanide, which modify the ethylene biosynthetic pathway. J. Plant Growth Regul. 2021, 40, 1257–1269. [Google Scholar] [CrossRef]
- Kępczyński, J. Gas-priming as a novel simple method of seed treatment with ethylene, hydrogen cyanide or nitric oxide. Acta Physiol. Plant. 2021, 43, 117. [Google Scholar] [CrossRef]
- Kara, Z.; Doğan, O.; Vergili, E. Sodium Nitroprusside and Gibberellin Effects on Seed Germination and Seedling Development of Grapevine (Vitis vinifera L.) Cvs. Ekşi Kara and Gök Üzüm. Erwerbs-Obstbau 2020, 62, 61–68. [Google Scholar] [CrossRef]
- Kępczyński, J.; Sznigir, P. Response of Amaranthus retroflexus L. seeds to gibberellic acid, ethylene and abscisic acid depending on duration of stratification and burial. Plant Growth Regul. 2013, 70, 15–26. [Google Scholar] [CrossRef]
- Kępczyński, J.; Sznigir, P. Participation of GA3, ethylene, NO and HCN in germination of Amaranthus retroflexus L. seeds with various dormancy levels. Acta Physiol. Plant. 2014, 36, 1463–1472. [Google Scholar] [CrossRef]
- Kępczyński, J.; Cembrowska-Lech, D.; Sznigir, P. Interplay between nitric oxide, ethylene, and gibberellic acid regulating the release of Amaranthus retroflexus seed dormancy. Acta Physiol. Plant. 2017, 39, 1–13. [Google Scholar] [CrossRef]
- Matilla, A.J. Polyamines and seed germination. Seed Sci. Res. 1996, 6, 81–93. [Google Scholar] [CrossRef]
- Silveira, V.; Santa-Catarina, C.; Tun, N.N.; Scherer, G.F.; Handro, W.; Guerra, M.P.; Floh, E.I. Polyamine effects on the endogenous polyamine contents, nitric oxide release, growth and differentiation of embryogenic suspension cultures of Araucaria angustifolia (Bert.) O. Ktze. Plant Sci. 2006, 171, 91–98. [Google Scholar] [CrossRef]
- Pal Bais, H.; Ravishankar, G.A. Role of polyamines in the ontogeny of plants and their biotechnological applications. Plant Cell, Tissue Organ Cult. 2002, 69, 1–34. [Google Scholar] [CrossRef]
- Cona, A.; Cenci, F.; Cervelli, M.; Federico, R.; Mariottini, P.; Moreno, S.; Angelini, R. Polyamine oxidase, a hydrogen peroxide-producing enzyme, is up-regulated by light and down-regulated by auxin in the outer tissues of the maize mesocotyl. Plant Physiol. 2003, 131, 803–813. [Google Scholar] [CrossRef]
- Krasuska, U.; Ciacka, K.; Bogatek, R.; Gniazdowska, A.J. Polyamines and Nitric Oxide Link in Regulation of Dormancy Removal and Germination of Apple (Malus domestica Borkh.) Embryos. J. Plant Growth Regul. 2014, 33, 590–601. [Google Scholar] [CrossRef]
- Krasuska, U.; Ciacka, K.; Gniazdowska, A. Nitric oxide-polyamines cross-talk during dormancy release and germination of apple embryos. Nitric Oxide 2017, 68, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.Q.; Okamoto, M.; Crawford, N.M. Identification of a plant nitric oxide synthase gene involved in hormonal signaling. Science 2003, 302, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.; Xiao, S.; Yu, L.; Fan, L.M. Expression of a rice gene OsNOA1 re-establishes nitric oxide synthesis and stress-related gene expression for salt tolerance in Arabidopsis nitric oxide-associated 1 mutant Atnoa1. Environ. Exp. Bot. 2009, 65, 90–98. [Google Scholar] [CrossRef]
- Patel, P.; Kadur Narayanaswamy, G.; Kataria, S.; Baghel, L. Involvement of nitric oxide in enhanced germination and seedling growth of magnetoprimed maize seeds. Plant Signal. Behav. 2017, 12, e1293217. [Google Scholar] [CrossRef] [PubMed]
- Kataria, S.; Anand, A.; Raipuria, R.K.; Kumar, S.; Jain, M.; Watts, A.; Brestic, M. Magnetopriming Actuates Nitric Oxide Synthesis to Regulate Phytohormones for Improving Germination of Soybean Seeds under Salt Stress. Cells 2022, 11, 2174. [Google Scholar] [CrossRef]
- Shi, Q.; Ding, F.; Wang, X.; Wei, M. Exogenous nitric oxide protect cucumber roots against oxidative stress induced by salt stress. Plant Physiol. Biochem. 2007, 45, 542–550. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Basalah, M.O. Role of nitric oxide in tolerance of plants to abiotic stress. Protoplasma 2011, 248, 447–455. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, W.; He, J.; Zhang, L.; Wei, Y.; Yang, M. Nitric oxide alleviates salt stress in seed germination and early seedling growth of pakchoi (Brassica chinensis L.) by enhancing physiological and biochemical parameters. Ecotoxicol. Environ. Saf. 2020, 187, 109785. [Google Scholar] [CrossRef]
- Zhai, J.; Liang, Y.; Zeng, S.; Yan, J.; Li, K.; Xu, H. Overexpression of tomato glutathione reductase (SlGR) in transgenic tobacco enhances salt tolerance involving the S-nitrosylation of GR. Plant Physiol. Biochem. 2023, 196, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.L.D.; Pinheiro, D.T.; Borges, E.E.D.L.; Silva, L.J.D.; Dias, D.C.F.D.S. Effect of cyanide by sodium nitroprusside (SNP) application on germination, antioxidative system and lipid peroxidation of Senna macranthera seeds under saline stress. J. Seed Sci. 2019, 41, 086–096. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Skalicky, M.; Brestic, M.; Pavla, V. Cross-talk between nitric oxide, hydrogen peroxide and calcium in salt-stressed Chenopodium quinoa Willd. At seed germination stage. Plant Physiol. Biochem. 2020, 154, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Ikeya, S.; Aoyanagi, T.; Ishizuka, I.; Takeuchi, A.; Kozaki, A. Nitrate Promotes Germination Under Inhibition by NaCl or High Concentration of Glucose. Plants 2020, 9, 707. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, F.; Naeem, M.; Akram, A.; Ashraf, M.Y.; Ahmad, K.S.; Zulfiqar, B.; Sardar, H.; Shabbir, R.N.; Majeed, S.; Shehzad, M.A.; et al. Seed priming with KNO3 mediates biochemical processes to inhibit lead toxicity in maize (Zea mays L.). J. Sci. Food Agric. 2017, 97, 4780–4789. [Google Scholar] [CrossRef]
- Castaings, L.; Camargo, A.; Pocholle, D.; Gaudon, V.; Texier, Y.; Boutet-Mercey, S.; Taconnat, L.; Renou, J.P.; Daniel-Vedele, F.; Fernandez, E.; et al. The nodule inception-like protein 7 modulates nitrate sensing and metabolism in Arabidopsis. Plant J. 2009, 57, 426–435. [Google Scholar] [CrossRef]
- Guan, P.; Wang, R.; Nacry, P.; Breton, G.; Kay, S.A.; Pruneda-Paz, J.L.; Davani, A.; Crawford, N.M. Nitrate foraging by Arabidopsis roots is mediated by the transcription factor TCP20 through the systemic signaling pathway. Proc. Natl. Acad. Sci. USA 2014, 111, 15267–15272. [Google Scholar] [CrossRef]
- Zhang, H.; Forde, B.G. An Arabidopsis MADS box gene that controls nutrient-induced changes in root architecture. Science 1998, 279, 407–409. [Google Scholar] [CrossRef]
- Konishi, M.; Yanagisawa, S. Arabidopsis NIN-like transcription factors have a central role in nitrate signalling. Nat. Commun. 2013, 4, 1617. [Google Scholar] [CrossRef]
- Fang, X.Z.; Fang, S.Q.; Ye, Z.Q.; Liu, D.; Zhao, K.L.; Jin, C.W. NRT1.1 Dual-Affinity Nitrate Transport/Signalling and its Roles in Plant Abiotic Stress Resistance. Front. Plant Sci. 2021, 12, 715694. [Google Scholar] [CrossRef]
- Álvarez-Aragón, R.; Haro, R.; Benito, B.; Rodríguez-Navarro, A. Salt intolerance in Arabidopsis: Shoot and root sodium toxicity, and inhibition by sodium-plus-potassium overaccumulation. Planta 2016, 243, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.X.; Zhu, Y.X.; Fang, X.Z.; Ye, J.Y.; Du, W.X.; Zhu, Q.Y.; Lin, X.Y.; Jin, C.W. Ammonium aggravates salt stress in plants by entrapping them in a chloride over-accumulation state in an NRT1.1-dependent manner. Sci. Total Environ. 2020, 746, 141244. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.H.; Yu, L.H.; Xiang, C.B. ARABIDOPSIS NITRATE REGULATED 1 acts as a negative modulator of seed germination by activating ABI3 expression. New Phytol. 2020, 225, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Cheng, Y.H.; Chen, K.E.; Tsay, Y.F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef]
- Konishi, M.; Okitsu, T.; Yanagisawa, S. Nitrate-responsive NIN-like protein transcription factors perform unique and redundant roles in Arabidopsis. J. Exp. Bot. 2021, 72, 5735–5750. [Google Scholar] [CrossRef]
- Liu, K.H.; Niu, Y.; Konishi, M.; Wu, Y.; Du, H.; Sun Chung, H.; Li, L.; Boudsocq, M.; McCormack, M.; Maekawa, S.; et al. Discovery of nitrate-CPK-NLP signalling in central nutrient-growth networks. Nature 2017, 545, 311–316. [Google Scholar] [CrossRef]
- Yan, D.; Easwaran, V.; Chau, V.; Okamoto, M.; Ierullo, M.; Kimura, M.; Endo, A.; Yano, R.; Pasha, A.; Gong, Y.; et al. NIN-like protein 8 is a master regulator of nitrate-promoted seed germination in Arabidopsis. Nat. Commun. 2016, 7, 13179. [Google Scholar] [CrossRef]
- Gupta, K.J.; Fernie, A.R.; Kaiser, W.M.; van Dongen, J.T. On the origins of nitric oxide. Trends Plant Sci. 2011, 16, 160–168. [Google Scholar] [CrossRef]
- Reda, M.; Golicka, A.; Kabała, K.; Janicka, M. Involvement of NR and PM-NR in NO biosynthesis in cucumber plants subjected to salt stress. Plant Sci. 2018, 267, 55–64. [Google Scholar] [CrossRef]
- Yi, Y.; Peng, Y.; Song, T.; Lu, S.; Teng, Z.; Zheng, Q.; Zhao, F.; Meng, S.; Liu, B.; Peng, Y.; et al. NLP2-NR Module Associated NO Is Involved in Regulating Seed Germination in Rice under Salt Stress. Plants 2022, 11, 795. [Google Scholar] [CrossRef]
- Marchive, C.; Roudier, F.; Castaings, L.; Bréhaut, V.; Blondet, E.; Colot, V.; Meyer, C.; Krapp, A. Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants. Nat. Commun. 2013, 4, 1713. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.; Ripoll, J.J.; Wang, R.; Vuong, L.; Bailey-Steinitz, L.J.; Ye, D.; Crawford, N.M. Interacting TCP and NLP transcription factors control plant responses to nitrate availability. Proc. Natl. Acad. Sci. USA 2017, 114, 2419–2424. [Google Scholar] [CrossRef] [PubMed]
- Le, Q.T.; Lee, W.J.; Choi, J.H.; Nguyen, D.T.; Truong, H.A.; Lee, S.A.; Hong, S.W.; Lee, H. The Loss of Function of the NODULE INCEPTION-Like PROTEIN 7 Enhances Salt Stress Tolerance in Arabidopsis Seedlings. Front. Plant Sci. 2021, 12, 743832. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Yang, L.; Paul, M.; Zu, Y.; Tang, Z. Ethylene promotes germination of Arabidopsis seed under salinity by decreasing reactive oxygen species: Evidence for the involvement of nitric oxide simulated by sodium nitroprusside. Plant Physiol. Biochem. 2013, 73, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Pan, Y.; Chang, B.; Wang, Y.; Tang, Z. NO Promotes Seed Germination and Seedling Growth Under High Salt May Depend on EIN3 Protein in Arabidopsis. Front. Plant Sci. 2015, 6, 1203. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, C.; Wang, X.; Shen, H.; Yang, L. Exogenous Ethylene Alleviates the Inhibition of Sorbus pohuashanensis Embryo Germination in a Saline-Alkali Environment (NaHCO3). Int. J. Mol. Sci. 2023, 24, 4244. [Google Scholar] [CrossRef]
- Garcia-Mata, C.; Lamattina, L. Abscisic acid (ABA) inhibits light-induced stomatal opening through calcium- and nitric oxide-mediated signaling pathways. Nitric Oxide 2007, 17, 143–151. [Google Scholar] [CrossRef]
- Foresi, N.; Mayta, M.L.; Lodeyro, A.F.; Scuffi, D.; Correa-Aragunde, N.; García-Mata, C.; Casalongué, C.; Carrillo, N.; Lamattina, L. Expression of the tetrahydrofolate-dependent nitric oxide synthase from the green alga Ostreococcus tauri increases tolerance to abiotic stresses and influences stomatal development in Arabidopsis. Plant J. 2015, 82, 891–903. [Google Scholar] [CrossRef]
- Albertos, P.; Tatematsu, K.; Mateos, I.; Sánchez-Vicente, I.; Fernández-Arbaizar, A.; Nakabayashi, K.; Nambara, E.; Godoy, M.; Franco, J.M.; Solano, R.; et al. Redox feedback regulation of ANAC089 signaling alters seed germination and stress response. Cell Rep. 2021, 35, 109263. [Google Scholar] [CrossRef]
- Malcheska, F.; Ahmad, A.; Batool, S.; Müller, H.M.; Ludwig-Müller, J.; Kreuzwieser, J.; Randewig, D.; Hänsch, R.; Mendel, R.R.; Hell, R.; et al. Drought-Enhanced Xylem Sap Sulfate Closes Stomata by Affecting ALMT12 and Guard Cell ABA Synthesis. Plant Physiol. 2017, 174, 798–814. [Google Scholar] [CrossRef]
- Lozano-Juste, J.; León, J. Enhanced abscisic acid-mediated responses in nia1nia2noa1-2 triple mutant impaired in NIA/NR- and AtNOA1-dependent nitric oxide biosynthesis in Arabidopsis. Plant Physiol. 2010, 152, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.; Griffiths, R.; Hancock, J.; Neill, S. A new role for an old enzyme: Nitrate reductase-mediated nitric oxide generation is required for abscisic acid-induced stomatal closure in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2002, 99, 16314–16318. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.M.; Desikan, R.; Bright, J.; Confraria, A.; Harrison, J.; Hancock, J.T.; Barros, R.S.; Neill, S.J.; Wilson, I.D. Differential requirement for NO during ABA-induced stomatal closure in turgid and wilted leaves. Plant Cell Environ. 2009, 32, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lau, E.; Lam, M.P.Y.; Chu, H.; Li, S.; Huang, G.; Guo, P.; Wang, J.; Jiang, L.; Chu, I.K.; et al. OsNOA1/RIF1 is a functional homolog of AtNOA1/RIF1: Implication for a highly conserved plant cGTPase essential for chloroplast function. New Phytol. 2010, 187, 83–105. [Google Scholar] [CrossRef]
- Hartman, S.; Liu, Z.; van Veen, H.; Vicente, J.; Reinen, E.; Martopawiro, S.; Zhang, H.; van Dongen, N.; Bosman, F.; Bassel, G.W.; et al. Ethylene-mediated nitric oxide depletion pre-adapts plants to hypoxia stress. Nat. Commun. 2019, 10, 4020. [Google Scholar] [CrossRef]
- Castillo, M.C.; Costa-Broseta, Á.; Gayubas, B.; León, J. NIN-like protein7 and PROTEOLYSIS6 functional interaction enhances tolerance to sucrose, ABA, and submergence. Plant Physiol. 2021, 187, 2731–2748. [Google Scholar] [CrossRef]
- Krasuska, U.; Gniazdowska, A. Nitric oxide and hydrogen cyanide as regulating factors of enzymatic antioxidant system in germinating apple embryos. Acta Physiol. Plant. 2012, 34, 683–692. [Google Scholar] [CrossRef]
- Oracz, K.; El-Maarouf-Bouteau, H.; Kranner, I.; Bogatek, R.; Corbineau, F.; Bailly, C. The mechanisms involved in seed dormancy alleviation by hydrogen cyanide unravel the role of reactive oxygen species as key factors of cellular signaling during germination. Plant Physiol. 2009, 150, 494–505. [Google Scholar] [CrossRef]
- Dębska, K.; Krasuska, U.; Budnicka, K.; Bogatek, R.; Gniazdowska, A. Dormancy removal of apple seeds by cold stratification is associated with fluctuation in H2O2, NO production and protein carbonylation level. J. Plant Physiol. 2013, 170, 480–488. [Google Scholar] [CrossRef]
- Ciacka, K.; Tyminski, M.; Gniazdowska, A.; Krasuska, U. Cold stratification-induced dormancy removal in apple (Malus domestica Borkh.) seeds is accompanied by an increased glutathione pool in embryonic axes. J. Plant Physiol. 2022, 274, 153736. [Google Scholar] [CrossRef]
- Broniowska, K.A.; Diers, A.R.; Hogg, N. S-nitrosoglutathione. Biochim. Biophys. Acta 2013, 1830, 3173–3181. [Google Scholar] [CrossRef] [PubMed]
- Durner, J.; Klessig, D.F. Nitric oxide as a signal in plants. Curr. Opin. Plant Biol. 1999, 2, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Jensen, D.E.; Belka, G.K.; Du Bois, G.C. S-Nitrosoglutathione is a substrate for rat alcohol dehydrogenase class III isoenzyme. Biochem. J. 1998, 331, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Ciacka, K.; Krasuska, U.; Otulak-Kozieł, K.; Gniazdowska, A. Dormancy removal by cold stratification increases glutathione and S-nitrosoglutathione content in apple seeds. Plant Physiol. Biochem. 2019, 138, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.H.; Sami, A.; Xu, Q.Q.; Wu, L.L.; Zheng, W.Y.; Chen, Z.P.; Jin, X.Z.; Zhang, H.; Li, Y.; Yu, Y.; et al. Effects of seed priming treatments on the germination and development of two rapeseed (Brassica napus L.) varieties under the co-influence of low temperature and drought. PLoS ONE 2021, 16, e0257236. [Google Scholar] [CrossRef]
- Amooaghaie, R.; Nikzad, K. The role of nitric oxide in priming-induced low-temperature tolerance in two genotypes of tomato. Seed Sci. Res. 2013, 23, 123–131. [Google Scholar] [CrossRef]
- Bian, L.; Yang, L.; Wang, J.A.; Shen, H.L. Effects of KNO3 pretreatment and temperature on seed germination of Sorbus pohuashanensis. J. For. Res. 2013, 24, 309–316. [Google Scholar] [CrossRef]
- Majláth, I.; Éva, C.; Hamow, K.Á.; Kun, J.; Pál, M.; Rahman, A.; Palla, B.; Nagy, Z.; Gyenesei, A.; Szalai, G.; et al. Methylglyoxal induces stress signaling and promotes the germination of maize at low temperature. Physiol. Plant. 2022, 174, e13609. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; Bhardwaj, A.; Bhandari, K.; Kumar, S.; Prasad, P.V.; Jha, U.; Siddique, K.H.M.; Nayyar, H. Nitric oxide secures reproductive efficiency in heat-stressed lentil (Lens culinaris Medik.) plants by enhancing the photosynthetic ability to improve yield traits. Physiol. Mol. Biol. Plants 2021, 27, 2549–2566. [Google Scholar] [CrossRef]
- Wei, W.; Hu, Y.; Yang, W.; Li, X.; Wei, J.; Hu, X.; Li, P. S-Nitrosoglutathion reductase activity modulates the thermotolerance of seeds germination by controlling ABI5 stability under high temperature. Phyton-Int. J. Exp. Bot. 2021, 90, 1075. [Google Scholar] [CrossRef]
Figure 1.
A model for the synthesis and decomposition of nitric oxide. NO is synthesized mainly through a nitrate/nitrite-dependent reduction pathway or through an NO synthase (NOS)-mediated oxidative pathway. NO is decomposed mainly by interacting with oxygen, ROS or GSH. NR; NO-forming nitrite reductase (NOFNiR); mitochondrial nitrite reductase electron transport chain (mETC); polyamines (PAs); GSNO reductase (GSNOR).
Figure 1.
A model for the synthesis and decomposition of nitric oxide. NO is synthesized mainly through a nitrate/nitrite-dependent reduction pathway or through an NO synthase (NOS)-mediated oxidative pathway. NO is decomposed mainly by interacting with oxygen, ROS or GSH. NR; NO-forming nitrite reductase (NOFNiR); mitochondrial nitrite reductase electron transport chain (mETC); polyamines (PAs); GSNO reductase (GSNOR).
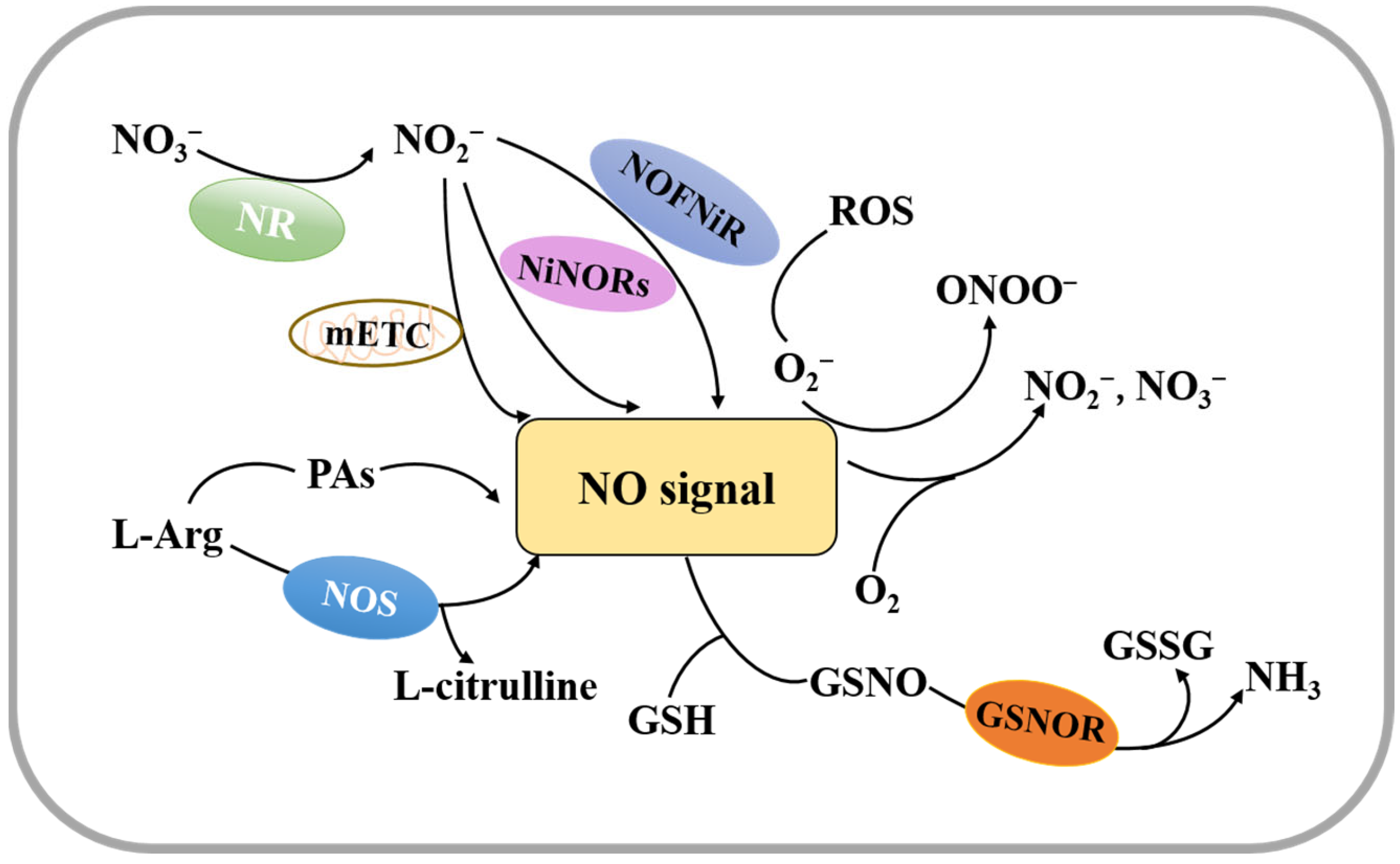
Figure 2.
The effect of nitric oxide on seed germination under light. In the presence of red light, PHYB moves from the cytoplasm to the nucleus and promotes seed germination promoting expression of the 26S proteasome pathway leading to PIF1 degradation. NO accumulation under red light conditions not only inhibited PIF1 transcription but also enhanced the HFR1-PIF1 interaction, ultimately leading to a weakened inhibitory effect of PIF1 on seed germination. SOM as a direct target gene downstream of PIF1, can affect the expression levels of related genes in the ABA and GA signaling pathways in photochrome-dependent photomorphogenesis. Light-mediated an antagonism relationship between NO and GA. During seed germination, NO content and PLD activity are increased by light. The arrows and bars indicate positive and inhibitory effects, respectively. PHYTOCHROME-INTERACTION FACTOR 1 (PIF1); LONG HYPOCOTYL IN FAR-RED (HFR1); phospholipase D (PLD); phosphatidic acid (PA); ABA-DEFICIENT1 (ABA1); NINE-CIS-EPOXYCAROTENOID DEOXYGENASE6 (NCED6); GIBBERELLIN 2-OXIDASE2 (GA2ox2); SOM (SOMNUS).
Figure 2.
The effect of nitric oxide on seed germination under light. In the presence of red light, PHYB moves from the cytoplasm to the nucleus and promotes seed germination promoting expression of the 26S proteasome pathway leading to PIF1 degradation. NO accumulation under red light conditions not only inhibited PIF1 transcription but also enhanced the HFR1-PIF1 interaction, ultimately leading to a weakened inhibitory effect of PIF1 on seed germination. SOM as a direct target gene downstream of PIF1, can affect the expression levels of related genes in the ABA and GA signaling pathways in photochrome-dependent photomorphogenesis. Light-mediated an antagonism relationship between NO and GA. During seed germination, NO content and PLD activity are increased by light. The arrows and bars indicate positive and inhibitory effects, respectively. PHYTOCHROME-INTERACTION FACTOR 1 (PIF1); LONG HYPOCOTYL IN FAR-RED (HFR1); phospholipase D (PLD); phosphatidic acid (PA); ABA-DEFICIENT1 (ABA1); NINE-CIS-EPOXYCAROTENOID DEOXYGENASE6 (NCED6); GIBBERELLIN 2-OXIDASE2 (GA2ox2); SOM (SOMNUS).
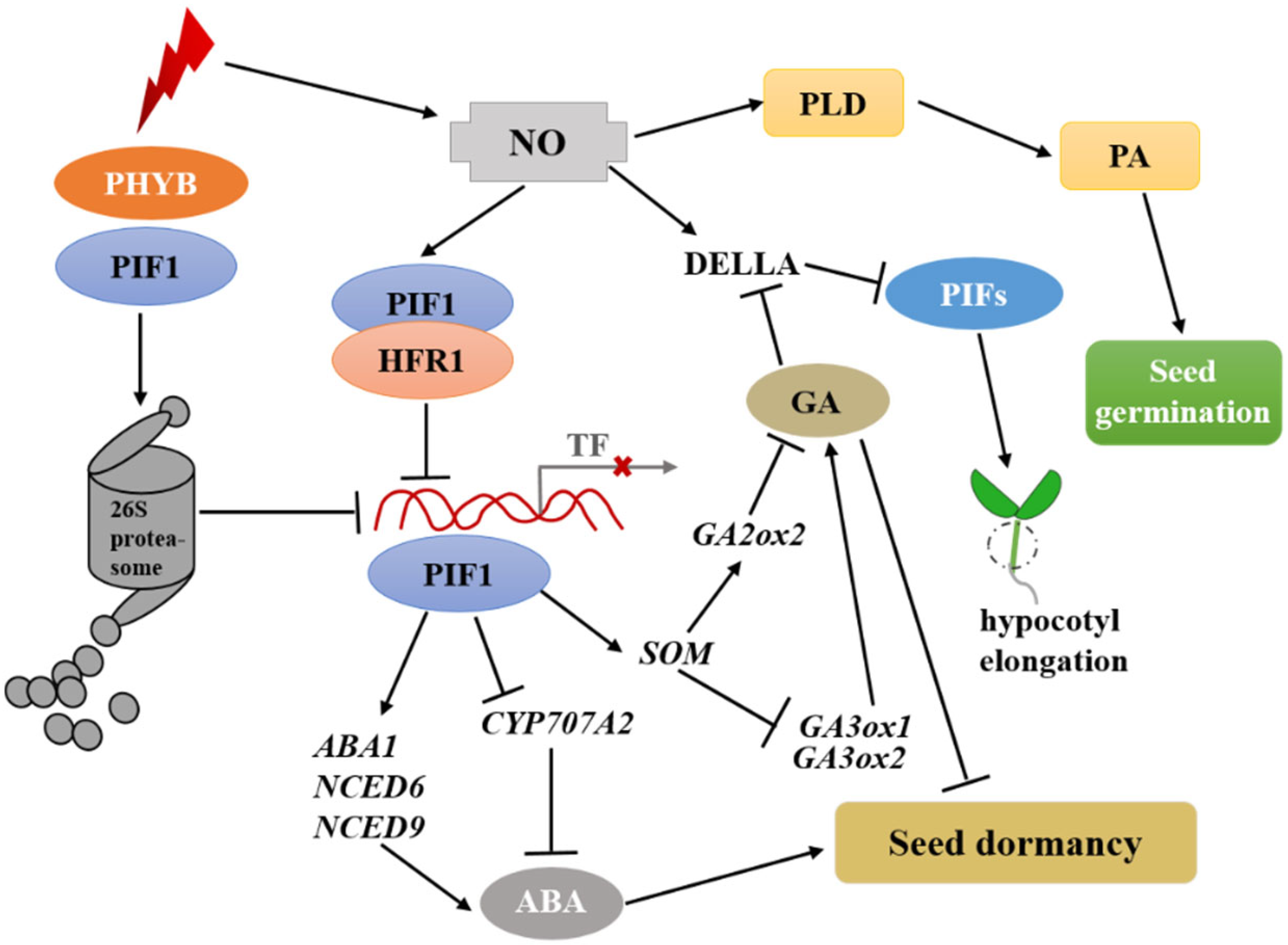
Figure 3.
A model of NO-ABA regulation on seed germination. NO induces ABA catabolism and inhibits ABA synthesis: nine-cis-epoxycarotenoid dioxygenase (NCED); short-chain dehydrogenase reductase (SDR1), abscisic aldehyde oxidase 3 (AAO3); ABA DEFICIENT3 (ABA3); phaseic acid (PA); dihydrophaseic acid (DPA); type 2C protein phosphatases (PP2C); SNF1-related protein kinase (SnRKs); ABSCISIC ACID INSENSITIVE5 (ABI5); Group VII of the ethylene response factor (ERF/AP2) family (ERFVIIs).
Figure 3.
A model of NO-ABA regulation on seed germination. NO induces ABA catabolism and inhibits ABA synthesis: nine-cis-epoxycarotenoid dioxygenase (NCED); short-chain dehydrogenase reductase (SDR1), abscisic aldehyde oxidase 3 (AAO3); ABA DEFICIENT3 (ABA3); phaseic acid (PA); dihydrophaseic acid (DPA); type 2C protein phosphatases (PP2C); SNF1-related protein kinase (SnRKs); ABSCISIC ACID INSENSITIVE5 (ABI5); Group VII of the ethylene response factor (ERF/AP2) family (ERFVIIs).

Figure 4.
Interaction of NO with various phytohormones in the regulation of seed germination. NO promoted the GA, JA and ethylene synthesis pathways and inhibited the ABA synthesis pathway, respectively. methionine (Met); S-adenosyl-L-methionine synthetase (SAM synthetase); S-adenosyl-methionine (S-AdoMet); 1-aminocyclopropane-1-carboxylic acid (ACC); 5′-methylthioadenosine (MTA); S-adenosyl-L-methionine methylthioadenosine-lyase (ACS), ACC oxidase (ACO); hydrogen cyanide (HCN); ethylene resistant 1 (ETR1); constitutive triple response 1 (CTR1); ETHYLENE INSENSITIVE2 (EIN2); ethylene-responsive factor (ERF); JA methylester (MeJA) and JA-isoleucine (JA-Ile); Polyamines (PAs); SAM (dcSAM); SAM decarboxylase (SAMDC); PA oxidase (PAO).
Figure 4.
Interaction of NO with various phytohormones in the regulation of seed germination. NO promoted the GA, JA and ethylene synthesis pathways and inhibited the ABA synthesis pathway, respectively. methionine (Met); S-adenosyl-L-methionine synthetase (SAM synthetase); S-adenosyl-methionine (S-AdoMet); 1-aminocyclopropane-1-carboxylic acid (ACC); 5′-methylthioadenosine (MTA); S-adenosyl-L-methionine methylthioadenosine-lyase (ACS), ACC oxidase (ACO); hydrogen cyanide (HCN); ethylene resistant 1 (ETR1); constitutive triple response 1 (CTR1); ETHYLENE INSENSITIVE2 (EIN2); ethylene-responsive factor (ERF); JA methylester (MeJA) and JA-isoleucine (JA-Ile); Polyamines (PAs); SAM (dcSAM); SAM decarboxylase (SAMDC); PA oxidase (PAO).
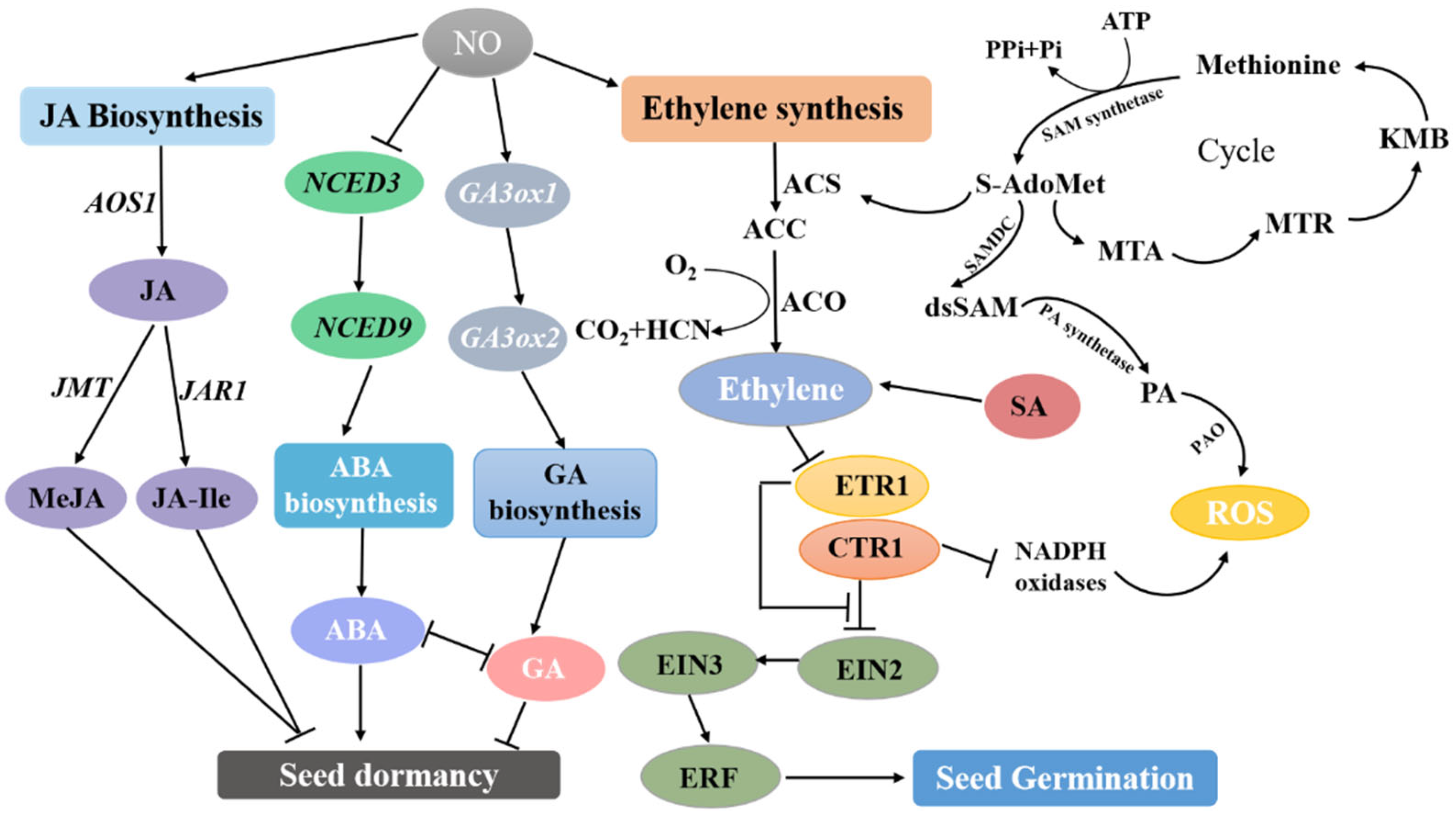
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Y.; Wang, R.; Wang, X.; Zhao, C.; Shen, H.; Yang, L. Nitric Oxide Regulates Seed Germination by Integrating Multiple Signalling Pathways. Int. J. Mol. Sci. 2023, 24, 9052. https://doi.org/10.3390/ijms24109052
AMA Style
Zhang Y, Wang R, Wang X, Zhao C, Shen H, Yang L. Nitric Oxide Regulates Seed Germination by Integrating Multiple Signalling Pathways. International Journal of Molecular Sciences. 2023; 24(10):9052. https://doi.org/10.3390/ijms24109052
Chicago/Turabian StyleZhang, Yue, Ruirui Wang, Xiaodong Wang, Caihong Zhao, Hailong Shen, and Ling Yang. 2023. "Nitric Oxide Regulates Seed Germination by Integrating Multiple Signalling Pathways" International Journal of Molecular Sciences 24, no. 10: 9052. https://doi.org/10.3390/ijms24109052
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.