
Transcription Factors Interact with ABA through Gene Expression and Signaling Pathways to Mitigate Drought and Salinity Stress

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
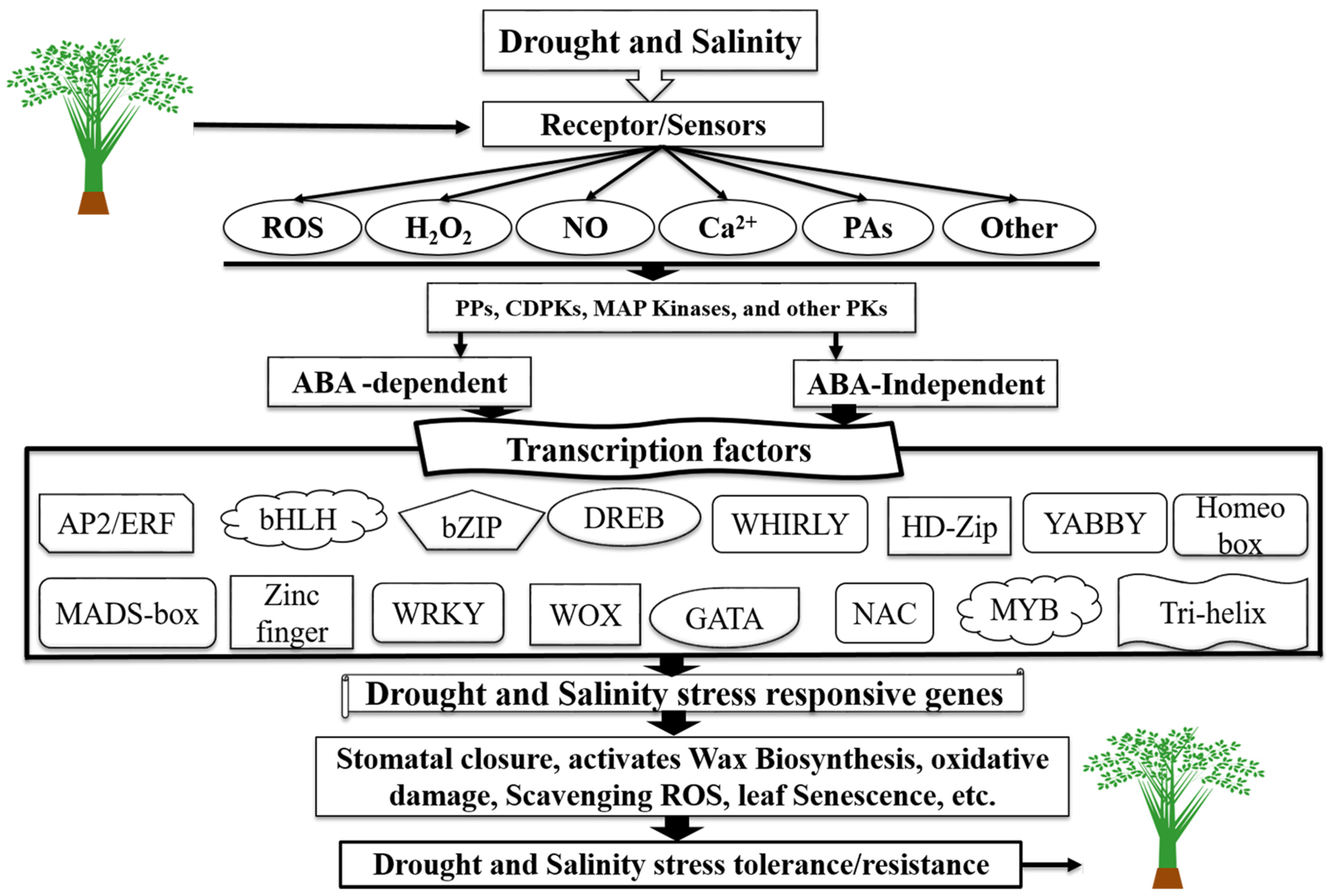
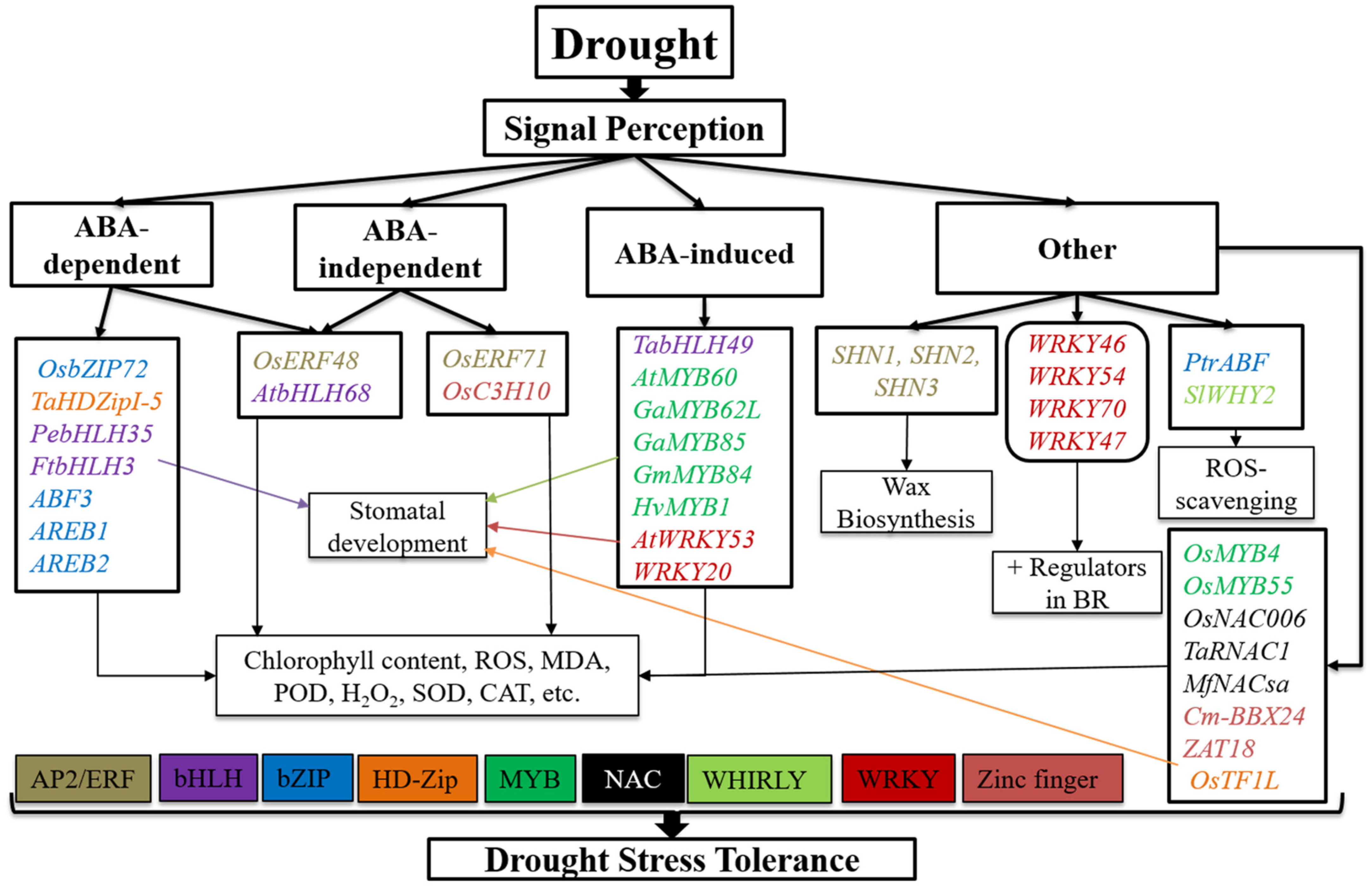
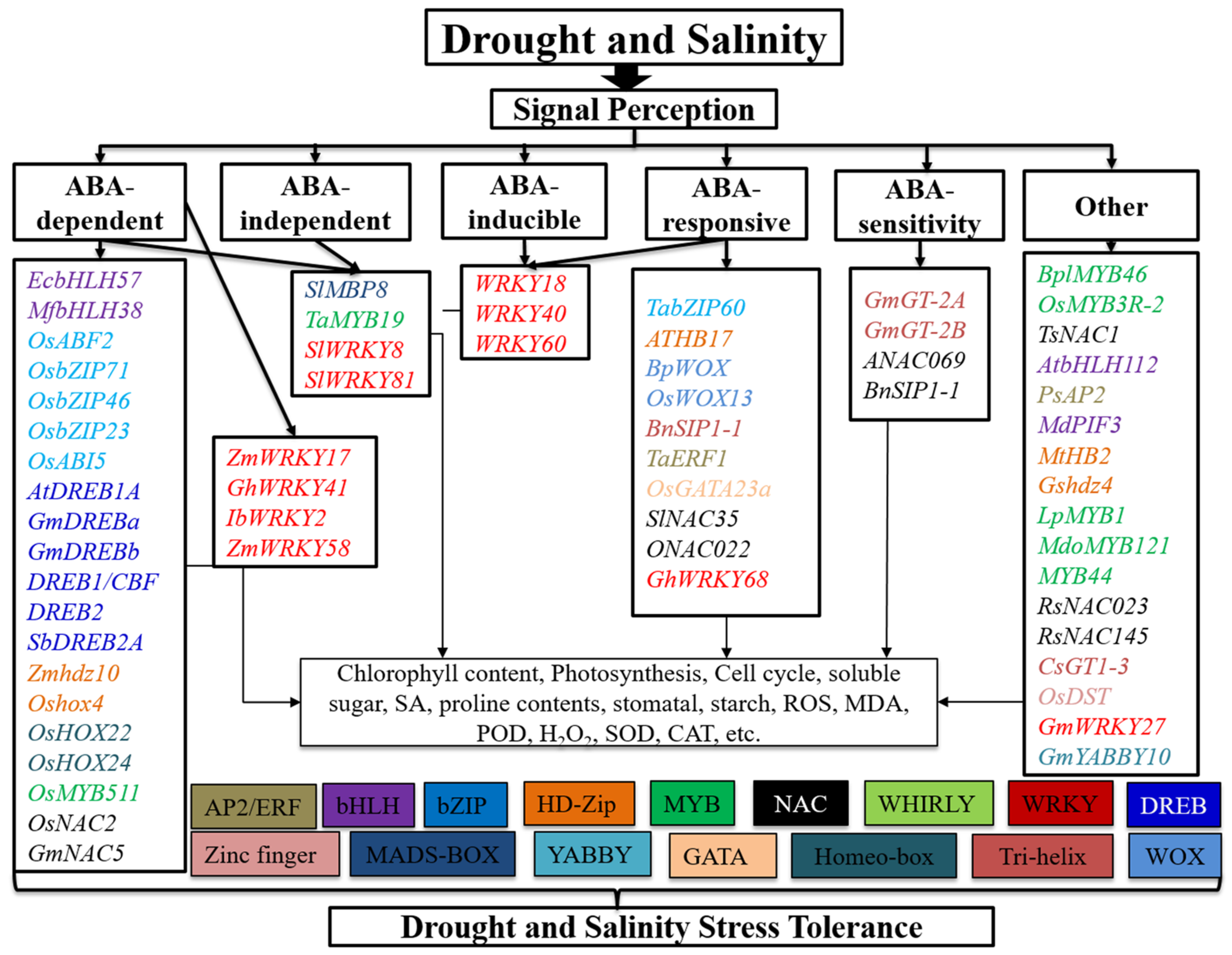
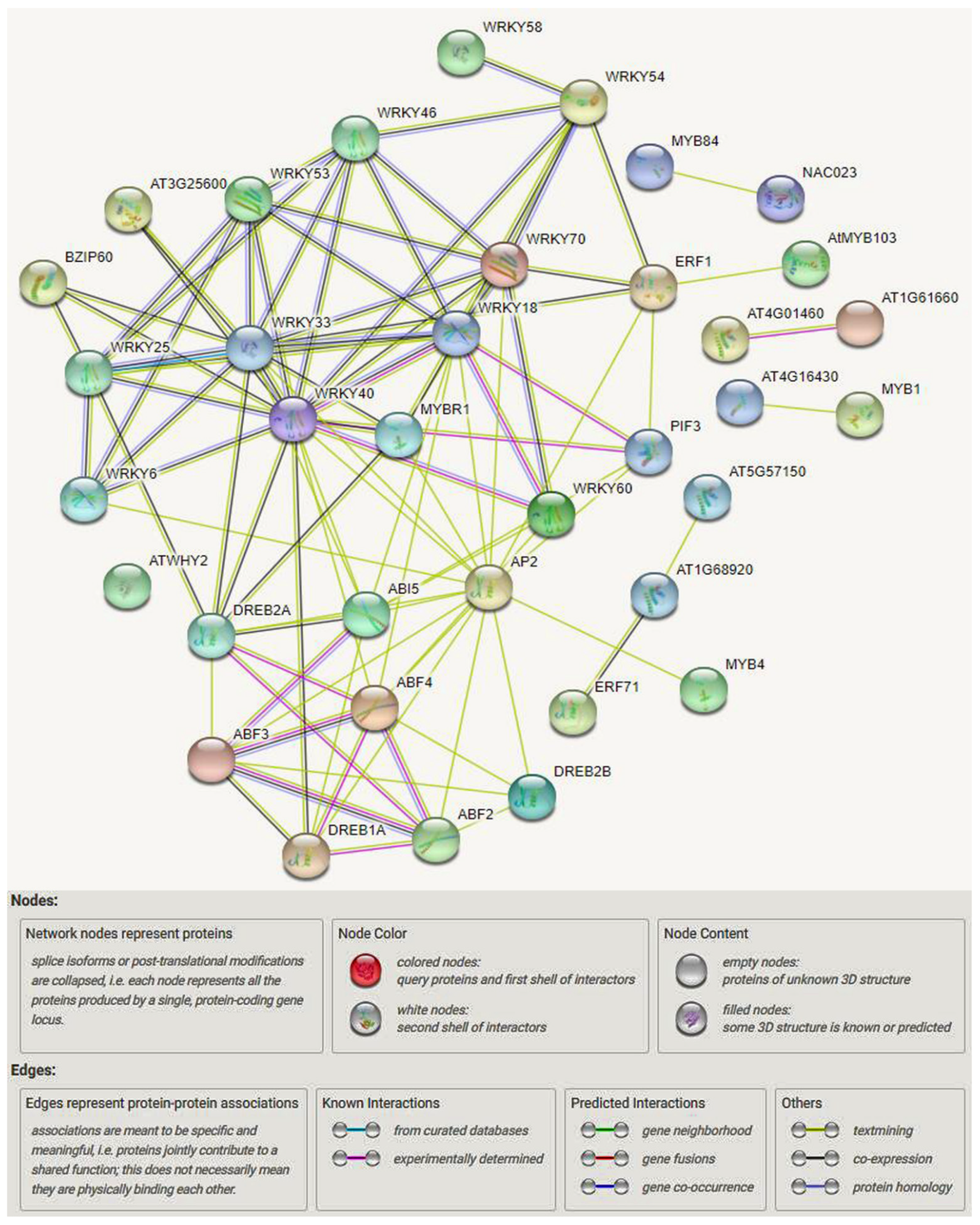
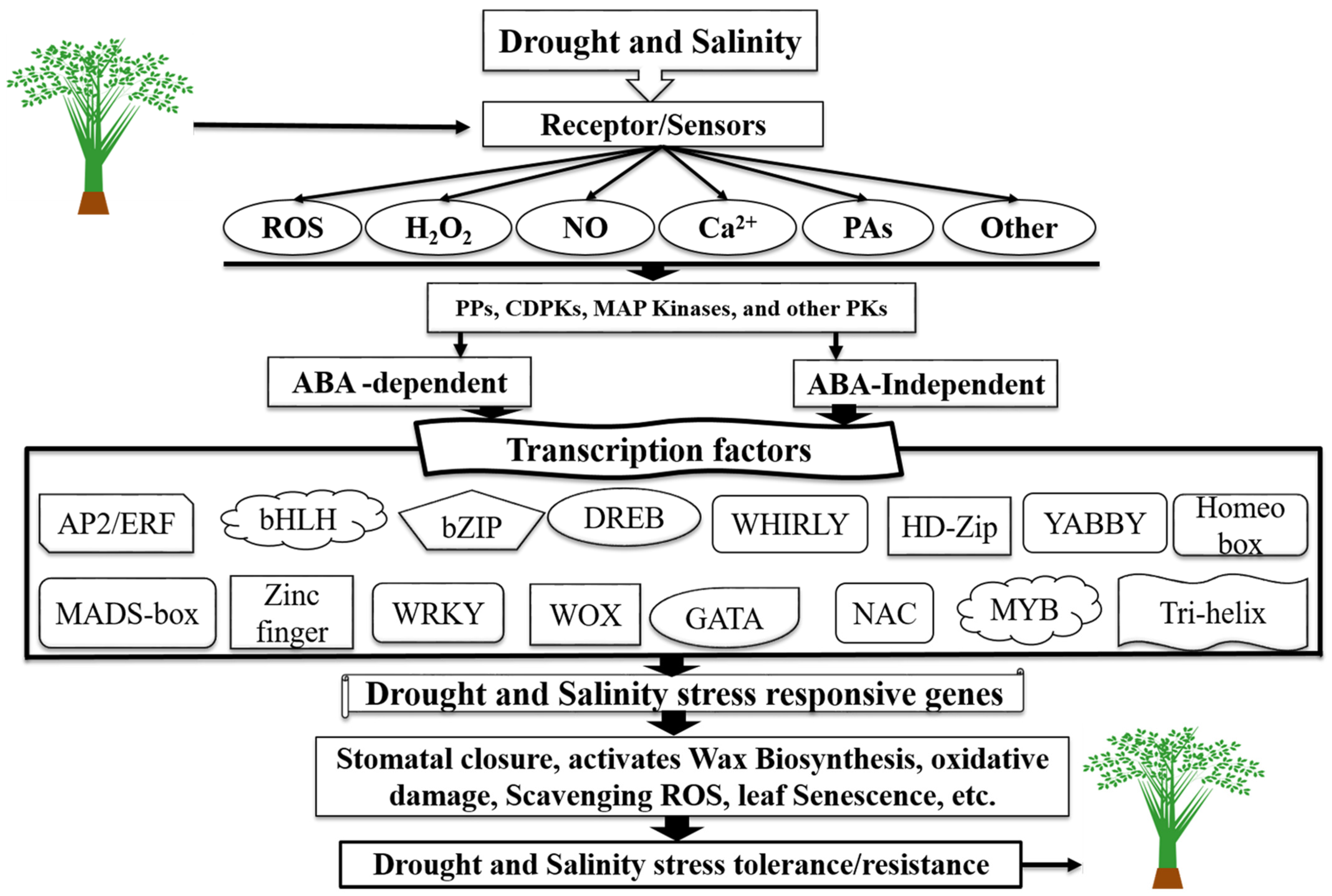
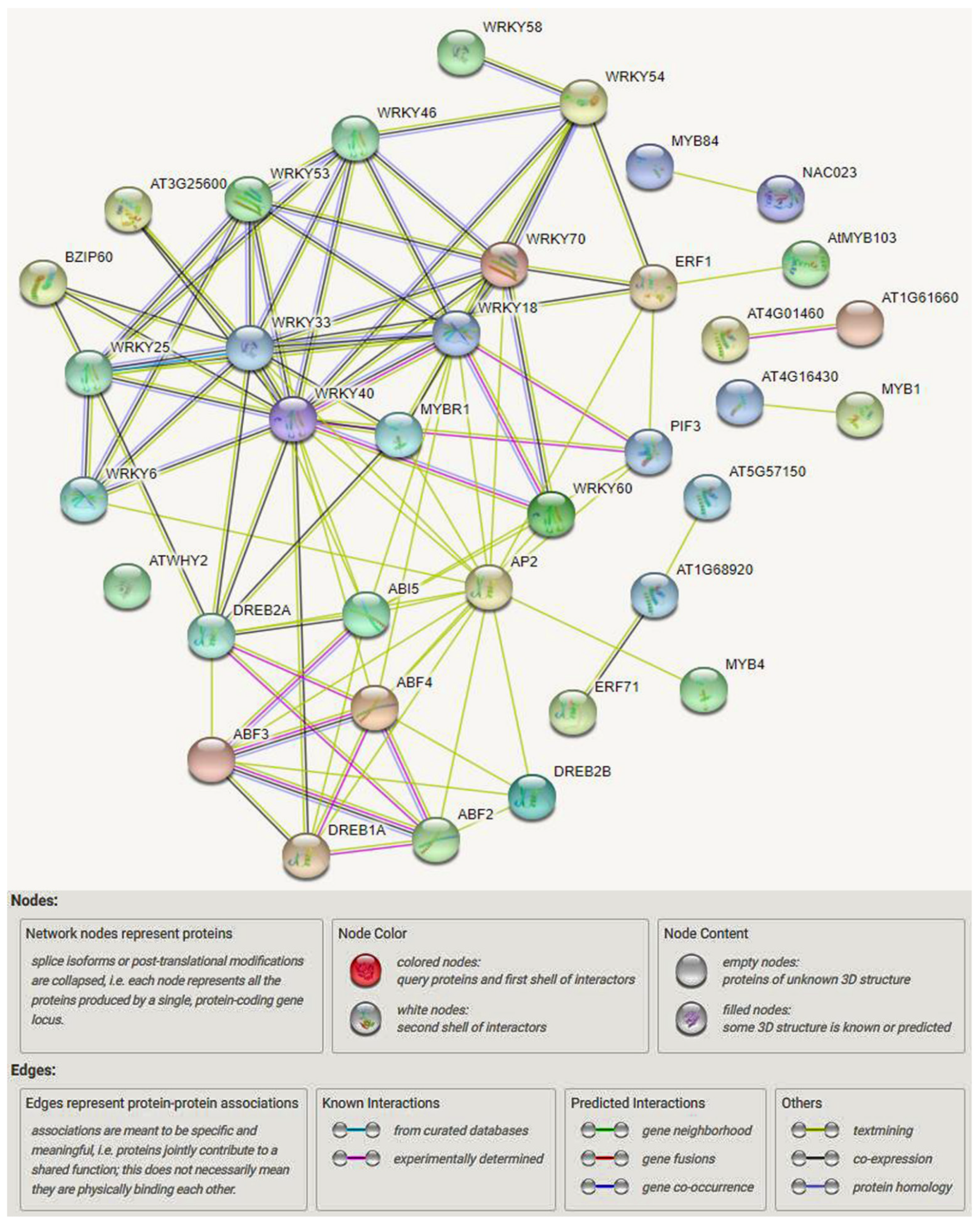
TFs Regulatory Network in Response to Drought and Salinity Stress
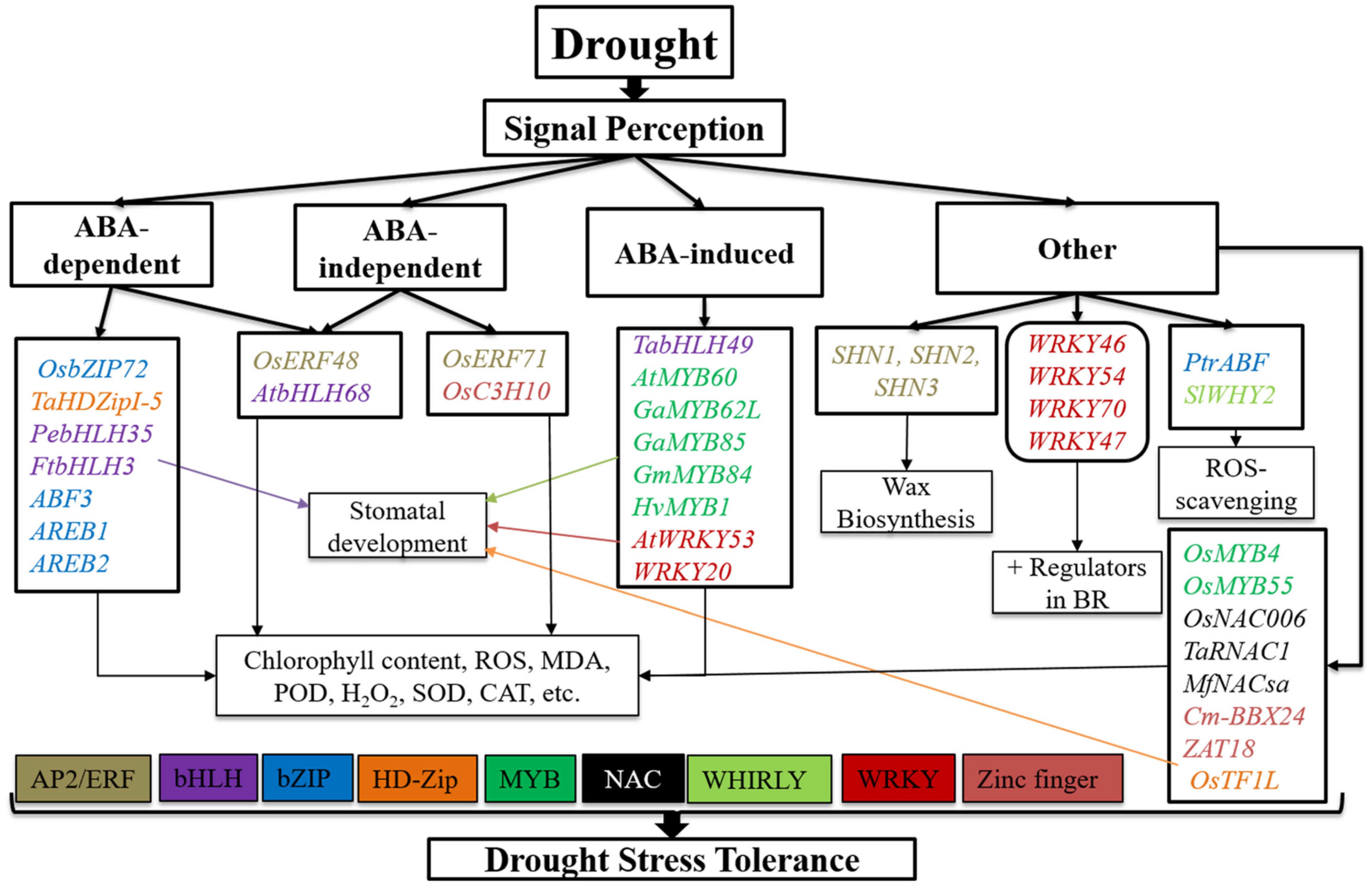
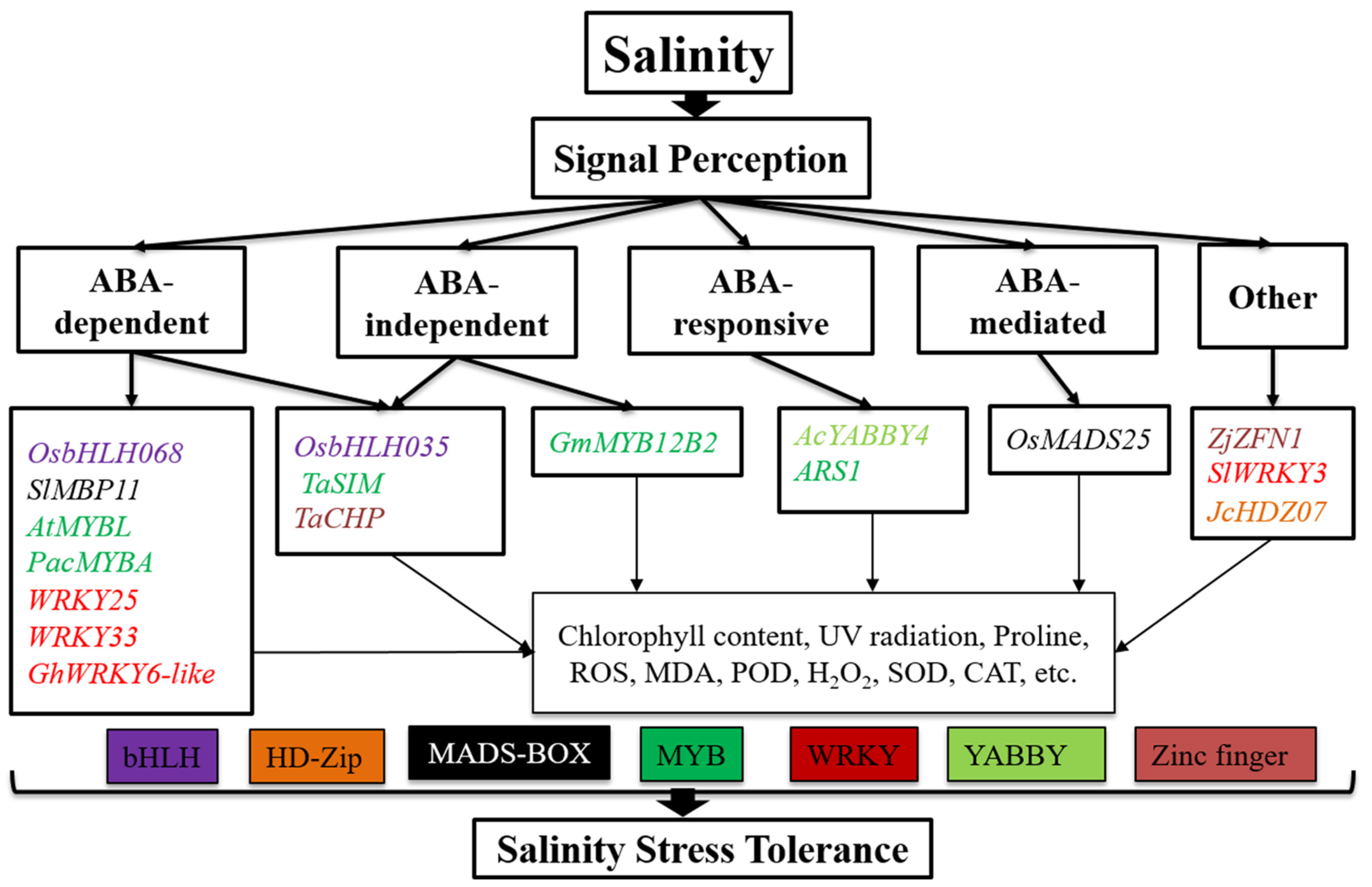
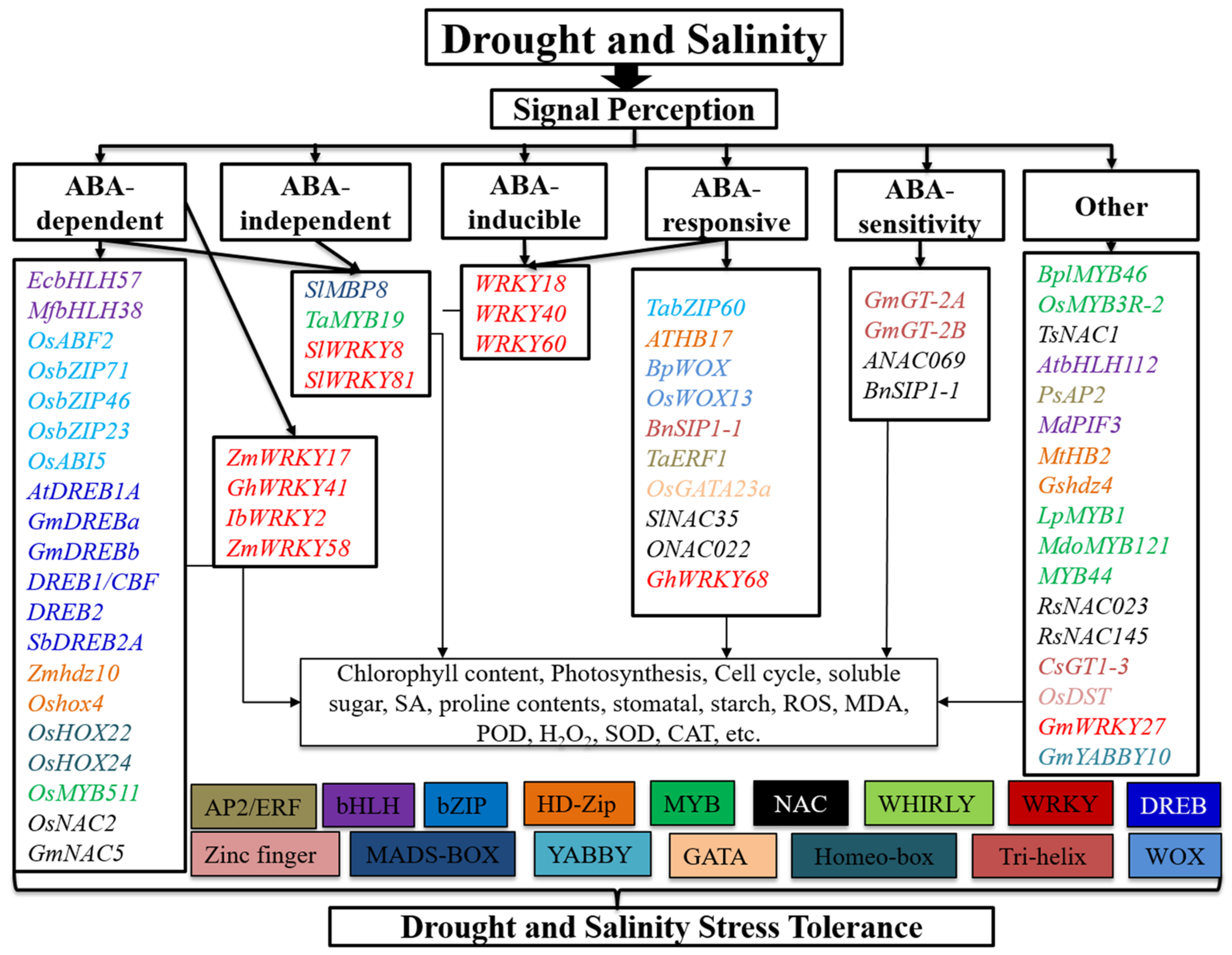
2. TFs Involved in Drought and Salinity Stress Responses
2.1. AP2/ERF
2.2. bHLH
2.3. bZIP
2.4. DREB
2.5. GATA
2.6. HD-Zip
2.7. Homeobox
2.8. MADS-Box
2.9. MYB
2.10. NAC
2.11. Trihelix
2.12. WHIRLY
2.13. WOX
2.14. WRKY
2.15. YABBY
2.16. Zinc Finger
2.17. Other
3. Conclusions and Future Research Priorities
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- He, M.; He, C.Q.; Ding, N.Z. Abiotic stresses: General defenses of land plants and chances for engineering multistress tolerance. Front. Plant Sci. 2018, 871, 1771. [Google Scholar] [CrossRef] [Green Version]
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waqas, M.A.; Kaya, C.; Riaz, A.; Farooq, M.; Nawaz, I.; Wilkes, A.; Li, Y. Potential mechanisms of abiotic stress tolerance in crop plants induced by thiourea. Front. Plant Sci. 2019, 10, 1336. [Google Scholar] [CrossRef] [PubMed]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K.; et al. Abiotic stress responses and microbe-mediated mitigation in plants: The omics strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.; Zhou, P.; Ma, X.; Zhou, L.; Wu, Y.; Ullah, Z.; Wang, S. Transcriptome profiling, biochemical and physiological analyses provide new insights towards drought tolerance in Nicotiana tabacum L. Genes 2019, 10, 1041. [Google Scholar] [CrossRef] [Green Version]
- Chantre Nongpiur, R.; Singla-Pareek, L.; Pareek, A. Genomics approaches for improving salinity stress tolerance in crop plants. Curr. Genom. 2016, 17, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Chen, Y.; Zhao, M.; Zhang, W.H. A novel Medicago truncatula HD-Zip gene, MtHB2, is involved in abiotic stress responses. Environ. Exp. Bot. 2012, 80, 1–9. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, Q.; Jin, X.; Peng, X.; Liu, J.; Deng, L.; Yan, H.; Sheng, L.; Jiang, H.; Cheng, B. A novel maize homeodomain-leucine zipper (HD-Zip) i gene, Zmhdz10, positively regulates drought and salt tolerance in both rice and arabidopsis. Plant Cell Physiol. 2014, 55, 1142–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.; Nutan, K.K.; Singla-pareek, S.L.; Pareek, A. Abiotic stresses cause differential regulation of alternative splice forms of GATA transcription factor in rice. Front. Plant Sci. 2017, 8, 1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Cui, R.; Xu, P.; Wu, J.; Mao, J.L.; Chen, Y.; Zhou, C.Z.; Yu, L.H.; Xiang, C.B. ATHB17 enhances stress tolerance by coordinating photosynthesis associated nuclear gene and ATSIG5 expression in response to abiotic stress. Sci. Rep. 2017, 7, 45492. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis -acting regulatory elements in osmotic- and cold-stress- responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Kazuo, S.; Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: Differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 2000, 3, 217–223. [Google Scholar]
- Ahn, T.S.; Ka, J.O.; Lee, G.H.; Song, H.G. Microcosm study for revegetation of barren land with wild plants by some plant growth-promoting rhizobacteria. J. Microbiol. Biotechnol. 2007, 17, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuteja, N. Abscisic acid and abiotic stress signaling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Liu, X.D.; Chi, X.J.; Wu, C.A.; Li, Y.Z.; Song, L.L.; Liu, X.M.; Wang, Y.F.; Wang, F.W.; Zhang, C.; et al. Dwarf apple MbDREB1 enhances plant tolerance to low temperature, drought, and salt stress via both ABA-dependent and ABA-independent pathways. Planta 2011, 233, 219–229. [Google Scholar] [CrossRef]
- Jakab, G.; Ton, J.; Flors, V.; Zimmerli, L.; Métraux, J.P.; Mauch-Mani, B. Enhancing Arabidopsis salt and drought stress tolerance by chemical priming for its abscisic acid responses. Plant Physiol. 2005, 139, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Hayat, K.; Iqbal, A.; Xie, L. Implications of abscisic acid in the drought stress tolerance of plants. Agronomy 2020, 10, 1323. [Google Scholar] [CrossRef]
- Singh, K.B. Transcriptional regulation in plants: The importance of combinatorial control. Plant Physiol. 1998, 118, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Yanagisawa, S. Transcription factors in rants: Physiological functions and regulation of expression. J. Plant Res. 1998, 111, 363–371. [Google Scholar] [CrossRef]
- Joshi, R.; Wani, S.H.; Singh, B.; Bohra, A.; Dar, Z.A.; Lone, A.A.; Pareek, A.; Singla-Pareek, S.L. Transcription factors and plants response to drought stress: Current understanding and future directions. Front. Plant Sci. 2016, 7, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabara, R.C.; Tripathi, P.; Rushton, P.J. The potential of transcription factor-based genetic engineering in improving crop tolerance to drought. OMICS J. Integr. Biol. 2014, 18, 601–614. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent advances in utilizing transcription factors to improve plant abiotic stress tolerance by transgenic technology. Front. Plant Sci. 2016, 7, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, R.K.; Wilkins, O. Single cell gene regulatory networks in plants: Opportunities for enhancing climate change stress resilience. Plant Cell Environ. 2021, 2006–2017. [Google Scholar] [CrossRef]
- Bartlett, A.; O’Malley, R.C.; Huang, S.S.C.; Galli, M.; Nery, J.R.; Gallavotti, A.; Ecker, J.R. Mapping genome-wide transcription-factor binding sites using DAP-seq. Nat. Protoc. 2017, 12, 1659–1672. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Oh, D.-H.; Duan1, L.; Prashanth Ramachandran, A.R.; Bartlett, A.; Dassanayake, M.; Dinneny, J.R. Divergence in a stress-associated gene regulatory network underlies differential 2 growth control in the Brassicaceae family. Biorxiv Prepr. 2020. [Google Scholar] [CrossRef]
- Zafar, S.A.; Zaidi, S.S.E.A.; Gaba, Y.; Singla-Pareek, S.L.; Dhankher, O.P.; Li, X.; Mansoor, S.; Pareek, A. Engineering abiotic stress tolerance via CRISPR/Cas-mediated genome editing. J. Exp. Bot. 2020, 71, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Erffelinck, M.L.; Ribeiro, B.; Perassolo, M.; Pauwels, L.; Pollier, J.; Storme, V.; Goossens, A. A user-friendly platform for yeast two-hybrid library screening using next generation sequencing. PLoS ONE 2018, 13, e0201270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleman, F.; Yazaki, J.; Lee, M.; Takahashi, Y.; Kim, A.Y.; Li, Z.; Kinoshita, T.; Ecker, J.R.; Schroeder, J.I. An ABA-increased interaction of the PYL6 ABA receptor with MYC2 Transcription Factor: A putative link of ABA and JA signaling. Sci. Rep. 2016, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Latchman, D.S. Transcription factors: An overview. Int. J. Biochem. Cell Biol. 1997, 29, 1305–1312. [Google Scholar] [CrossRef] [Green Version]
- Wärnmark, A.; Treuter, E.; Wright, A.P.H.; Gustafsson, J.Å. Activation functions 1 and 2 of nuclear receptors: Molecular strategies for transcriptional activation. Mol. Endocrinol. 2003, 17, 1901–1909. [Google Scholar] [CrossRef]
- Khan, S.A.; Li, M.Z.; Wang, S.M.; Yin, H.J. Revisiting the role of plant transcription factors in the battle against abiotic stress. Int. J. Mol. Sci. 2018, 19, 1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, M.A.; Dangl, J.L. Functions of the respiratory burst oxidase in biotic interactions, abiotic stress and development. Curr. Opin. Plant Biol. 2005, 8, 397–403. [Google Scholar] [CrossRef]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Huang, C.; Deng, X.; Zhou, S.; Chen, L.; Li, Y.; Wang, C.; Ma, Z.; Yuan, Q.; Wang, Y.; et al. TaASR1, a transcription factor gene in wheat, confers drought stress tolerance in transgenic tobacco. Plant Cell Environ. 2013, 36, 1449–1464. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Ito, Y.; Yamaguchi-Shinozaki, K. Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol. 2009, 149, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Zhou, J.; Deng, R.Y.; Zhao, H.X.; Li, C.L.; Chen, H.; Suzuki, T.; Park, S.U.; Wu, Q. Overexpression of a tartary buckwheat R2R3-MYB transcription factor gene, FtMYB9, enhances tolerance to drought and salt stresses in transgenic Arabidopsis. J. Plant Physiol. 2017, 214, 81–90. [Google Scholar] [CrossRef]
- Cominelli, E.; Galbiati, M.; Tonelli, C. Transcription factors controlling stomatal movements and drought tolerance. Transcription 2010, 1, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, R.K.; Gupta, A.; Ranjan, S.; Singh, R.; Pathre, U.V.; Nath, P.; Sane, A.P. The EAR motif controls the early flowering and senescence phenotype mediated by over-expression of SlERF36 and is partly responsible for changes in stomatal density and photosynthesis. PLoS ONE 2014, 9, e101995. [Google Scholar] [CrossRef] [Green Version]
- Hussain, Q.; Shi, J.; Scheben, A.; Zhan, J.; Wang, X.; Liu, G.; Yan, G.; King, G.J.; Edwards, D.; Wang, H. Genetic and signalling pathways of dry fruit size: Targets for genome editing-based crop improvement. Plant Biotechnol. J. 2020, 18, 1124–1140. [Google Scholar] [CrossRef]
- Mishra, S.; Phukan, U.J.; Tripathi, V.; Singh, D.K. PsAP2 an AP2/ERF family transcription factor from Papaver somniferum enhances abiotic and biotic stress tolerance in transgenic tobacco. Plant Mol. Biol. 2015, 89, 173–186. [Google Scholar] [CrossRef]
- Licausi, F.; Ohme-Takagi, M.; Perata, P. APETALA2/Ethylene Responsive Factor (AP2/ERF) transcription factors: Mediators of stress responses and developmental programs. New Phytol. 2013, 199, 639–649. [Google Scholar] [CrossRef]
- Rashid, M.; Guangyuan, H.; Guangxiao, Y.; Hussain, J.; Xu, Y. AP2/ERF transcription factor in rice: Genome-wide anvas and yntenic relationships between monocots and udicots. Evol. Bioinform. 2012, 2012, 321–355. [Google Scholar] [CrossRef]
- Jung, H.; Chung, P.J.; Park, S.H.; Redillas, M.C.F.R.; Kim, Y.S.; Suh, J.W.; Kim, J.K. Overexpression of OsERF48 causes regulation of OsCML16, a calmodulin-like protein gene that enhances root growth and drought tolerance. Plant Biotechnol. J. 2017, 15, 1295–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.K.; Jung, H.; Jang, G.; Jeong, J.S.; Kim, Y.S.; Ha, S.H.; Do Choi, Y.; Kim, J.K. Overexpression of the OsERF71 transcription factor alters rice root structure and drought resistance. Plant Physiol. 2016, 172, 575–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; Van Arkel, G.; Pereira, A. The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Chen, X.; Liu, J.; Ye, J.; Guo, Z. The rice ERF transcription factor OsERF922 negatively regulates resistance to Magnaporthe oryzae and salt methylation and chromatin patterning tolerance. J. Exp. Bot. 2012, 63, 3899–3912. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.-S.; Xia, L.-Q.; Chen, M.; Cheng, X.-G.; Zhang, R.-Y. Isolation and molecular characterization of the Triticum aestivum L. ethylene-responsive factor 1 (TaERF1) that increases multiple stress tolerance. Plant Mol. Biol. 2007, 1, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Le Hir, R.; Castelain, M.; Chakraborti, D.; Moritz, T.; Dinant, S.; Bellini, C. AtbHLH68 transcription factor contributes to the regulation of ABA homeostasis and drought stress tolerance in Arabidopsis thaliana. Physiol. Plant. 2017, 160, 312–327. [Google Scholar] [CrossRef] [Green Version]
- Yao, P.F.; Li, C.L.; Zhao, X.R.; Li, M.F.; Zhao, H.X.; Guo, J.Y.; Cai, Y.; Chen, H.; Wu, Q. Overexpression of a tartary buckwheat gene, FtbHLH3, enhances drought/oxidative stress tolerance in transgenic Arabidopsis. Front. Plant Sci. 2017, 8, 625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Wang, C.; Han, X.; Tang, S.; Liu, S.; Xia, X.; Yin, W. A novel bHLH transcription factor PebHLH35 from Populus euphratica confers drought tolerance through regulating stomatal development, photosynthesis and growth in Arabidopsis. Biochem. Biophys. Res. Commun. 2014, 450, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, Y.; Liu, D.; Wang, X.; Zhang, L. Transcription factor TabHLH49 positively regulates dehydrin WZY2 gene expression and enhances drought stress tolerance in wheat. BMC Plant Biol. 2020, 20, 259. [Google Scholar] [CrossRef]
- Chen, H.C.; Hsieh-Feng, V.; Liao, P.C.; Cheng, W.H.; Liu, L.Y.; Yang, Y.W.; Lai, M.H.; Chang, M.C. The function of OsbHLH068 is partially redundant with its homolog, AtbHLH112, in the regulation of the salt stress response but has opposite functions to control flowering in Arabidopsis. Plant Mol. Biol. 2017, 94, 531–548. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.C.; Cheng, W.H.; Hong, C.Y.; Chang, Y.S.; Chang, M.C. The transcription factor OsbHLH035 mediates seed germination and enables seedling recovery from salt stress through ABA-dependent and ABA-independent pathways, respectively. Rice 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Tian, X.; Li, S.; Fu, Y.; Xu, J.; Wang, G. The aabhlh35 transcription factor identified from anthurium andraeanum is involved in cold and drought tolerance. Plants 2019, 8, 216. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ji, X.; Nie, X.; Qu, M.; Zheng, L.; Tan, Z.; Zhao, H.; Huo, L.; Liu, S.; Zhang, B.; et al. Arabidopsis AtbHLH112 regulates the expression of genes involved in abiotic stress tolerance by binding to their E-box and GCG-box motifs. New Phytol. 2015, 207, 692–709. [Google Scholar] [CrossRef] [PubMed]
- Babitha, K.C.; Vemanna, R.S.; Nataraja, K.N.; Udayakumar, M. Overexpression of EcbHLH57 transcription factor from Eleusine coracana L. in tobacco confers tolerance to salt, oxidative and drought stress. PLoS ONE 2015, 10, e0137098. [Google Scholar] [CrossRef] [Green Version]
- Zheng, P.F.; Yang, Y.Y.; Zhang, S.; You, C.X.; Zhang, Z.L.; Hao, Y.J. Identification and functional characterization of MdPIF3 in response to cold and drought stress in Malus domestica. Plant Cell Tissue Organ Cult. 2020. [Google Scholar] [CrossRef]
- Qiu, J.; Huang, Z.; Xiang, X.; Xu, W.; Wang, J.; Chen, J.; Song, L.; Xiao, Y. MfbHLH38, a Myrothamnus flabellifolia bHLH transcription factor, confers tolerance to drought and salinity stresses in Arabidopsis. BMC Plant Biol. 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Huang, X.; Liu, J.; Chen, X. Overexpression of PtrABF gene, a bZIP transcription factor isolated from Poncirus trifoliata, enhances dehydration and drought tolerance in tobacco via scavenging ROS and modulating expression of stress-responsive genes. BMC Plant Biol. 2010, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 672–685. [Google Scholar] [CrossRef]
- Lu, G.; Gao, C.; Zheng, X.; Han, B. Identification of OsbZIP72 as a positive regulator of ABA response and drought tolerance in rice. Planta 2009, 229, 605–615. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, L.; Xia, C.; Zhao, G.; Liu, J.; Jia, J.; Kong, X. A novel wheat bZIP transcription factor, TabZIP60, confers multiple abiotic stress tolerances in transgenic Arabidopsis. Physiol. Plant. 2015, 538–554. [Google Scholar] [CrossRef]
- Hossain, A.; Cho, J.; Han, M.; Ahn, C.; Jeon, J.; An, G.; Bum, P. The ABRE-binding bZIP transcription factor OsABF2 is a positive regulator of abiotic stress and ABA signaling in rice. J. Plant Physiol. 2010, 167, 1512–1520. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Mao, B.; Ou, S.; Wang, W.; Liu, L.; Wu, Y.; Chu, C.; Wang, X. OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant Mol. Biol. 2014, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Ma, S.; Zong, W.; Yang, N.; Lv, Y.; Yan, C.; Guo, Z.; Li, J.; Li, X.; Xiang, Y.; et al. MODD mediates deactivation and degradation of OsbZIP46 to negatively regulate ABA signaling and drought resistance in rice. Plant Cell 2016, 28, 2161–2177. [Google Scholar] [CrossRef] [Green Version]
- Tolerance, D.; Xiang, Y.; Tang, N.; Du, H.; Ye, H.; Xiong, L. Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity. Plant Physiol. 2008, 148, 1938–1952. [Google Scholar] [CrossRef] [Green Version]
- Zou, M.; Guan, Y.; Ren, H.; Zhang, F.; Chen, F. A bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance. Plant Mol. Biol. 2008, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Lata, C.; Prasad, M. Role of DREBs in regulation of abiotic stress responses in plants. J. Exp. Bot. 2011, 62, 4731–4748. [Google Scholar] [CrossRef] [Green Version]
- Gujjar, R.S.; Aktar, M.; Singh, M. Transcription factors in abiotic stress tolerance Transcription factors in abiotic stress tolerance. Ind. J. Plant Physiol. 2014, 4, 306–316. [Google Scholar] [CrossRef]
- Li, X.-P.; Tian, A.-G.; Luo, G.-Z.; Gong, Z.-Z.; Zhang, J.-S.; Chen, S.-Y. Soybean DRE-binding transcription factors that are responsive to abiotic stresses. Theor. Appl. Genet. 2004. [Google Scholar] [CrossRef]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta Gene Regul. Mech. 2012, 1819, 86–96. [Google Scholar] [CrossRef]
- Gupta, K.; Agarwal, K.; Reddy, M.K.; Bhavanath, J. SbDREB2A, an A-2 type DREB transcription factor from extreme halophyte Salicornia brachiata confers abiotic stress tolerance in Escherichia coli. Plant Cell Rep. 2010, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wu, T.; Li, Z.; Huang, K.; Kim, N.E.; Ma, Z.; Kwon, S.W.; Jiang, W.; Du, X. OsGATA16, a GATA transcription factor, confers cold tolerance by repressing OsWRKY45–1 at the seedling stage in rice. Rice 2021, 14. [Google Scholar] [CrossRef]
- Yang, Y.; Luang, S.; Harris, J.; Riboni, M.; Li, Y.; Bazanova, N.; Hrmova, M.; Haefele, S.; Kovalchuk, N.; Lopato, S. Overexpression of the class I homeodomain transcription factor TaHDZipI-5 increases drought and frost tolerance in transgenic wheat. Plant Biotechnol. J. 2018, 16, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.W.; Lee, D.K.; Jung, H.; Chung, P.J.; Kim, Y.S.; Do Choi, Y.; Suh, J.W.; Kim, J.K. Overexpression of OsTF1L, a rice HD-Zip transcription factor, promotes lignin biosynthesis and stomatal closure that improves drought tolerance. Plant Biotechnol. J. 2019, 17, 118–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Bao, X.; Wang, S.; Liu, Y.; Tan, J.; Yang, M.; Zhang, M.; Dai, R.; Yu, X. A physic nut stress-responsive HD-Zip transcription factor, JcHDZ07, confers enhanced sensitivity to salinity stress in transgenic arabidopsis. Front. Plant Sci. 2019, 10, 942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.Q.; Zhou, Y.Q.; He, C.Y.; Mou, B.Q.; Zhou, W. Over-expression of oshox4 enhances drought and salinity tolerance in rice. Russ. J. Plant Physiol. 2020, 67, 1152–1162. [Google Scholar] [CrossRef]
- Cao, L.; Yu, Y.; DuanMu, H.; Chen, C.; Duan, X.; Zhu, P.; Chen, R.; Li, Q.; Zhu, Y.; Ding, X. A novel Glycine soja homeodomain-leucine zipper (HD-Zip) I gene, Gshdz4, positively regulates bicarbonate tolerance and responds to osmotic stress in Arabidopsis. BMC Plant Biol. 2016, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, A.; Khurana, J.P.; Jain, M. Characterization of rice homeobox genes, OsHOX22 and OsHOX24, and over-expression of OsHOX24 in transgenic arabidopsis suggest their role in abiotic stress response. Front. Plant Sci. 2016, 7, 627. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Chen, G.; Cui, B.; Gao, Q.; Guo, J.E.; Li, A.; Zhang, L.; Hu, Z. Solanum lycopersicum agamous-like MADS-box protein AGL15-like gene, SlMBP11, confers salt stress tolerance. Mol. Breed. 2016, 36, 125. [Google Scholar] [CrossRef]
- Wu, J.; Yu, C.; Hunag, L.; Wu, M.; Liu, B.; Liu, Y.; Song, G.; Liu, D. Overexpression of MADS—Box transcription factor OsMADS25 enhances salt stress tolerance in rice and arabidopsis. Plant Growth Regul. 2020, 163–171. [Google Scholar] [CrossRef]
- Yin, W.; Hu, Z.; Hu, J.; Zhu, Z.; Yu, X.; Cui, B. Tomato (Solanum lycopersicum) MADS-box transcription factor SlMBP8 regulates drought, salt tolerance and stress-related genes. Plant Growth Regul. 2017, 83, 55–68. [Google Scholar] [CrossRef]
- Cominelli, E.; Galbiati, M.; Vavasseur, A.; Conti, L.; Sala, T.; Vuylsteke, M.; Leonhardt, N.; Dellaporta, S.L.; Tonelli, C. A guard-cell-specific MYB transcription factor regulates stomatal movements and plant drought tolerance. Curr. Biol. 2005, 15, 1196–1200. [Google Scholar] [CrossRef]
- Pasquali, G.; Biricolti, S.; Locatelli, F.; Baldoni, E.; Mattana, M. Osmyb4 expression improves adaptive responses to drought and cold stress in transgenic apples. Plant Cell Rep. 2008, 27, 1677–1686. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, W.; Qin, M.; Li, S.; Qiao, M.; Liu, Z.; Xiang, F. Drought tolerance conferred in soybean (Glycine max. L.) by GmMYB84, a novel R2R3-MYB transcription factor. Plant Cell Physiol. 2017, 58, 1764–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casaretto, J.A.; El-Kereamy, A.; Zeng, B.; Stiegelmeyer, S.M.; Chen, X.; Bi, Y.M.; Rothstein, S.J. Expression of OsMYB55 in maize activates stress-responsive genes and enhances heat and drought tolerance. BMC Genom. 2016, 17, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butt, H.I.; Yang, Z.; Chen, E.; Zhao, G.; Gong, Q.; Yang, Z.; Zhang, X.; Li, F. Functional characterization of cotton GaMYB62L, a novel R2R3 TF in transgenic arabidopsis. PLoS ONE 2017, 12, e0170578. [Google Scholar] [CrossRef] [PubMed]
- Butt, H.I.; Yang, Z.; Gong, Q.; Chen, E.; Wang, X.; Zhao, G.; Ge, X.; Zhang, X.; Li, F. GaMYB85, an R2R3 MYB gene, in transgenic Arabidopsis plays an important role in drought tolerance. BMC Plant Biol. 2017, 17, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ju, H.; Chung, M.; Huang, P.; Ahn, S.; Kim, C.S. The R-R-Type MYB-like transcription factor, AtMYBL, is involved in promoting leaf senescence and modulates an abiotic stress response in arabidopsis. Plant Cell Physiol. 2011, 52, 138–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, J.F.; Cara, B.; Fernando, P.; Pineda, B.; Egea, I.; Flores, F.B. The tomato mutant ars1 (altered response to salt stress 1) identifies an R1-type MYB transcription factor involved in stomatal closure under salt acclimation T-DNA. Plant Biotechnol. J. 2016, 1, 1345–1356. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Guo, X.; Guo, X.; Zhao, D.; Zhao, W.; Chen, J.; Li, T. PacMYBA, a sweet cherry R2R3-MYB transcription factor, is a positive regulator of salt stress tolerance and pathogen resistance. Plant Physiol. Biochem. 2017. [Google Scholar] [CrossRef]
- Li, X.W.; Wang, Y.; Yan, F.; Li, J.W.; Zhao, Y.; Zhao, X.; Zhai, Y. Overexpression of soybean R2R3-MYB transcription factor, GmMYB12B2, and tolerance to UV radiation and salt stress in transgenic Arabidopsis. Genet. Mol. Res. 2016, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ni, Z.; Chen, Q.; Qu, Y. The wheat salinity-induced R2R3-MYB transcription factor TaSIM confers salt stress tolerance in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2017. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Jiang, Y.; Lu, X.; Wang, B.; Zhou, P. A R2R3-MYB transcription factor from Lablab purpureus induced by drought increases tolerance to abiotic stress in Arabidopsis. Mol. Biol. Rep. 2016, 43, 1089–1100. [Google Scholar] [CrossRef]
- Cao, Z.; Zhang, S.; Wang, R.; Zhang, R.; Hao, Y. Genome wide analysis of the apple MYB transcription factor family allows the identification of MdoMYB121 gene confering abiotic stress tolerance in plants. PLoS ONE 2013, 8, e69955. [Google Scholar] [CrossRef]
- Persak, H.; Pitzschke, A. Dominant repression by arabidopsis transcription factor MYB44 causes oxidative damage and hypersensitivity to abiotic stress. Int. J. Mol. Sci. 2014, 15, 2517. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, G.; Zhao, G.; Xia, C.; Jia, J.; Liu, X.; Kong, X. Characterization of a wheat R2R3-MYB transcription factor gene, TaMYB19, involved in enhanced abiotic stresses in arabidopsis. Plant Cell Physiol. 2014, 55, 1802–1812. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Wang, Y.; Wang, L.; Hu, P.; Wang, Y.; Jia, Y.; Zhang, C.; Zhang, Y.; Zhang, Y.; Wang, C.; et al. Expression of the MYB transcription factor gene BplMYB46 affects abiotic stress tolerance and secondary cell wall deposition in Betula platyphylla. Plant Biotechnol. J. 2017, 107–121. [Google Scholar] [CrossRef]
- Huang, P.; Chen, H.; Mu, R.; Yuan, X.; Zhang, H.S.; Huang, J. OsMYB511 encodes a MYB domain transcription activator early regulated by abiotic stress in rice. Genet. Mol. Res. 2015, 14, 9506–9517. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Dai, X.; Xu, Y.; Guo, J.; Liu, Y.; Chen, N.; Xiao, J.; Zhang, D.; Xu, Z.; Zhang, X.; et al. Enhanced tolerance to chilling stress in OsMYB3R-2 transgenic rice is mediated by alteration in cell cycle and ectopic expression of stress genes. Plant Physiol. 2009, 150, 244–256. [Google Scholar] [CrossRef]
- Du, Y.T.; Zhao, M.J.; Wang, C.T.; Gao, Y.; Wang, Y.X.; Liu, Y.W.; Chen, M.; Chen, J.; Zhou, Y.B.; Xu, Z.S.; et al. Identification and characterization of GmMYB118 responses to drought and salt stress. BMC Plant Biol. 2018, 18, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.D.; Wendelboe-Nelson, C.; Morris, P.C. The barley transcription factor HvMYB1 is a positive regulator of drought tolerance. Plant Physiol. Biochem. 2019, 142, 246–253. [Google Scholar] [CrossRef]
- Wang, B.; Zhong, Z.; Wang, X.; Han, X.; Yu, D.; Wang, C.; Song, W.; Zheng, X.; Chen, C.; Zhang, Y. Knockout of the OsNAC006 transcription factor causes drought and heat sensitivity in rice. Int. J. Mol. Sci. 2020, 21, 2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Chai, S.; McIntyre, C.L.; Xue, G.P. Overexpression of a predominantly root-expressed NAC transcription factor in wheat roots enhances root length, biomass and drought tolerance. Plant Cell Rep. 2018, 37, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Duan, M.; Zhang, R.; Zhu, F.; Zhang, Z.; Gou, L.; Wen, J.; Dong, J.; Wang, T. A lipid-anchored NAC transcription factor is translocated into the nucleus and activates glyoxalase i expression during drought stress. Plant Cell 2017, 29, 1748–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Zhang, S.; Ma, X.; Wang, Y.; Kong, F.; Meng, Q. A stress-associated NAC transcription factor ( SlNAC35 ) from tomato plays a positive role in biotic and abiotic stresses A stress-associated NAC transcription factor ( SlNAC35 ) from tomato plays a positive role in biotic and abiotic stresses. Physiol. Plant. 2017, 158, 45–64. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Lv, B.; Luo, L.; He, J.; Mao, C.; Xi, D.; Ming, F. The NAC-type transcription factor OsNAC2 regulates ABA-dependent genes and abiotic stress tolerance in rice. Sci. Rep. 2017, 7, 40641. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, B.; Li, Z.; Peng, Z.; Zhang, J. TsNAC1 Is a key transcription factor in abiotic stress resistance and growth 1. Plant Physiol. 2018, 176, 742–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Xu, G.; Meng, Q.; Huang, F.; Yu, D. Overexpression of the GmNAC2 gene, an NAC transcription factor, reduces GmNAC5, a NAC transcription factor, is a transient response regulator induced by abiotic stress in soybean. Plant Mol. Biol. Rep. 2012. [Google Scholar] [CrossRef]
- Karanja, B.K.; Xu, L.; Wang, Y. Genome-wide characterization and expression profiling of NAC transcription factor genes under abiotic stresses in radish (Raphanus sativus L.). PeerJ 2017, 5, e4172. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Shi, X.; Wang, Y.; Guo, Y.; Yang, K.; Wang, Y. Arabidopsis ANAC069 binds to C[A/G]CG[T/G] sequences to negatively regulate salt and osmotic stress tolerance. Plant Mol. Biol. 2017, 93, 369–387. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Zhang, H.; Huang, L.; Li, D.; Song, F. Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice. Front. Plant Sci. 2016, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, W.; Liu, Z.; Wu, Z.; Zhuang, J. Trihelix family transcription factors in tea plant (Camellia sinensis): Identification, classification, and expression profiles response to abiotic Trihelix family transcription factors in tea plant (Camellia sinensis): Identification, classificati. Acta Physiol. Plant. 2017, 39, 217. [Google Scholar] [CrossRef]
- Xie, Z.; Zou, H.; Lei, G.; Wei, W.; Zhou, Q.; Niu, C. Soybean trihelix transcription factors GmGT-2A and GmGT-2B improve plant tolerance to abiotic stresses in transgenic arabidopsis. PLoS ONE 2009, 4, e6898. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Tang, S.; Mei, F.; Peng, X.; Li, J.; Li, X. BnSIP1-1, a trihelix family gene, mediates abiotic stress tolerance and ABA signaling in brassica napus. Front. Plant Sci. 2017, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wu, D.; Shan, T.; Xu, S.; Qin, R.; Li, H.; Negm, M.; Wu, D.; Li, J. The trihelix transcription factor OsGTγ-2 is involved adaption to salt stress in rice. Plant Mol. Biol. 2020. [Google Scholar] [CrossRef]
- Xi, J.; Qiu, Y.; Du, L.; Poovaiah, B.W. Plant science plant-specific trihelix transcription factor AtGT2L interacts with calcium/calmodulin and responds to cold and salt stresses. Plant Sci. 2012, 185–186, 274–280. [Google Scholar] [CrossRef]
- Xu, H.; Shi, X.; He, L.; Guo, Y.; Zang, D.; Li, H.; Zhang, W.; Wang, Y. Arabidopsis thaliana trihelix transcription factor AST1 mediates salt and osmotic stress tolerance by binding to a novel AGAG-box and some GT motifs. Plant Cell Physiol. 2018, 59, 946–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Wang, G.; Zhao, W.; Zhang, S.; Kong, F.; Dong, X. Overexpression of tomato WHIRLY protein enhances tolerance to drought stress and resistance to Pseudomonas solanacearum in transgenic tobacco. Biol. Plant. 2018, 62, 55–68. [Google Scholar] [CrossRef]
- Tang, F.; Chen, N.; Zhao, M.; Wang, Y.; He, R.; Peng, X.; Shen, S. Identification and functional divergence analysis of WOX gene family in paper mulberry. Int. J. Mol. Sci. 2017, 1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh-Thu, P.-T.; Kim, J.S.; Chae, S.; Jun, K.M.; Lee, G.-S.; Kim, D.-E.; Cheong, J.-J.; Song, S.I.; Nahm, B.H.; Kim, Y.-K. Molecules and cells minireview a WUSCHEL homeobox transcription factor, OsWOX13, enhances drought tolerance and triggers early flowering in rice. Mol. Cell. Biol. 2018, 41, 781–798. [Google Scholar]
- Chen, J.; Nolan, T.M.; Ye, H.; Zhang, M.; Tong, H.; Xin, P.; Chu, J. Arabidopsis WRKY46, WRKY54, and WRKY70 transcription factors are involved in brassinosteroid-regulated plant growth and drought responses. Plant Cell 2017, 29, 1425–1439. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yu, D. Activated expression of AtWRKY53 negatively regulates drought tolerance by mediating stomatal movement. Plant Cell Rep. 2015. [Google Scholar] [CrossRef]
- Raineri, J.; Wang, S.; Peleg, Z.; Blumwald, E.; Chan, R.L. The rice transcription factor OsWRKY47 is a positive regulator of the response to water deficit stress. Plant Mol. Biol. 2015. [Google Scholar] [CrossRef]
- Luo, X.; Bai, X.; Sun, X.; Zhu, D.; Liu, B.; Ji, W.; Cai, H.; Cao, L.; Wu, J.; Hu, M. Expression of wild soybean WRKY20 in Arabidopsis enhances drought tolerance and regulates ABA signalling. J. Exp. Bot. 2013. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Deyholos, M.K. Functional characterization of Arabidopsis NaCl-inducible WRKY25 and WRKY33 transcription factors in abiotic stresses. Plant Mol. Biol. 2009, 69, 91–105. [Google Scholar] [CrossRef]
- Ullah, A.; Sun, H.; Yang, X. A novel cotton WRKY gene, GhWRKY6 -like, improves salt tolerance by activating the ABA signaling pathway and. Physiol. Plant. 2018, 439–454. [Google Scholar] [CrossRef] [PubMed]
- Hichri, I.; Muhovski, Y.; Žižková, E.; Dobrev, P.I.; Gharbi, E.; Franco-zorrilla, J.M.; Lopez-vidriero, I.; Solano, R.; Clippe, A.; Errachid, A.; et al. The solanum lycopersicum WRKY3 transcription factor SlWRKY3 is involved in salt stress tolerance in tomato. Front. Plant Sci. 2017, 8, 1343. [Google Scholar] [CrossRef]
- Chen, H.; Lai, Z.; Shi, J.; Xiao, Y.; Chen, Z.; Xu, X. Roles of arabidopsis WRKY18, WRKY40 and WRKY60 transcription factors in plant responses to abscisic acid and abiotic stress. BMC Plant Biol. 2010, 10, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, R.; Dai, W.; Zhang, C.; Wang, Y.; Wu, M.; Zhao, Y. The maize WRKY transcription factor ZmWRKY17 negatively regulates salt stress tolerance in transgenic Arabidopsis plants. Planta 2017, 246, 1215–1231. [Google Scholar] [CrossRef]
- Chu, X.; Wang, C.; Chen, X.; Lu, W.; Li, H.; Wang, X.; Hao, L. The Cotton WRKY gene GhWRKY41 positively regulates salt and drought stress tolerance in transgenic nicotiana benthamiana. PLoS ONE 2015, 10, e0143022. [Google Scholar] [CrossRef]
- Jia, H.; Wang, C.; Wang, F.; Liu, S.; Li, G.; Guo, X. GhWRKY68 reduces resistance to salt and drought in transgenic nicotiana benthamiana. PLoS ONE 2015, 10, e0120646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Zhang, Z.; Dong, J.; Wang, T. Overexpression of MtWRKY76 increases both salt and drought tolerance in Medicago truncatula. Environ. Exp. Bot. 2016, 123, 50–58. [Google Scholar] [CrossRef]
- Wang, F.; Chen, H.W.; Li, Q.T.; Wei, W.; Li, W.; Zhang, W.K.; Ma, B.; Bi, Y.D.; Lai, Y.C. GmWRKY27 interacts with GmMYB174 to reduce expression of GmNAC29 for stress tolerance in soybean plants. Plant J. 2015, 224–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Zhou, Y.; Zhai, H.; He, S.; Zhao, N.; Liu, Q. A novel sweetpotato WRKY transcription factor, IBWRKY2, positively regulates drought and salt tolerance in transgenic arabidopsis. Biomolecules 2020, 10, 506. [Google Scholar] [CrossRef] [Green Version]
- Cai, R.; Zhao, Y.; Wang, Y.; Lin, Y.; Peng, X. Overexpression of a maize WRKY58 gene enhances drought and salt tolerance in transgenic rice. Plant Cell Tissue Organ Cult. 2014. [Google Scholar] [CrossRef]
- Gao, Y.F.; Liu, J.K.; Yang, F.M.; Zhang, G.Y.; Wang, D.; Zhang, L.; Ou, Y.B.; Yao, Y.A. The WRKY transcription factor WRKY8 promotes resistance to pathogen infection and mediates drought and salt stress tolerance in Solanum lycopersicum. Physiol. Plant. 2020, 168, 98–117. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Li, X.; Yang, Y.; Liu, C.; Zhou, G.; Wan, H.; Cheng, Y. Tomato WRKY81 acts as a negative regulator for drought tolerance by modulating guard cell H2O2–mediated stomatal closure. Environ. Exp. Bot. 2020, 171, 103960. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Li, X.; Mao, Q.; Wan, H.; Zhou, G.; Cheng, Y. The SlWRKY81 transcription factor inhibits stomatal closure by attenuating nitric oxide accumulation in the guard cells of tomato under drought. Physiol. Plant. 2021, 172, 885–895. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Li, X.; Wan, H.; Zhou, G.; Cheng, Y. SlWRKY81 reduces drought tolerance by attenuating proline biosynthesis in tomato. Sci. Hortic. 2020, 270, 109444. [Google Scholar] [CrossRef]
- Li, Z.; Li, G.; Cai, M.; Priyadarshani, S.V.G.N.; Aslam, M.; Zhou, Q.; Huang, X.; Wang, X.; Liu, Y.; Qin, Y. Genome-wide analysis of the YABBY transcription factor family in pineapple and functional identification of AcYABBY4 involvement in salt stress. Int. J. Mol. Sci. 2019, 20, 5863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.P.; Lu, D.; Yu, T.F.; Ji, Y.J.; Zheng, W.J.; Zhang, S.X.; Chai, S.C.; Chen, Z.Y.; Cui, X.Y. Genome-wide analysis of the YABBY family in soybean and functional identification of GmYABBY10 involvement in high salt and drought stresses. Plant Physiol. Biochem. 2017, 119, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ma, C.; Xu, Y.; Wei, Q.; Imtiaz, M.; Lan, H.; Gao, S.; Cheng, L.; Wang, M.; Fei, Z.; et al. A zinc finger protein regulates flowering time and abiotic stress tolerance in chrysanthemum by modulating gibberellin biosynthesis. Plant Cell 2014, 26, 2038–2054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seong, S.Y.; Shim, J.S.; Bang, S.W.; Kim, J.K. Overexpression of osc3h10, a ccch-zinc finger, improves drought tolerance in rice by regulating stress-related genes. Plants 2020, 9, 1298. [Google Scholar] [CrossRef]
- Yin, M.; Wang, Y.; Zhang, L.; Li, J.; Quan, W.; Yang, L.; Wang, Q.; Chan, Z. The arabidopsis Cys2/His2 zinc finger transcription factor ZAT18 is a positive regulator of plant tolerance to drought stress. J. Exp. Bot. 2017, 68, 2991–3005. [Google Scholar] [CrossRef]
- Teng, K.; Tan, P.; Guo, W.; Yue, Y.; Fan, X.; Wu, J. Heterologous expression of a novel zoysia japonica c2H2 zinc finger gene, ZjZFN1, improved salt tolerance in arabidopsis. Front. Plant Sci. 2018, 9, 1159. [Google Scholar] [CrossRef]
- Li, C.; Lv, J.; Zhao, X.; Ai, X.; Zhu, X.; Wang, M.; Zhao, S.; Xia, G. TaCHP: A wheat zinc finger protein gene down-regulated by abscisic acid and salinity stress plays a positive role in stress tolerance. Plant Physiol. 2010, 154, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Santosh Kumar, V.V.; Verma, R.K.; Yadav, S.K.; Yadav, P.; Watts, A.; Rao, M.V.; Chinnusamy, V. CRISPR-Cas9 mediated genome editing of drought and salt tolerance (OsDST) gene in indica mega rice cultivar MTU1010. Physiol. Mol. Biol. Plants 2020, 26, 1099–1110. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, N.; Zhou, G.; Hussain, S.; Ahmed, S.; Tian, H.; Wang, S. Knockout of the entire family of AITR genes in Arabidopsis leads to enhanced drought and salinity tolerance without fitness costs. BMC Plant Biol. 2021, 21, 1–15. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | TFs | Function | Expression | Species | References |
|---|---|---|---|---|---|
| OsERF48 | AP2/ERF | Enhances root growth | Seedlings roots | Rice | [46] |
| OsERF71 | AP2/ERF | Cell wall modification, root structure | Root meristem | Rice | [47] |
| SHN1 | AP2/ERF | Activates Wax Biosynthesis | Flower | Arabidopsis | [48] |
| SHN2 | AP2/ERF | Activates Wax Biosynthesis | Flower | Arabidopsis | [48] |
| SHN3 | AP2/ERF | Activates Wax Biosynthesis | Flower | Arabidopsis | [48] |
| OsERF922 | AP2/ERF | Modulation of the ABA levels | Shoot | Rice | [49] |
| PsAP2 | AP2/ERF | Regulate the level of RNS and ROS | Leaves, floral bud, root | Papaver somniferum | [43] |
| TaERF1 | AP2/ERF | Stress signal transduction pathways | Leaf | Wheat | [50] |
| AtbHLH68 | bHLH | Regulation of lateral root elongation | Shoot and root | Arabidopsis | [51] |
| FtbHLH3 | bHLH | Activating the antioxidant system | Root, stem, flower, and leaves | Fagopyrum tataricum | [52] |
| PebHLH35 | bHLH | Regulating stomatal density and aperture | Root and leaf | Arabidopsis | [53] |
| TabHLH49 | bHLH | Regulates dehydrin WZY2 gene expression | Leaves, stem and roots | wheat | [54] |
| OsbHLH068 | bHLH | Control flowering | Leaves and aerial tissues | Arabidopsis | [55] |
| OsbHLH035 | bHLH | Reduces ABA levels | Germinating seeds, seedlings | Rice | [56] |
| AabHLH35 | bHLH | Improved tolerance to drought stress | Leaf | Anthurium andraeanum | [57] |
| AtbHLH112 | bHLH | Proline biosynthesis and ROS scavenging | Root, leaves | Arabidopsis | [58] |
| EcbHLH57 | bHLH | Improved root growth | Leaf, root | Finger millet | [59] |
| MdPIF3 | bHLH | Positively regulates the drought resistance | Root | Malus domestica | [60] |
| MfbHLH38 | bHLH | Regulating osmotic balance | Leaves, roots | Myrothamnus flabellifolia | [61] |
| PtrABF | bZIP | Scavenging ROS and enhances dehydration | Leaves | Poncirus trifoliate | [62] |
| ABF3 | bZIP | Activate target genes in ABA signaling | Root | Arabidopsis | [63] |
| AREB1 | bZIP | Activate target genes in ABA signaling | Root | Arabidopsis | [63] |
| AREB2 | bZIP | Activate target genes in ABA signaling | Root | Arabidopsis | [63] |
| OsbZIP72 | bZIP | Positive regulator of ABA response | Seedlings | Rice | [64] |
| TabZIP60 | bZIP | Increased plant sensitivity to ABA | Spikes, leaves, stems | Wheat | [65] |
| OsABF2 | bZIP | Positive regulator of ABA signaling | Various rice tissues | Rice | [66] |
| OsbZIP71 | bZIP | Important role in ABA-mediated | Root, shoot | Rice | [67] |
| OsbZIP46 | bZIP | Negatively regulate ABA signaling | Leaf | Rice | [68] |
| OsbZIP23 | bZIP | Improved sensitivity to ABA | Leaves, root, shoot | Rice | [69] |
| OsABI5 | bZIP | Low fertility | Mature pollen | Rice | [70] |
| AtDREB1A | DREB | Higher proline and SOD activity | Leaves | Arabidopsis | [72] |
| GmDREBa | DREB | Transcriptional activation ability | Leaves, seedlings | Soybean | [73] |
| GmDREBb | DREB | Transcriptional activation ability | Leaves, seedlings | Soybean | [73] |
| DREB1/CBF | DREB | Acquisition of stress tolerance | Seed maturation | Arabidopsis | [74] |
| DREB2 | DREB | Acquisition of stress tolerance | Seed maturation | Arabidopsis | [74] |
| SbDREB2A | DREB | Response to stress | Leaves and root | Salicornia brachiata | [75] |
| OsGATA23a | GATA | Response to environmental signals | Seedling, stem | Rice | [11] |
| OsGATA16 | GATA | Enhanced drought tolerance | Panicles, guard cells | Rice | [76] |
| TaHDZipI-5 | HD-Zip | Delayed flowering and a grain yield decrease | Flowers and grains | Wheat | [77] |
| OsTF1L | HD-Zip | Lignin biosynthesis and stomatal closure | Root, shoot, flower | Rice | [78] |
| JcHDZ07 | HD-Zip | Changes in physiological indexes | Roots, leaves, seeds | Arabidopsis/Nut | [79] |
| MtHB2 | HD-Zip | Negative role in regulation of abiotic stress | Pods, leaves, root, stem | Medicago truncatula | [9] |
| Zmhdz10 | HD-Zip | ABA signal transduction pathway | Root, stem, tassels, ears, leaf | Maize | [10] |
| ATHB17 | HD-Zip | Alleviating the damage to chloroplast | Root, leaves | Arabidopsis | [12] |
| Oshox4 | HD-Zip | Controlling ABA signal perception | Leaves | Rice | [80] |
| Gshdz4 | HD-Zip | Positively regulates bicarbonate | Leaves, stem, root | Soybean | [81] |
| OsHOX22 | Homeobox | Higher sensitivity to ABA and hormones | Root, fresh weight | Rice | [82] |
| OsHOX24 | Homeobox | Higher sensitivity to ABA and hormones | Root, fresh weight | Rice | [82] |
| SlMBP11 | MADS-box | Higher chlorophyll content, higher MDA | Root and shoot | Arabidopsis | [83] |
| OsMADS25 | MADS-box | Higher proline contents, MDA | Seedling, shoot and root | Rice | [84] |
| SlMBP8 | MADS-box | Negative regulator in stress response | Root, sepals and fruits | Tomato | [85] |
| AtMYB60 | MYB | Stomatal Movements | Seedling, stem, leaves, flower | Arabidopsis | [86] |
| OsMYB4 | MYB | Improved physiological and biochemical adaptation | Leaves, root, stem, flower, seed | Rice/Transgenic Apple | [87] |
| GmMYB84 | MYB | Improves drought stress response and promotes root growth | Root and flower | Soybean | [88] |
| OsMYB55 | MYB | Encoding proteins involved in general defense responses and abiotic stress | Seedlings | Rice/Maize | [89] |
| GaMYB62L | MYB | Enhanced the expression of ABA | Root and leaves | Arabidopsis | [90] |
| GaMYB85 | MYB | Reduced stomatal density, with greater stomatal size | Seedlings | Cotton | [91] |
| AtMYBL | MYB | Promoting leaf senescence | Leaves | Arabidopsis | [92] |
| ARS1 | MYB | Stomatal closure | Root, flower, leaves | Tomato | [93] |
| PacMYBA | MYB | Pathogen resistance | Leaf | Sweet cherry | [94] |
| GmMYB12B2 | MYB | Regulates UV radiation | Seedlings | Soybean | [95] |
| TaSIM | MYB | Improve crop resistance to salt stresses | Root, leaf, and stem | Wheat | [96] |
| LpMYB1 | MYB | Improve the drought and salt tolerance | Seedling, root, seeds | Lablab purpureus | [97] |
| MdoMYB121 | MYB | Roles in secondary metabolism | Seed germination, seedling | Tomato/Apple | [98] |
| MYB44 | MYB | Oxidative damage and hypersensitivity | Leaves, seedlings | Arabidopsis | [99] |
| TaMYB19 | MYB | Leads to improved stress tolerance | Root, seedlings | Wheat | [100] |
| BplMYB46 | MYB | Affects secondary cell wall deposition | Stem, leaves, root | Betula platyphylla | [101] |
| OsMYB511 | MYB | Panicle development | Panicles at an earlier stage | Rice | [102] |
| OsMYB3R-2 | MYB | Mediated by regulating the cell cycle | Seedling | Rice | [103] |
| GmMYB118 | MYB | Reducing the contents of ROS and MDA | Root | Soybean | [104] |
| HvMYB1 | MYB | Acting as a mediator of ABA action | Roots and leaves | Barley | [105] |
| OsNAC006 | NAC | Responses to stimuli, cofactor binding | Stems and leaves | Rice | [106] |
| TaRNAC1 | NAC | Enlargement of the root system | Root | Wheat | [107] |
| MfNACsa | NAC | Oxidation-reduction and lipid transport | Root and leaves | Medicago falcata | [108] |
| SlNAC35 | NAC | Involving auxin and SA signaling | Roots | Tomato | [109] |
| OsNAC2 | NAC | Regulates both abiotic stress responses and ABA-dependent | Root and leaves | Rice | [110] |
| TsNAC1 | NAC | Regulates the expansion of cells | Root, mature tissues, shoot | T. halophila | [111] |
| GmNAC5 | NAC | Involved in seed development and abiotic stress responses | Roots and immature seeds | Soybean | [112] |
| RsNAC023 | NAC | Reacted to all stresses except ABA | Roots, flowers, and leaves | Radish | [113] |
| RsNAC145 | NAC | Reacted to all stresses except ABA | Root, flower, and leaves | Radish | [113] |
| ANAC069 | NAC | Decreased ROS scavenging capability and proline biosynthesis | Leaves, stems, siliques | Arabidopsis | [114] |
| ONAC022 | NAC | Modulating an ABA-mediated pathway | Seedling and panicles | Rice | [115] |
| CsGT1-3 | Tri-helix | Stress tolerance | Leaves | Tea Plant | [116] |
| CsGT2-1 | Tri-helix | Stress tolerance | Leaves | Tea Plant | [116] |
| GmGT-2A | Tri-helix | Regulate plant stress responses | Stem, pods | Soybean | [117] |
| GmGT-2B | Tri-helix | Regulate plant stress responses | Stem, pods | Soybean | [117] |
| BnSIP1-1 | Tri-helix | Roles in ABA synthesis and signaling | Roots, stems, leaves, pollens | Brassica napus | [118] |
| OsGTγ-2 | Tri-helix | Regulating salinity adaptation | Roots, stems and seeds | Rice | [119] |
| AtGT2L | Tri-helix | Interacts with calcium/calmodulin | Flowers and leaves | Arabidopsis | [120] |
| AST1 | Tri-helix | Reduced ROS accumulation | Leaves, stems, and anthers | Arabidopsis | [121] |
| SlWHY2 | WHIRLY | Reducing ROS accumulation | Pollens | Tomato | [122] |
| BpWOX | WOX | Plant development and stress responses | Apical bud, stem, and root | Paper mulberry | [123] |
| OsWOX13 | WOX | Triggers early flowering | Leaves | Rice | [124] |
| WRKY46 | WRKY | BR-regulated plant growth | Leaves | Arabidopsis | [125] |
| WRKY54 | WRKY | BR-regulated plant growth | Leaves | Arabidopsis | [125] |
| WRKY70 | WRKY | BR-regulated plant growth | Leaves | Arabidopsis | [125] |
| AtWRKY53 | WRKY | Mediating stomatal movement | Guard cells | Arabidopsis | [126] |
| OsWRKY47 | WRKY | Reduction in photosynthesis and high yields | Leaves | Rice | [127] |
| WRKY20 | WRKY | Regulates ABA signaling | Seedlings | Soybean | [128] |
| WRKY25 | WRKY | Increasing sensitivity to ABA | Leaves, siliques, flower, root | Arabidopsis | [129] |
| WRKY33 | WRKY | Increasing sensitivity to ABA | Leaves, siliques, flower, root | Arabidopsis | [129] |
| GhWRKY6-like | WRKY | Activating the ABA signaling pathway, scavenging of ROS | Roots, stem, leaves, flowers, and anthers | Cotton | [130] |
| SlWRKY3 | WRKY | Regulation of senescence related process | Leaves and mature fruit | Tomato | [131] |
| WRKY18 | WRKY | Plant defense and stress responses | Seed germination and root | Arabidopsis | [132] |
| WRKY40 | WRKY | Plant defense and stress responses | Seed germination and root | Arabidopsis | [132] |
| WRKY60 | WRKY | Plant defense and stress responses | Seed germination and root | Arabidopsis | [132] |
| ZmWRKY17 | WRKY | Decreased ABA sensitivity | Tassels, Leaf, root | Maize | [133] |
| GhWRKY41 | WRKY | Enhanced stomatal closure | Stomata | Cotton | [134] |
| GhWRKY68 | WRKY | Regulating ABA signaling | Leaf | Cotton | [135] |
| MtWRKY76 | WRKY | Increased salt and drought tolerance | Root, seedling | Medicago truncatula | [136] |
| GmWRKY27 | WRKY | Improvements in stress tolerance | Root, cotyledons | Soybean | [137] |
| IbWRKY2 | WRKY | Enhancing the tolerance to abiotic stress | Seedling, leaves, germination | Sweet potato | [138] |
| ZmWRKY58 | WRKY | Positive regulator of stress tolerance | Root, leaf, germination | Rice/Maize | [139] |
| SlWRKY8 | WRKY | Resistance to pathogen infection | Stem, roots, flowers | Tomato | [140] |
| SlWRKY81 | WRKY | Regulator of stomatal closure | Leaves | Tomato | [141,142,143] |
| AcYABBY4 | YABBY | Important role in response to ABA | Sepal and petal | Pineapple | [144] |
| GmYABBY10 | YABBY | Highly sensitive in drought | Seedling, root, germination | Soybean | [145] |
| Cm-BBX24 | Zinc finger | Modulating gibberellin biosynthesis | Root, leaves, stem | Chrysanthemum | [146] |
| OsC3H10 | Zinc finger | Response to drought | Seeds | Rice | [147] |
| ZAT18 | Zinc finger | Positive drought stress regulator | Stems, siliques, leaves | Arabidopsis | [148] |
| ZjZFN1 | Zinc finger | Stress responses in seed germination | Leaf, stem, root | Zoysia japonica | [149] |
| TaCHP | Zinc finger | Enhances stress tolerance | Roots, leaf | Wheat | [150] |
| OsDST | Zinc finger | Stomatal density | Flag leaf | Rice | [151] |
| OsRR22 | Other | Enhanced the tolerance to salinity | Shoot | Rice | [152] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, Q.; Asim, M.; Zhang, R.; Khan, R.; Farooq, S.; Wu, J. Transcription Factors Interact with ABA through Gene Expression and Signaling Pathways to Mitigate Drought and Salinity Stress. Biomolecules 2021, 11, 1159. https://doi.org/10.3390/biom11081159
Hussain Q, Asim M, Zhang R, Khan R, Farooq S, Wu J. Transcription Factors Interact with ABA through Gene Expression and Signaling Pathways to Mitigate Drought and Salinity Stress. Biomolecules. 2021; 11(8):1159. https://doi.org/10.3390/biom11081159
Chicago/Turabian StyleHussain, Quaid, Muhammad Asim, Rui Zhang, Rayyan Khan, Saqib Farooq, and Jiasheng Wu. 2021. "Transcription Factors Interact with ABA through Gene Expression and Signaling Pathways to Mitigate Drought and Salinity Stress" Biomolecules 11, no. 8: 1159. https://doi.org/10.3390/biom11081159
APA StyleHussain, Q., Asim, M., Zhang, R., Khan, R., Farooq, S., & Wu, J. (2021). Transcription Factors Interact with ABA through Gene Expression and Signaling Pathways to Mitigate Drought and Salinity Stress. Biomolecules, 11(8), 1159. https://doi.org/10.3390/biom11081159