
Amelioration of Chlorpyrifos-Induced Toxicity in Brassica juncea L. by Combination of 24-Epibrassinolide and Plant-Growth-Promoting Rhizobacteria

 , ,
, ,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Treatment with Microbes
2.2. Plant Material and Experimental Plot
2.3. Analysis of Growth Parameters
2.4. Cell Injury
2.5. Analysis of Oxidative Stress Markers
2.5.1. Superoxide Anion (O2−)
2.5.2. Hydrogen Peroxide (H2O2)
2.6. Estimation of MDA Content
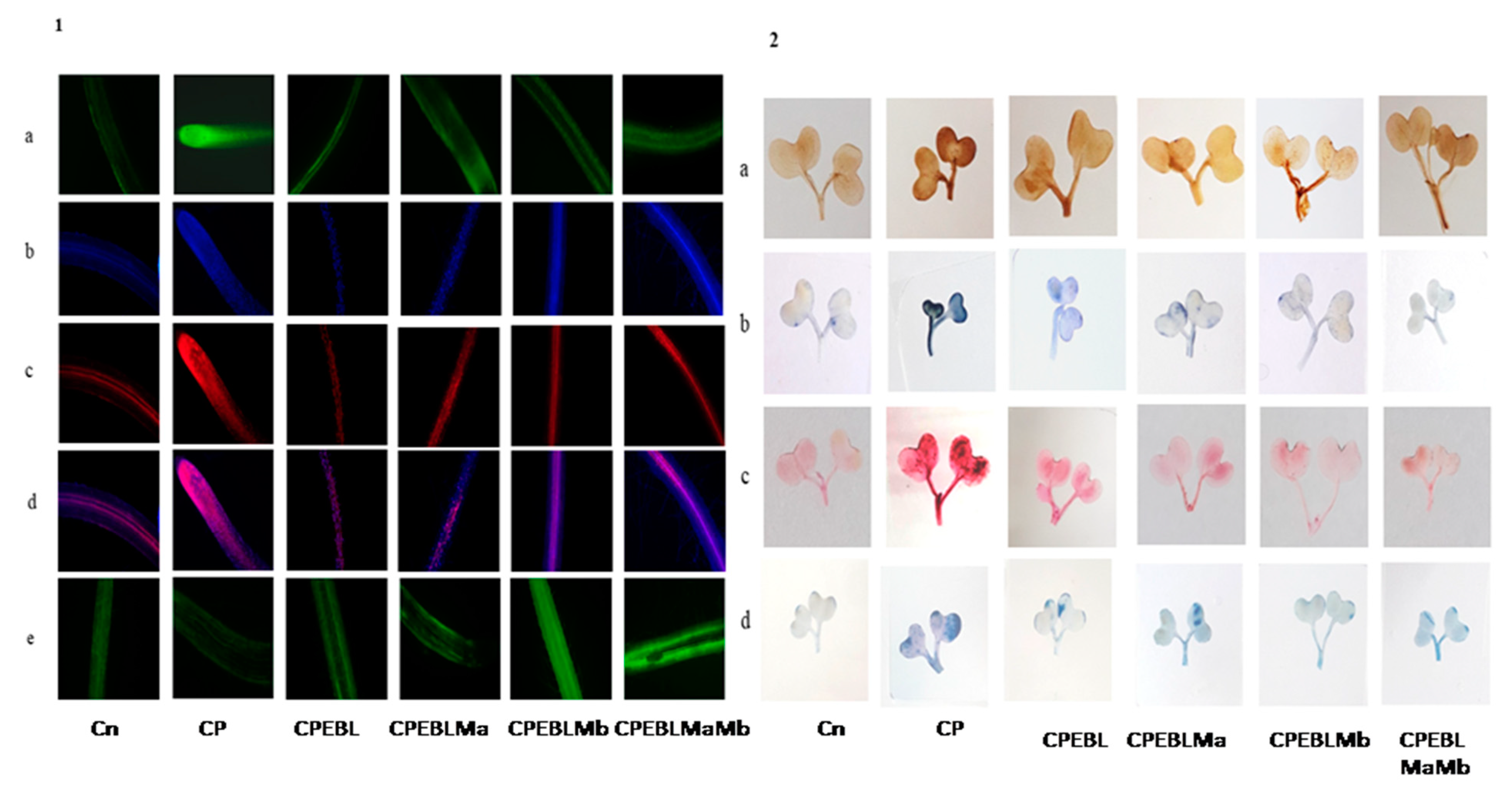
2.7. Fluorescent Imaging of Nuclear and Membrane Damage
2.8. Estimation of Nitrate Reductase and NO Content
2.9. Antioxidative Defence System
2.10. Analysis of Gene Expression through qRT-PCR
2.11. Statistical Analysis
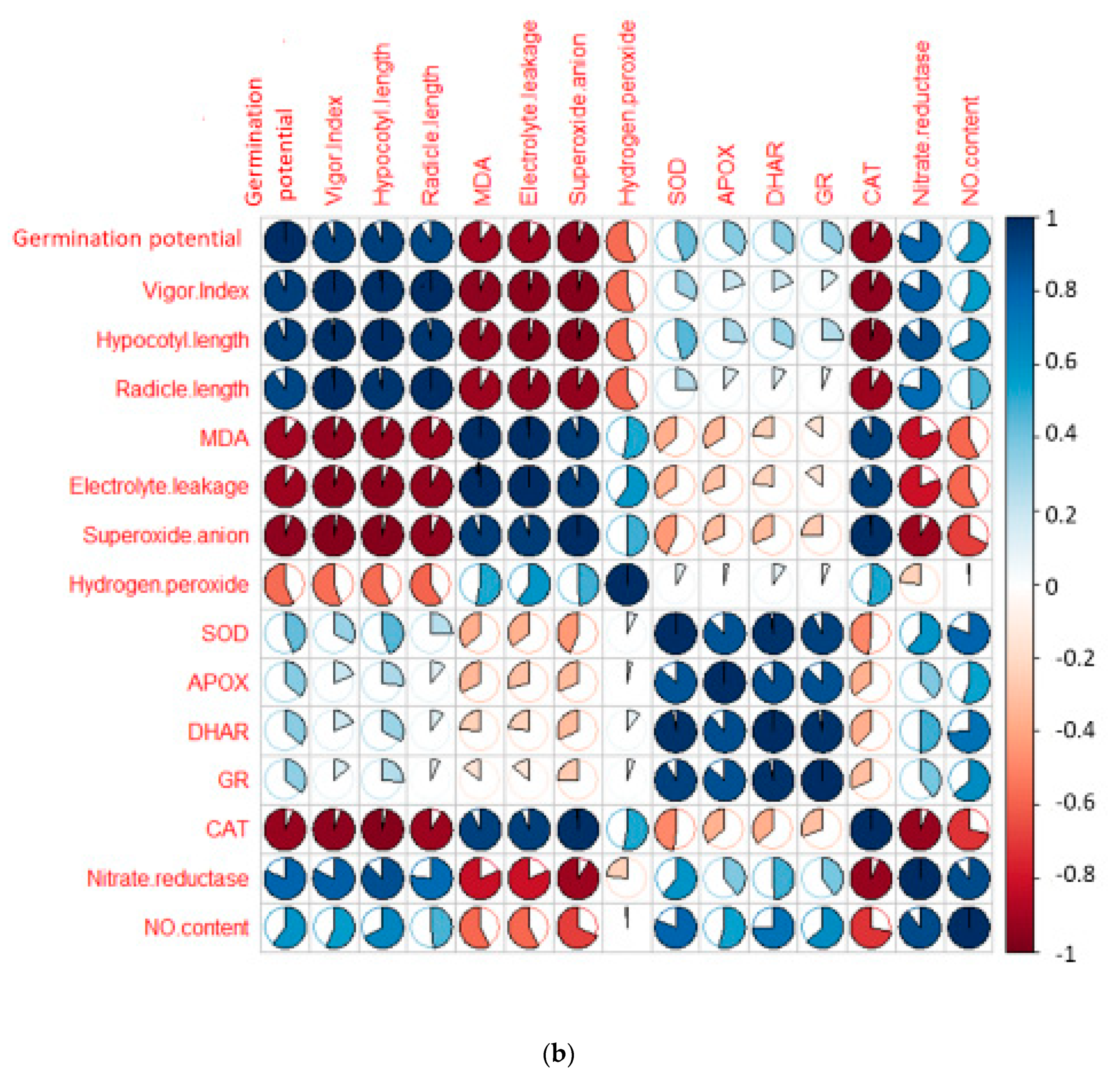
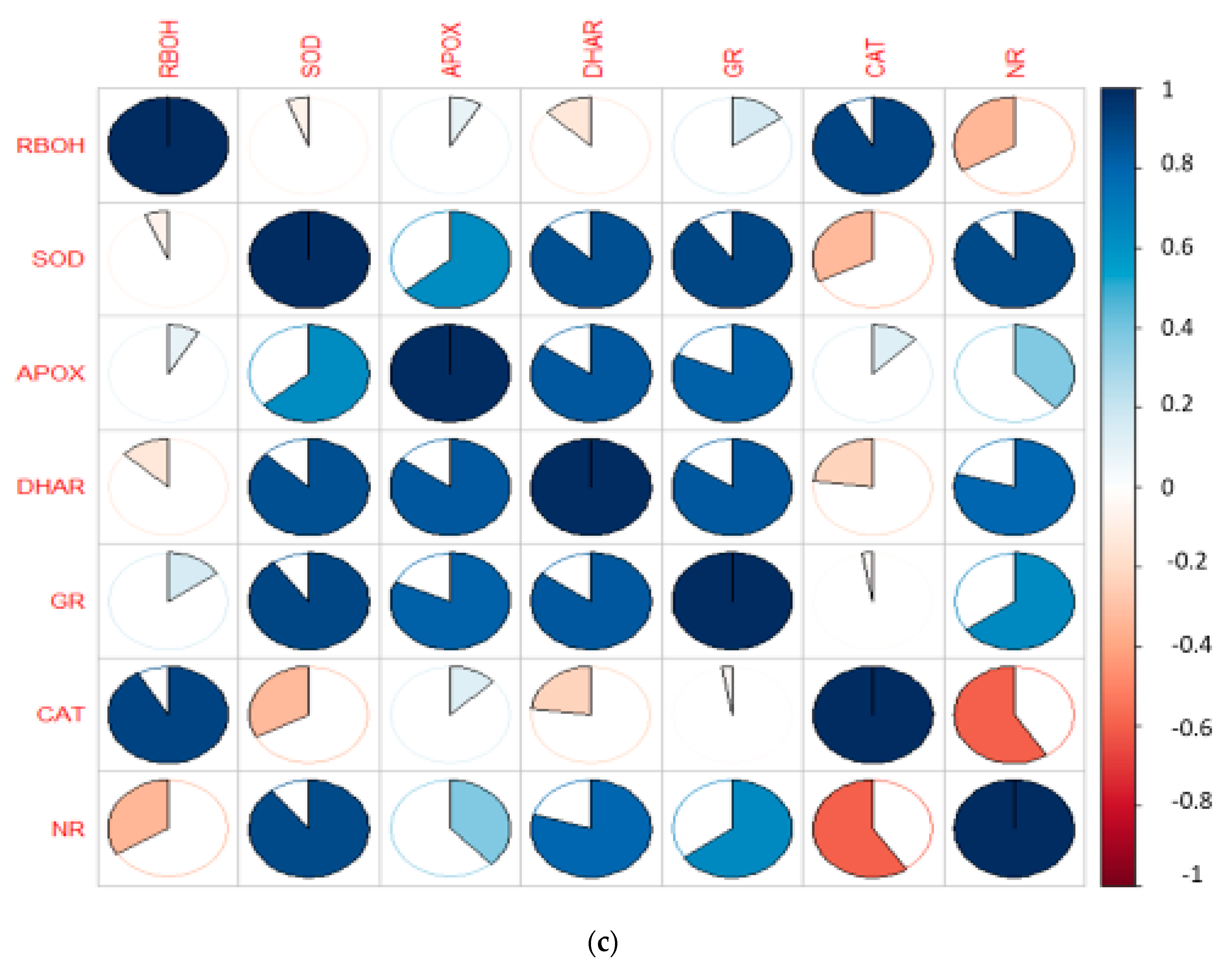
3. Results
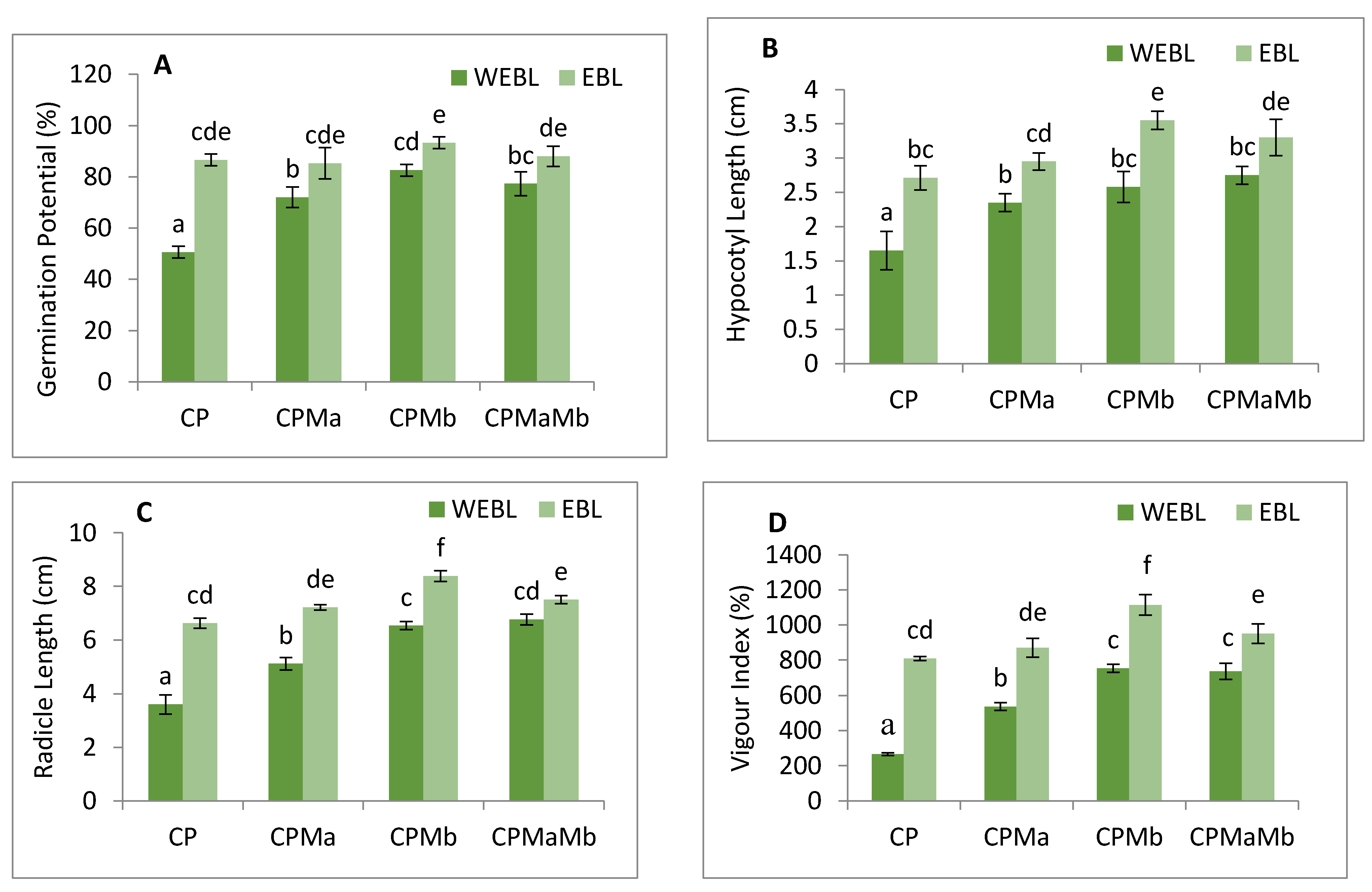
3.1. Effect of EBL and PGPRs on Growth of Brassica juncea Seedlings Treated with CP
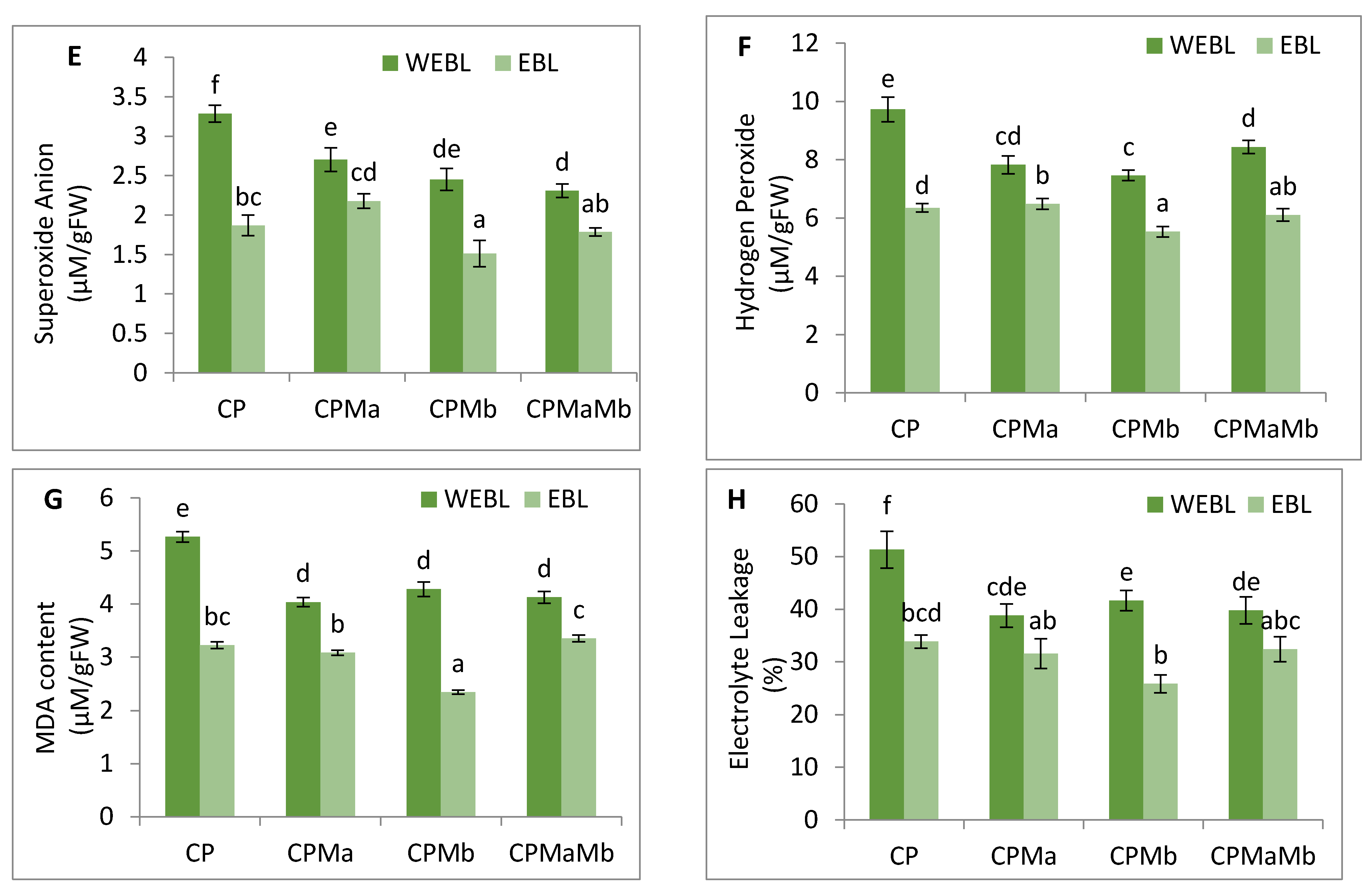
3.2. Effect of EBL and PGPRs on Oxidative Damage in Brassica juncea Seedlings Treated with CP
3.3. Effect of EBL and PGPRs on Lipid Peroxidation and Membrane Permeability in Brassica juncea Seedlings Treated with CP
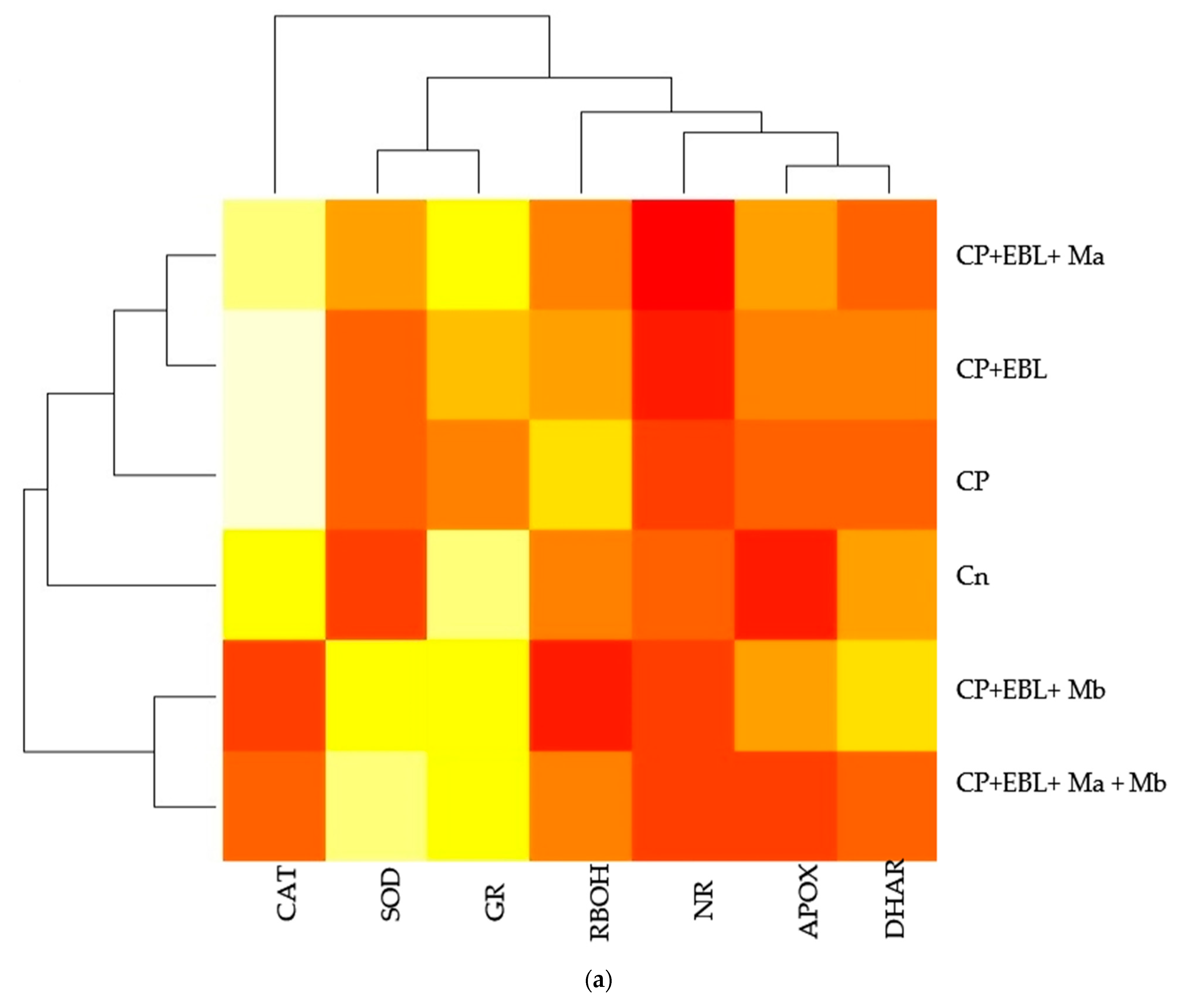
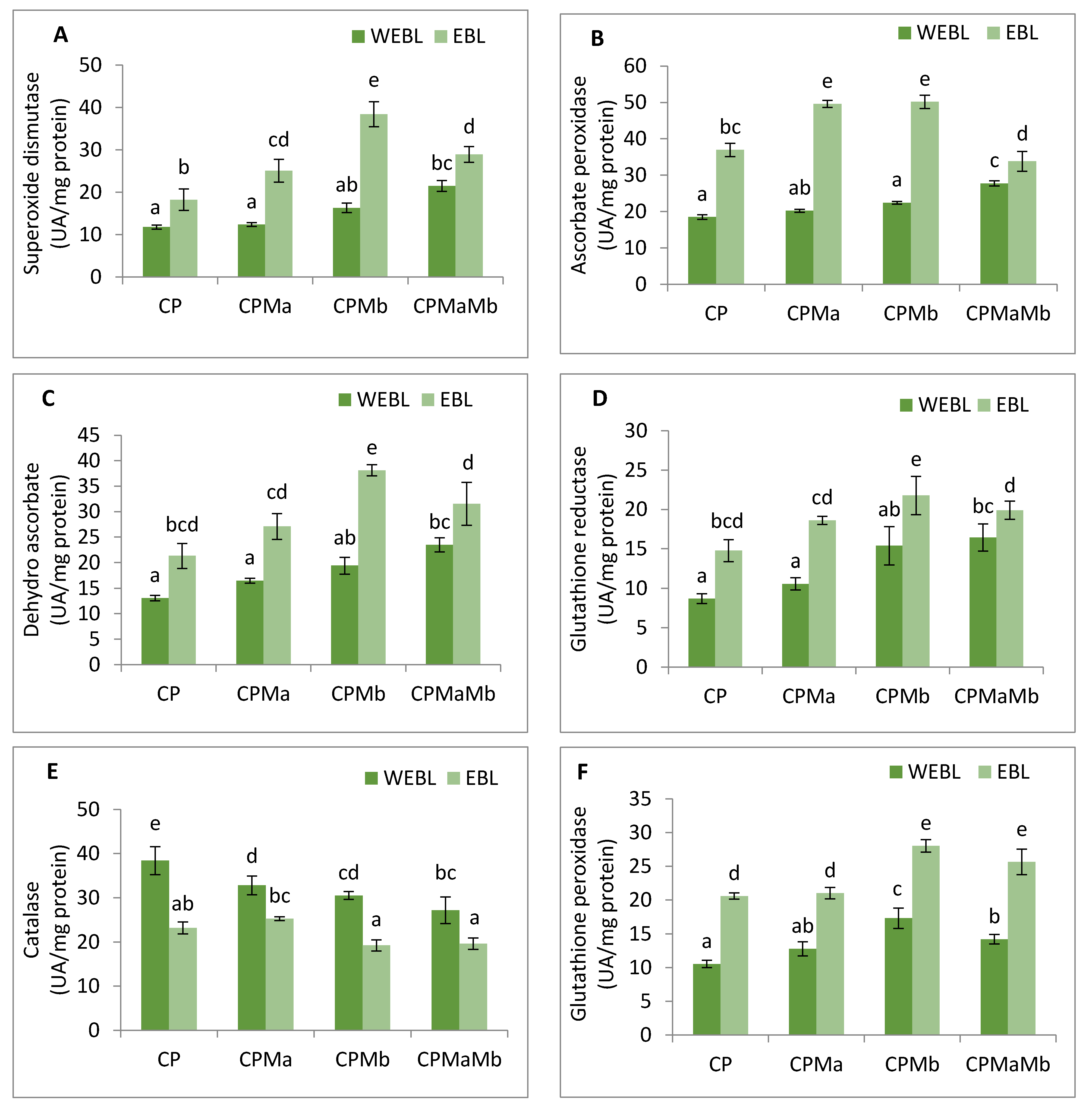
3.4. Effect of EBL and PGPRs on Antioxidative Enzymes in Brassica juncea Seedlings Treated with CP
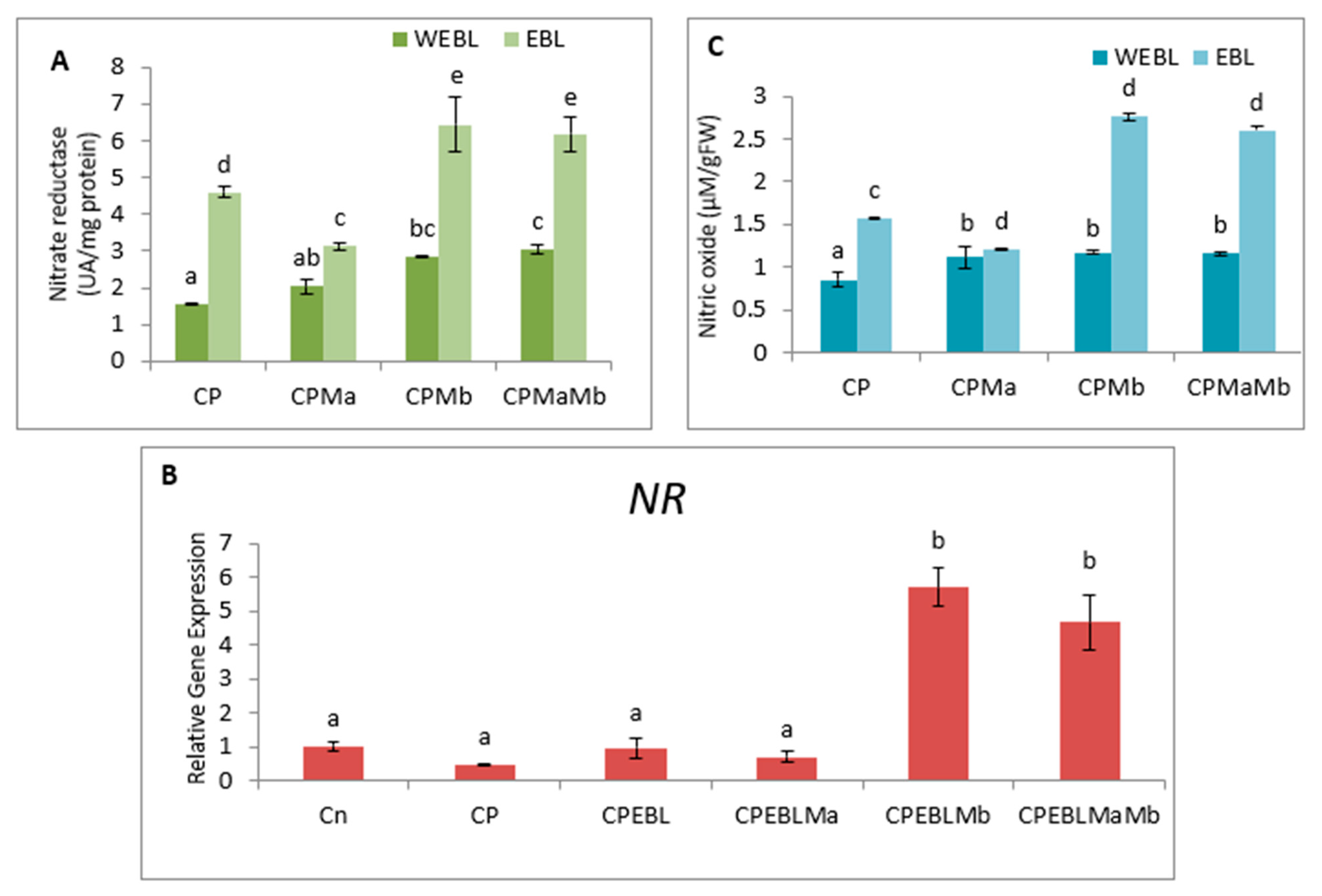
3.5. Effect of EBL and PGPRs on NO and NR Activity in Brassica juncea Seedlings Treated with CP
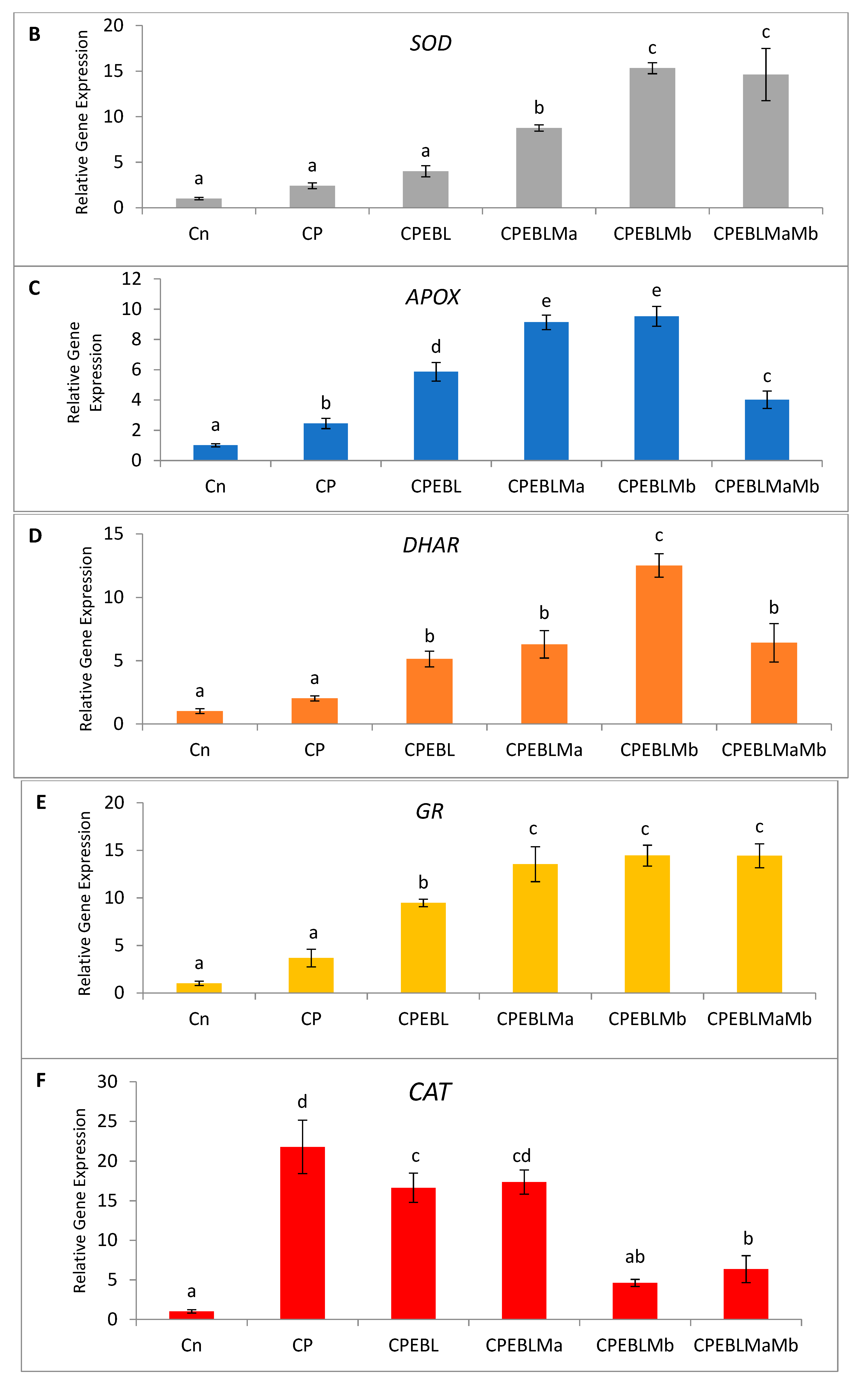
3.6. Effect of EBL and PGPRs in Regulating the Expression of Antioxidative Defence-Related Genesin CP-Treated Brassica juncea Seedlings
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- UN World Population Prospects: The 2017 Revision, Key Findings and Advance Tables; Working Paper No. ESA/P/WP/248; United Nation: New York, NY, USA, 2017.
- Stephens, E.C.; Jones, A.D.; Parsons, D. Agricultural systems research and global food security in the 21st century: An overview and roadmap for future opportunities. Agric. Syst. 2018, 163, 1–6. [Google Scholar] [CrossRef]
- Abhilash, P.C.; Tripathi, V.; Edrisi, S.A.; Dubey, R.K.; Bakshi, M.; Dubey, P.K.; Singh, H.B.; Ebbs, S.D. Sustainability of crop production from polluted lands. Energy Ecol. Environ. 2016, 1, 54–65. [Google Scholar] [CrossRef]
- EL-Saeid, M.H.; Alghamdi, A.G. Identification of Pesticide Residues and Prediction of Their Fate in Agricultural Soil. Water Air Soil Pollut. 2020, 231, 284. [Google Scholar] [CrossRef]
- Liu, N.; Zhu, L. Metabolomic and Transcriptomic Investigation of Metabolic Perturbations in Oryza sativa L. Triggered by Three Pesticides. Environ. Sci. Technol. 2020, 54, 6115–6124. [Google Scholar] [CrossRef]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Briceno, G.; Fuentes, M.S.; Palma, G.; Jorquera, M.A.; Amoroso, M.J.; Diez, M.C. Chlorpyrifos biodegradation and 3, 5, 6-trichloro-2-pyridinol production by actinobacteria isolated from soil. Int. Biodeterior. Biodegrad. 2012, 73, 1–7. [Google Scholar] [CrossRef]
- Grube, A.; Donaldson, D.; Kiely, T.; Wu, L. Pesticides Industry Sales and Usage; US EPA: Washington, DC, USA, 2011.
- Singh, B.K.; Walker, A. Microbial degradation of organophosphorus compounds. FEMS Microbiol. Rev. 2006, 30, 428–471. [Google Scholar] [CrossRef] [PubMed]
- Rayu, S.; Nielsen, U.N.; Nazaries, L.; Singh, B.K. Isolation and molecular characterization of novel chlorpyrifos and 3, 5, 6-trichloro-2-pyridinol-degrading bacteria from sugarcane farm soils. Front. Microbiol. 2017, 8, 518. [Google Scholar] [CrossRef]
- Chandra, R.; Bharagava, R.N.; Yadav, S.; Mohan, D. Accumulation and distribution of toxic metals in wheat (Triticum aestivum L.) and Indian mustard (Brassica campestris L.) irrigated with distillery and tannery effluents. J. Hazard. Mater. 2010, 162, 1514–1521. [Google Scholar] [CrossRef]
- Sharma, I.; Bhardwaj, R.; Pati, P.K. Mitigation of adverse effects of chlorpyrifos by 24-epibrassinolide and analysis of stress markers in a rice variety Pusa Basmati-1. Ecotoxicol. Environ. Saf. 2012, 85, 72–81. [Google Scholar] [CrossRef]
- Yu, C.; Hu, S.; He, P.; Sun, G.; Zhang, C.; Yu, Y. Inducing male sterility in Brassica napus L. by a sulphonylurea herbicide, tribenuron—Methyl. Plant Breed. 2006, 125, 61–64. [Google Scholar] [CrossRef]
- Alavanja, M.C.; Ross, M.K.; Bonner, M.R. Increased cancer burden among pesticide applicators and others due to pesticide exposure. CA Cancer J. Clin. 2013, 63, 120–142. [Google Scholar] [CrossRef] [PubMed]
- Kana, R.; Špundová, M.; Ilık, P.; Lazár, D.; Klem, K.; Tomek, P.; Prášil, O. Effect of herbicide clomazone on photosynthetic processes in primary barley (Hordeum vulgare L.) leaves. Pestic Biochem. Physiol. 2004, 78, 161–170. [Google Scholar] [CrossRef]
- Parween, T.; Jan, S.; Mahmooduzzafar, S.; Fatma, T.; Siddiqui, Z.H. Selective effect of pesticides on plant—A review. Crit. Rev. Food Sci. Nutr. 2016, 56, 160–179. [Google Scholar] [CrossRef]
- Mahapatra, K.; De, S.; Banerjee, S.; Roy, S. Pesticide mediated oxidative stress induces genotoxicity and disrupts chromatin structure in fenugreek (Trigonella foenum-graecum L.) seedlings. J. Hazard. Mater. 2019, 369, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kumar, V.; Yuan, H.; Kanwar, M.K.; Bhardwaj, R.; Thukral, A.K.; Zheng, B. Jasmonic acid seed treatment stimulates insecticide detoxification in Brassica juncea L. Front. Plant Sci. 2018, 9, 1609. [Google Scholar] [CrossRef]
- Zeyad, M.T.; Kumar, M.; Malik, A. Mutagenicity, genotoxicity and oxidative stress induced by pesticide industry wastewater using bacterial and plant bioassays. Biotechnol. Rep. 2019, 24, e00389. [Google Scholar] [CrossRef]
- Fatma, F.; Kamal, A.; Srivastava, A. Exogenous application of salicylic acid mitigates the toxic effect of pesticides in Vigna radiata (L.) Wilczek. J. Plant Growth Regul. 2018, 37, 1185–1194. [Google Scholar] [CrossRef]
- Shakir, S.K.; Irfan, S.; Akhtar, B.; ur Rehman, S.; Daud, M.K.; Taimur, N.; Azizullah, A. Pesticide-induced oxidative stress and antioxidant responses in tomato (Solanum lycopersicum) seedlings. Ecotoxicology 2018, 27, 919–935. [Google Scholar] [CrossRef]
- Gupta, P.; Seth, C.S. Interactive role of exogenous 24 Epibrassinolide and endogenous NO in Brassica juncea L. under salinity stress: Evidence for NR-dependent NO biosynthesis. Nitric Oxide 2020, 97, 33–47. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Prasad, S.M. Interplay of calcium and nitric oxide in improvement of growth and arsenic-induced toxicity in mustard seedlings. Sci. Rep. 2020, 10, 6900. [Google Scholar]
- Idrees, N.; Tabassum, B.; Sarah, R.; Hussain, M.K. Natural Compound from Genus Brassica and Their Therapeutic Activities. In Natural Bio-Active Compounds; Springer: Singapore, 2019; pp. 477–491. [Google Scholar]
- Razaq, M.; Mehmood, A.; Aslam, M.; Ismail, M.; Afzal, M.; Shad, S.A. Losses in yield and yield components caused by aphids to late sown Brassica napus L., Brassica juncea L. and Brassica carrinata A. Braun at Multan, Punjab (Pakistan). Pak. J. Bot. 2011, 43, 319–324. [Google Scholar]
- Ashraf, M.; Akram, N.A.; Arteca, R.N.; Foolad, M.R. The physiological, biochemical and molecular roles of brassinosteroids and salicylic acid in plant processes and salt tolerance. Crit. Rev. Plant Sci. 2010, 29, 162–190. [Google Scholar] [CrossRef]
- Clouse, S.D. Brassinosteroid signal transduction: From receptor kinase activation to transcriptional networks regulating plant development. Plant Cell 2011, 23, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. The Plant Cell. 2020, 32, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; He, B.B.; Qian, X.J.; Zhou, Y.H.; Shi, K.; Zhou, J.; Yu, J.Q.; Xia, X.J. 24-Epibrassinolide alleviates organic pollutants-retarded root elongation by promoting redox homeostasis and secondary metabolism in Cucumis sativus L. Environ. Pollut. 2017, 229, 922–931. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.A.; Hayat, S.; Ahmad, A. Brassinosteroids protect photosynthetic machinery against the cadmium induced oxidative stress in two tomato cultivars. Chemosphere 2011, 84, 1446–1451. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Ahmed, S.; Ali, A.; Yasin, N.A. 2-Hydroxymelatonin mitigates cadmium stress in cucumis sativus seedlings: Modulation of antioxidant enzymes and polyamines. Chemosphere 2020, 243, 125308. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. The role of nitrate reductase in brassinosteroid-induced endogenous nitric oxide generation to improve cadmium stress tolerance of pepper plants by upregulating the ascorbate-glutathione cycle. Ecotoxicol. Environ. Saf. 2020, 196, 110483. [Google Scholar] [CrossRef]
- Pangesti, N.; Pineda, A.; Pieterse, C.M.; Dicke, M.; Van Loon, J.J. Two-way plant mediated interactions between root-associated microbes and insects: From ecology to mechanisms. Front. Plant Sci. 2013, 4, 414. [Google Scholar] [CrossRef]
- Dennis, P.G.; Miller, A.J.; Hirsch, P.R. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities? FEMS Microbiol. Ecol. 2010, 72, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, V.; Kitto, S.L.; Caplan, J.L.; Hsueh, Y.H.; Kearns, D.B.; Wu, Y.S.; Bais, H.P. Microbe-associated molecular patterns-triggered root responses mediate beneficial rhizobacterial recruitment in Arabidopsis. Plant Physiol. 2012, 160, 1642–1661. [Google Scholar] [CrossRef] [PubMed]
- Kroll, S.; Agler, M.T.; Kemen, E. Genomic dissection of host–microbe and microbe–microbe interactions for advanced plant breeding. Curr. Opin. Plant Biol. 2017, 36, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Dar, M.A.; Kaushik, G.; Villarreal-Chiu, J.F. Pollution status and bioremediation of chlorpyrifos in environmental matrices by the application of bacterial communities: A review. J. Environ. Manag. 2019, 239, 124–136. [Google Scholar] [CrossRef]
- Sun, T.; Li, M.; Saleem, M.; Zhang, X.; Zhang, Q. The fungicide “fluopyram” promotes pepper growth by increasing the abundance of P-solubilizing and N-fixing bacteria. Ecotoxicol. Environ. Saf. 2020, 188, 109947. [Google Scholar] [CrossRef]
- Briceño, G.; Vergara, K.; Schalchli, H.; Palma, G.; Tortella, G.; Fuentes, M.S.; Diez, M.C. Organophosphorus pesticide mixture removal from environmental matrices by a soil Streptomyces mixed culture. Environ. Sci. Pollut. Res. 2018, 25, 21296–21307. [Google Scholar] [CrossRef]
- Chawla, N.; Suneja, S.; Kukreja, K. Isolation and characterization of chlorpyriphos degrading bacteria. Ind. J. Agric. Res. 2013, 47, 381–391. [Google Scholar]
- Lakshmi, C.V.; Kumar, M.; Khanna, S. Biodegradation of chlorpyrifos in soil by enriched cultures. Curr. Microbiol. 2009, 58, 35–38. [Google Scholar] [CrossRef]
- Maya, K.; Singh, R.S.; Upadhyay, S.N.; Dubey, S.K. Kinetic analysis reveals bacterial efficacy for biodegradation of chlorpyrifos and its hydrolyzing metabolite TCP. Process Biochem. 2011, 46, 2130–2136. [Google Scholar] [CrossRef]
- Ghanem, I.; Orfi, M.; Shamma, M. Biodegradation of chlorphyrifos by Klebsiella sp. isolated from an activated sludge sample of waste water treatment plant in damascus. Folia Microbiol. 2007, 52, 423–427. [Google Scholar] [CrossRef]
- Anwar, S.; Liaquat, F.; Khan, Q.M.; Khalid, Z.M.; Iqbal, S. Biodegradation of chlorpyrifos and its hydrolysis product 3, 5, 6-trichloro-2-pyridinol by Bacillus pumilus strain C2A1. J. Hazard. Mater. 2009, 168, 400–405. [Google Scholar] [CrossRef]
- El-Helow, E.R.; Badawy, M.E.; Mabrouk, M.E.; Mohamed, E.A.; El-Beshlawy, Y.M. Biodegradation of chlorpyrifos by a newly isolated Bacillus subtilis strain, Y242. Bioremediat J. 2013, 17, 113–123. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Chen, X.; Shi, Y.; Su, Z.C. Bacterial Degradation of Chlorpyrifos by Bacillus Cereus. In Advanced Materials Research; Trans Tech Publications Ltd.: Bach, Switzerland, 2012; pp. 676–680. [Google Scholar]
- Kumar, D. Biodegradation of γ-Hexachlorocyclohexane by Burkholderia sp. IPL04. Biocatal. Agric. Biotechnol. 2018, 16, 331–339. [Google Scholar] [CrossRef]
- Khanna, K.; Jamwal, V.L.; Kohli, S.K.; Gandhi, S.G.; Ohri, P.; Bhardwaj, R.; Abd Allah, E.F.; Hashem, A.; Ahmad, P. Plant growth promoting rhizobacteria induced Cd tolerance in Lycopersicon esculentum through altered antioxidative defense expression. Chemosphere 2019, 217, 463–474. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, F.M.; de Assis Pereira, T.; Souza, T.P.; Guimarães, P.H.S.; Martins, A.D.; Schwan, R.F.; Pasqual, M.; Dória, J. Beneficial effects of inoculation of growth-promoting bacteria in strawberry. Microbiol. Res. 2019, 223, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, C.; Banerjee, S.; Acharya, U.; Mitra, A.; Mallick, I.; Haldar, A.; Ghosh, A.; Ghosh, A. Evaluation of plant growth promotion properties and induction of antioxidative defense mechanism by tea rhizobacteria of Darjeeling, India. Sci. Rep. 2020, 10, 15536. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Yasmeen, T.; Ali, Q.; Ali, S.; Arif, M.S.; Hussain, S.; Rizvi, H. Influence of Pseudomonas aeruginosa as PGPR on oxidative stress tolerance in wheat under Zn stress. Ecotoxicol.Environ. Saf. 2014, 104, 285–293. [Google Scholar] [CrossRef]
- Pozo, M.J.; López-Ráez, J.A.; Azcón-Aguilar, C.; García-Garrido, J.M. Phytohormones as integrators of environmental signals in the regulation of mycorrhizal symbioses. New Phytol. 2015, 205, 1431–1436. [Google Scholar] [CrossRef]
- Selosse, M.A.; Bessis, A.; Pozo, M.J. Microbial priming of plant and animal immunity: Symbionts as developmental signals. Trends Microbiol. 2014, 22, 607–613. [Google Scholar] [CrossRef]
- Carvalhais, L.C.; Schenk, P.M.; Dennis, P.G. Jasmonic acid signalling and the plant holobiont. Curr. Opin. Microbiol. 2017, 37, 42–47. [Google Scholar] [CrossRef]
- Yu, M.H.; Zhao, Z.Z.; He, J.X. Brassinosteroid signaling in plant–microbe interactions. Int. J. Mol. Sci. 2018, 19, 4091. [Google Scholar] [CrossRef]
- Eichmann, R.; Richards, L.; Schäfer, P. Hormones as go-betweens in plant microbiome assembly. Plant J. 2021, 105, 518–541. [Google Scholar] [CrossRef] [PubMed]
- Pati, U.K.; Chowdhury, A. A comparison of phytotoxic potential among the crude extracts from Parthenium hysterophorus L. extracted with solvents of increasing polarity. Int. Lett. Nat. Sci. 2015, 33, 73–81. [Google Scholar] [CrossRef]
- Batool, A.; Ziaf, K.; Amjad, M. Effect of halo-priming on germination and vigor index of cabbage (Brassica oleracea var. capitata). J. Environ. Agric. Sci. 2015, 2, 1–8. [Google Scholar]
- Lu, S.; Wang, Z.; Niu, Y.; Guo, Z.; Huang, B. Antioxidant responses of radiation-induced dwarf mutants of Bermuda grass to drought stress. J. Am. Soc. Hortic. Sci. 2008, 133, 360–366. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; Liu, W.; Ye, X.F.; Zai, H.F.; Zhao, L.J. Alleviation of salt stress in citrus seedlings inoculated with mycorrhiza: Changes in leaf antioxidant defense systems. Plant Soil Environ. 2010, 56, 470–475. [Google Scholar] [CrossRef]
- Frahry, G.; Schopfer, P. NADH-stimulated, cyanide-resistant superoxide production in maize coleoptiles analyzed with a tetrazolium-based assay. Planta 2001, 212, 175–183. [Google Scholar] [CrossRef]
- Patterson, B.D.; MacRae, E.A.; Ferguson, I.B. Estimation of hydrogen peroxide in plant extracts using titanium (IV). Anal. Biochem. 1984, 139, 487–492. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley—Powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Rodriguez-Serrano, M.; Romero-Puertas, M.C.; Pazmino, D.M.; Testillano, P.S.; Risueño, M.C.; Luis, A.; Sandalio, L.M. Cellular response of pea plants to cadmium toxicity: Cross talk between reactive oxygen species, nitric oxide, and calcium. Plant Physiol. 2009, 150, 229–243. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Pompella, A.; Maellaro, E.; Casini, A.F.; Comporti, M. Histochemical detection of lipid peroxidation in the liver of bromobenzene-poisoned mice. Am. J. Pathol. 1987, 129, 295–301. [Google Scholar] [PubMed]
- Yamamoto, Y.; Kobayashi, Y.; Matsumoto, H. Lipid peroxidation is an early symptom triggered by aluminum, but not the primary cause of elongation inhibition in pea roots. Plant Physiol. 2001, 125, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Alcala, G.; Gotor, C.; Meyer, A.J.; Fricker, M.; Vega, J.M.; Romero, L.C. Glutathione biosynthesis in Arabidopsis trichome cells. Proc. Natl. Acad. Sci. USA 2000, 97, 11108–11113. [Google Scholar] [CrossRef]
- Callard, D.; Axelos, M.; Mazzolini, L. Novel molecular markers for late phases of the growth cycle of Arabidopsis thaliana cell-suspension cultures are expressed during organ senescence. Plant Physiol. 1996, 112, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, E.G. Nitrate reductase assay in intact plant tissues. Biochem. Biophys. Res. Commun. 1971, 43, 1274–1279. [Google Scholar] [CrossRef]
- Zhou, B.; Guo, Z.; Xing, J.; Huang, B. Nitric oxide is involved in abscisic acid-induced antioxidant activities in Stylosanthes guianensis. J. Exp. Bot. 2005, 56, 3223–3228. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzym. Anal. 1984, 105, 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Flohe, L.; Gunzler, W.A. Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar] [PubMed]
- Carlberg, I.N.C.E.R.; Mannervik, B.E.N.G.T. Purification and characterization of the flavoenzyme glutathione reductase from rat liver. J. Biol. Chem. 1975, 250, 5475–5480. [Google Scholar] [CrossRef]
- Dalton, D.A.; Russell, S.A.; Hanus, F.J.; Pascoe, G.A.; Evans, H.J. Enzymatic reactions of ascorbate and glutathione that prevent peroxide damage in soybean root nodules. Proc. Natl. Acad. Sci. USA 1986, 83, 3811–3815. [Google Scholar] [CrossRef]
- Kono, Y. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef]
- Awasthi, P.; Mahajan, V.; Jamwal, V.L.; Kapoor, N.; Rasool, S.; Bedi, Y.S.; Gandhi, S.G. Cloning and expression analysis of chalcone synthase gene from Coleus forskohlii. J. Genet. 2016, 95, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Yuan, H.; Kumar, V.; Ramakrishnan, M.; Kohli, S.K.; Kaur, R.; Thukral, A.K.; Bhrardwaj, R.; Zheng, B. Castasterone attenuates insecticide induced phytotoxicity in mustard. Ecotoxicol. Environ. Saf. 2019, 179, 50–61. [Google Scholar] [CrossRef]
- Sharma, A.; Thakur, S.; Kumar, V.; Kanwar, M.K.; Kesavan, A.K.; Thukral, A.K.; Bhardwaj, R.; Parvaiz, A.; Ahmad, P. Pre-sowing seed treatment with 24-epibrassinolide ameliorates pesticide stress in Brassica juncea L. through the modulation of stress markers. Front. Plant Sci. 2016, 7, 1569. [Google Scholar] [CrossRef] [PubMed]
- Eleiwa, M.E.; Bafeel, S.O.; Ibrahim, S.A. Influence of brassinosteroids on wheat plant (Triticum aestivum L.) production under salinity stress conditions. I-Growth parameters and photosynthetic pigments. Aust. J. Basic Appl. Sci. 2011, 5, 58–65. [Google Scholar]
- Kaur, R.; Yadav, P.; Sharma, A.; Thukral, A.K.; Kumar, V.; Kohli, S.K.; Bhardwaj, R. Castasterone and citric acid treatment restores photosynthetic attributes in Brassica juncea L. under Cd (II) toxicity. Ecotoxicol. Environ. Saf. 2017, 145, 466–475. [Google Scholar] [CrossRef]
- Yadav, P.; Kaur, R.; Kanwar, M.K.; Sharma, A.; Verma, V.; Sirhindi, G.; Bhardwaj, R. Castasterone confers copper stress tolerance by regulating antioxidant enzyme responses, antioxidants, and amino acid balance in B. juncea seedlings. Ecotoxicol. Environ. Saf. 2018, 147, 725–734. [Google Scholar] [CrossRef]
- Ahmad, P.; Abd_Allah, E.F.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Bhardwaj, R.; Siddique, K.H. Exogenous application of calcium to 24-epibrassinosteroid pre-treated tomato seedlings mitigates NaCl toxicity by modifying ascorbate–glutathione cycle and secondary metabolites. Sci. Rep. 2018, 8, 13515. [Google Scholar] [CrossRef]
- Wu, C.; Li, F.; Xu, H.; Zeng, W.; Yu, R.; Wu, X.; Shen, L.; Liu, Y.; Li, J. The potential role of brassinosteroids (BRs) in alleviating antimony (Sb) stress in Arabidopsis thaliana. Plant Physiol. Biochem. 2019, 141, 51–59. [Google Scholar] [CrossRef]
- Rattan, A.; Kapoor, D.; Kapoor, N.; Bhardwaj, R.; Sharma, A. Brassinosteroids regulate functional components of antioxidative defense system in salt stressed maize seedlings. Plant Growth Regul. 2020, 39, 1465–1475. [Google Scholar] [CrossRef]
- Cerana, R.; Bonetti, A.; Marre, M.T.; Romani, G.; Lado, P.; Marre, E. Effects of a brassinosteroid on growth and electrogenic proton extrusion in Azuki bean epicotyls. Physiol. Plant. 1983, 59, 23–27. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Yusuf, M.; Ahmad, I.; Ahmad, A. Brassinosteroids and their role in response of plants to abiotic stresses. Biol. Plant. 2014, 58, 9–17. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Khan, E.A. 24-Epibrassinolide application in plants: An implication for improving drought stress tolerance in plants. Plant Physiol Biochem. 2019, 135, 295–303. [Google Scholar] [CrossRef]
- Kutschera, U.; Wang, Z.Y. Growth-limiting proteins in maize coleoptiles and the auxin-brassinosteroid hypothesis of mesocotyl elongation. Protoplasma 2016, 253, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, D.K.; Verma, J.P.; Krishna, R.; Gaurav, A.K.; Yadav, J. Molecular characterization of monocrotophos and chlorpyrifos tolerant bacterial strain for enhancing seed germination of vegetable crops. Chemosphere 2019, 223, 636–650. [Google Scholar] [CrossRef] [PubMed]
- Asari, S.; Tarkowská, D.; Rolčík, J.; Novák, O.; Palmero, D.V.; Bejai, S.; Meijer, J. Analysis of plant growth-promoting properties of Bacillus amyloliquefaciens UCMB5113 using Arabidopsis thaliana as host plant. Planta 2017, 245, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Akbar, S.; Sultan, S. Soil bacteria showing a potential of chlorpyrifos degradation and plant growth enhancement. Braz. J. Microbiol. 2016, 47, 563–570. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef]
- Singh, D.; Ghosh, P.; Kumar, J.; Kumar, A. Plant Growth-Promoting Rhizobacteria (PGPRs): Functions and Benefits. In Microbial Interventions in Agriculture and Environment; Springer: Singapore, 2019; pp. 205–227. [Google Scholar]
- Ambreetha, S.; Chinnadurai, C.; Marimuthu, P.; Balachandar, D. Plant-associated Bacillus modulates the expression of auxin-responsive genes of rice and modifies the root architecture. Rhizosphere 2018, 5, 57–66. [Google Scholar] [CrossRef]
- Jatan, R.; Chauhan, P.S.; Lata, C. Pseudomonas putida modulates the expression of miRNAs and their target genes in response to drought and salt stresses in chickpea (Cicer arietinum L.). Genomics 2019, 111, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Foo, E.; Plett, J.M.; Lopez-Raez, J.A.; Reid, D. The Role of plant hormones in plant-microbe symbioses. Front Plant Sci. 2019, 10, 1391. [Google Scholar] [CrossRef] [PubMed]
- McGuiness, P.N.; Reid, J.B.; Foo, E. The role of gibberellins and brassinosteroids in nodulation and arbuscular mycorrhizal associations. Front. Plant Sci. 2019, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- McGuiness, P.N.; Reid, J.B.; Foo, E. Brassinosteroids play multiple roles in nodulation of pea via interactions with ethylene and auxin. Planta 2020, 252, 1–8. [Google Scholar] [CrossRef] [PubMed]
- McGuiness, P.N.; Reid, J.B.; Foo, E. The influence of ethylene, gibberellins and brassinosteroids on energy and nitrogen-fixation metabolites in nodule tissue. Plant Sci. 2021, 305, 110846. [Google Scholar] [CrossRef]
- Foo, E.; McAdam, E.L.; Weller, J.L.; Reid, J.B. Interactions between ethylene, gibberellins, and brassinosteroids in the development of rhizobial and mycorrhizal symbioses of pea. J. Exp. Bot. 2016, 67, 2413–2424. [Google Scholar] [CrossRef] [PubMed]
- Tofighi, C.; Khavari-Nejad, R.A.; Najafi, F.; Razavi, K.; Rejali, F. Brassinosteroid (BR) and arbuscular mycorrhizal (AM) fungi alleviate salinity in wheat. J. Plant Nutr. 2017, 40, 1091–1098. [Google Scholar] [CrossRef]
- Sharma, I.; Bhardwaj, R.; Pati, P.K. Exogenous application of 28-homobrassinolide modulates the dynamics of salt and pesticides induced stress responses in an elite rice variety Pusa Basmati-1. Plant Growth Regul. 2015, 34, 509–518. [Google Scholar] [CrossRef]
- Chen, Q.; Yang, G. Signal function studies of ROS, especially RBOH-dependent ROS, in plant growth, development and environmental stress. Plant Growth Regul. 2019, 39, 1–15. [Google Scholar] [CrossRef]
- Khafi, A.S.; Iranbakhsh, A.; Afshar, A.S.; Nejad, R.A.K. RBOH expression and ROS metabolism in Citrullus colocynthis under cadmium stress. Braz. J. Bot. 2020, 43, 35–43. [Google Scholar] [CrossRef]
- Zhou, Y.; Xia, X.; Yu, G.; Wang, J.; Wu, J.; Wang, M.; Yang, Y.; Yu, K.S.Y.; Chen, Z.; Gan, J.; et al. Brassinosteroids play a critical role in the regulation of pesticide metabolism in crop plants. Sci. Rep. 2015, 5, 9018. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Yüzbaşıoğlu, E.; Dalyan, E. Salicylic acid alleviates thiram toxicity by modulating antioxidant enzyme capacity and pesticide detoxification systems in the tomato (Solanum lycopersicum Mill.). Plant Physiol. Biochem. 2019, 135, 322–330. [Google Scholar] [CrossRef]
- Homayoonzadeh, M.; Moeini, P.; Talebi, K.; Roessner, U.; Hosseininaveh, V. Antioxidant system status of cucumber plants under pesticides treatment. Acta Physiol Plant. 2020, 42, 161. [Google Scholar] [CrossRef]
- Wang, Z.; Jiang, Y.; Peng, X.; Xu, S.; Zhang, H.; Gao, J.; Xi, Z. Exogenous 24-epibrassinolide regulates antioxidant and pesticide detoxification systems in grapevine after chlorothalonil treatment. Plant Growth Regul. 2017, 81, 455–466. [Google Scholar] [CrossRef]
- Liu, S.; He, Y.; Tian, H.; Yu, C.; Tan, W.; Li, Z.; Duan, L. Application of brassinosteroid mimetics improves growth and tolerance of maize to nicosulfuron toxicity. Plant Growth Regul. 2019, 38, 701–712. [Google Scholar] [CrossRef]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K.; Abid, M.; Rizvan, M.; Shahid, M.R.; Alotaibi, M.; et al. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci. Rep. 2020, 10, 16975. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Singh, S.P.; Upadhyay, S.K. Role of Superoxide Dismutases (SODs) in Stress Tolerance in Plants. In Molecular Approaches in Plant Biology and Environmental Challenges. Energy, Environment, and Sustainability; Singh, S., Upadhyay, S., Pandey, A., Kumar, S., Eds.; Springer: Singapore, 2019. [Google Scholar] [CrossRef]
- Hakeem, K.R.; Alharby, H.F.; Rehman, R. Antioxidative defense mechanism against lead-induced phytotoxicity in Fagopyrum kashmirianum. Chemosphere 2019, 216, 595–604. [Google Scholar] [CrossRef]
- Sahu, G.K.; Upadhyay, S.; Sahoo, B.B. Mercury induced phytotoxicity and oxidative stress in wheat (Triticum aestivum L.) plants. Physiol. Mol. Biol. Plants. 2012, 18, 21–31. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Cuypers, A.; Karen, S.; Jos, R.; Kelly, O.; Els, K.; Tony, R.; Nele, H.; Nathalie, V.; Suzy, V.S.; Frank, V.B.; et al. The cellular redox state as a modulator in cadmium and copper responses in Arabidopsis thaliana seedlings. J. Plant Physiol. 2011, 168, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Ruan, Y.P.; Zhou, J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Brassinosteroid alleviates polychlorinated biphenyls-induced oxidative stress by enhancing antioxidant enzymes activity in tomato. Chemosphere 2013, 90, 2645–2653. [Google Scholar] [CrossRef]
- Kasotia, A.; Varma, A.; Choudhary, D.K. Pseudomonas-mediated mitigation of salt stress and growth promotion in Glycine max. Agric. Res. 2015, 4, 31–41. [Google Scholar] [CrossRef]
- Rais, A.; Jabeen, Z.; Shair, F.; Hafeez, F.Y.; Hassan, M.N. Bacillus spp., a bio-control agent enhances the activity of antioxidant defense enzymes in rice against Pyricularia oryzae. PLoS ONE 2017, 12, e0187412. [Google Scholar]
- Tripathi, D.K.; Mishra, R.K.; Singh, S.; Singh, S.; Singh, V.P.; Singh, P.K.; Prasad, S.M.; Dubey, N.K.; Pandey, A.C.; Sahi, S.; et al. Nitric oxide ameliorates zinc oxide nanoparticles phytotoxicity in wheat seedlings: Implication of the ascorbate-glutathione cycle. Front. Plant Sci. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.X.; Zhou, Y.H.; Ding, J.G.; Xia, X.J.; Shi, K.A.I.; Chen, S.C.; Asami, T.; Chen, J.Q.; Yu, J.Q. Role of nitric oxide in hydrogen peroxide-dependent induction of abiotic stress tolerance by brassinosteroids in cucumber. Plant Cell Environ. 2011, 34, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Hichri, I.; Boscari, A.; Meilhoc, E.; Catalá, M.; Barreno, E.; Bruand, C.; Lanfranco, L.; Brouquisse, R. Nitric oxide: A multitask player in plant–microorganism symbioses. Gasotransmitters Plant. 2016, 239–268. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Tripathi, D.K.; Sharma, S. Silicon and plant growth promoting rhizobacteria differentially regulate AgNP-induced toxicity in Brassica juncea: Implication of nitric oxide. J. Hazard. Mater. 2020, 390, 121806. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene and Gene ID | Sequences |
| Actin (KM881,428.1) | Forward Primer 5′ ACTGGTATTGTGCTTGACTCTG3′ Reverse Primer 5′ AGCTTCTCTTTAATGTCACGGAC3′ |
| SOD (AF540,558.1) | Forward Primer 5′ CACATTTCAACCCTGATGGTAA3′ Reverse Primer 5′ ACAGCCCTTCCGACAATA3′ |
| APOX (AF038,839.1) | Forward Primer 5′ CCACTTGAGACAGGTGTTACTA3′ Reverse Primer 5′ TCCTTGAAGTAAGAGTTGTCGAAA3′ |
| DHAR (AF536,330.1) | Forward Primer 5′ CTGGATGAGCTTAGTACATTCAAC3′ Reverse Primer 5′ GGAAAGAAAGTGAATCTGGAACA3′ |
| GR (AF349,449.1) | Forward Primer 5′ GATGCAGCGCTTGATTTAC3′ Reverse Primer 5′ TCCCTAACGTCTTCATCAAACC3′ |
| CAT (AF104,451.1) | Forward Primer 5′ GTTCGACTTTGACCCACT3′ Reverse Primer 5′ ATCCCAGGAACAATGATAGC3′ |
| NR (XM_022711045.1) | Forward Primer 5′ GGTGGAGGTGACTCTAGATG3′ Reverse Primer 5′ TCGAACCGCAACGTCTTTA3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakshi, P.; Chouhan, R.; Sharma, P.; Mir, B.A.; Gandhi, S.G.; Landi, M.; Zheng, B.; Sharma, A.; Bhardwaj, R. Amelioration of Chlorpyrifos-Induced Toxicity in Brassica juncea L. by Combination of 24-Epibrassinolide and Plant-Growth-Promoting Rhizobacteria. Biomolecules 2021, 11, 877. https://doi.org/10.3390/biom11060877
Bakshi P, Chouhan R, Sharma P, Mir BA, Gandhi SG, Landi M, Zheng B, Sharma A, Bhardwaj R. Amelioration of Chlorpyrifos-Induced Toxicity in Brassica juncea L. by Combination of 24-Epibrassinolide and Plant-Growth-Promoting Rhizobacteria. Biomolecules. 2021; 11(6):877. https://doi.org/10.3390/biom11060877
Chicago/Turabian StyleBakshi, Palak, Rekha Chouhan, Pooja Sharma, Bilal Ahmad Mir, Sumit G. Gandhi, Marco Landi, Bingsong Zheng, Anket Sharma, and Renu Bhardwaj. 2021. "Amelioration of Chlorpyrifos-Induced Toxicity in Brassica juncea L. by Combination of 24-Epibrassinolide and Plant-Growth-Promoting Rhizobacteria" Biomolecules 11, no. 6: 877. https://doi.org/10.3390/biom11060877
APA StyleBakshi, P., Chouhan, R., Sharma, P., Mir, B. A., Gandhi, S. G., Landi, M., Zheng, B., Sharma, A., & Bhardwaj, R. (2021). Amelioration of Chlorpyrifos-Induced Toxicity in Brassica juncea L. by Combination of 24-Epibrassinolide and Plant-Growth-Promoting Rhizobacteria. Biomolecules, 11(6), 877. https://doi.org/10.3390/biom11060877