
Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging


Abstract
1. Introduction
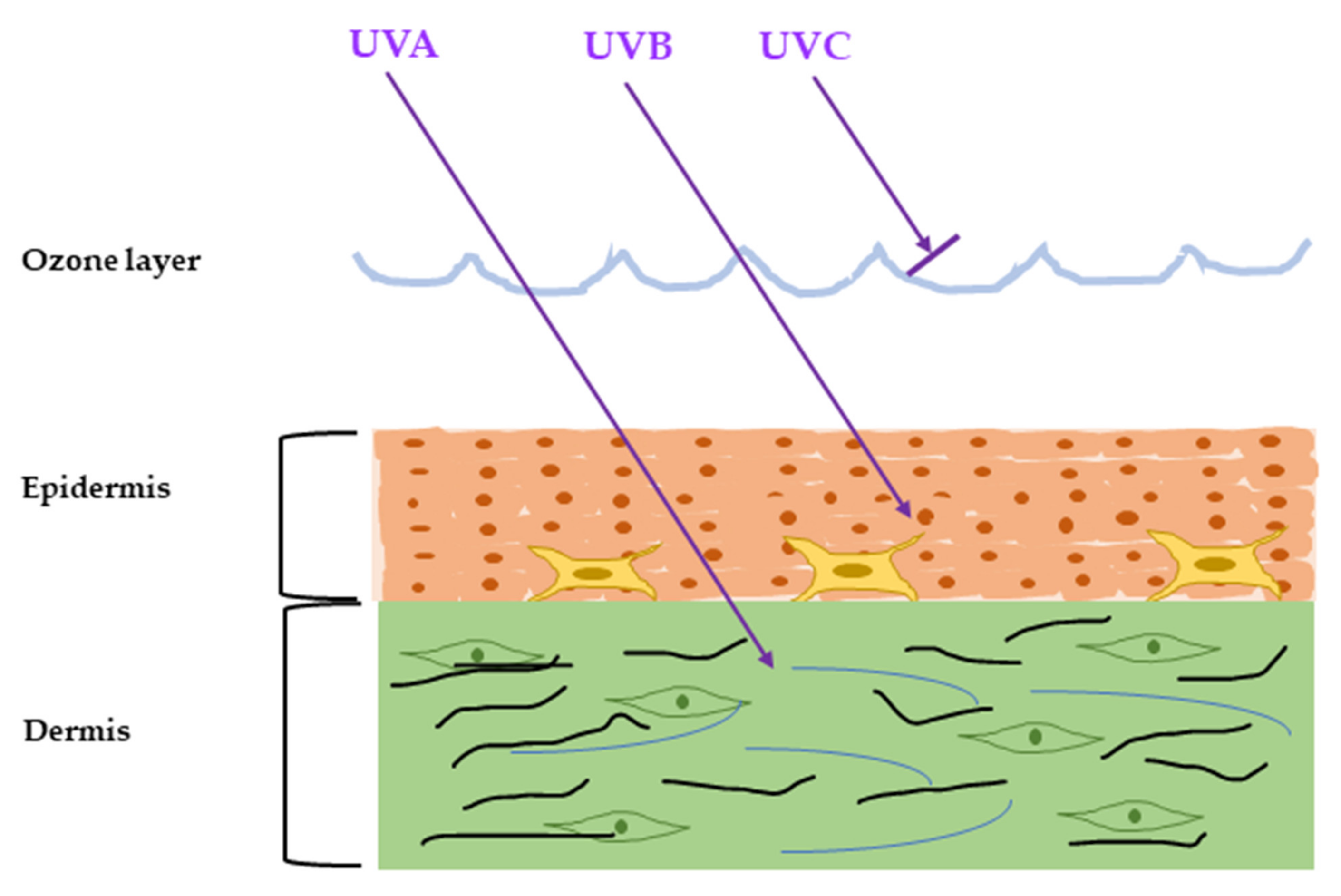
2. Inflammatory Response Caused by UVR
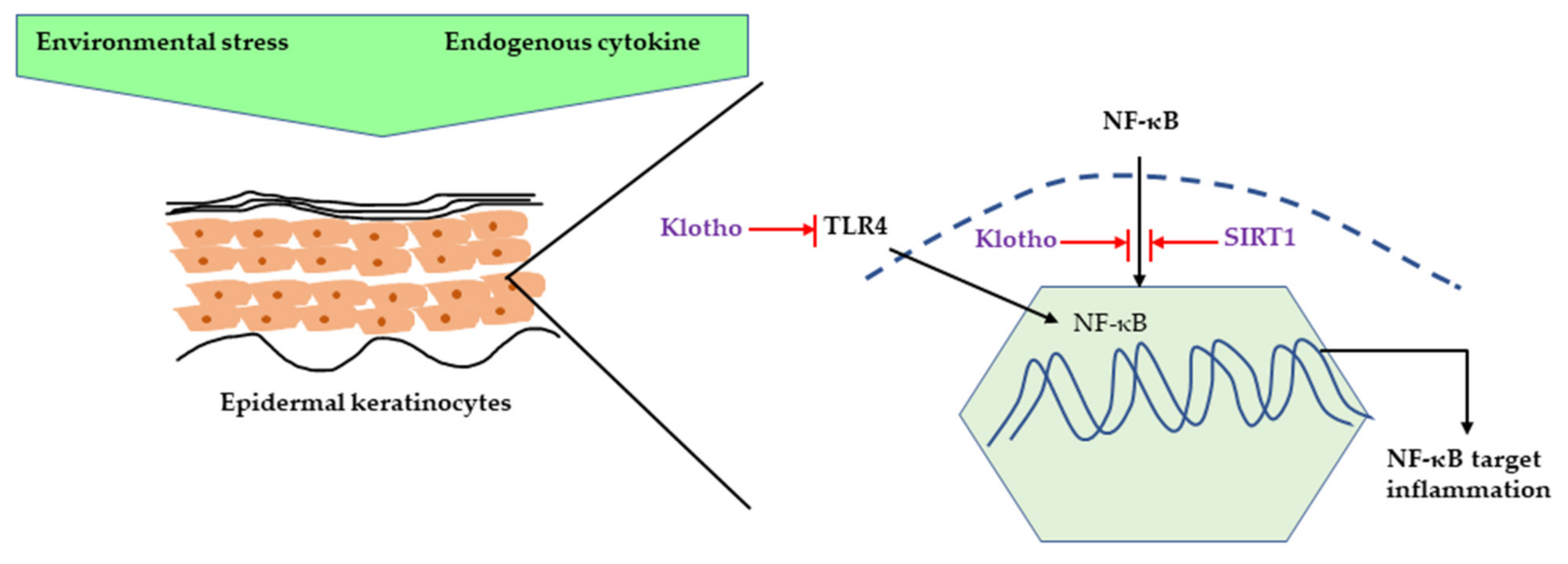
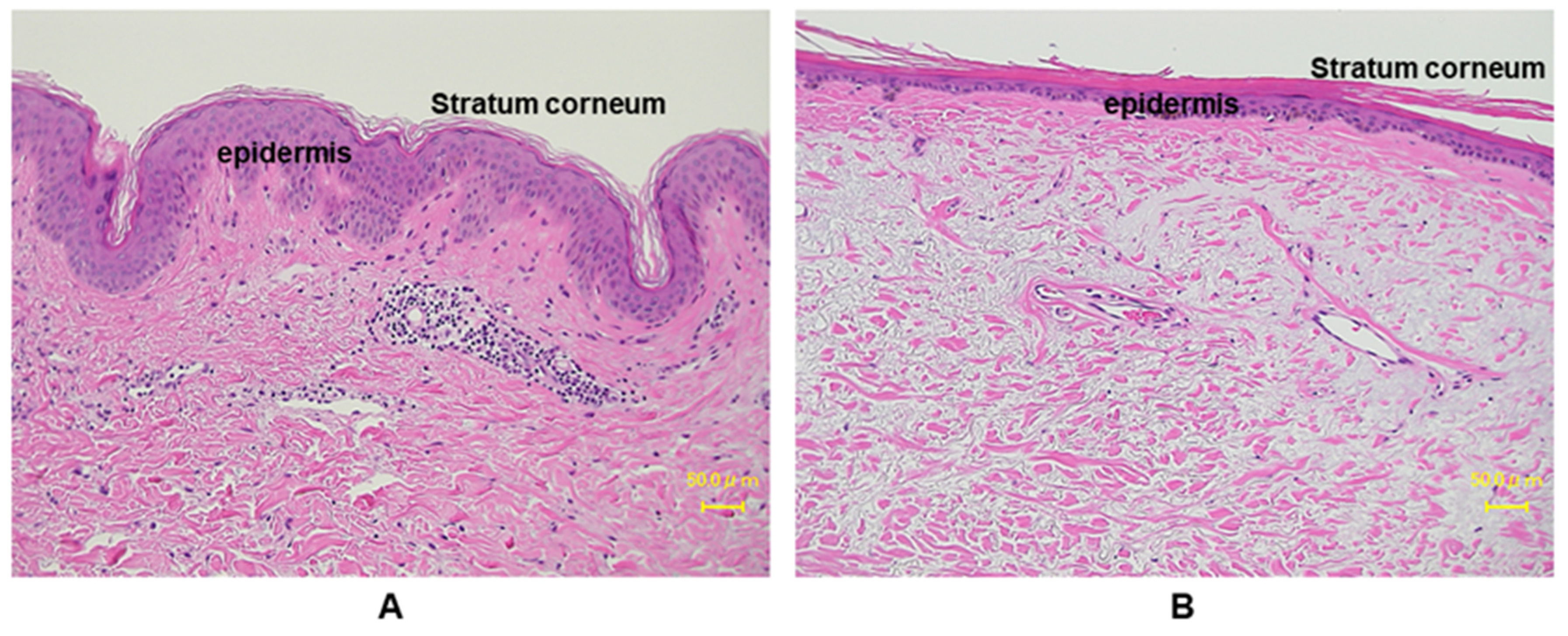
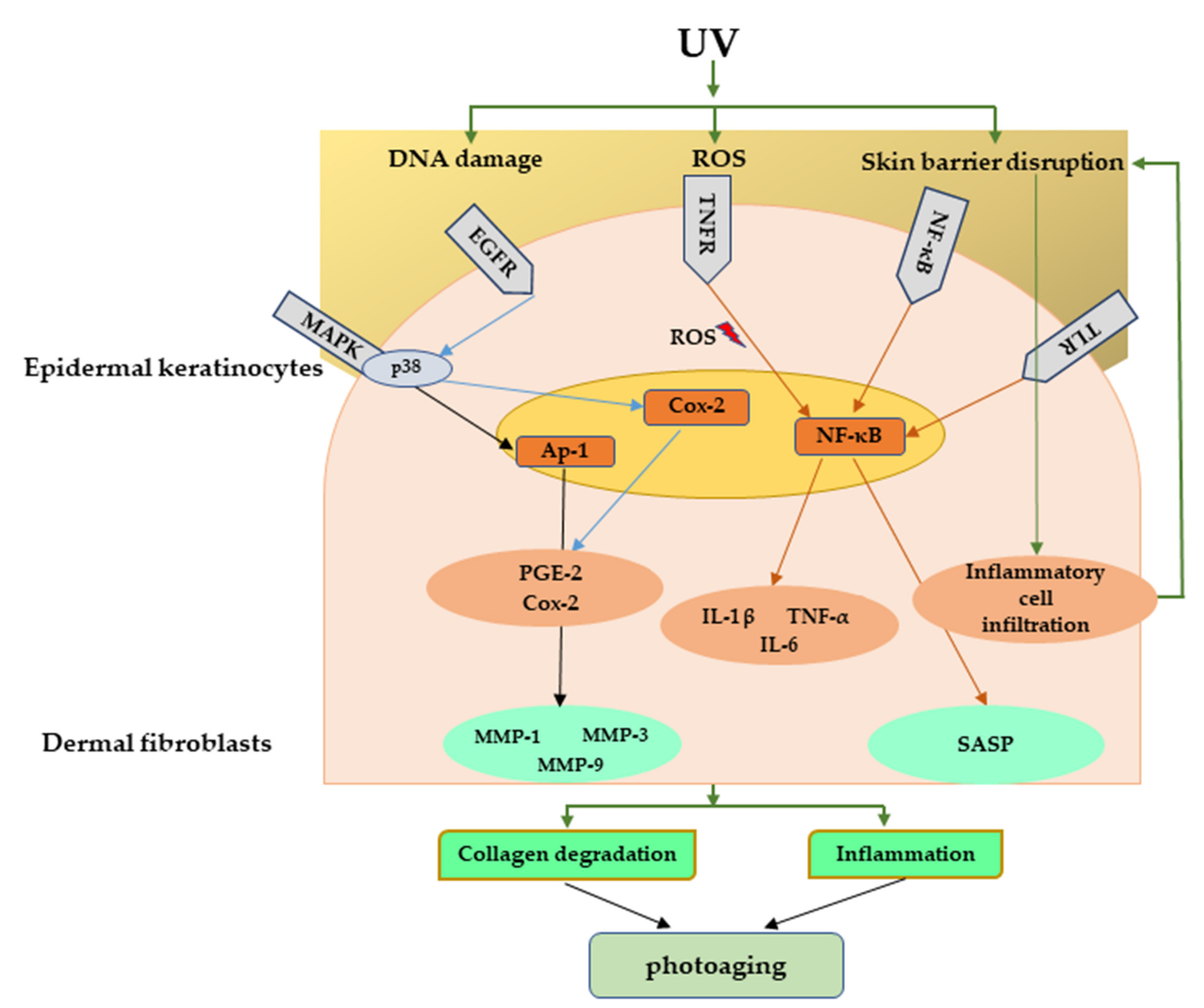
2.1. Major Inflammatory Responses in Epidermis upon UV Exposure
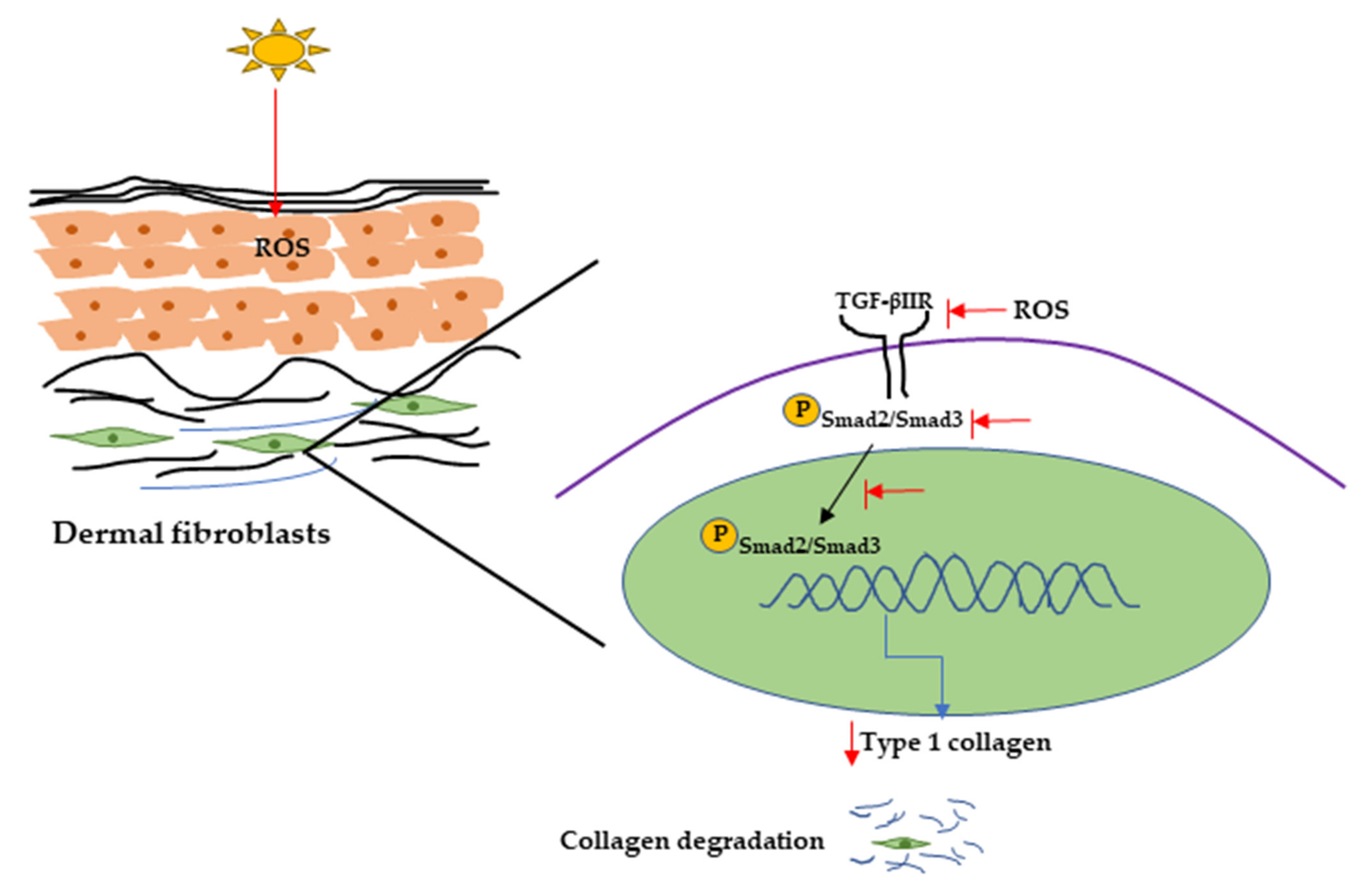
2.2. Major Inflammatory Responses in Dermis upon UV Exposure
3. Conclusions
Funding
Conflicts of Interest
Abbreviations
| UVR | Ultraviolet radiation |
| ROS | Reactive oxygen species |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| HaCaT | Human epidermal keratinocytes |
| IL | Interleukin |
| TNF- α | Tumor necrosis factor α |
| NF-kB | Nuclear factor kappa light chain enhancer of activated B |
| HIF-1a | Hypoxia-inducible factor 1-alpha |
| Nrf-2 | Nuclear factor erythroid 2-related factor 2 |
| AP-1 | Activator protein 1 |
| SC | Stratum corneum |
| TEWL | Transepidermal water loss |
| EGFR | Epidermal growth factor receptor |
| TGFR | Transforming growth factor receptors |
| TLRs | Toll-like receptors |
| TNFR | TNF-α receptor |
| CSF | Colony stimulating factors |
| TGF-α | Transforming growth factor α |
| TGF- β | Transforming growth factor β |
| HMGB1 | High-mobility group box 1 |
| PDGF | Platelet-derived growth factor |
| COX2 | Cyclooxygenases 2 |
| MAP | Mitogen-activated protein |
| PGE2 | Prostaglandin 2 |
| TβRII | TGF-β type II receptor |
| ECM | Extracellular matrix |
| HDFs | Human dermal fibroblasts |
| NHDFs | Normal human dermal fibroblasts |
| HSF2 | Human skin fibroblasts |
| MED | Minimal erythemal dose |
| iNOS | Inducible nitric oxide synthase |
| FGF | Fibroblast growth factors |
| MMPs | Matrix-degrading metalloproteinases |
| MMP-1 | Collagenase-1 |
| MMP-3 | Stromelysin-1 |
| MMP-9 | Gelatinase B |
| COL1A2 | Type I collagen gene |
| SASP | Senescence-associated secretory phenotype |
References
- Zhang, S.; Duan, E. Fighting against Skin Aging. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Intrinsic and extrinsic factors in skin ageing: A review. Int. J. Cosmet. Sci. 2008, 30, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Tsatsou, F.; Trakatelli, M.; Patsatsi, A.; Kalokasidis, K.; Sotiriadis, D. Extrinsic aging. Dermato-Endocrinology 2012, 4, 285–297. [Google Scholar] [CrossRef]
- Lephart, E.D. Equol’s Anti-Aging Effects Protect against Environmental Assaults by Increasing Skin Antioxidant Defense and ECM Proteins While Decreasing Oxidative Stress and Inflammation. Cosmetics 2018, 5, 16. [Google Scholar] [CrossRef]
- O’Connor, C.; Courtney, C.; Murphy, M. Shedding light on the myths of ultraviolet radiation in the COVID-19 pandemic. Clin. Exp. Dermatol. 2021, 46, 187–188. [Google Scholar] [CrossRef] [PubMed]
- Strzałka, W.; Zgłobicki, P.; Kowalska, E.; Bażant, A.; Dziga, D.; Banaś, A.K. The Dark Side of UV-Induced DNA Lesion Repair. Genes 2020, 11, 1450. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Arango, A.; De Antioquia, U.; Flórez-Muñoz, S.; Sanclemente, G. Mechanisms of skin aging. IATREIA 2017, 30, 160–170. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Richa; Kumar, A.; Tyagi, M.B.; Sinha, R.P. Molecular Mechanisms of Ultraviolet Radiation-Induced DNA Damage and Repair. J. Nucleic Acids 2010, 2010, 1–32. [Google Scholar] [CrossRef]
- Krutmann, J.; Morita, A.; Chung, J.H. Sun Exposure: What Molecular Photodermatology Tells Us About Its Good and Bad Sides. J. Investig. Dermatol. 2012, 132, 976–984. [Google Scholar] [CrossRef]
- Fuller, B. Role of PGE-2 and Other Inflammatory Mediators in Skin Aging and Their Inhibition by Topical Natural Anti-Inflammatories. Cosmetics 2019, 6, 6. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxidants Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Yasui, H.; Sakurai, H. Chemiluminescent Detection and Imaging of Reactive Oxygen Species in Live Mouse Skin Exposed to UVA. Biochem. Biophys. Res. Commun. 2000, 269, 131–136. [Google Scholar] [CrossRef]
- Ichihashi, M.; Ueda, M.; Budiyanto, A.; Bito, T.; Oka, M.; Fukunaga, M.; Tsuru, K.; Horikawa, T. UV-induced skin damage. Toxicology 2003, 189, 21–39. [Google Scholar] [CrossRef]
- Kammeyer, A.; Luiten, R. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Davinelli, S.; Bertoglio, J.C.; Polimeni, A.; Scapagnini, G. Cytoprotective Polyphenols Against Chronological Skin Aging and Cutaneous Photodamage. Curr. Pharm. Des. 2018, 24, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G. Intracellular Signaling Mechanisms Involved in the Biological Effects of the Xanthophyll Carotenoid Astaxanthin to Prevent the Photo-aging of the Skin in a Reactive Oxygen Species Depletion-independent Manner: The Key Role of Mitogen and Stress-activated Protein Kinase 1. Photochem. Photobiol. 2018, 95, 480–489. [Google Scholar] [CrossRef]
- Hui, H.; Zhai, Y.; Ao, L.; CLeveland, J.C., Jr.; Liu, H.; Fullerton, D.A.; Meng, X. Klotho suppresses the inflammatory responses and ameliorates cardiac dysfunction in aging endotoxemic mice. Oncotarget 2017, 8, 15663–15676. [Google Scholar] [CrossRef]
- Zhang, B.; Xu, J.; Quan, Z.; Qian, M.; Liu, W.; Zheng, W.; Yin, F.; Du, J.; Zhi, Y.; Song, N. Klotho Protein Protects Human Keratinocytes from UVB-Induced Damage Possibly by Reducing Expression and Nuclear Translocation of NF-κB. Med Sci. Monit. 2018, 24, 8583–8591. [Google Scholar] [CrossRef]
- Bi, F.; Liu, W.; Wu, Z.; Ji, C.; Chang, C. Antiaging Factor Klotho Retards the Progress of Intervertebral Disc Degeneration through the Toll-Like Receptor 4-NF-κB Pathway. Int. J. Cell Biol. 2020, 2020, 8319516. [Google Scholar] [CrossRef]
- Cao, C.; Lu, S.; Kivlin, R.; Wallin, B.; Card, E.; Bagdasarian, A.; Tamakloe, T.; Wang, W.-J.; Song, X.; Chu, W.-M.; et al. SIRT1 confers protection against UVB- and H2O2-induced cell death via modulation of p53 and JNK in cultured skin keratinocytes. J. Cell. Mol. Med. 2008, 13, 3632–3643. [Google Scholar] [CrossRef]
- Lee, D.; Goldberg, A.L. Muscle Wasting in Fasting Requires Activation of NF-κB and Inhibition of AKT/Mechanistic Target of Rapamycin (mTOR) by the Protein Acetylase, GCN5. J. Biol. Chem. 2015, 290, 30269–30279. [Google Scholar] [CrossRef]
- Yousef, H.; Alhajj, M.; Sharma, S. Anatomy, Skin (Integument), Epidermis; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Kubo, A.; Nagao, K.; Amagai, M. Epidermal barrier dysfunction and cutaneous sensitization in atopic diseases. J. Clin. Investig. 2012, 122, 440–447. [Google Scholar] [CrossRef]
- Banerjee, S.; Leptin, M. Systemic Response to Ultraviolet Radiation Involves Induction of Leukocytic IL-1β and Inflammation in Zebrafish. J. Immunol. 2014, 193, 1408–1415. [Google Scholar] [CrossRef]
- Ansel, J.C.; Luger, T.A.; Green, I. The Effect of In Vitro and In Vivo UV Irradiation on the Production of ETAF Activity by Human and Murine Keratinocytes. J. Investig. Dermatol. 1983, 81, 519–523. [Google Scholar] [CrossRef]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV Radiation and the Skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef]
- Biniek, K.; Levi, K.; Dauskardt, R.H. Solar UV radiation reduces the barrier function of human skin. Proc. Natl. Acad. Sci. USA 2012, 109, 17111–17116. [Google Scholar] [CrossRef]
- Hwang, I.S.; Kim, J.E.; Choi, S.I.; Lee, H.R.; Lee, Y.J.; Jang, M.J.; Son, H.J.; Lee, H.S.; Oh, C.H.; Kim, B.H.; et al. UV radiation-induced skin aging in hairless mice is effectively prevented by oral intake of sea buckthorn (Hippophae rhamnoides L.) fruit blend for 6 weeks through MMP suppression and increase of SOD activity. Int. J. Mol. Med. 2012, 30, 392–400. [Google Scholar] [CrossRef]
- El-Domyati, M.; Attia, S.; Saleh, F.; Brown, D.; Birk, D.E.; Gasparro, F.; Ahmad, H.; Uitto, J. Intrinsic aging vs. photoaging: A comparative histopathological, immunohistochemical, and ultrastructural study of skin. Exp. Dermatol. 2002, 11, 398–405. [Google Scholar] [CrossRef]
- Pearse, A.D.; Gaskell, S.A.; Marks, R. Epidermal Changes in Human Skin Following Irradiation With Either UVB or UVA. J. Investig. Dermatol. 1987, 88, 83–87. [Google Scholar] [CrossRef]
- Takema, Y.; Yorimoto, Y.; Kawai, M.; Imokawa, G. Age-related changes in the elastic properties and thickness of human facial skin. Br. J. Dermatol. 1994, 131, 641–648. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, Z.; Voorhees, J.; Fisher, G. EGF receptor crosstalks with cytokine receptors leading to the activation of c-Jun kinase in response to UV irradiation in human keratinocytes. Cell. Signal. 2001, 13, 139–144. [Google Scholar] [CrossRef]
- Giangreco, A.; Goldie, S.J.; Failla, V.; Saintigny, G.; Watt, F.M. Human Skin Aging Is Associated with Reduced Expression of the Stem Cell Markers β1 Integrin and MCSP. J. Investig. Dermatol. 2010, 130, 604–608. [Google Scholar] [CrossRef]
- Jiang, S.J.; Chu, A.W.; Lu, Z.F.; Pan, M.H.; Che, D.F.; Zhou, X.J. Ultraviolet B-induced alterations of the skin barrier and epidermal calcium gradient. Exp. Dermatol. 2007, 16, 985–992. [Google Scholar] [CrossRef]
- Haratake, A.; Uchida, Y.; Schmuth, M.; Tanno, O.; Yasuda, R.; Epstein, J.H.; Elias, P.M.; Holleran, W.M. UVB-Induced Alterations in Permeability Barrier Function: Roles for Epidermal Hyperproliferation and Thymocyte-Mediated Response. J. Investig. Dermatol. 1997, 108, 769–775. [Google Scholar] [CrossRef]
- Kim, B.E.; Leung, D.Y. Significance of Skin Barrier Dysfunction in Atopic Dermatitis. Allergy Asthma Immunol. Res. 2018, 10, 207–215. [Google Scholar] [CrossRef]
- Abe, T.; Mayuzumi, J. The change and recovery of human skin barrier functions after ultraviolet light irradiation. Chem. Pharm. Bull. 1979, 27, 458–462. [Google Scholar] [CrossRef]
- Cestone, E.; Michelotti, A.; Zanoletti, V.; Zanardi, A.; Mantegazza, R.; Dossena, M. Acne RA-1,2, a novel UV-selective face cream for patients with acne: Efficacy and tolerability results of a randomized, placebo-controlled clinical study. J. Cosmet. Dermatol. 2017, 16, 265–270. [Google Scholar] [CrossRef]
- Suh, M.G.; Bae, G.Y.; Jo, K.; Kim, J.M.; Hong, K.-B.; Suh, H.J. Photoprotective Effect of Dietary Galacto-Oligosaccharide (GOS) in Hairless Mice via Regulation of the MAPK Signaling Pathway. Molecules 2020, 25, 1679. [Google Scholar] [CrossRef]
- Choi, H.J.; Song, B.R.; Kim, J.E.; Bae, S.J.; Choi, Y.J.; Lee, S.J.; Gong, J.E.; Lee, H.S.; Lee, C.Y.; Kim, B.-H.; et al. Therapeutic Effects of Cold-Pressed Perilla Oil Mainly Consisting of Linolenic acid, Oleic Acid and Linoleic Acid on UV-Induced Photoaging in NHDF Cells and SKH-1 Hairless Mice. Molecules 2020, 25, 989. [Google Scholar] [CrossRef]
- Piepkorn, M.; Predd, H.; Underwood, R.; Cook, P. Proliferation-differentiation relationships in the expression of heparin-binding epidermal growth factor-related factors and erbB receptors by normal and psoriatic human keratinocytes. Arch. Dermatol. Res. 2003, 295, 93–101. [Google Scholar] [CrossRef]
- Matsuura, H.; Myokai, F.; Arata, J.; Noji, S.; Taniguchi, S. Expression of type II transforming growth factor-β receptor mRNA in human skin, as revealed by in situ hybridization. J. Dermatol. Sci. 1994, 8, 25–32. [Google Scholar] [CrossRef]
- Schmid, P.; Itin, P.; Rufli, T. In situ analysis of transforming growth factors-beta (TGF-beta 1, TGF-beta 2, TGF-beta 3) and TGF-beta type II receptor expression in basal cell carcinomas. Br. J. Dermatol. 1996, 134, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Iram, N.; Mildner, M.; Prior, M.; Petzelbauer, P.; Fiala, C.; Hacker, S.; Schöppl, A.; Tschachler, E.; Elbe-Bürger, A. Age-related changes in expression and function of Toll-like receptors in human skin. Dev. 2012, 139, 4210–4219. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.; Kupper, T.S. Inflammatory Skin Diseases, T Cells, and Immune Surveillance. N. Engl. J. Med. 1999, 341, 1817–1828. [Google Scholar] [CrossRef]
- Kupper, T.S. Immune and inflammatory processes in cutaneous tissues. Mechanisms and speculations. J. Clin. Investig. 1990, 86, 1783–1789. [Google Scholar] [CrossRef]
- Lee, R.T.; Briggs, W.H.; Cheng, G.C.; Rossiter, H.B.; Libby, P.; Kupper, T. Mechanical deformation promotes secretion of IL-1 al-pha and IL-1 receptor antagonist. J. Immunol. 1997, 159, 5084–5088. [Google Scholar]
- Ansel, J.; Perry, P.; Brown, J.; Damm, D.; Phan, T.; Hart, C.; Luger, T.; Hefeneider, S. Cytokine Modulation of Keratinocyte Cytokines. J. Investig. Dermatol. 1990, 94, s101–s107. [Google Scholar] [CrossRef]
- Johnson, K.E.; Wulff, B.C.; Oberyszyn, T.M.; Wilgus, T.A. Ultraviolet light exposure stimulates HMGB1 release by keratinocytes. Arch. Dermatol. Res. 2013, 305, 805–815. [Google Scholar] [CrossRef]
- Meephansan, J.; Komine, M.; Tsuda, H.; Tominaga, S.-I.; Ohtsuki, M. Ultraviolet B irradiation induces the expression of IL-33 mRNA and protein in normal human epidermal keratinocytes. J. Dermatol. Sci. 2012, 65, 72–74. [Google Scholar] [CrossRef]
- Choi, S.; Jung, T.; Cho, B.; Choi, S.; Sim, W.; Han, X.; Lee, S.J.; Kim, Y.; Lee, O. Anti-photoaging effect of fermented agricultural by-products on ultraviolet B-irradiated hairless mouse skin. Int. J. Mol. Med. 2019, 44, 559–568. [Google Scholar] [CrossRef]
- Choi, S.-H.; Choi, S.-I.; Jung, T.-D.; Cho, B.-Y.; Lee, J.-H.; Kim, S.-H.; Yoon, S.-A.; Ham, Y.-M.; Yoon, W.-J.; Cho, J.-H.; et al. Anti-Photoaging Effect of Jeju Putgyul (Unripe Citrus) Extracts on Human Dermal Fibroblasts and Ultraviolet B-induced Hairless Mouse Skin. Int. J. Mol. Sci. 2017, 18, 2052. [Google Scholar] [CrossRef]
- Wang, X.-F.; Huang, Y.-F.; Wang, L.; Xu, L.-Q.; Yu, X.-T.; Liu, Y.-H.; Li, C.-L.; Zhan, J.Y.-X.; Su, Z.-R.; Chen, J.-N.; et al. Photo-protective activity of pogostone against UV-induced skin premature aging in mice. Exp. Gerontol. 2016, 77, 76–86. [Google Scholar] [CrossRef]
- Kwak, C.S.; Yang, J.; Shin, C.-Y.; Chung, J.H. Rosa multiflora Thunb Flower Extract Attenuates Ultraviolet-Induced Photoaging in Skin Cells and Hairless Mice. J. Med. Food 2020, 23, 988–997. [Google Scholar] [CrossRef]
- Martinez, R.M.; Fattori, V.; Saito, P.; Pinto, I.C.; Rodrigues, C.C.A.; Melo, C.P.B.; Bussmann, A.J.C.; Staurengo-Ferrari, L.; Bezerra, J.R.; Vignoli, J.A.; et al. The Lipoxin Receptor/FPR2 Agonist BML-111 Protects Mouse Skin Against Ultraviolet B Radiation. Molecules 2020, 25, 2953. [Google Scholar] [CrossRef]
- Rui, Y.; Zhaohui, Z.; Wenshan, S.; Bafang, L.; Hu, H. Protective effect of MAAs extracted from Porphyra tenera against UV irradiation-induced photoaging in mouse skin. J. Photochem. Photobiol. B Biol. 2019, 192, 26–33. [Google Scholar] [CrossRef]
- Moon, K.-C.; Yang, J.-P.; Lee, J.-S.; Jeong, S.-H.; Dhong, E.-S.; Han, S.-K. Effects of Ultraviolet Irradiation on Cellular Senescence in Keratinocytes Versus Fibroblasts. J. Craniofacial Surg. 2019, 30, 270–275. [Google Scholar] [CrossRef]
- Bora, N.S.; Mazumder, B.; Mandal, S.; Patowary, P.; Goyary, D.; Chattopadhyay, P.; Dwivedi, S.K. Amelioration of UV radiation-induced photoaging by a combinational sunscreen formulation via aversion of oxidative collagen degradation and promotion of TGF-β-Smad-mediated collagen production. Eur. J. Pharm. Sci. 2019, 127, 261–275. [Google Scholar] [CrossRef]
- Qin, D.; Lee, W.-H.; Gao, Z.; Zhang, W.; Peng, M.; Sun, T.; Gao, Y. Protective effects of antioxidin-RL from Odorrana livida against ultraviolet B-irradiated skin photoaging. Peptides 2018, 101, 124–134. [Google Scholar] [CrossRef]
- Kong, S.-Z.; Li, D.-D.; Luo, H.; Li, W.-J.; Huang, Y.-M.; Li, J.-C.; Hu, Z.; Huang, N.; Guo, M.-H.; Chen, Y.; et al. Anti-photoaging effects of chitosan oligosaccharide in ultraviolet-irradiated hairless mouse skin. Exp. Gerontol. 2018, 103, 27–34. [Google Scholar] [CrossRef]
- Subedi, L.; Lee, T.H.; Wahedi, H.M.; Baek, S.-H.; Kim, S.Y. Resveratrol-Enriched Rice Attenuates UVB-ROS-Induced Skin Aging via Downregulation of Inflammatory Cascades. Oxidative Med. Cell. Longev. 2017, 2017, 1–15. [Google Scholar] [CrossRef]
- Kang, S.M.; Han, S.; Oh, J.-H.; Lee, Y.M.; Park, C.-H.; Shin, C.-Y.; Lee, D.H.; Chung, J.H. A synthetic peptide blocking TRPV1 activation inhibits UV-induced skin responses. J. Dermatol. Sci. 2017, 88, 126–133. [Google Scholar] [CrossRef][Green Version]
- Schneider, L.A.; Raizner, K.; Wlaschek, M.; Brenneisen, P.; Gethöffer, K.; Scharffetter-Kochanek, K. UVA-1 exposure in vivo leads to an IL-6 surge within the skin. Exp. Dermatol. 2017, 26, 830–832. [Google Scholar] [CrossRef]
- Misawa, E.; Tanaka, M.; Saito, M.; Nabeshima, K.; Yao, R.; Yamauchi, K.; Abe, F.; Yamamoto, Y.; Furukawa, F. Protective effects ofAloesterols against UVB-induced photoaging in hairless mice. Photodermatol. Photoimmunol. Photomed. 2017, 33, 101–111. [Google Scholar] [CrossRef]
- Kim, E.J.; Kim, Y.K.; Kim, M.-K.; Kim, S.; Kim, J.Y.; Lee, D.H.; Chung, J.H. UV-induced inhibition of adipokine production in subcutaneous fat aggravates dermal matrix degradation in human skin. Sci. Rep. 2016, 6, 25616. [Google Scholar] [CrossRef]
- Zhan, J.Y.-X.; Wang, X.-F.; Liu, Y.-H.; Zhang, Z.-B.; Wang, L.; Chen, J.-N.; Huang, S.; Zeng, H.-F.; Lai, X.-P. Andrographolide Sodium Bisulfate Prevents UV-Induced Skin Photoaging through Inhibiting Oxidative Stress and Inflammation. Mediat. Inflamm. 2016, 2016, 1–12. [Google Scholar] [CrossRef]
- Afnan, Q.; Kaiser, P.J.; Rafiq, R.A.; Nazir, L.A.; Bhushan, S.; Bhardwaj, S.C.; Sandhir, R.; Tasduq, S.A. Glycyrrhizic acid prevents ultraviolet-B-induced photodamage: A role for mitogen-activated protein kinases, nuclear factor kappa B and mitochondrial apoptotic pathway. Exp. Dermatol. 2016, 25, 440–446. [Google Scholar] [CrossRef]
- Yoo, J.H.; Kim, J.K.; Yang, H.J.; Park, K.M. Effects of Egg Shell Membrane Hydrolysates on UVB-radiation-induced Wrinkle Formation in SKH-1 Hairless Mice. Food Sci. Anim. Resour. 2015, 35, 58–70. [Google Scholar] [CrossRef]
- Kim, W.; Kim, E.; Yang, H.J.; Kwon, T.; Han, S.; Lee, S.; Youn, H.; Jung, Y.H.; Kang, C.; Youn, B. Inhibition of hedgehog signalling attenuates UVB-induced skin photoageing. Exp. Dermatol. 2015, 24, 611–617. [Google Scholar] [CrossRef]
- Zhang, X.; Xie, Y.-L.; Yu, X.-T.; Su, Z.-Q.; Yuan, J.; Li, Y.-C.; Su, Z.-R.; Zhan, J.Y.-X.; Lai, X.-P. Protective Effect of Super-Critical Carbon Dioxide Fluid Extract from Flowers and Buds of Chrysanthemum indicum Linnén Against Ultraviolet-Induced Photo-Aging in Mice. Rejuvenation Res. 2015, 18, 437–448. [Google Scholar] [CrossRef]
- Kong, S.-Z.; Chen, H.-M.; Yu, X.-T.; Zhang, X.; Feng, X.-X.; Kang, X.-H.; Li, W.-J.; Huang, N.; Luo, H.; Su, Z.-R. The protective effect of 18β-Glycyrrhetinic acid against UV irradiation induced photoaging in mice. Exp. Gerontol. 2015, 61, 147–155. [Google Scholar] [CrossRef]
- Kim, M.-J.; Woo, S.W.; Kim, M.-S.; Park, J.-E.; Hwang, J.-K. Anti-photoaging effect of aaptamine in UVB-irradiated human dermal fibroblasts and epidermal keratinocytes. J. Asian Nat. Prod. Res. 2014, 16, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.E.; Kim, M.S.; Jeon, W.-K.; Seo, Y.K.; Kim, B.-C.; Hahn, J.H.; Park, C.S. A nuclear factor kappa B-derived inhibitor tripeptide inhibits UVB-induced photoaging process. J. Dermatol. Sci. 2014, 76, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.-X.; Yu, X.-T.; Li, W.-J.; Kong, S.-Z.; Liu, Y.-H.; Zhang, X.; Xian, Y.-F.; Zhang, X.-J.; Su, Z.-R.; Lin, Z.-X. Effects of topical application of patchouli alcohol on the UV-induced skin photoaging in mice. Eur. J. Pharm. Sci. 2014, 63, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-C.; Chiang, A.-N.; Liu, H.-N.; Chang, Y.-T. EGb-761 prevents ultraviolet B-induced photoaging via inactivation of mitogen-activated protein kinases and proinflammatory cytokine expression. J. Dermatol. Sci. 2014, 75, 55–62. [Google Scholar] [CrossRef]
- Chiu, H.-W.; Chen, C.-H.; Chen, Y.-J.; Hsu, Y.-H. Far-infrared suppresses skin photoaging in ultraviolet B-exposed fibroblasts and hairless mice. PLoS ONE 2017, 12, e0174042. [Google Scholar] [CrossRef]
- El-Abaseri, T.B.; Hammiller, B.; Repertinger, S.K.; Hansen, L.A. The Epidermal Growth Factor Receptor Increases Cytokine Production and Cutaneous Inflammation in Response to Ultraviolet Irradiation. ISRN Dermatol. 2013, 2013, 1–11. [Google Scholar] [CrossRef]
- Kim, A.L.; Labasi, J.M.; Zhu, Y.; Tang, X.; McClure, K.; Gabel, C.A.; Athar, M.; Bickers, D.R. Role of p38 MAPK in UVB-Induced Inflammatory Responses in the Skin of SKH-1 Hairless Mice. J. Investig. Dermatol. 2005, 124, 1318–1325. [Google Scholar] [CrossRef]
- Murata, K.; Oyama, M.; Ogata, M.; Fujita, N.; Takahashi, R. Oral administration of Jumihaidokuto inhibits UVB-induced skin damage and prostaglandin E2 production in HR-1 hairless mice. J. Nat. Med. 2021, 75, 142–155. [Google Scholar] [CrossRef]
- Gerber, P.A.; Buhren, B.A.; Schrumpf, H.; Hevezi, P.; Bölke, E.; Sohn, D.; Jänicke, R.U.; Belum, V.R.; Robert, C.; Lacouture, M.E.; et al. Mechanisms of skin aging induced by EGFR inhibitors. Support. Care Cancer 2016, 24, 4241–4248. [Google Scholar] [CrossRef]
- O’Dea, E.L.; Kearns, J.D.; Hoffmann, A. UV as an Amplifier Rather Than Inducer of NF-κB Activity. Mol. Cell 2008, 30, 632–641. [Google Scholar] [CrossRef]
- Vicentini, F.T.; He, T.; Shao, Y.; Fonseca, M.J.; Verri, W.A.; Fisher, G.J.; Xu, Y. Quercetin inhibits UV irradiation-induced inflammatory cytokine production in primary human keratinocytes by suppressing NF-κB pathway. J. Dermatol. Sci. 2011, 61, 162–168. [Google Scholar] [CrossRef]
- Xia, J.; Song, X.; Bi, Z.; Chu, W.; Wan, Y. UV-induced NF-kappaB activation and expression of IL-6 is attenuated by (-)-epigallocatechin-3-gallate in cultured human keratinocytes in vitro. Int. J. Mol. Med. 2005, 16, 943–950. [Google Scholar]
- Bernard, J.J.; Cowing-Zitron, C.; Nakatsuji, T.; Muehleisen, B.; Muto, J.; Borkowski, A.W.; Martinez, L.; Greidinger, E.L.; Yu, B.D.; Gallo, R.L. Ultraviolet radiation damages self noncoding RNA and is detected by TLR3. Nat. Med. 2012, 18, 1286–1290. [Google Scholar] [CrossRef]
- Jo, J.; Im, S.H.; Babcock, D.T.; Iyer, S.C.; Gunawan, F.; Cox, D.N.; Galko, M.J. Drosophila caspase activity is required independently of apoptosis to produce active TNF/Eiger during nociceptive sensitization. Cell Death Dis. 2017, 8, e2786. [Google Scholar] [CrossRef]
- Deshmukh, J.; Pofahl, R.; Haase, I. Epidermal Rac1 regulates the DNA damage response and protects from UV-light-induced keratinocyte apoptosis and skin carcinogenesis. Cell Death Dis. 2017, 8, e2664. [Google Scholar] [CrossRef]
- Schwarz, A.; Bhardwaj, R.; Aragane, Y.; Mahnke, K.; Riemann, H.; Metze, D.; Luger, T.A.; Schwarz, T. Ultraviolet-B-Induced Apoptosis of Keratinocytes: Evidence for Partial Involvement of Tumor Necrosis Factor-α in the Formation of Sunburn Cells. J. Investig. Dermatol. 1995, 104, 922–927. [Google Scholar] [CrossRef]
- Liarte, S.; Bernabé-García, Á.; Nicolás, F.J. Role of TGF-β in Skin Chronic Wounds: A Keratinocyte Perspective. Cells 2020, 9, 306. [Google Scholar] [CrossRef]
- Quan, T.; He, T.; Kang, S.; Voorhees, J.J.; Fisher, G.J. Ultraviolet Irradiation Alters Transforming Growth Factor β/Smad Pathway in Human Skin In Vivo. J. Investig. Dermatol. 2002, 119, 499–506. [Google Scholar] [CrossRef]
- Xu, D.; Yuan, R.; Gu, H.; Liu, T.; Tu, Y.; Yang, Z.; He, L. The effect of ultraviolet radiation on the transforming growth factor beta 1/Smads pathway and p53 in actinic keratosis and normal skin. Arch. Dermatol. Res. 2013, 305, 777–786. [Google Scholar] [CrossRef]
- Brown, T.M.; Krishnamurthy, K. Histology, Dermis; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Prost-Squarcioni, C.; Fraitag, S.; Heller, M.; Boehm, N. Histologie fonctionnelle du derme. Annales de Dermatologie et de Vénéréologie 2008, 135, 5–20. [Google Scholar] [CrossRef]
- Won, H.-R.; Lee, P.; Oh, S.-R.; Kim, Y.-M. Epigallocatechin-3-Gallate Suppresses the Expression of TNF-α-Induced MMP-1 via MAPK/ERK Signaling Pathways in Human Dermal Fibroblasts. Biol. Pharm. Bull. 2021, 44, 18–24. [Google Scholar] [CrossRef]
- Mueller, S.N.; Zaid, A.; Carbone, F.R. Tissue-Resident T Cells: Dynamic Players in Skin Immunity. Front. Immunol. 2014, 5, 332. [Google Scholar] [CrossRef]
- Nigam, Y.; Knight, J. Exploring the anatomy and physiology of ageing. Part 11—The skin. Nurs. Times 2009, 104, 24–25. [Google Scholar]
- Manicone, A.M.; McGuire, J.K. Matrix metalloproteinases as modulators of inflammation. Semin. Cell Dev. Biol. 2008, 19, 34–41. [Google Scholar] [CrossRef]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Shin, J.-W.; Kwon, S.-H.; Choi, J.-Y.; Na, J.-I.; Huh, C.-H.; Choi, H.-R.; Park, K.-C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef]
- Quan, T.; Little, E.; Quan, H.; Qin, Z.; Voorhees, J.J.; Fisher, G.J. Elevated Matrix Metalloproteinases and Collagen Fragmentation in Photodamaged Human Skin: Impact of Altered Extracellular Matrix Microenvironment on Dermal Fibroblast Function. J. Investig. Dermatol. 2013, 133, 1362–1366. [Google Scholar] [CrossRef]
- Kruger, T.E.; Miller, A.H.; Wang, J. Collagen Scaffolds in Bone Sialoprotein-Mediated Bone Regeneration. Sci. World J. 2013, 2013, 1–6. [Google Scholar] [CrossRef]
- Fisher, G.J.; Choi, H.-C.; Bata-Csorgo, Z.; Shao, Y.; Datta, S.; Wang, Z.-Q.; Kang, S.; Voorhees, J.J. Ultraviolet Irradiation Increases Matrix Metalloproteinase-8 Protein in Human Skin In Vivo. J. Investig. Dermatol. 2001, 117, 219–226. [Google Scholar] [CrossRef]
- Quan, T.; He, T.; Kang, S.; Voorhees, J.J.; Fisher, G.J. Solar Ultraviolet Irradiation Reduces Collagen in Photoaged Human Skin by Blocking Transforming Growth Factor-β Type II Receptor/Smad Signaling. Am. J. Pathol. 2004, 165, 741–751. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Yuan, W.; Mori, Y.; Varga, J. Smad-dependent stimulation of type I collagen gene expression in human skin fibroblasts by TGF-β involves functional cooperation with p300/CBP transcriptional coactivators. Oncogene 2000, 19, 3546–3555. [Google Scholar] [CrossRef] [PubMed]
- Park, B.; Hwang, E.; Seo, S.A.; Zhang, M.; Park, S.-Y.; Yi, T.-H. Dietary Rosa damascena protects against UVB-induced skin aging by improving collagen synthesis via MMPs reduction through alterations of c-Jun and c-Fos and TGF-β1 stimulation mediated smad2/3 and smad7. J. Funct. Foods 2017, 36, 480–489. [Google Scholar] [CrossRef]
- Fridlyanskaya, I.; Alekseenko, L.; Nikolsky, N. Senescence as a general cellular response to stress: A mini-review. Exp. Gerontol. 2015, 72, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Ogata, Y.; Yamada, T.; Hasegawa, S.; Sanada, A.; Iwata, Y.; Arima, M.; Nakata, S.; Sugiura, K.; Akamatsu, H. SASP-induced macrophage dysfunction may contribute to accelerated senescent fibroblast accumulation in the dermis. Exp. Dermatol. 2021, 30, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Wlaschek, M.; Maity, P.; Makrantonaki, E.; Scharffetter-Kochanek, K. Connective Tissue and Fibroblast Senescence in Skin Aging. J. Investig. Dermatol. 2021, 141, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Lupa, D.M.W.; Kalfalah, F.; Safferling, K.; Boukamp, P.; Poschmann, G.; Volpi, E.; Götz-Rösch, C.; Bernerd, F.; Haag, L.; Huebenthal, U.; et al. Characterization of Skin Aging–Associated Secreted Proteins (SAASP) Produced by Dermal Fibroblasts Isolated from Intrinsically Aged Human Skin. J. Investig. Dermatol. 2015, 135, 1954–1968. [Google Scholar] [CrossRef]
- Victorelli, S.; Lagnado, A.; Halim, J.; Moore, W.; Talbot, D.; Barrett, K.; Chapman, J.; Birch, J.; Ogrodnik, M.; Meves, A.; et al. Senescent human melanocytes drive skin ageing via paracrine telomere dysfunction. EMBO J. 2019, 38, e101982. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Olivieri, F.; Prattichizzo, F.; Grillari, J.; Balistreri, C.R. Cellular Senescence and Inflammaging in Age-Related Diseases. Mediat. Inflamm. 2018, 2018, 1–6. [Google Scholar] [CrossRef]
- Basisty, N.; Kale, A.; Jeon, O.H.; Kuehnemann, C.; Payne, T.; Rao, C.; Holtz, A.; Shah, S.; Sharma, V.; Ferrucci, L.; et al. A proteomic atlas of senescence-associated secretomes for aging biomarker development. PLoS Biol. 2020, 18, e3000599. [Google Scholar] [CrossRef]
- Childs, B.G.; Baker, D.J.; Wijshake, T.; Conover, C.A.; Campisi, J.; Van Deursen, J.M. Senescent intimal foam cells are deleterious at all stages of atherosclerosis. Science 2016, 354, 472–477. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Type | Study Subject | UV Dose | Inflammatory Cytokine | Aging Phenotype | Reference |
|---|---|---|---|---|---|
| In-vivo | DBA/2 mice | 180 mJ/cm2 | TNF-α | Increase epidermal thickness, neutrophil infiltration | [47] |
| In-vivo | HR-1 hairless mice | 100 mJ/cm2 | TNF-α, COX-2, iNOS, IL-6, IL-1β | Skin wrinkle formation, increase epidermal thickness, collagen degradation, mast cell infiltration | [51] |
| In-vivo | HR-1 hairless mice | 100 mJ/cm2 | TNF-α, IL-6, IL-1β | Skin wrinkle formation, increase epidermal thickness, collagen degradation, trans epidermal water loss (TEWL) of dorsal skin | [52] |
| In-vivo | Chinese Kun Ming mice | 100–400 mJ/cm2 | IL-1β, IL-6, TNF-α, COX-2, PGE2, MMP-1, MMP-3 | Coarse wrinkles, erythema, edema, thickening, leathery appearance, epidermal hyperplasia, reduced collagen fibers | [53] |
| In-vivo | SKH-1 hairless mice | 100 mJ/cm2 | TNF-α, MMP-13, IL-1 β, IL-6 | Increase epidermal thickness, collagen degradation | [54] |
| In-vivo | hairless mice (HRS/J) | 0.384 mW/ cm2 | TNF-α, IL-6, IL-1β | Increase epidermal thickness, collagen degradation, mast cell infiltration | [55] |
| In-vivo | hairless mice | 312 nm and 790 µW/cm2 intensity | IL-6, IL-12, TNF-α | Skin wrinkle formation, increase epidermal thickness, collagen degradation, collagen and elastic fibers’ (elastin) degradation | [39] |
| In-vivo | ICR mice | UVA: 20.81 J/cm2, UVB: 0.47 J/cm2 | MMP-1, MMP-3, TNF-α | Thicker scarf-skin, and a disruption of the skin tissue in dermis | [56] |
| In-vitro | HaCaT and HDFs | 200 mJ/cm2 | TNF-α, IL-1β | Increase senescence-associated β -galactosidase activity, increase ROS production | [57] |
| In-vivo | Wister rat | 280–400 nm | IL-6, IL-1β, TNF-α | Increase epidermal thickness, disrupted stratum corneum, abnormal hair follicles and loss of histological architecture, uneven sebaceous glands dermis, reduced skin collagen and elastic fibers | [58] |
| In-vivo and In-vitro | HaCaT | 20 mJ/cm2 | COX-2, MMP-1, FGF, TNF-α, IL-6, and decreased TGF-β1 | Increase ROS production, DNA oxidative damage, reduce procollagen type I | [59] |
| Kunming mice | 150–300 mJ/cm2 | Not investigated | Increase epidermal and dermis thickness, infiltrationof inflammatory cells, decrease collagen fibers | ||
| In-vivo | hairless BALB/c | 55 mJ/cm2 | TNF-α, IL-1β, IL-6 | Increase epidermal and dermis thickness, skin erythema, dry, thickening, sagging, coarse wrinkles, and reduced skin collagen type I | [60] |
| In-vitro and ex-vivo | NHDFs | 144 mJ/cm2 | TNF-α, IL-6, iNOS, COX-2 | Increase ROS production | [61] |
| Reconstructed human skin (Keraskin™ FT) | 100 mJ/cm2 | Not investigated | Wrinkle formation, disruption and decomposition of collagen fibers in skin tissues exposed to UVB | ||
| In-vivo and In-vitro | HaCaT | 75 mJ/cm2 | TNF-α, IL-6, MMP-1 | Not investigated | [62] |
| hairless mice | 100 mJ/cm2 | MMP-9, MMP-13 | Increase epidermal and dermis thickness | ||
| In-vivo | Human | 25–50 J/ cm2 | IL-6 | Not investigated | [63] |
| In-vivo | hairless mice | 45–210 mJ/cm2 | MMP-2, 9, 12 and 13, TNF-α, IL-1β | Increase epidermal thickness, decrease Type I collagen, deep wrinkle formation | [64] |
| In-vivo | Human | 70 and 90 mJ/cm2 | TNF-α, MMP-1 | Decrease Type I procollagen | [65] |
| In-vivo | Kunming mice | 1 to 4 MED, 1 MED = 70 mJ/cm2 | IL-1β, IL-6, IL-10, TNF- α, MMP-1, MMP-3 | Reduce skin elasticity, coarse wrinkle formation, erythema, increase epidermal thickness, decrease collagen content | [66] |
| In-vivo and In-vitro | HaCaT cell | 10 mJ/cm2 | IL-1α, IL-1β, IL-6, TNF-α, MMP-2/9 | Increase ROS production | [67] |
| BALB/c mice | 180 mJ/cm2 | Not Investigated | Increase epidermal thickness, infiltration of leucocyte | ||
| In-vivo | SKH-1 hairless mice | 30 to 120 mJ/cm2 | MMP-1, 2, 9, COX2, IL-1β, IL-6 | Increase epidermal and dermal thickness, thick and deep wrinkle formation, decrease collagen content | [68] |
| In-vivo and In-vitro | HaCaT cell | 50 J/m2 | COX-2 IL-1β, IL-6, TNF-α, MMP1, MMP2, MMP9 | Increase cellular senescence | [69] |
| BALB/c athymic nude mice | 60 to 120 mJ/cm2 | COX-2, IL-1β | Increase epidermal thickness, wrinkle formation | ||
| In-vivo | Kunming mice | 1 to 4 MED, 1 MED = 70 mJ/cm2 | IL-1β, IL-6, IL-10, TNF-α | Deep wrinkles, erythema, edema, and skin burn, increase epidermal thickness, degrade dermal collagen fibers, suppress antioxidant enzymes | [70] |
| In-vivo | Kunming mice | 75 to 300 mJ/cm2 | MMP-1, MMP-3, IL-6, TNF-α, IL-1 | Edema, erythema, thickening and coarse wrinkles, epidermal hyperplasia, disorganized collagen fibers | [71] |
| In-vitro | HaCaT cell | 15 mJ/cm2 | COX-2, TNF-α, IL-1β | Increase ROS production, suppress antioxidant enzymes | [72] |
| In-vivo and In-vitro | HaCaT cell | 12.5 mJ/cm2 | TNF-α, IL-1 α, MMP-1, COX-2 | Not investigated | [73] |
| SKH-1 hairless mice | 36–122 mJ/cm2 | MMP-1 | Increase epidermal and dermal thickness, decrease Type I collagen | ||
| In-vivo | KM mice | 1–4 MED, 1 MED = 100 mJ/cm2 | IL-10, IL-6, TNF-α, MMP-1, MMP-3 | Severe wrinkles, increase epidermal thickness, decease antioxidant enzymes | [74] |
| In-vivo and In-vitro | NHDFs | 90 mJ/cm2 | MMP-1, IL-1 α, IL-1β, IL-6, TNF-α | Increase ROS production | [75] |
| BALB/c mice | 90 mJ/cm2 | IL-8 | Wrinkle formation, epidermal hyperplasia | ||
| In-vivo | SKH-1 hairless mice | 100–200 mJ/cm2 | MMP-1, MMP-9, decreased TGF-β1 | Increase epidermal thickness, reduce procollagen type I | [76] |
| In-vivo | SKH-1 hairless mice | 280–320 nm | Reduced klotho | Hyper-thickened epidermis | [18] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ansary, T.M.; Hossain, M.R.; Kamiya, K.; Komine, M.; Ohtsuki, M. Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging. Int. J. Mol. Sci. 2021, 22, 3974. https://doi.org/10.3390/ijms22083974
Ansary TM, Hossain MR, Kamiya K, Komine M, Ohtsuki M. Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging. International Journal of Molecular Sciences. 2021; 22(8):3974. https://doi.org/10.3390/ijms22083974
Chicago/Turabian StyleAnsary, Tuba M., Md. Razib Hossain, Koji Kamiya, Mayumi Komine, and Mamitaro Ohtsuki. 2021. "Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging" International Journal of Molecular Sciences 22, no. 8: 3974. https://doi.org/10.3390/ijms22083974
APA StyleAnsary, T. M., Hossain, M. R., Kamiya, K., Komine, M., & Ohtsuki, M. (2021). Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging. International Journal of Molecular Sciences, 22(8), 3974. https://doi.org/10.3390/ijms22083974